Introduction
An ideal dental implant requires adequate alveolar
ridge dimensions following tooth extraction. In recent years,
alveolar bone regeneration has been widely investigated by filling
the extraction socket with bone substitutes or growth factors to
protect alveolar bone against resorption. These substitutes include
deproteinized bovine bone (Bio-Oss) particles (1), deproteinized bovine bone and collagen
matrix Bio-Oss Collagen (2) and
demineralized freeze-dried bone allograft (3). The growth factors include recombinant
human bone morphogenetic protein-2 (4), recombinant human platelet-derived
growth factor (5) and plasma-rich
growth factor (6), among others.
These growth factors exert different effects during the bone
formation, such as inflammatory reactions, soft and hard callus
formation, and bone remodelling. Despite the osteoinductive agents
of these growth factors, their use in bone regeneration are limited
due to the drawbacks, including osteolysis, heterotopic
ossification, easy degradation, safety and efficiency (7,8). As a
result, they are far from being ready for extensive clinical use. A
new treatment aim for the preservation of the alveolar ridge
includes how to achieve optimal osteoinduction effects and provide
sufficient nutrition and oxygen to the osteoblasts during the
process of new bone formation.
Molecular oxygen is essential for metabolic
processes, including oxidative phosphorylation, in which oxygen
serves as an electron acceptor during ATP formation (9). Lack of oxygen may result in failure to
generate sufficient ATP to maintain essential cellular functions.
Hypoxia-inducible factor 1 (HIF-1), which is composed of HIF-1α and
HIF-1β, is a heterodimeric basic helix-loop-helix transcription
factor that regulates hypoxia-inducible genes, including the human
erythropoietin (EPO) gene (10).
HIF-1α is not expressed in normal cells, but can be induced under
hypoxic conditions; it acts as one of the key transcription factors
that regulate numerous downstream responses to hypoxia, including
the expression of several angiogenic factors (11). Previous findings have demonstrated
that hypoxia transcriptionally upregulates the expression of
HIF-1α, vascular endothelial growth factor (VEGF), transforming
growth factor (TGF)-β and insulin-like growth factor-2, which may
serve an important role in angiogenesis during osteogenesis
(12). It has long been recognized
that angiogenesis is essential for osteogenesis. Under hypoxic
conditions, the HIF-1α protein is stabilized as it dimerizes with
HIF-1β and binds to the hypoxia response elements in target gene
promoter sequences (13). The
oxygen-dependent prolyl hydroxylases, which act upon HIF-1α, are
members of the same protein family that contains the
procollagenprolyl 4-hydroxylase, an enzyme essential for collagen
formation (14). However, the
application of HIF-1α for bone regeneration remains controversial
due to its seemingly contradictory dual role in osteoblast
differentiation.
Endochondral bone formation and fracture repair are
dependent upon blood vessel invasion (1) and exercise-induced bone formation,
which is associated with increased blood supply. However, the
effect of oxygenation on the function of osteoblasts has not been
extensively investigated to date. Previous results have
demonstrated that VEGF, a potent angiogenic peptide with specific
mitogenic and chemotactic properties, has a key role in
angiogenesis in cartilage and temporomandibular joint
osteoarthritis (2). As an important
transcription factor in angiogenesis, HIF-1α is one of the most
important regulators of VEGF expression. Therefore, it was
hypothesized that the application of HIF-1α in tooth extraction
sites may exert osteoinductive and angiogenic effects during the
bone healing process. Bio-Oss has been extensively examined in
vitro and in vivo in a variety of clinical and
preclinical studies (15–17). To examine the function of HIF-1α in
the preservation of the alveolar ridge in a dog model, HIF-1α was
applied to the tooth extraction sites by using Bio-Oss as the
protein carrier.
Materials and methods
Preparation of HIF-1α protein
carrier
Bio-Oss (Geistlich Pharma AG, Wolhusen, Switzerland)
was used as a carrier for the HIF-1α protein (AnaSpec Inc.,
Fremont, CA, USA). Pieces of Bio-Oss, 1 mm in diameter, were
submerged in a concentration of 100 mg/ml HIF-1α protein prior to
use.
Animal protocol
Six beagle dogs (age, 1.5 years; weight, 15–20 kg)
were obtained from the Animal Centre of the Ninth People's
Hospital, Shanghai Jiao Tong University. The Institutional Animal
Care and Use Committee of the Ninth People's Hospital, Shanghai
Jiao Tong University, approved the protocol used in the present
study (ethics approval number, 2013-304). All animals were handled
according to the guidelines established by the Institutional Animal
Care Committee.
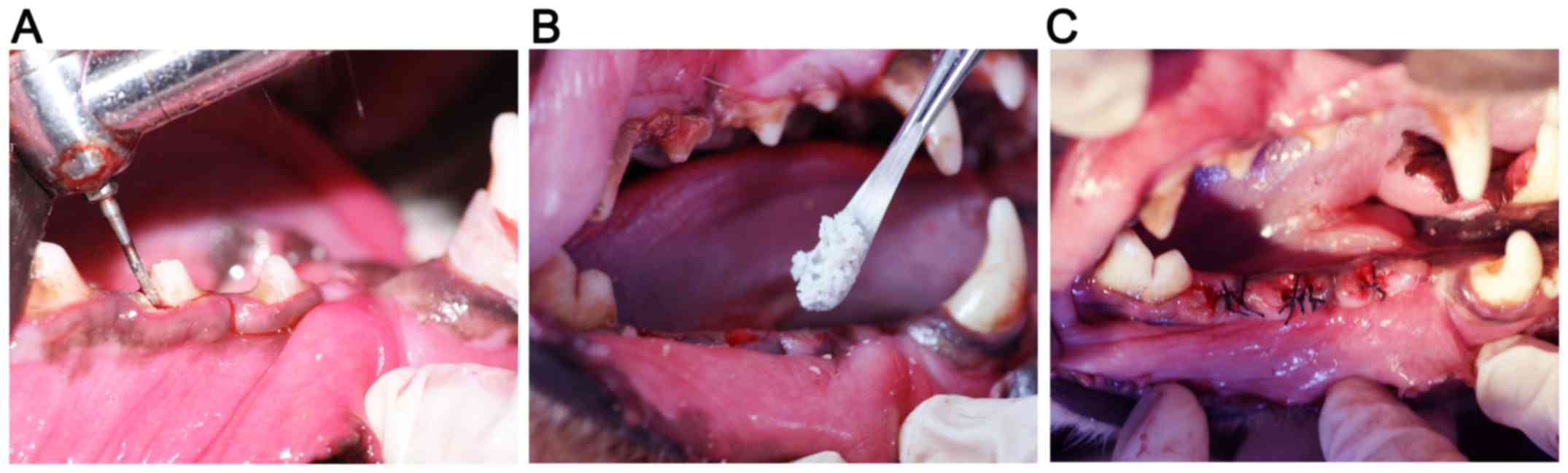
Surgical procedures
Each animal was anesthetized with an intramuscular
injection of ketamine (10 mg/kg; Fujian Gutian Pharma Co., Ltd.,
Fujian, China) and xylazine (1 mg/kg; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). According to a previous study (18), the second and fourth premolar teeth
of the lower jaws were carefully extracted following vertical tooth
separation. Two randomly selected extraction sockets on one side
were filled with Bio-Oss alone or Bio-Oss + HIF-1α, while the
contralateral sockets remained unfilled. There were three groups of
extraction sites in the present study: Blank group (no fillings in
the sockets), Bio-Oss group (Bio-Oss filling), and Bio-Oss + HIF-1α
group (Bio-Oss + HIF-1α filling). The wounds were closed following
vertical flap elevation. All the operative procedures were
performed under sterile conditions (Fig.
