Introduction
Diabetes mellitus (DM) is a systemic metabolic
disorder characterized by impaired glucose metabolism, the
incidence of which continues to grow at an alarming rate from 10.6%
in 1989 to 32.1% in 2009 in Saudi Arabia based (1) A severe microvascular complication of
DM, diabetic retinopathy (DR), is one of the four major blinding
diseases that has become the leading cause of adult blindness,
according to surveys conducted in other developed countries,
including the United States and Japan (2). DR is typically diagnosed at a late
stage when treatment options are limited. Due to its asymptomatic
onset and progressive disease course, DR often results in
irreversible blindness, which has a shortage of effective treatment
due to the complex nature of the disease and limited knowledge of
its pathogenic mechanism (3–5). Therefore, finding an effective
treatment for DR has important clinical significance.
Poor blood glucose control, hypertension,
hyperlipidemia and obesity may be involved in the development of
DR, and may also contribute to the development of neuropathy
(6). Studies (7,8)
assessing the pathogenetic pathways that lead to microangiopathy
indicate that many genes may be involved in its development. Known
biochemical mechanisms that underlie DR include advanced glycation
end-product (AGE) formation, oxidative stress, the activation of
protein kinase C (PKC) isoforms and polyol and hexosamine pathway
activity (7). As a result of
impaired pathway activity, a number of issues may arise, including
retinal tissue hypoxia, endothelial cell dysfunction, vasodilation
impairment, angiogenic factor hyperactivation and changes to the
extracellular matrix (7). At
present, the treatment of DR primarily targets its proliferation,
but the restoration of lost visual function remains difficult
(8).
Carnosine is an endogenous dipeptide consisting of
beta-alanine and L-histidine that is produced in skeletal muscle
and the nervous system, which is influenced by diet (9,10). It
has been reported that many biological effects of carnosine,
including anti-inflammation, anti-aging, immune-regulation and
anti-diabetes are associated with its anti-oxidative activities
(11–13). Carnosine has been revealed to be an
effective scavenger for the removal of reactive carbonyl species,
as determined in a mouse model of diabetic nephropathy, and the
reactive species located in pancreatic β-cells (10,14).
Javadi et al (15)
demonstrated that carnosine primarily prevents dehydroascorbic
acid-induced unfolding and the aggregation of lens proteins and
significant lens opacity. Furthermore, other studies have revealed
that carnosine exerts a positive effect on the prevention and
treatment of DR, but it has no association with anti-oxidation and
anti-glycosylation (16,17).
Mitogen-activated protein kinase (MAPK) is a signal
transduction pathway that is involved in various physiological
processes, including gene expression and the proliferation,
differentiation, death and survival of various cells (18,19).
Members of the MAPK family are regulated by a cascade of
phosphorylation and are activated by extracellular stimulus
(20). In the process of ERK1/2
phosphorylation, the extracellular signal related kinase 1/2
(ERK1/2) cascade is involved in the MAPK pathway, while MAPK
signaling is activated by MAPK kinase 1 (21). The activation of the MAPK/ERK
signaling pathway has been used frequently for the study of
diabetic complications (22).
Previous studies have also revealed that the PKC signaling pathway
is involved in diabetic wound healing (23) and diabetic myocardial injury
(24). The current study assessed
the alteration of PKC, ERK, and phosphorylated (p)-ERK expression
in diabetic rats following treatment with carnosine, PD98059 (an
inhibitor of MAPK/ERK) or U46619 (an activator of MAPK/ERK).
Reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blotting was performed to assess the
association between carnosine and the MAPK/ERK signaling pathway.
The results revealed that suppressing the activation of the
MAPK/ERK signaling pathway may serve an important role in the
prevention and treatment carnosine-induced DR. However, the PKC
pathway may not be involved in this process.
Materials and methods
Reagents and animals
Streptozotocin (STZ) was purchased from
Sigma-Aldrich (Merck KGaA; Darmstadt, Germany). Cluster of
differentiation (CD)31 antibodies were purchased from Abcam
(Cambridge, UK), and Goat anti-Mouse Immunoglobulin G (IgG)
H&L-cyanine 3 (Cy3) antibodies were purchased from ProteinTech
Group, Inc. (Chicago, IL, USA). A total of 25 male sprague dawley
rats (age, 7 weeks; weight, 180–185 g) were purchased from the
Experimental Animal Center of Southern Medical University
(Guangzhou, China). All rats were housed in a specific pathogen
free room at a temperature of 25°C and a humidity of 50% under a 12
h light/dark cycle, with ad libitum access to food and
water.
Ethics statement
All animal experiments were performed in accordance
with the recommendations included in the Guide for the Care and Use
of Laboratory Animals of the National Institutes of Health.
Furthermore, the protocol of the current study was approved by the
Committee on the Ethics of Animal Experiments of Tangdu Hospital,
Air Force Medical University (Xi'an, China).
Establishment of diabetic rat
model
Following a week of feeding, the body weight of rats
was measured. A total of 10 rats were randomly divided into 2
groups: The DM group (n=5; weight, 200–220 g) and the normal group
(n=5; weight, 180–200 g). Rats in the DM group were
intraperitoneally injected once with 2% STZ at a dose of 60 mg/kg.
Animals were considered diabetic when glucose levels were >16 mM
at 72 h following injection. Normal rats were administered the same
quantity (100 µl) of citrate buffer solution (0.02 mol/l; pH=4.5).
Rats were euthanized via an intraperitoneal injection of 240 mg/kg
sodium pentobarbital followed by cervical dislocation. Retinas were
then harvested for cell isolation.
Rat retinal vascular endothelial cell
(RVEC) isolation and cell culture
To establish a primary cell culture of rat RVECs,
retinas isolated from rats of each group were digested with trypsin
at 37°C for 30 min. Residual retinas were then minced and fragments
were filtered through a 100 µm cell strainer. Following harvest,
fragments from the strainer were incubated in 3 ml of mixed
collagenase (Sigma-Aldrich) for 30 min at room temperature.
