Introduction
Inflammation is a complex biological response of
body tissues to harmful stimuli, such as pathogens, damaged cells
or irritants; it is a protective response, involving immune cells,
blood vessels, and molecular mediators (1). Inflammation may be cause of various
diseases (1). The factors involved
in the anti-inflammatory effect may play a therapeutic role for
diseases implicated in inflammation. Anti-inflammatory effects have
reportedly been exhibited by botanical factors, including catechin
(2,3), baicalin (4,5), and
β-caryophyllene (6,7). β-Caryophyllene, a sesquiterpene, is a
natural dietary ingredient found in many edible plants, which can
be ingested daily as an essential oil and is approved as a food
additive by the Food and Drug Administration (FDA). Interestingly,
β-caryophyllene is known to be a selective agonist of cannabinoid
receptor type-2 and to exert cannabimimetic anti-inflammatory and
analgesic effects in animals (8,9) for both
acute and chronic pain with inflammation (10–14).
However, the mechanism by which β-caryophyllene exhibits
anti-inflammatory effects is poorly understood. Previously, we
investigated whether β-caryophyllene interacted with various
botanical molecules, which revealed anti-inflammatory effects in
mouse macrophages RAW264.7 cells in vitro (15). It was found that the combination of
(+)-catechin, baicalin and β-caryophyllene with concentrations,
which did not independently exhibit a significant effect on
RAW264.7 cells, showed a synergistic-suppressive effect on the
proliferation and a synergistic-stimulatory effect on the death of
RAW267.4 cells in vitro (15), supporting the view that these three
factors in combination could potentially eliminate activated
macrophages. This may serve as a usefulness therapeutic tool
against inflammation.
RAW264.7 cells are a murine macrophage that produces
various inflammatory cytokines including TNF-α, IL-6 and IL-1β
(16). The present study was
undertaken to investigate the suppressive effects of (+)-catechin,
baicalin and β-caryophyllene, which does not have significant
effects on cell number, on the production of inflammatory
cytokines, including TNF-α, IL-6 and IL-1β, which is enhanced by
the treatment with lipopolysaccharide (LPS) in mouse macrophage
RAW264.7 cells in vitro. The productions of TNF-α, IL-6 and
IL-1β was not altered by the addition of (+)-catechin, baicalin,
β-caryophyllene or by the three combined factors in RAW264.7 cells
cultured without LPS. The treatment with LPS caused a remarkable
production of TNF-α, IL-6 and IL-1β. This enhancement was
suppressed by the addition of (+)-catechin, baicalin or
β-caryophyllene. Interestingly, the production of these cytokines
was suppressed in RAW264.7 cells by the combination of all three
factors. Thus, the combination of (+)-catechin, baicalin and
β-caryophyllene was found to have a potent-suppressive effect on
macrophages in vitro. This composition may be a useful tool
as anti-inflammatory agent.
Materials and methods
Materials
Dulbecco's modification of Eagle's medium (DMEM)
with 4.5 g/l glucose, L-glutamine and sodium pyruvate and
antibiotics (penicillin and streptomycin) were purchased from
Corning (Mediatech, Inc., Manassas, VA, USA). Fetal bovine serum
(FBS) was from Hyclone (Logan, UT, USA). (+)-catechin, baicalin,
and β-caryophyllene were obtained from Cayman Chemical (Ann Arbor,
MI, USA). These reagents were dissolved in dimethylsulfoxide (DMSO)
and stored in the dark at −20°C until use experiment. All other
reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA)
unless otherwise specified. We used the following doses:
(+)-Catechin [1µg/ml (3.45 µM) of medium], baicalin [1 µg/ml (2.24
µM)], and β-caryophyllene [1 µg/ml (5 µM)]. At these doses, the
combinations exhibited a synergistic effect on the number of
RAW264.7 cells with culture for 3 days (15).
RAW264.7 cells
Mouse RAW264.7 cells (murine macrophage) were
obtained from the American Type Culture Collection (Rockville, MD,
USA) (17,18). RAW264.7 cells were cultured in DMEM
containing 10% FBS and 1% P/S.
Assay of cell proliferation
To determine the effect of various factors on cell
growth, RAW264.7 cells (1×105/ml per well) were cultured
in a water-saturated atmosphere containing 5% CO2 and
95% air at 37°C using a 24-well plate (8,19). Cells
were cultured in DMEM containing 10% FBS and 1% P/S in the presence
of either vehicle (1% DMSO as a final concentration), (+)-catechin
(1 µg/ml), baicalin (1 µg/ml), or β-caryophyllene (1 µg/ml) for 3
days upon reaching confluency. To determine the increase in cell
number, RAW264.7 cells, which were attached on each dish, were then
detached from each culture dish by adding a sterile solution (0.1
ml per well) of 0.05% trypsin plus EDTA in
Ca2+/Mg2+-free PBS (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) with incubation for 2 min at 37°C. Each
well was then added 0.9 ml of DMEM containing 10% FBS and 1% P/S.