1A-C).
Sample harvest
At 12 weeks post-surgery, all the animals were
euthanized with intravenous injection of pentobarbital (100 mg/kg;
Sigma-Aldrich; Merck KGaA). The alveolar bone surrounding the
extraction socket was dissected and cut into blocks. Protein and
RNA were isolated from the tissues around the extraction sites. The
bone samples from the extraction sites were fixed in 4%
paraformaldehyde overnight at 4°C and were then dehydrated in
ethanol solution.
Micro-computed tomography (CT)
measurement
Specimens were harvested and assessed with the
microtomographic imaging system (MicroCT-80, Siemens Healthineers,
Erlangen, Germany), as previously reported (19). All micro-CT scans were performed with
the following acquisition parameters: Tube voltage: 80 kV; tube
current: 0.50 mA; scan time: 1,500 msec; tomographic rotation:
360°; a rotation step of 360; and a cool-down time of 2,000 msec.
The setting of the charge-coupled Device (CCD) readout was as
follows: Transaxial, 2048; axial, 2048; and binning, 1. The
settings of the effective scope CCD-FOV were as follows:
Transaxial, 19.03 mm; axial, 19.03 mm; and effective pixel size,
9.29 µm. The morphology of reconstructed mandible was assessed
using an animal micro-CT scanner (Inveon Research Workplace,
Siemens Healthineers). Micro-CT measurements included bone volume
to tissue volume (BV/TV), bone mineral density and trabecular
number of the extraction sockets.
Histological and histomorphometric
observations
Following micro-CT examination, all samples were
decalcified in 10% EDTA for 3 weeks and were then dehydrated in 50
and 70% alcohol for 30 min each. Following paraffin embedding, the
samples were cut into 6-µm sections with a Leica microtome and
mounted on slides (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). For hematoxylin and eosin (H&E) staining, the sections
were deparaffinized and hydrated through a xylene and graded
ethanol series (100, 90, 70 and 50%) for 5 min each, then stained
with hematoxylin for 5 min and eosin for 3 min at room temperature.
The samples were observed under a Leica microscope (Leica
Microsystems GmbH, Wetzlar, Germany).
Immunohistochemical (IHC)
staining
IHC staining was performed to analyze
osteoblast-associated factors. First, bone sections were processed
for antigen retrieval by digestion with 0.05% trypsin at 37°C for
15 min, and then incubated with primary antibodies against
osteocalcin (OCN; 1:200; ab93876, Abcam, Cambridge, UK) and
runt-related transcription factor 2 (Runx2; 1:200; ab23981, Abcam)
overnight at 4°C. Subsequently, the sections were incubated with
the horseradish peroxidase (HRP)-conjugated anti-rabbit secondary
antibodies (1:1,000; ab150077; Abcam, Cambridge, UK) for 1 h at
room temperature, and developed in 3,3′-diaminobenzidine chromogen
(8801-4965-72; Invitrogen; Thermo Fisher Scientific, Inc.). The
sections were counterstained with hematoxylin for 5 min at room
temperature and observed under a light microscope (magnification,
×20; Leica Microsystems GmbH).
Western blotting
For protein extraction, the frozen samples were
transferred to a tube prefilled with beads (Nextadvance, Inc.,
Troy, NY, USA) and immersed with 100 µl 2X SDS lysis buffer (100 mM
Tris-HCl, 4% SDS, 0.1% bromophenol blue, 20% glycerol, supplemented
with DTT and protease inhibitors), lysed via centrifugation at
15,000 × g for 15 min at 4°C using an electronic homogenizer
(Nextadvance, Inc.). The supernatants were then collected and
protein concentrations were measured using a bicinchoninic acid
assay protein assay kit (23225; Thermo Fisher Scientific, Inc.).
Protein was denatured at 95°C for 10 min, then cooled to room
temperature. Samples (10 µl) were resolved on a 10% SDS-PAGE gel
and transferred to polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked with 5%
non-fat dry milk in Tris-buffered saline with Tween 20 for 1 h at
room temperature, and incubated overnight at 4°C with primary
antibodies against HIF-1α (1:800; ab228649, Abcam), OCN (1:800,
ab93876, Abcam), Runx2 (1:1,000; ab23981, Abcam), βactin (1:3,000;
ab8226; Acbam) and osteoblast-specific transcription factor osterix
(OSX; 1:500; SC22538, Santa Cruz Biotechnology, Inc., Dallas, TX,
USA); respectively. Membranes were subsequently washed 3 times
using phosphate buffered saline with Tween 20 and incubated with
the following secondary antibodies for 1 h at room temperature: HRP
conjugated- Goat anti-Rabbit Immunoglobulin G (IgG; 1:8,000;
ab6721; Abcam) and HRP conjugated Goat anti-Mouse IgG (1:5,000;
ab6789; Abcam). Antibody-antigen complexes were detected using the
Luminata Forte HRP substrate (EMD Millipore). Membranes were
visualized and quantified using a blot scanner (LI-COR Biosciences,
Lincoln, NE, USA) and Image Studio™ Lite software (Li-COR
Biosciences).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
RNA samples from bone tissues were transferred to
the aforementioned tube prefilled with beads (Nextadvance, Inc.)
and homogenized using a Bullet blender (Nextadvance, Inc.). RNA
extraction was performed using the TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturers'
protocol. Isolated RNA was dissolved in RNAse-free water and
quantified by measuring the absorbance at 260 and 280 nm with a
spectrophotometer. The RNA samples were then treated with DNAse I,
and cDNA was prepared for each sample using RevertAid H Minus
Reverse Transcriptase (EP0452, Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The primers of the target
genes were indicated in (Table I).