Digestion was stopped using complete medium (Dulbecco's Modified
Eagles medium supplemented with 10% fetal bovine serum, 100 U/ml
penicillin and 100 µg/ml streptomycin, all Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at room temperature. Following
centrifugation at 252 × g at room temperature for 5 min, cells were
suspended in complete medium. The above suspension was transferred
to a culture flask precoated with 1% fibronectin (Sigma-Aldrich)
and cultured at 37°C in a 5% CO2 humidified incubator.
The medium was changed every 2.5 days until all cells formed
confluent monolayers. Cells were passaged at a split ratio of
1:2.
Cell identification
Rat RVECs were seeded (7×103 cells) in
12-well plates containing coverslips for 24 h at 37°C and then
fixed using 4% paraformaldehyde for 10 min at room temperature and
washed in triplicate with PBS. Samples were then blocked with
normal goat serum (1:50; cat. no. ab7481; Abcam, Cambridge, UK) for
15 min at room temperature. Subsequently cell slides were incubated
with primary antibodies (anti-CD31; cat. no. ab28364; 1:200; Abcam)
at 37°C for 2 h. Samples were washed with PBS in triplicate and
probed with Goat anti-Mouse immunoglobulin G H&L-Cy3 (cat. no.
SA00009-1; 1:1,000; ProteinTech Group, Inc.) at 37°C for 1 h.
Hoechst 33258 (Beyotime Institute of Biotechnology, Haimen, China)
was used to stain nuclei and images were captured using a
fluorescence microscope (magnification, ×50; Olympus Corporation,
Tokyo, Japan) at room temperature for 15 min. Cell number analysis
was performed using Image J 1.45 software (National Institutes of
Health, Bethesda, MD) as described previously (25). The number of positive cells (with red
coloration) and blue stained nuclei were counted separately.
Positive rate was determined to be the number of positive red
stained cells/the number of blue stained nuclei.
Drug treatment
Rat RVECs from each group were plated in 6-well
plates containing 2 ml of complete DMEM medium at a density of
2×105 cells/well. Following cell culture for 24 h at
37°C, medium was exchanged for fresh complete DMEM supplemented
with PD98059 (25 µmol/l; Merck KGaA), U46619 (80 µmol/l; Merck
KGaA), PD98059 (25 µmol/l) in combination with carnosine (1%; cat.
no. C9625; Sigma-Aldrich; Merck KGaA), and U46619 (80 µmol/l) in
combination with carnosine (1%). The control group was treated with
equal quantities of deionized solvent (DMSO or ethanol). The
expression of PKC, ERK and p-ERK in each group was detected 24 h
following treatment.
Retinal immunofluorescence
staining
Of the 25 rats used in the current study, a total of
15 rats (weight, 235–245 g) were randomly divided into 3 groups
(n=5 in each group): A Diabetes mellitus (DM) group, a DM+
carnosine (100 mg/kg/d by gavage for 10 days) group and a control
group. Retinas were isolated from the DM group, the DM+ carnosine
group and the control group, and fixed in 4% para-formaldehyde
solution at room temperature for 15 min. Retinas were subsequently
sectioned (0.3 mm thick), stained with CD31 antibodies and Hochest
as aforementioned and examined using fluorescence microscopy
(magnification, ×50; Olympus Corporation).
RT-qPCR
RT-qPCR analysis was performed as described
previously (26). Total cellular RNA
was extracted from rat retinas using a TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) and cDNA was prepared using the
M-MLV reverse transcriptase kit (Promega Corporation, Madison, WI)
with random primers according to the manufacturer's protocol. The
thermocycling conditions for PCR were as follows: Pre-denaturation
for 10 min at 94°C, denaturation for 20 sec at 94°C, annealing for
20 sec at 55°C, and extension for 20 sec at 72°C. Each PCR reaction
was performed for 40 cycles. RT-qPCR analysis was performed using a
Biosystems StepOne™ real-time PCR apparatus (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using standard procedures and
analyzed using ABI Prism 7500 SDS software (Applied Biosystems;
Thermo Fisher Scientific, Inc.). A Fast SYBR-Green Master Mix was
obtained from Thermo Fisher Scientific, Inc. Data were presented as
the relative expression following normalization to the expression
of GAPDH using the 2−∆∆Cq method (27). The PCR primer sequences were as
follows GAPDH forward, 5′-ACAGCAACAGGGTGGTGGAC-3′ and reverse,
5′-TTTGAGGGTGCAGCGAACTT-3′; PKC forward (5′-3′),
5′-TGGAGTCCTGCTGTATGAGATGTTG-3′ and reverse,
5′-CGCTTTCCTGGGTGCTTGG-3′; ERK forward, 5′-GAAAGCATTACCTTGACCAG-3′
and reverse, 5′-CTTTGGAGTCAGCATTTGG-3′.
Western blotting
The procedure for western blotting was performed as
described previously (28). Total
cellular protein was extracted using radioimmunoprecipitation assay
lysis buffer containing phenylmethylsulfonyl fluoride Protease
Inhibitor (Sigma-Aldrich; Merck KGaA). A BCA Protein Assay kit was
utilized for protein determination (cat. no. 23225; Thermo Fisher
Scientific). Proteins (30 µg) were resolved in 5 or 12% SDS-PAGE
gels and then transferred onto ployvinylidene difluoride (PVDF)
membranes following electrophoresis. Subsequently, PVDF membranes
were blocked using 5% skimmed milk in TBST (20 mM Tris, pH 7.6, 150
mM NaCl, 0.1% Tween-20; wt/vol) for 2 h at room temperature.
Membranes were immunoblotted with Mouse anti-PKC (cat. no. P5704;
1:1,000; Sigma-Aldrich; Merck KGaA), ERK1/2 Rabbit Polyclonal (cat.
no. 16443-1-AP; 1:1,000; ProteinTech Group, Inc.) and anti-p-ERK
antibodies (cat. no. ab65142; 1:1,000; Abcam) at 4°C overnight.
Samples were then incubated with IRDyeTM-800 conjugated anti-mouse
(cat. no. P/N 925-32210; 1:10,000; Li-COR Biosciences, Lincoln, NE)
and IRDye® 800CW Goat anti-Rabbit IgG (H+L)secondary
antibodies (cat. no. P/N 925-32211; 1:10,000; Li-COR Biosciences)
for 30 min at room temperature. The specific protein bands were
detected using Odyssey Infrared Imaging System (Li-COR Biosciences)
and analyzed with Image Studio™ Software 4.0 (Li-COR Biosciences).
The quantification of each protein was normalized to β-actin.
Statistical analysis
All data were expressed as the mean ± standard error
of the mean and analyzed using GraphPad Prism5 software (GraphPad
Software, Inc., La Jolla, CA). One-way analysis of variance
followed by a Tukey's test was used to analyze the significant
differences between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
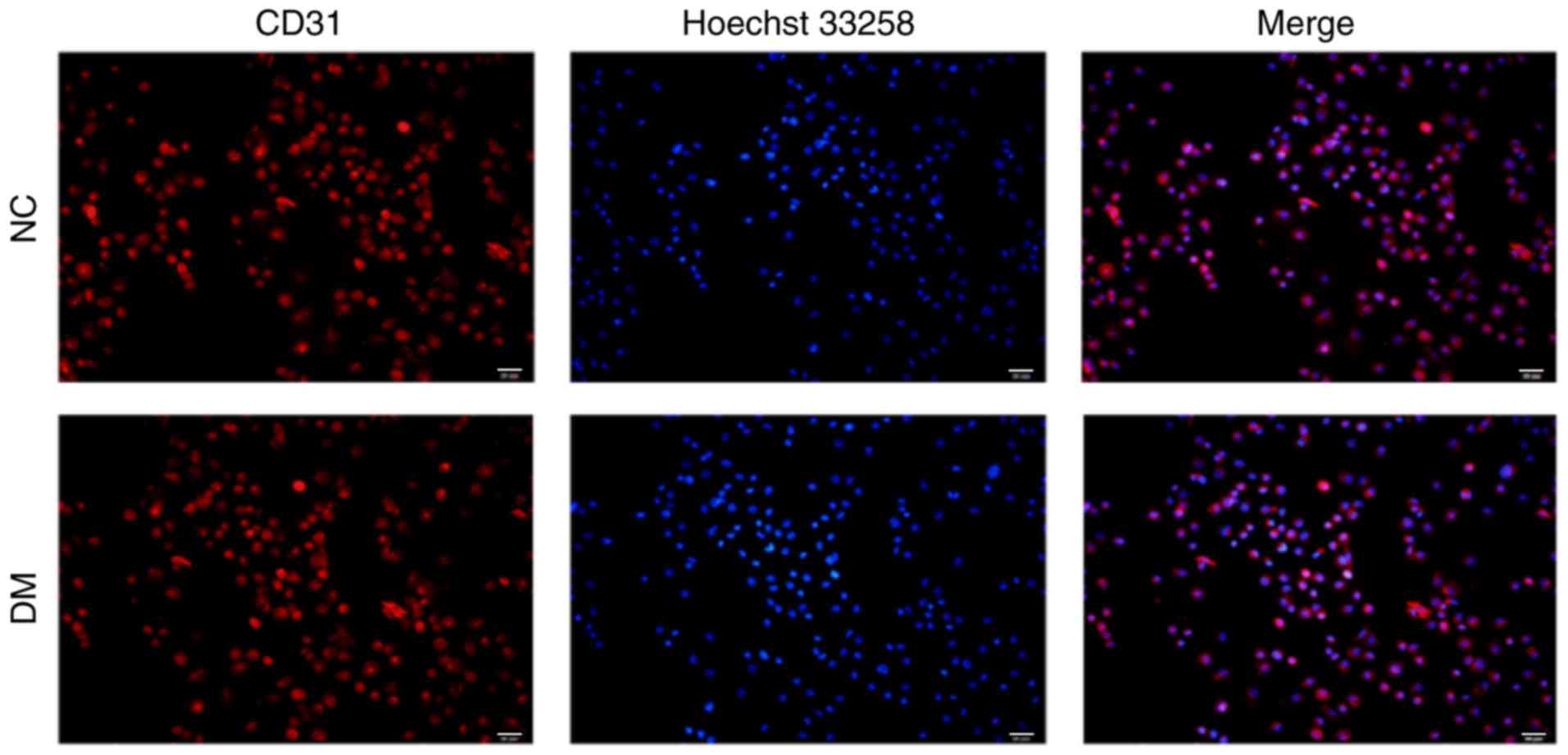
RVEC determination
To determine whether the isolated and cultivated
cells were RVECs, cells were fixed with 4% paraformaldehyde and
subsequently stained with CD31, a specific marker of endothelial
cells. As presented in Fig. 1,
>90% (Data not shown) of the isolated cells stained positive for
CD31, thereby confirming that they were RVECs.