To evaluate the gene expression levels, qPCR was performed with
SYBR Green PCR Kit using iCycler (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). PCR reactions were performed at 0.5 µM for each
primer in a 25-µl volume containing 1 µl cDNA sample. The reaction
was initiated by activating the polymerase with a 5-min
pre-incubation at 95°C. Amplification was achieved with 45 cycles
of 15 sec denaturation at 94°C, 20 sec annealing at 65°C and 10 sec
extension at 72°C. The program was concluded by a melting curve
analysis. All experiments were performed in triplicate. The copy
numbers of each gene were determined with the quantification cycle
(2−ΔΔCq) method (20).
The means of the copy numbers of glyceraldehyde 3-phosphate
dehydrogenase GAPDH were used as internal controls. Standard curves
of all primers were prepared from total normal cDNA, amplified by
semi-quantitative PCR and cloned using the TOPO II TA Cloning Kit
(Thermo Fisher Scientific, Inc.) following the manufacturer's
recommendations.
 | Table I.Primer sequences utilized for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Primer sequences utilized for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Primer sequences
(5′-3′) |
|---|
| Runx2 | F:
TCCAGACCAGCAGCACTCCATA |
|
| R:
TTCCATCAGCGTCAACACCATC |
| ALP | F:
CTGCGGGACTCAACGACACT |
|
| R:
AGGAGGGTACTCATTGGC ATAGC |
| OCN | F:
TCACAGACCCAGACAGAACCG |
|
| R:
AGCCCAGAGTCCAGGTAGCG |
| HIF-1α | F:
GTGTACCCTAACTAGCCGAGGA |
|
| R:
GTTCACAAATCAGCACCAAGC |
| VEGF | F:
AAGAGCGATCCCCACGTCAA |
|
| R:
TTCGTTTCAGTGCCACATACCA |
| GAPDH | F:
ATGTTTGTGATGGGCGTGAAGGTC |
|
| R:
TTCTGGGTGGCAGTGAT |
Statistical analysis
Data are presented as mean ± standard deviation
(n>3). Individual pairwise comparisons were analyzed by
2-sample, 2-tailed Student's t-tests. The statistical significance
of differences among more than 2 groups was analyzed using one-way
analysis of variance. All statistical analyses were performed using
SPSS 18.0 (SPSS, Inc., Chicago. IL, USA) and P<0.05 was
considered to indicate a statistically significant difference.
Results
General observations
Following surgery, all 6 dogs remained in good
health and had no complications. All extraction sockets healed
uneventfully and there were no signs of inflammation or other
adverse reactions.
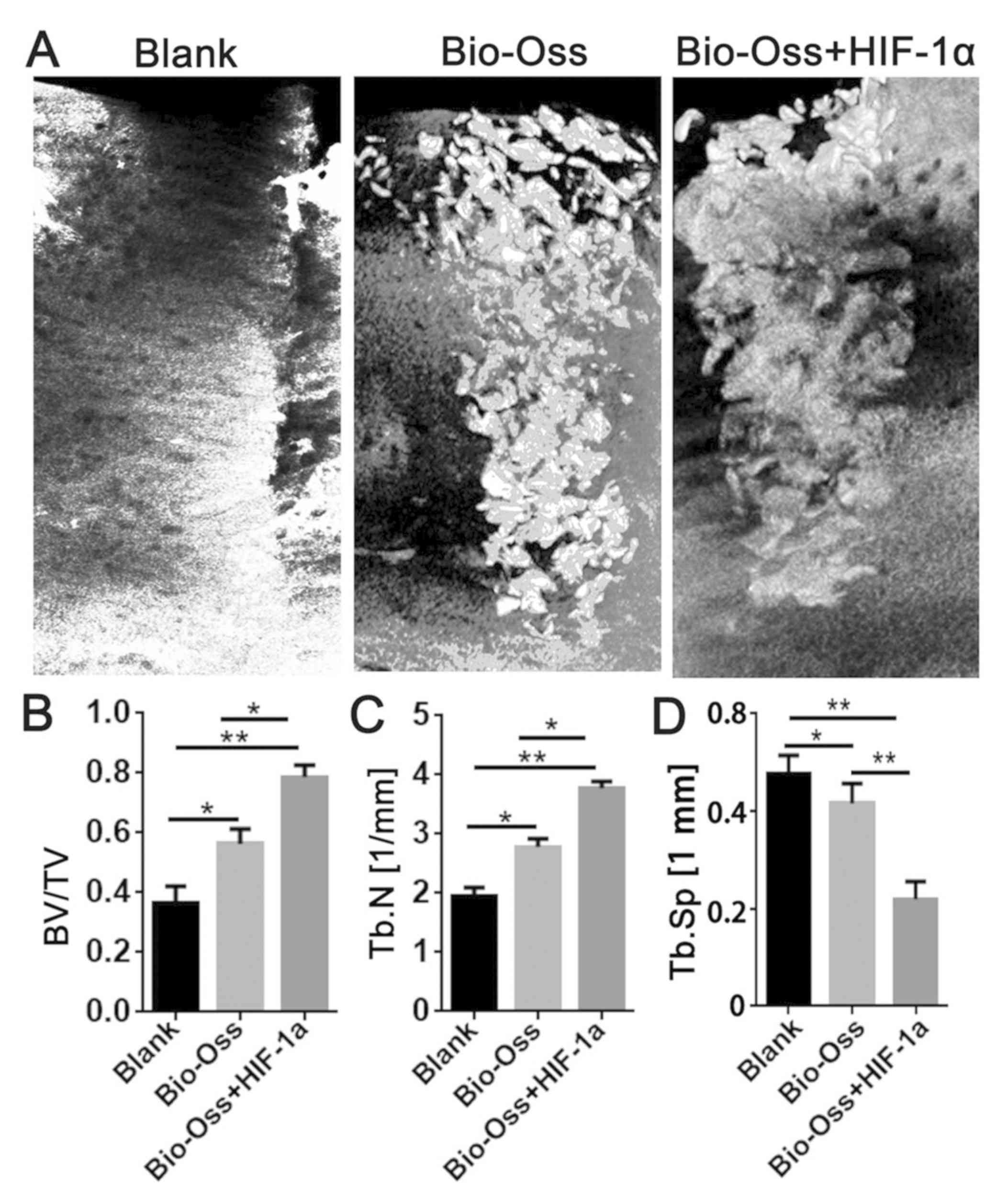
Micro-CT measurement
Micro-CT software was used for the three-dimensional
reconstruction of the mandibular alveolar sockets, and the alveolar
bone regeneration in the hollow of the extraction site was further
evaluated. The results of micro-CT examination revealed that
Bio-Oss combined with HIF-1α resulted in a significantly higher
BV/TV (P<0.05 and P<0.01; Fig.