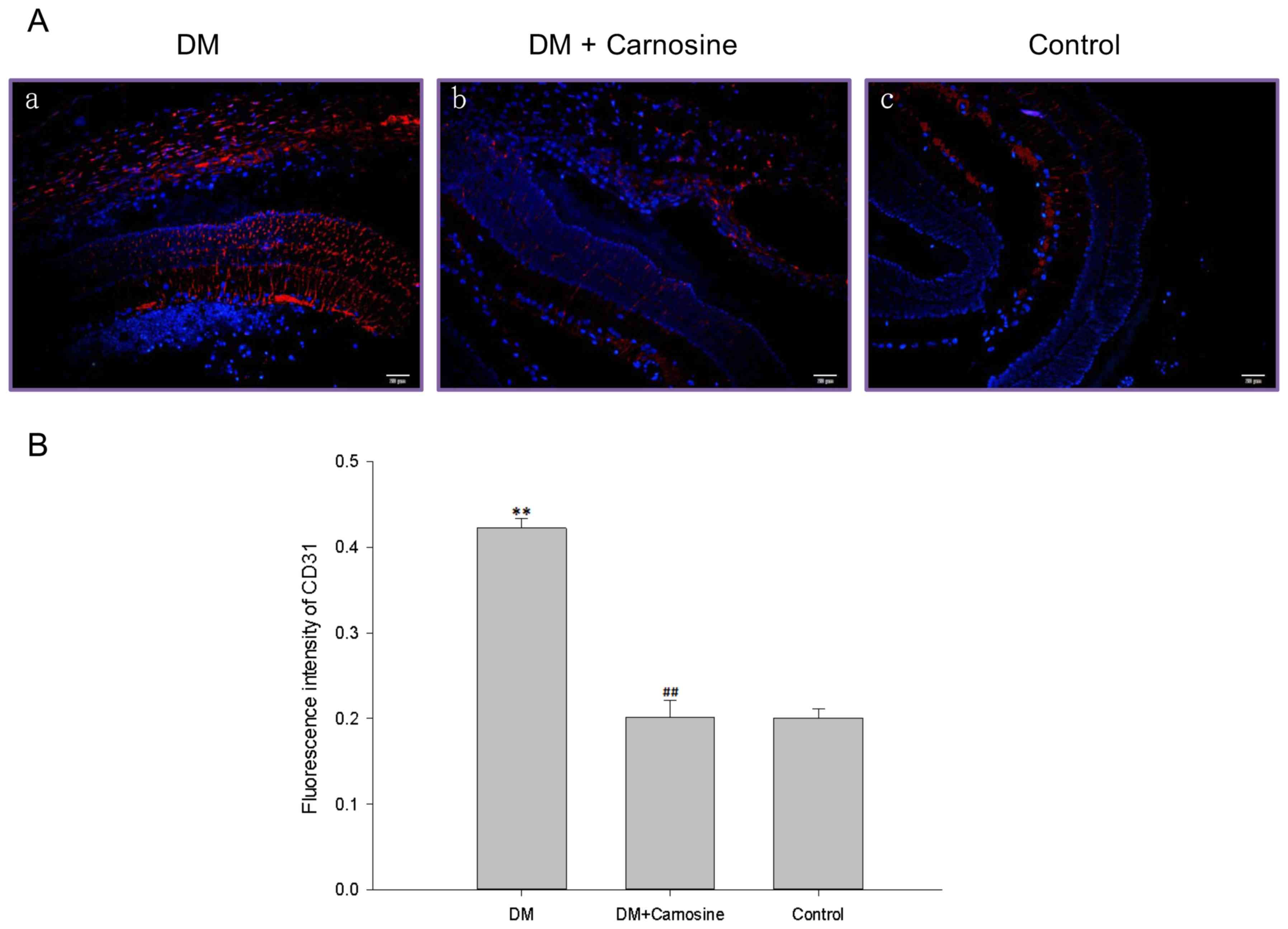
Carosine inhibited DM-caused retinal
overvascularization
CD31 immunostaining revealed that the density of
retinal microvessels were significantly increased in DM rats
compared with control rats and carosine markedly inhibited this
increase of density in DM rats (Fig.
2).
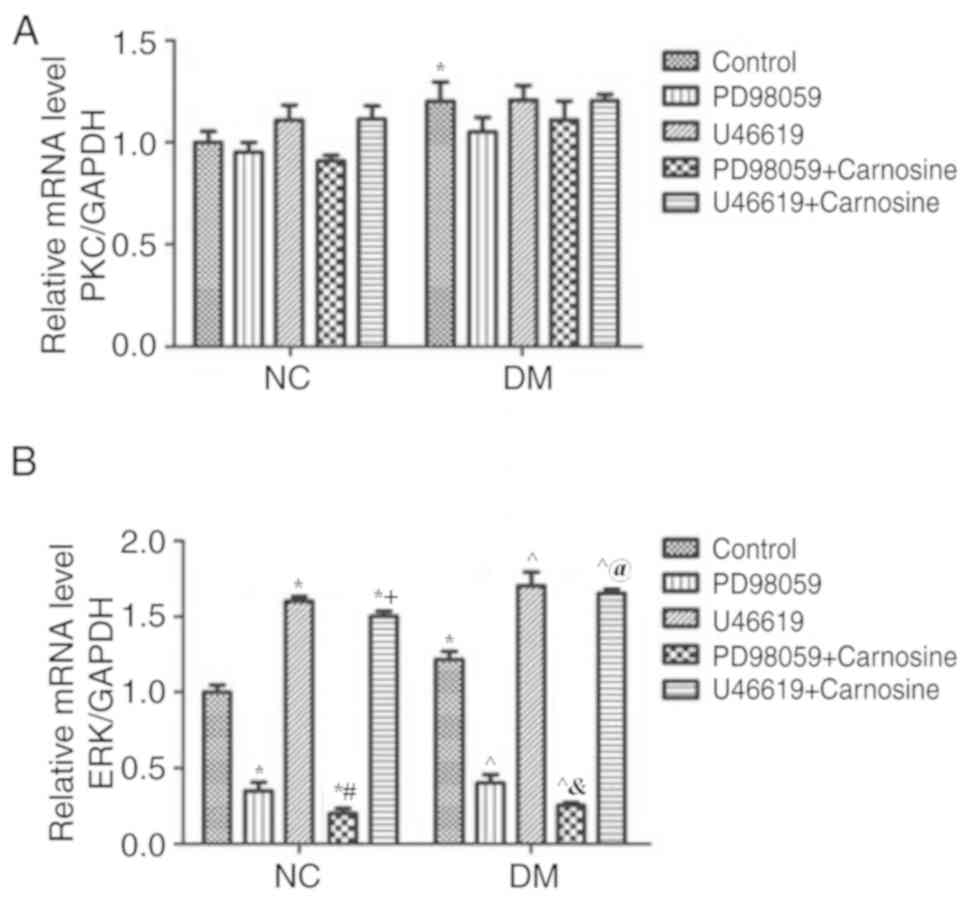
The MAPK/ERK pathway is involved in
the prevention and treatment of DR, as caused by carnosine
Changes of PKC, ERK and p-ERK expression in diabetic
rats following treatment with carnosine, PD98059 and U46619 were
assessed via RT-qPCR and western blotting. The mRNA levels of PKC
(Fig. 3A) and ERK (Fig. 3B) of diabetic RVECs were
significantly higher than those of normal RVECs. However, PKC mRNA
levels in normal or diabetic RVECs were not substantially impacted
by PD98059 and U46619 with or without carosine (Fig. 3A). Furthermore, ERK mRNA levels of
RVECs was increased by U46619 and reduced by PD98059 and these
changes were further enhanced by carosine (Fig. 3B).