2B) and Tb.N (P<0.05 and P<0.01; Fig. 2C), and a significantly lower
trabecular separation (Tb.Sp; each, P<0.01; Fig. 2D) compared with the other two
groups.
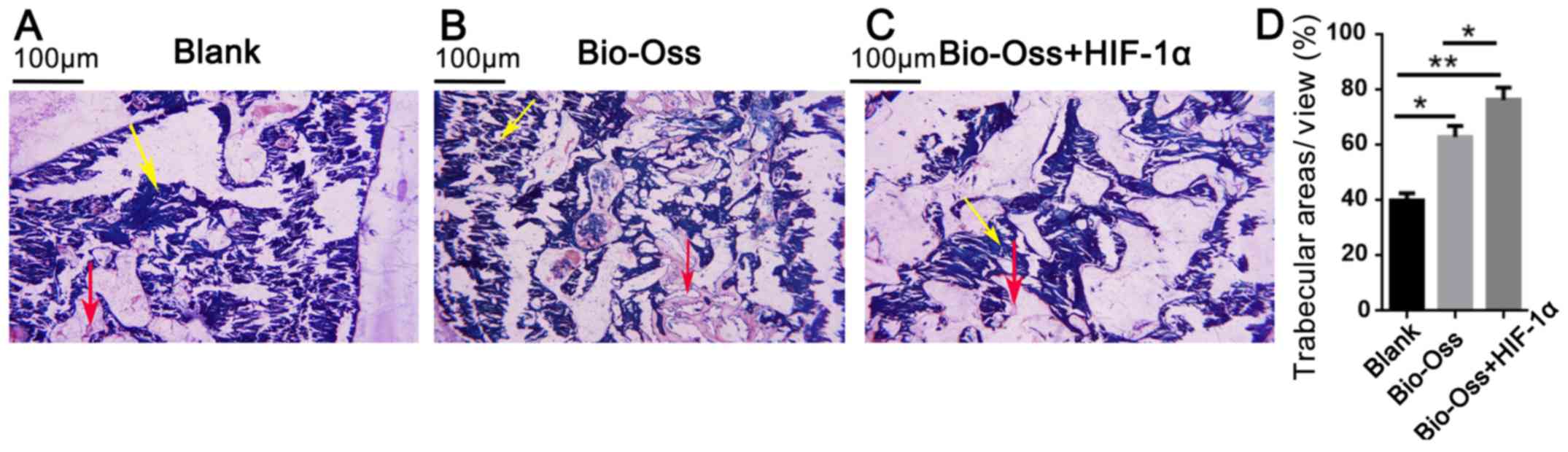
Histological and histomorphometric
analysis
A total of 12 weeks post-surgery, all the extraction
sockets were filled with new bone, and there were clear boundaries
between old and new bone in each group. New bone consisted
primarily of mature woven bone, a part of which was transformed
into lamellar bone (Fig. 3A-C). In
the Bio-Oss and Bio-Oss + HIF-1α groups, there was more active new
bone formation, trabecular bone maturation and calcification
compared with the blank group (Fig.
3). Furthermore, the trabecular bone areas were significantly
more extensive in the Bio-Oss + HIF-1α group as compared with those
in the blank and Bio-Oss groups (P<0.05 and P<0.01; Fig. 3D).
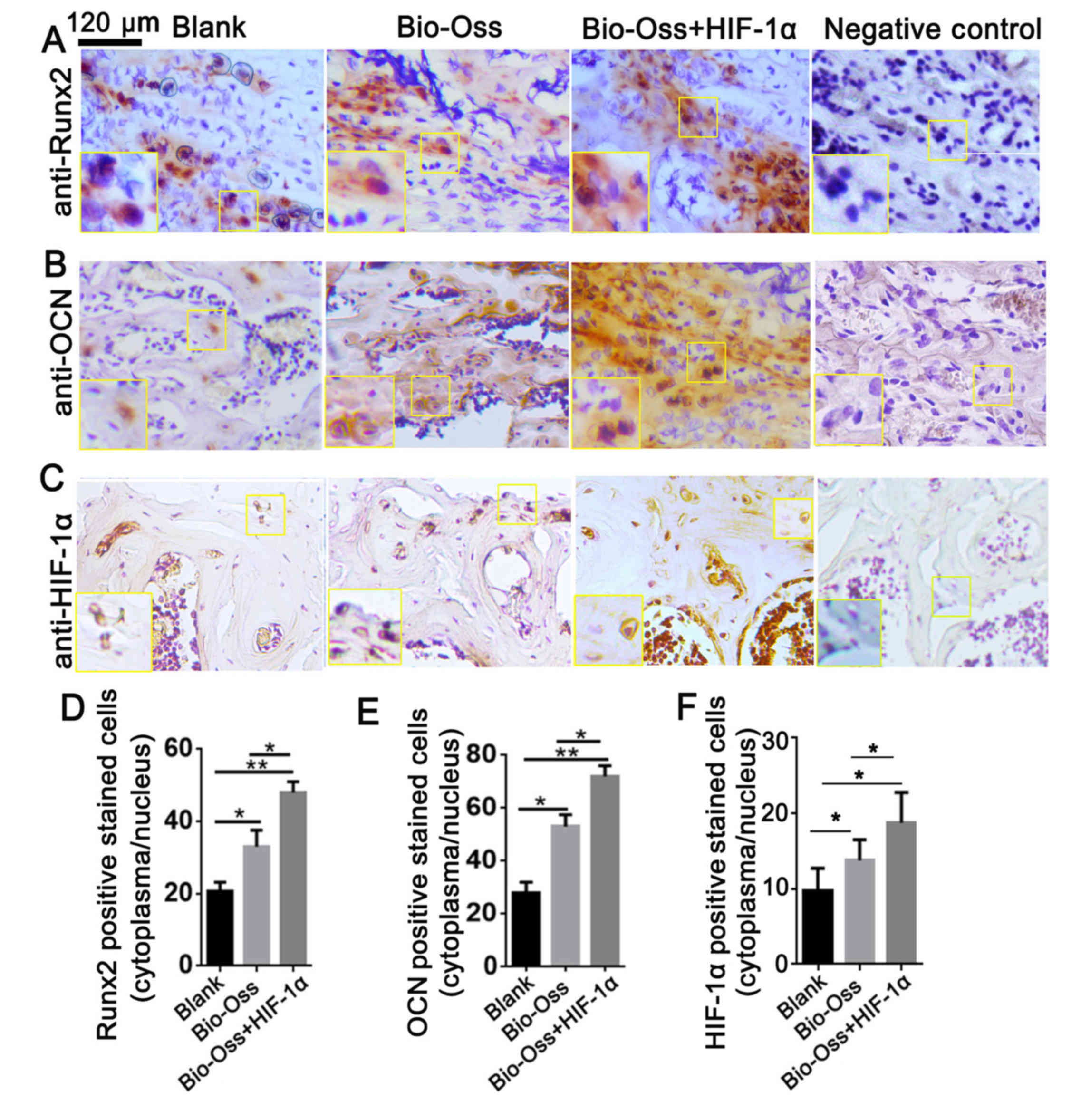
HIF-1α combined with Bio-Oss improves
the expression of Runx2 and OCN in vivo
To further investigate the role of HIF-1α in
osteoblast differentiation in the bone marrow, IHC staining was
performed to analyze the protein expression level of Runx2, OCN and
HIF-1α in bone sections (Fig. 4).