The protein levels of PKC, ERK, and p-ERK in RVECs
after treatment with PD98059, U46619, and carnosine were assessed
via western blotting. Levels of PKC, ERK and phosphorylated ERK
protein were significantly increased in diabetic RVECs compared
with normal RVECs (Fig. 4A-D).
PD98059 and U46619 increased the PKC protein level of normal RVECs
but had little effect on the PKC protein level of diabetic RVECs.
Furthermore, carosine inhibited PD98059 but augmented the U46619
induced increase of PKC. However, it did not effect the PKC levels
of diabetic RVECs (Fig. 4A and B).
ERK and phosphorylated ERK levels were reduced by PD98059 and
increased by U46619 in normal and diabetic RVECs. Carosine
treatment reduced ERK and phosphorylated ERK levels of normal and
diabetic RVECs treated with PD98059 and U46619. Normal RVECs
treated with U46619 however, were not affected (Fig. 4A, C and D). Taken together, these
data suggest that the MAPK/ERK signaling pathway, rather than the
PKC pathway, mediates carnosine-induced microvascular changes in
DR.
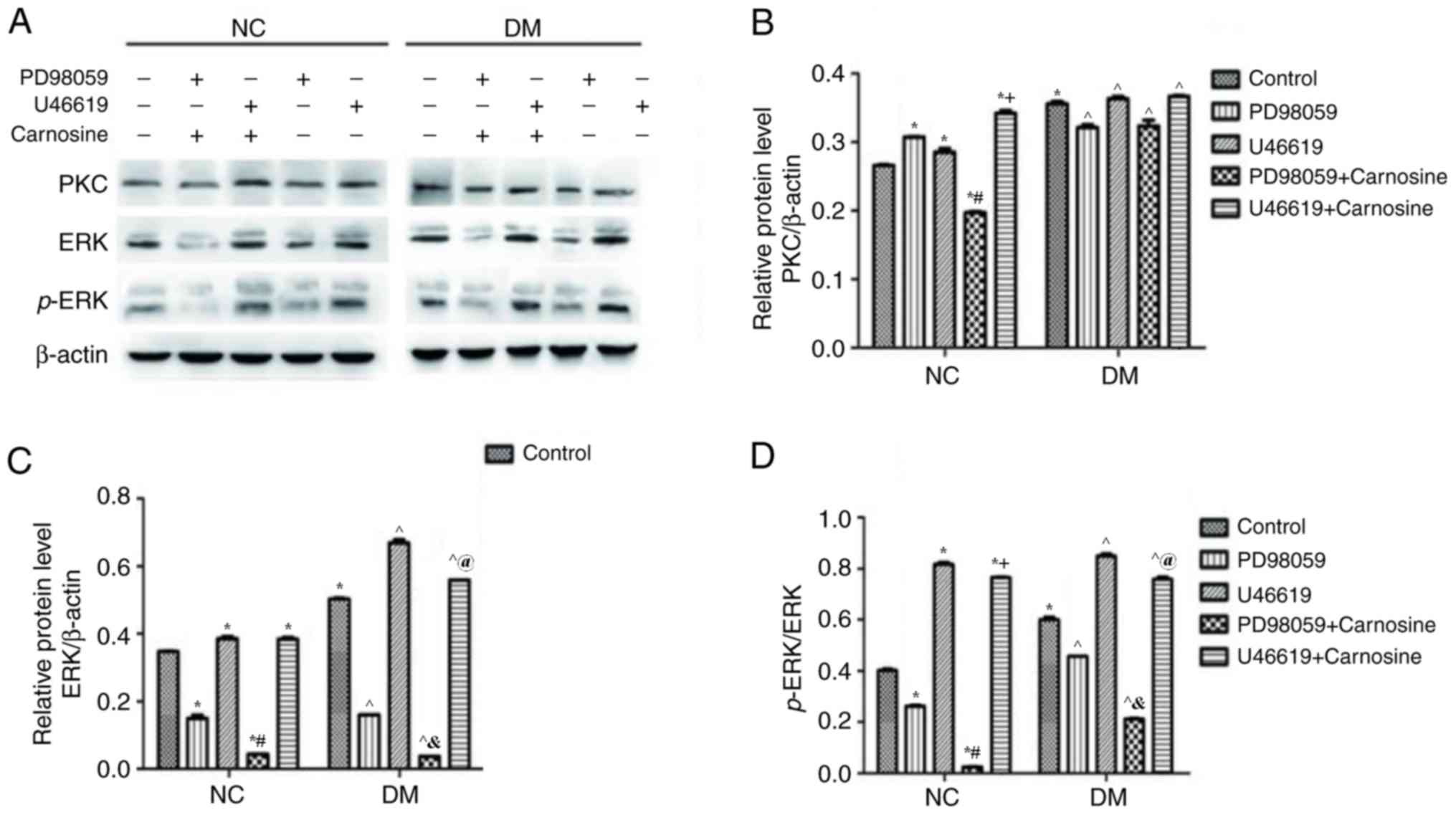 | Figure 4.Carnosine regulates the level of
PKC/ERK/p-ERK proteins in RVECs. (A) Western blotting determined
the expression of PKC, ERK and p-ERK following treatment with or
without PD98059, U46619 and carnosine for 24 h (A). The
quantification of (B) PKC, (C) ERK, (D) p-ERK and (E) ERK/p-ERK
expression was determined by normalizing to β-actin. Data are
presented as the mean ± standard error of the mean from 3
independent experiments. *P<0.05 vs. NC Control;
#P<0.05 vs. NC PD98059; +P<0.05 vs.