Runx2 is the first transcription factor required for the
determination of osteoblast commitment, and is predominantly
expressed in preosteoblasts and immature osteoblasts (21). The present results demonstrated that
the Runx2 expression level was significantly upregulated in the
Bio-Oss + HIF-1α group compared with the other two groups
(P<0.05 and P<0.01; Fig. 4A and
D). The results also indicated that the number of
HIF-1α-positive stained cells was significantly increased in the
Bio-Oss + HIF-1α group compared with the other groups (both
P<0.01; Fig. 4C and F). Notably,
the expression level of OCN, which is primarily expressed in mature
osteoblasts (22), was also
significantly increased in the Bio-Oss + HIF-1α group compared with
the other two groups (P<0.05 and P<0.01; Fig. 4B and E). These results indicated that
HIF-1α treatment in the extraction sites promoted osteoblast
differentiation and mineralization in bone tissues.
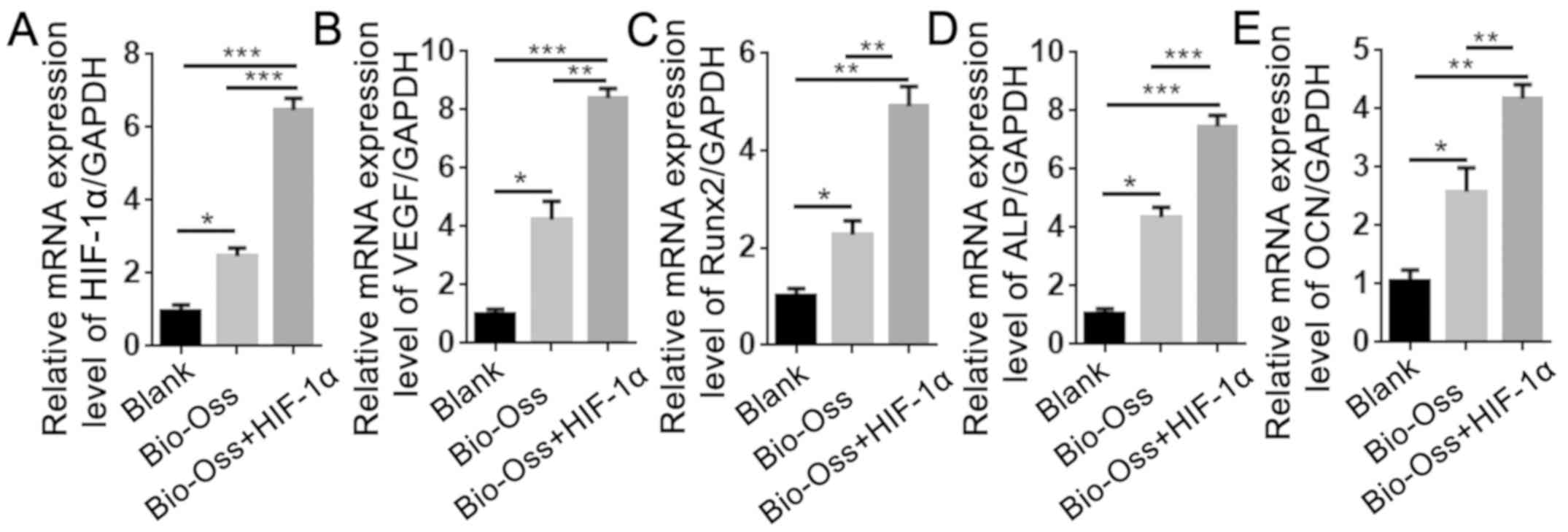
HIF-1α combined with Bio-Oss activates
the mRNA expression levels of angiogenesis- and
osteogenesis-associated genes in extraction sites
To examine the effects of HIF-1α treatment on the
expression of angiogenesis- and osteogenesis-associated genes, RNA
was isolated from the bone tissues around the extraction sites. The
results of RT-qPCR revealed that the mRNA expression levels of
HIF-1α and VEGF in the Bio-Oss + HIF-1α group were 7-fold and
9-fold higher, respectively, compared with the blank group, and
2-fold and 1.5-fold higher, respectively, compared with the Bio-Oss
group (Fig. 5A and B). In addition,
the expression of Runx2, alkaline phosphatase (ALP) and OCN in the
Bio-Oss + HIF-1α group was significantly upregulated compared with
the other two groups (P<0.05; Fig.
5C-E), which was consistent with the results of IHC staining.
These results demonstrated that HIF-1α enhanced the function of
Bio-Oss in promoting angiogenesis and osteogenesis in the
extraction sites.