U46619; ^P<0.05 vs. DM Control;
&P<0.05 vs. DM PD98059; @P<0.05 vs.
DM U46619. PKC, protein kinase C; ERK, extracellular signal related
kinase; p, phosphorylated; RVEC, retinal vascular endothelial
cells; NC, negative control; DM, diabetes mellitus. |
Discussion
Carnosine (β-Ala-l-His) is an imidazole dipeptide
that has many biological functions. Numerous studies have
demonstrated that the carnosine-carnosinase system serves a key
role in the pathogenesis of diabetes and that carnosine may be used
as a novel therapeutic agent against type 2 diabetes (10,29,30).
Cripps et al (10) revealed
that following treatment with carnosine, insulin secretion from
isolated mouse islets or insulinoma cell line β-cells was
increased. They further revealed that the inhibition of insulin
secretion caused by glucolipotoxic was reversed and that glucose
uptake into skeletal muscle cells was enhanced. AGEs and oxidative
stress serve a role in the development of diabetic complications
(31). It has been demonstrated that
carnosine can treat diabetic complications by decreasing oxidation
and glycation products in the serum and livers of diabetic rats
(30).
Visual impairment as a result of DR has a
significant negative impact on patient quality of life and their
ability to successfully manage their disease (32). Due to its asymptomatic onset and
progressive disease course, identifying an effective treatment of
DR is clinically significant. Pfister et al (33) determined that treatment with oral
carnosine prevents vascular damage in experimental DR, independent
of its biochemical function, which includes reactive oxygen species
(ROS) or advanced glycation end (AGE) inhibition (33).
The MAPK/ERK signaling pathway has been determined
to serve an important role in various pathogenetic pathways and
physiological processes, including the regulation of cell cycle
entry and proliferation, as well as in the development of DR by
upregulating chronic inflammation (34). The MAPK/ERK signaling pathway
includes various MAPKs, including ERKs, which integrate multiple
biochemical and environmental signals (including Ras activation and
the kinase cascade) via phosphorylation cascades (35). Gogg et al (36) demonstrated that the phosphorylation
of ERK1/2 was significantly upregulated in Type 2 diabetes. A
previous study revealed that after the degradation of the insulin
receptor substrate (IRS)-1 protein, p-MAPK levels are upregulated
following the increase of ERK1/2 phosphorylation in diabetes
(37). However, the basic
phosphorylation of ERK1/2 has also been demonstrated to be
upregulated in adipose cells in type 2 diabetes (38). Additionally, IRS-1 has also been
revealed to be decreased (39).
ERK is strongly activated in primary neuroretinal
tissue and has been determined to be a neuroprotective regulator in
the neuroretina, which is involved in the maintenance of the
neuroretina- retinal pigment epithelium interaction (40,41). It
has been reported that the activation of ERK serves an important
role in the maintenance of the inner blood-retinal barrier, similar
to that of the RPE outer blood-retinal barrier (42). Van Dijk et al (43) demonstrated that the reversible
suppression of ERK activity leads to disturbances in the
neuroretina-RPE interaction and in the occurrence of SRF, which
results in retinopathy.
The glycation of proteins and the formation of AGEs
serve an important role in DM and DR. An increase in AGE level has
been identified in the retinas of patients with diabetes and has
been determined to be positively correlated with the increase of
blood glucose and the development of DR (44). Previous studies have demonstrated
that the combination of AGE and the AGE receptor causes the
activation of the MAPK/ERK signaling pathway, the activation of the
oxidative stress response and the overexpression of certain
cytokines (including inflammatory cytokines, lymphocyte adhesion
molecules, vascular regulators and coagulants), which may lead to
retinal lesions and vascular endothelial cell damage in
vitro (45,46). The current study revealed that the
expression of p-ERK/ERK significantly changed following treatment
with PD98059, U46619 and carnosine, respectively or in combination.
The current study therefore hypothesized that the protective effect
of carnosine in DR may be associated with AGEs and the MAPK/ERK
signaling pathway. However, further studies are required to clarify
how the interaction between carnosine and the MAPK/ERK signaling
pathway improves DR.
A previous study has determined that the activity of
PKC, MEK and ERK1/2 is significantly increased in glomerular
mesangial cells in STZ-induced and glucose-induced diabetic rats,
and that the activation of MEK and ERK1/2 is PKC-dependent
(46). The results of the current
study revealed that there was almost no effect on the expression of
PKC following treatment with PD98059, U46619 and carnosine,
respectively or in combination, indicating that the suppression of
the PKC-independent MAPK/ERK signaling pathway, rather than the PKC
pathway, may serve an important role in the prevention and
treatment of carnosine-induced DR.
In summary, the present study demonstrated that the
MAPK/ERK signaling pathway may mediate carnosine-induced
microvascular changes in DR.
Acknowledgements
Not applicable.
Funding
The current study was supported a grant obtained
from the Innovation fund of the Air Force Medical University (grant
no. 2014JCR010).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YG designed the current study, wrote the manuscript
and established the diabetic rat model. CG performed western
blotting and statistical analysis and RT-qPCR. WH performed retinal
immunofluorescence staining, drug treatment and cell
identification. ZD performed RVEC isolation and cell culture.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the recommendations included in the Guide for the Care and Use
of Laboratory Animals of the National Institutes of Health.