HIF-1α combined with Bio-Oss
upregulates the protein expression levels of osteogenesis- and
angiogenesis-associated factors in extraction sites
To further examine the effect of HIF-1α on
osteogenesis and angiogenesis in the extraction sites at the
protein level, protein was isolated from the bone tissues around
the extraction sites. The results of western blotting indicated
that the protein level of VEGF, OSX and Runx2 were significantly
increased in the Bio-Oss + HIF-1α group compared with those in the
blank group and Bio-Oss group. Furthermore, the protein level of
OCN in the Bio-Oss + HIF-1α group was ~3-fold and 7-fold higher
than those in the Bio-Oss group and Blank group (Fig. 6A and B). These results confirmed that
application of HIF-1α in the extraction sites increased the protein
expression levels of angiogenesis- and osteogenesis-associated
factors.
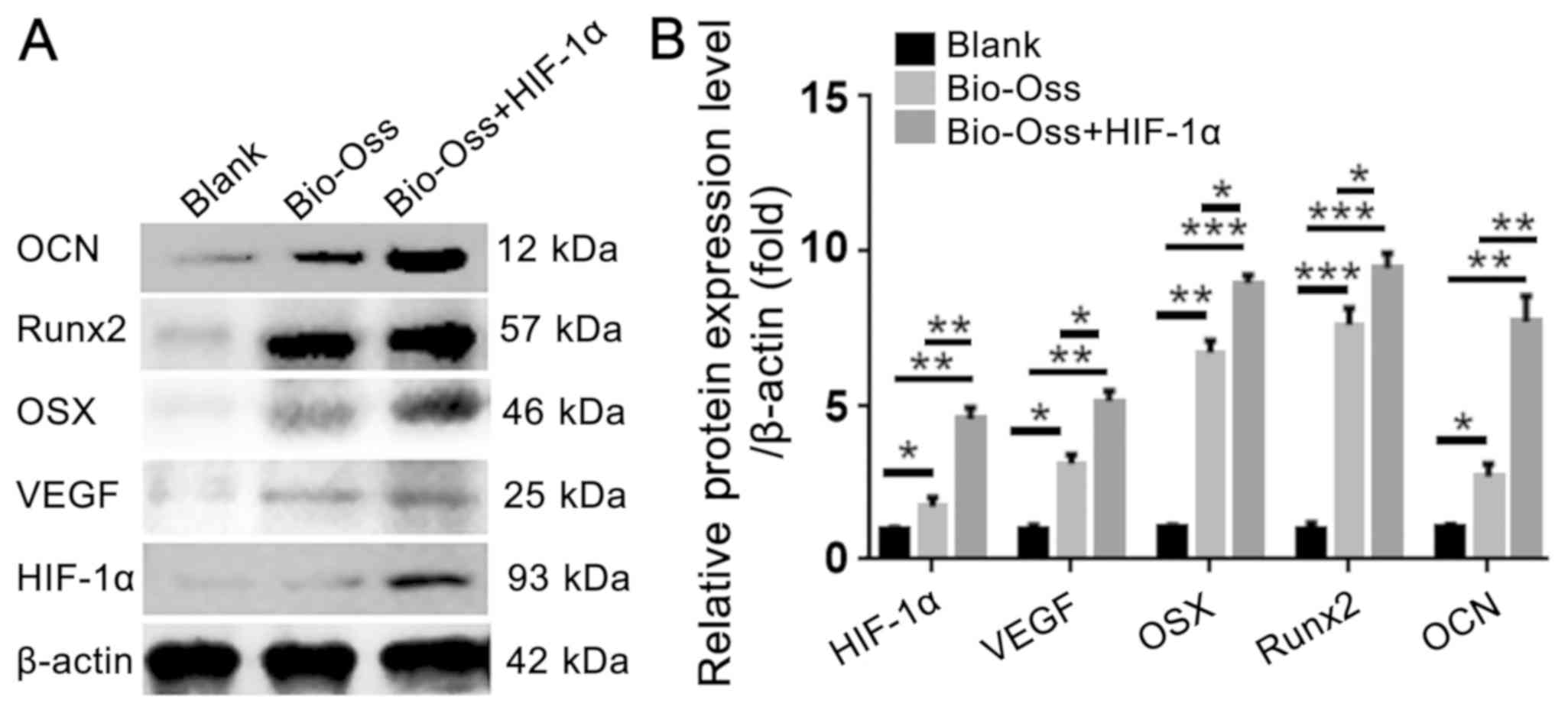 | Figure 6.HIF-1α combined with Bio-Oss
upregulates the protein expression levels of osteogenesis- and
angiogenesis-associated factors in extraction sites. (A) Western
blotting was used to detect the protein levels of OCN, Runx2, OSX,
VEGF and HIF-1α in the bone tissues around the extraction sites.
(B) Quantification of the protein levels in (A) by normalizing to
β-actin. The results are presented as mean ± standard deviation,
n>3. *P<0.05, **P<0.01 and ***P<0.005 as indicated.
HIF-1α, hypoxia-inducible factor-1α; VEGF, vascular endothelial
growth factor; Runx2, runt-related transcription factor 2; OCN,
osteocalcin; OSX, osteoblast-specific transcription factor
osterix. |
Discussion
A variety of studies have demonstrated that HIF-1α
is subjected to proteasomal degradation under non-hypoxic
conditions (23,24). The present study investigated the
effect of HIF-1α on alveolar ridge preservation. The present
results demonstrated that the HIF-1α protein provided a superior
micro-environment for alveolar ridge preservation compared with the
Bio-Oss alone and blank groups. Therefore, Bio-Oss + HIF-1α appears
to be an effective treatment for enhancing new bone formation in
tooth extraction sites, as it promotes osteogenesis and
angiogenesis.
A good substitute for alveolar ridge preservation
should not only repair the bone defect, but must also be able to
preserve the alveolar shape (15).
The most frequently used method is the alveolar fossa plug, which,
however, only fills the space rather than promoting the formation
of new bone. In the present study, Bio-Oss in combination with
HIF-1α was used to fill the extraction socket, which is a hypoxic
environment following tooth extraction, similar to humans (25). The activation of HIF-1α is a specific
and sensitive response to hypoxia, which may induce the expression
of several pro-angiogenic factors, such as VEGF, inducible nitric
oxidase, heme oxygenase-1, angiotensin-2 and EPO (26). Furthermore, HIF-1α can increase the
number of osteoblasts, improve their ability to survive under
hypoxic and ischemic conditions, and increase angiogenesis in
hypoxic tissues (27). In addition,
as an ideal support, Bio-Oss is similar to the human bone structure
and has a large internal surface area, which makes it suitable for
the adsorption of growth factors and biological proteins (28). The different pore sizes of the
internal structure of Bio-Oss can promote the regeneration of blood
vessels and maintain the stability of blood clots (29). Bio-Oss consists of small granules,
with a diameter of 10–60 nm. In addition, its physical structure is
similar to that of human bone, and can be absorbed during bone
regeneration (30). As angiogenesis
and osteogenesis are tightly coupled during bone regeneration, the
combination of HIF-1α and Bio-Oss appears to be an optimal choice
for alveolar ridge preservation following tooth extraction.
However, the application of HIF-1α in bone
regeneration remains controversial. According to a previous report,
HIF-1α is a pro-osteogenic factor in woven bone formation following
damaging loading, but is considered an anti-osteogenic factor in
lamellar bone formation following non-damaging mechanical loading
(31). In the present study, the
HIF-1α protein combined with Bio-Oss significantly increased
trabecular bone formation in the extraction sockets compared with
the other two groups, as indicated by micro-CT examination and
H&E staining. Furthermore, the arrangement of the vascular
network was more concentrated in the HIF-1α + Bio-Oss group
compared with the other two groups. These results demonstrated that
HIF-1a combined with Bio-Oss not only promoted osteogenesis in the
extraction socket, but also stimulated the formation of new blood
vessels around the bone. The results can be explained by the fact
that tooth extraction may represent damaging mechanical loading
that activates the supportive function of HIF-1α in trabecular bone
formation. In addition, Tomlinson and Silva (32) also revealed that the
osteogenesis-promoting function of HIF-1α in sites with damaging
mechanical loading may be associated with its role as a
transcription factor during angiogenesis. VEGF is the primary
target of HIF-1α; it serves an important role in the
vascularization of bone and it is upregulated after damaging bone
loading (33). Additionally,
inhibition of angiogenesis results in reduced woven bone formation
following damaging mechanical loading (34). Consistently, the present results
demonstrated that the expression levels of osteogenesis-associated
genes (Runx2, OSX and OCN) and an angiogenesis-associated gene
(VEGF) were significantly upregulated at the mRNA and protein
levels in the Bio-Oss + HIF-1α group compared with the other two
groups. The results indicate that HIF-1α may be used as an enhancer
for trabecular bone formation in tooth extraction sites.
As previous reports have indicated, HIF-1α promotes
the expression of osteogenic and angiogenic related genes in
hypoxia, which indicates HIF-1α is one of the major regulators of
angiogenic-osteogenic coupling in bone formation (35,36).
Based on the results of the present study, the increased bone
formation in the HIF-1α + Bio-Oss group may have occurred via the
several mechanisms. First, HIF-1α may transactivates the expression
of VEGF, which stimulates blood vessel formation in the extraction
sites. The increased blood supply not only delivers the oxygen and
nutrients to the bone, but also introduces more numbers of
mesenchymal stem cells in the extraction sites that then commit to
osteoblasts. The other is that, HIF-1α may act directly on the
osteoblast-associated genes to promote osteoblast differentiation
and mineralization. It has been demonstrated that in some
experimental settings, HIF-1α and VEGF can regulate bone cell
function without stimulating angiogenesis (37). The results from the present IHC
staining and qPCR analysis also revealed increased expression
levels of Runx2 and OCN in the HIF-1α+Bio-Oss group.
Taken together, the findings of the present study
demonstrated that HIF-1α combined with Bio-Oss promotes trabecular
bone formation in the extraction sockets by activating the
expression of osteogenesis- and angiogenesis-associated genes. This
study provides important insights into the role of HIF-1α under
conditions of damaging mechanical loading during bone development.
Further elucidation of the mechanisms underlying the regulatory
effect of HIF-1α on the expression of Runx2, OSX and OCN may
provide a useful tool for its application in bone regeneration.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (NSFC) (grant no. 81470768).
and by grants from the Leading Academic Discipline Project of
Pudong New Area Health System Discipline (grant no. PWRd2016-09)
and Jiangxi Natural Science Foundation (grant no.
20181BAB2050380).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL, YHL and LT conceived and designed the
experiments of the current study. LT, YZ and YL performed the
experiments. YH and YHL analyzed the data. YL and YHL wrote and
revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The Institutional Animal Care and Use Committee of
the Ninth People's Hospital, Shanghai Jiao Tong University,
approved the protocol used in this study. All animals were handled
according to the guidelines established by the Institutional Animal
Care Committee.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests to disclose.
References
|
1
|
Mardas N, Chadha V and Donos N: Alveolar
ridge preservation with guided bone regeneration and a synthetic
bone substitute or a bovine-derived xenograft: A randomized,
controlled clinical trial. Clin Oral Implants Res. 21:688–698.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Araújo MG, Liljenberg B and Lindhe J:
Dynamics of Bio-Oss Collagen incorporation in fresh extraction
wounds: An experimental study in the dog. Clin Oral Implants Res.
21:55–64. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Becker W, Clokie C, Sennerby L, Urist MR
and Becker BE: Histologic findings after implantation and
evaluation of different grafting materials and titanium micro
screws into extraction sockets: Case reports. J Periodontol.
69:414–421. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jung RE, Glauser R, Schärer P, Hämmerle
CH, Sailer HF and Weber FE: Effect of rhBMP-2 on guided bone
regeneration in humans. Clin Oral Implants Res. 14:556–568. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simion M, Rocchietta I, Monforte M and
Maschera E: Three-dimensional alveolar bone reconstruction with a
combination of recombinant human platelet-derived growth factor BB
and guided bone regeneration: A case report. Int J Periodontics
Restorative Dent. 28:239–243. 2008.PubMed/NCBI
|
|
6
|
Anitua E: The use of plasma-rich growth
factors (PRGF) in oral surgery. Pract Proced Aesthet Dent.
13:487–493. 2001.PubMed/NCBI
|
|
7
|
Cao J, Wang L, Lei DL, Liu YP, Du ZJ and
Cui FZ: Local injection of nerve growth factor via a hydrogel
enhances bone formation during mandibular distraction osteogenesis.