Furthermore, the protocol of the current study was approved by the
Committee on the Ethics of Animal Experiments of Tangdu Hospital,
Air Force Medical University.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Alotaibi A, Perry L, Gholizadeh L and
Al-Ganmi A: Incidence and prevalence rates of diabetes mellitus in
Saudi Arabia: An overview. J Epidemiol Glob Health. 7:211–218.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mrozikiewicz-Rakowska B, Łukawska M,
Nehring P, Szymański K, Sobczyk-Kopcioł A, Krzyżewska M, Maroszek
P, Płoski R and Czupryniak L: Genetic predictors associated with
diabetic retinopathy in patients with diabetic foot. Pol Arch
Intern Med. 128:35–42. 2018.PubMed/NCBI
|
|
3
|
Tan GS, Cheung N, Simó R, Cheung GC and
Wong TY: Diabetic macular oedema. Lancet Diabetes Endocrinol.
5:143–155. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tolentino MS, Tolentino AJ and Tolentino
MJ: Current and investigational drugs for the treatment of diabetic
retinopathy. Expert Opin Investig Drugs. 25:1011–1022. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brownlee M, Aiello LP, Cooper ME, Vinik
AI, Plutzky J and Boulton AJ: Complications of diabetes mellitus.
In Williams Textbook of Endocrinology (Thirteenth Edition)
Elsevier. 1484–1581. 2017.
|
|
6
|
Happich M, Breitscheidel L, Meisinger C,
Ulbig M, Falkenstein P, Benter U and Watkins J: Cross-sectional
analysis of adult diabetes type 1 and type 2 patients with diabetic
microvascular complications from a German retrospective
observational study. Curr Med Res Opin. 23:1367–1374. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brownlee M: Biochemistry and molecular
cell biology of diabetic complications. Nature. 414:813–820. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ramasamy R, Vannucci SJ, Yan SS, Herold K,
Yan SF and Schmidt AM: Advanced glycation end products and RAGE: A
common thread in aging, diabetes, neurodegeneration, and
inflammation. Glycobiology. 15:16R–28R. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Katakura Y, Totsuka M, Imabayashi E,
Matsuda H and Hisatsune T: Anserine/carnosine supplementation
suppresses the expression of the inflammatory chemokine CCL24 in
peripheral blood mononuclear cells from elderly people. Nutrients.
9:E11992017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cripps MJ, Hanna K, Lavilla C Jr, Sayers
SR, Caton PW, Sims C, De Girolamo L, Sale C and Turner MD:
Carnosine scavenging of glucolipotoxic free radicals enhances
insulin secretion and glucose uptake. Sci Rep. 7:133132017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baye E, Ukropcova B, Ukropec J, Hipkiss A,
Aldini G and de Courten B: Physiological and therapeutic effects of
carnosine on cardiometabolic risk and disease. Amino Acids.
48:1131–1149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tan RR, Li YF, Zhang SJ, Huang WS, Tsoi B,
Hu D, Wan X, Yang X, Wang Q, Kurihara H and He RR: Abnormal
O-GlcNAcylation of Pax3 occurring from hyperglycemia-induced neural
tube defects is ameliorated by carnosine but not folic acid in
chicken embryos. Mol Neurobiol. 54:281–294. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li YF, He RR, Tsoi B, Li XD, Li WX, Abe K
and Kurihara H: Anti-stress effects of carnosine on
restraint-evoked immunocompromise in mice through spleen lymphocyte
number maintenance. PLoS One. 7:e331902012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Albrecht T, Schilperoort M, Zhang S, Braun
JD, Qiu J, Rodriguez A, Pastene DO, Krämer BK, Köppel H, Baelde H,
et al: Carnosine attenuates the development of both type 2 diabetes
and diabetic nephropathy in BTBR ob/ob mice. Sci Rep. 7:444922017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Javadi S, Yousefi R, Hosseinkhani S,
Tamaddon AM and Uversky VN: Protective effects of carnosine on
dehydroascorbate-induced structural alteration and opacity of lens
crystallins: Important implications of carnosine pleiotropic
functions to combat cataractogenesis. J Biomol Struct Dyn.
35:1766–1784. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Klein BE: Overview of epidemiologic
studies of diabetic retinopathy. Ophthalmic Epidemiol. 14:179–183.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Coleman SK, Rebalka IA, D'Souza DM and
Hawke TJ: Skeletal muscle as a therapeutic target for delaying type
1 diabetic complications. World J Diabetes. 6:1323–1336. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cargnello M and Roux PP: Activation and
function of the MAPKs and their substrates, the MAPK-activated
protein kinases. Microbiol Mol Biol Rev. 75:50–83. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pearson G, Robinson F, Beers Gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: Regulation and physiological functions.
Endocr Rev. 22:153–183. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cervellini I, Galino J, Zhu N, Allen S,
Birchmeier C and Bennett DL: Sustained MAPK/ERK activation in adult
schwann cells impairs nerve repair. J Neurosci. 38:679–690. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Danquah A, de Zelicourt A, Colcombet J and
Hirt H: The role of ABA and MAPK signaling pathways in plant
abiotic stress responses. Biotechnol Adv. 32:40–52. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hirata K and Kubo K: Relationship between
blood levels of N-carboxymethyl-lysine and pentosidine and the
severity of microangiopathy in type 2 diabetes. Endocr J.