Oral Surg Oral Med Oral Pathol Oral Radiol. 113:48–53. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu K and Olsen BR: The roles of vascular
endothelial growth factor in bone repair and regeneration. Bone.
91:30–38. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci
USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Semenza GL, Roth PH, Fang HM and Wang GL:
Transcriptional regulation of genes encoding glycolytic enzymes by
hypoxia-inducible factor 1. J Biol Chem. 269:23757–23763.
1994.PubMed/NCBI
|
|
11
|
Salceda S and Caro J: Hypoxia-inducible
factor 1alpha (HIF-1alpha) protein is rapidly degraded by the
ubiquitin-proteasome system under normoxic conditions. Its
stabilization by hypoxia depends on redox-induced changes. J Biol
Chem. 272:22642–22647. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Warren SM, Steinbrech DS, Mehrara BJ,
Saadeh PB, Greenwald JA, Spector JA, Bouletreau PJ and Longaker MT:
Hypoxia regulates osteoblast gene expression. J Surg Res.
99:147–155. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ahluwalia A and Tarnawski AS: Critical
role of hypoxia sensor-HIF-1α in VEGF gene activation. Implications
for angiogenesis and tissue injury healing. Curr Med Chem.
19:90–97. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Utting JC, Robins SP, Brandao-Burch A,
Orriss IR, Behar J and Arnett TR: Hypoxia inhibits the growth,
differentiation and bone-forming capacity of rat osteoblasts. Exp
Cell Res. 312:1693–1702. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aludden HC, Mordenfeld A, Hallman M,
Dahlin C and Jensen T: Lateral ridge augmentation with Bio-Oss
alone or Bio-Oss mixed with particulate autogenous bone graft: A
systematic review. Int J Oral Maxillofac Surg. 46:1030–1038. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Paolantonio M, Scarano A, Di Placido G,
Tumini V, D'Archivio D and Piattelli A: Periodontal healing in
humans using anorganic bovine bone and bovine peritoneum-derived
collagen membrane: A clinical and histologic case report. Int J
Periodontics Restorative Dent. 21:505–515. 2001.PubMed/NCBI
|
|
17
|
Shabanovich A: P.067 application of
BIO-OSS® and BIO-GIDE® at carrying out immediate dental
implantology. J Cranio-Maxillofac Surg. 34 (Suppl 1):S148–S149.
2006. View Article : Google Scholar
|
|
18
|
He F, Ren W, Tian X, Liu W, Wu S and Chen
X: Comparative study on in vivo response of porous calcium
carbonate composite ceramic and biphasic calcium phosphate ceramic.
Mater Sci Eng C Mater Biol Appl. 64:117–123. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Roldán JC, Jepsen S, Miller J, Freitag S,
Rueger DC, Açil Y and Terheyden H: Bone formation in the presence
of platelet-rich plasma vs. bone morphogenetic protein-7. Bone.
34:80–90. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Komori T: Roles of Runx2 in skeletal
development. Adv Exp Med Biol. 962:83–93. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Komori T: Regulation of skeletal
development by the Runx family of transcription factors. J Cell
Biochem. 95:445–453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zou D, Zhang Z, He J, Zhu S, Wang S, Zhang
W, Zhou J, Xu Y, Huang Y, Wang Y, et al: Repairing critical-sized
calvarial defects with BMSCs modified by a constitutively active
form of hypoxia-inducible factor-1α and a phosphate cement
scaffold. Biomaterials. 32:9707–9718. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Majmundar AJ, Wong WJ and Simon MC:
Hypoxia-inducible factors and the response to hypoxic stress. Mol
Cell. 40:294–309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Daly B, Sharif MO, Newton T, Jones K and
Worthington HV: Local interventions for the management of alveolar
osteitis (dry socket). Cochrane Database Syst Rev.
12:CD0069682012.PubMed/NCBI
|
|
26
|
Eskandani M, Vandghanooni S, Barar J,
Nazemiyeh H and Omidi Y: Cell physiology regulation by hypoxia
inducible factor-1: Targeting oxygen-related nanomachineries of
hypoxic cells. Int J Biol Macromol. 99:46–62. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ceradini DJ, Yao D, Grogan RH, Callaghan
MJ, Edelstein D, Brownlee M and Gurtner GC: Decreasing
intracellular superoxide corrects defective ischemia-induced new
vessel formation in diabetic mice. J Biol Chem. 283:10930–10938.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yeo EJ, Chun YS and Park JW: New
anticancer strategies targeting HIF-1. Biochem Pharmacol.
68:1061–1069. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Luo G, Huang Y and Gu F: rhBMP2-loaded
calcium phosphate cements combined with allogenic bone marrow
mesenchymal stem cells for bone formation. Biomed Pharmacother.
92:536–543. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kellner J, Sivajothi S and McNiece I:
Differential properties of human stromal cells from bone marrow,
adipose, liver and cardiac tissues. Cytotherapy. 17:1514–1523.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Agarwal S, Loder S, Brownley C, Cholok D,
Mangiavini L, Li J, Breuler C, Sung HH, Li S, Ranganathan K, et al:
Inhibition of Hif1α prevents both trauma-induced and genetic
heterotopic ossification. Proc Natl Acad Sci USA. 113:E338–E347.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tomlinson RE and Silva MJ: HIF-1α
regulates bone formation after osteogenic mechanical loading. Bone.
73:98–104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hankenson KD, Gagne K and Shaughnessy M:
Extracellular signaling molecules to promote fracture healing and
bone regeneration. Adv Drug Deliv Rev. 94:3–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Segar CE, Ogle ME and Botchwey EA:
Regulation of angiogenesis and bone regeneration with natural and
synthetic small molecules. Curr Pharm Des. 19:3403–3419. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Y, Wan C, Deng L, Liu X, Cao X,
Gilbert SR, Bouxsein ML, Faugere MC, Guldberg RE, Gerstenfeld LC,
et al: The hypoxia-inducible factor alpha pathway couples
angiogenesis to osteogenesis during skeletal development. J Clin
Invest. 117:1616–1626. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suzuki N, Gradin K, Poellinger L and
Yamamoto M: Regulation of hypoxia-inducible gene expression after
HIF activation. Exp Cell Res. 356:182–186. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Riddle RC, Khatri R, Schipani E and
Clemens TL: Role of hypoxia-inducible factor-1alpha in
angiogenic-osteogenic coupling. J Mol Med (Berl). 87:583–590. 2009.
View Article : Google Scholar : PubMed/NCBI
|