51:537–544. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schaaf G, Dynowski M, Mousley CJ, Shah SD,
Yuan P, Winklbauer EM, de Campos MK, Trettin K, Quinones MC,
Smirnova TI, et al: Resurrection of a functional
phosphatidylinositol transfer protein from a pseudo-Sec14 scaffold
by directed evolution. Mol Biol Cell. 22:892–905. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maas M, Wang R, Paddock C, Kotamraju S,
Kalyanaraman B, Newman PJ and Newman DK: Reactive oxygen species
induce reversible PECAM-1 tyrosine phosphorylation and SHP-2
binding. Am J Physiol Heart Circ Physiol. 285:H2336–H2344. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Grishagin IV: Automatic cell counting with
ImageJ. Anal Biochem. 473:63–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mao L, Wang H, Ma F, Guo Z, He H, Zhou H
and Wang N: Exposure to static magnetic fields increases insulin
secretion in rat INS-1 cells by activating the transcription of the
insulin gene and up-regulating the expression of vesicle-secreted
proteins. Int J Radiat Biol. 93:831–840. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liao XH, Wang N, Liu QX, Qin T, Cao B, Cao
DS and Zhang TC: Myocardin-related transcription factor-A induces
cardiomyocyte hypertrophy. IUBMB Life. 63:54–61. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
DeLisser HM, Newman PJ and Albelda SM:
Molecular and functional aspects of PECAM-1/CD31. Immunol Today.
15:490–495. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aydın AF, Bingül İ, Küçükgergin C,
Doğan-Ekici I, Doğru Abbasoğlu S and Uysal M: Carnosine decreased
oxidation and glycation products in serum and liver of high-fat
diet and low-dose streptozotocin-induced diabetic rats. Int J Exp
Pathol. 98:278–288. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Goldin A, Beckman JA, Schmidt AM and
Creager MA: Advanced glycation end products: Sparking the
development of diabetic vascular injury. Circulation. 114:597–605.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hendrick AM, Gibson MV and Kulshreshtha A:
Diabetic retinopathy. Prim Care. 42:451–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pfister F, Riedl E, Wang Q, vom Hagen F,
Deinzer M, Harmsen MC, Molema G, Yard B, Feng Y and Hammes HP: Oral
carnosine supplementation prevents vascular damage in experimental
diabetic retinopathy. Cell Physiol Biochem. 28:125–136. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang L, Zhang ZK and Liang S:
Epigallocatechin-3-gallate protects retinal vascular endothelial
cells from high glucose stress in vitro via the MAPK/ERK-VEGF
pathway. Genet Mol Res. 15:2016.
|
|
35
|
Hu L, Zhang Y, Chen L, Zhou W, Wang Y and
Wen J: MAPK and ERK polymorphisms are associated with PCOS risk in
Chinese women. Oncotarget. 8:100261–100268. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gogg S, Smith U and Jansson PA: Increased
MAPK activation and impaired insulin signaling in subcutaneous
microvascular endothelial cells in type 2 diabetes: The role of
endothelin-1. Diabetes. 58:2238–2245. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aguirre V, Werner ED, Giraud J, Lee YH,
Shoelson SE and White MF: Phosphorylation of Ser307 in insulin
receptor substrate-1 blocks interactions with the insulin receptor
and inhibits insulin action. J Biol Chem. 277:1531–1537. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Carlson CJ, Koterski S, Sciotti RJ,
Poccard GB and Rondinone CM: Enhanced basal activation of
mitogen-activated protein kinases in adipocytes from type 2
diabetes: Potential role of p38 in the downregulation of GLUT4
expression. Diabetes. 52:634–641. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rondinone CM, Wang LM, Lonnroth P, Wesslau
C, Pierce JH and Smith U: Insulin receptor substrate (IRS) 1 is
reduced and IRS-2 is the main docking protein for
phosphatidylinositol 3-kinase in adipocytes from subjects with
non-insulin-dependent diabetes mellitus. Proc Natl Acad Sci USA.
94:4171–4175. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ulbrich F, Kaufmann KB, Coburn M, Lagrèze
WA, Roesslein M, Biermann J, Buerkle H, Loop T and Goebel U:
Neuroprotective effects of Argon are mediated via an ERK-1/2
dependent regulation of heme-oxygenase-1 in retinal ganglion cells.
J Neurochem. 134:717–727. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marra C, Gomes Moret D, de Souza Corrêa A,
Chagas da Silva F, Moraes P, Linden R and Sholl-Franco A: Protein
kinases JAK and ERK mediate protective effect of interleukin-2 upon
ganglion cells of the developing rat retina. J Neuroimmunol.
233:120–126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
LoRusso PM, Krishnamurthi SS, Rinehart JJ,
Nabell LM, Malburg L, Chapman PB, DePrimo SE, Bentivegna S, Wilner
KD, Tan W and Ricart AD: Phase I pharmacokinetic and
pharmacodynamic study of the oral MAPK/ERK kinase inhibitor
PD-0325901 in patients with advanced cancers. Clin Cancer Res.
16:1924–1937. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
van Dijk EH, Duits DE, Versluis M, Luyten
GP, Bergen AA, Kapiteijn EW, de Lange MJ, Boon CJ and van der
Velden PA: Loss of MAPK pathway activation in post-mitotic retinal
cells as mechanism in MEK inhibition-related retinopathy in cancer
patients. Medicine (Baltimore). 95:e34572016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wautier JL and Guillausseau PJ: Advanced
glycation end products, their receptors and diabetic angiopathy.
Diabetes Metab. 27:535–542. 2001.PubMed/NCBI
|
|
45
|
Genuth S, Sun W, Cleary P, Sell DR, Dahms
W, Malone J, Sivitz W, Monnier VM and DCCT Skin Collagen Ancillary
Study Group: Glycation and carboxymethyllysine levels in skin
collagen predict the risk of future 10-year progression of diabetic
retinopathy and nephropathy in the diabetes control and
complications trial and epidemiology of diabetes interventions and
complications participants with type 1 diabetes. Diabetes.
54:3103–3111. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Xu T, Wang NS, Fu LL, Ye CY, Yu SQ and Mei
CL: Celecoxib inhibits growth of human autosomal dominant
polycystic kidney cyst-lining epithelial cells through the
VEGF/Raf/MAPK/ERK signaling pathway. Mol Biol Rep. 39:7743–7753.
2012. View Article : Google Scholar : PubMed/NCBI
|