The cell number in the cell suspension was counted as described
below in the section ‘cell counting’.
Assay of cell death
To determine the effect of various factors on cell
death including necrotic cell death and apoptotic cell death,
RAW264.7 cells (1×105/ml per well) were cultured using a
24-well plate in DMEM containing 10% FBS and 1% P/S for 3 days in
reaching upon confluency in the absence of drugs (8,20). After
reaching on subconfluence with culture for 3 days, the cells were
additionally cultured for 24 h after treatment of either vehicle
(1% DMSO as a final concentration), (+)-catechin (1 µg/ml of
medium), baicalin (1 µg/ml), β-caryophyllene (1 µg/ml), or their
combination. After culture, the viable cells, which were attached
on each dish, were detached from each culture dishes by adding a
sterile solution (0.1 ml per well) of 0.05% trypsin plus EDTA in
Ca2+/Mg2+-free PBS (Thermo Fisher Scientific,
Inc.) with incubation for 2 min at 37°C. Each well was then added
0.9 ml of DMEM containing 10% FBS and 1% P/S. The cell number in
the cell suspension was counted as described below in the section
‘cell counting’.
Counting of cell number
To detach cells on each well after culturing in
order to assay the proliferation and death of RAW264.7 cells, the
culture dishes were incubated for 2 min at 37°C after the addition
of a solution (0.1 ml per well) containing 0.05% trypsin plus EDTA
in Ca2+/Mg2+-free PBS, and the cells were
detached through pipetting after the addition of DMEM (0.9 ml)
containing 10% FBS and 1% P/S (8,19,20).
Medium containing the suspended cells (0.1 ml) was mixed by the
addition of 0.1 ml of 0.5% trypan blue staining solution, which can
look living cells but not death cells. The number of viable cells
was counted under a microscope (Olympus MTV-3) using a
Hemocytometer (Sigma-Aldrich) and a cell counter (Line Seiki
H-102P, Tokyo, Japan). For each dish, we took the average of two
counts. Cell numbers are shown as number per well.
Assay of cytokine production
RAW264.7 cells (1×105/ml per well) were
cultured in a 24-well plate in DMEM containing 10% FBS and 1% P/S
for 3 days upon reaching subconfluency (8,19), and
then the cells were further cultured for 5 h after the addition of
either vehicle (1% DMSO as a final concentration), (+)-catechin (1
µg/ml of medium), baicalin (1 µg/ml), (β-caryophyllene (1 µg/ml),
or the three factors in combination (1 µg/ml of each factor) with
or without LPS (100 ng/ml). After incubation, medium was collected
to assay cytokines, and then the cells were detached from each of
the culture dishes to determine the number of cells as described in
‘cell counting’ (21). The
concentrations of TNF-α, IL-6 or IL-1β in the medium were analyzed
with the ELISA kits (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) of mouse TNF-α (cat. no. KHC301), IL-6 (cat. no. EH2IL6), and
IL-1β (cat. no. EM2IL1B) according to the manufactur's
instructions. Data regarding the production of each cytokine are
presented as pictogram (pg) secreted into culture medium (ml).
Statistical analysis
Data are presented as the mean ± standard deviation
(SD). Statistical significance was determined using GraphPad InStat
version3 for Windows XP (GraphPad Software, Inc., La Jolla, CA,
USA). Multiple comparisons were performed by one-way analysis of
variance (ANOVA) with the Tukey-Kramer multiple comparisons post
hoc test for parametric data as indicated, and we reanalyzed data
without using t-tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
The effects of (+)-catechin, baicalin and
β-caryophyllene, which have anti-inflammatory effects, on the
proliferation of RAW264.7 cells in vitro were investigated.
To determine the effect on cell proliferation, RAW264.7 cells were
cultured for 3 days in the presence of either vehicle (1% DMSO as a
final concentration), (+)-catechin [1 µg/ml (3.45 µM)], baicalin [1
µg/ml (2.24 µM)], or β-caryophyllene [1 µg/ml (5 µM)] (Fig. 1). Cell growth was not altered in the
presence of (+)-catechin, baicalin, β-caryophyllene, or baicalin (1
µg/ml) plus β-caryophyline (1 µg/ml), and catechin (1 µg/ml) plus
β-caryophyline (1 µg/ml). Of note, the three combination of
(+)-catechin, baicalin and β-caryophyllene revealed a
potential-suppressive effect on cell proliferation (Fig. 1). Suppression of cell proliferation
caused during 3 days-culture with combined compounds showed
approximately 45% as compared with that of control.
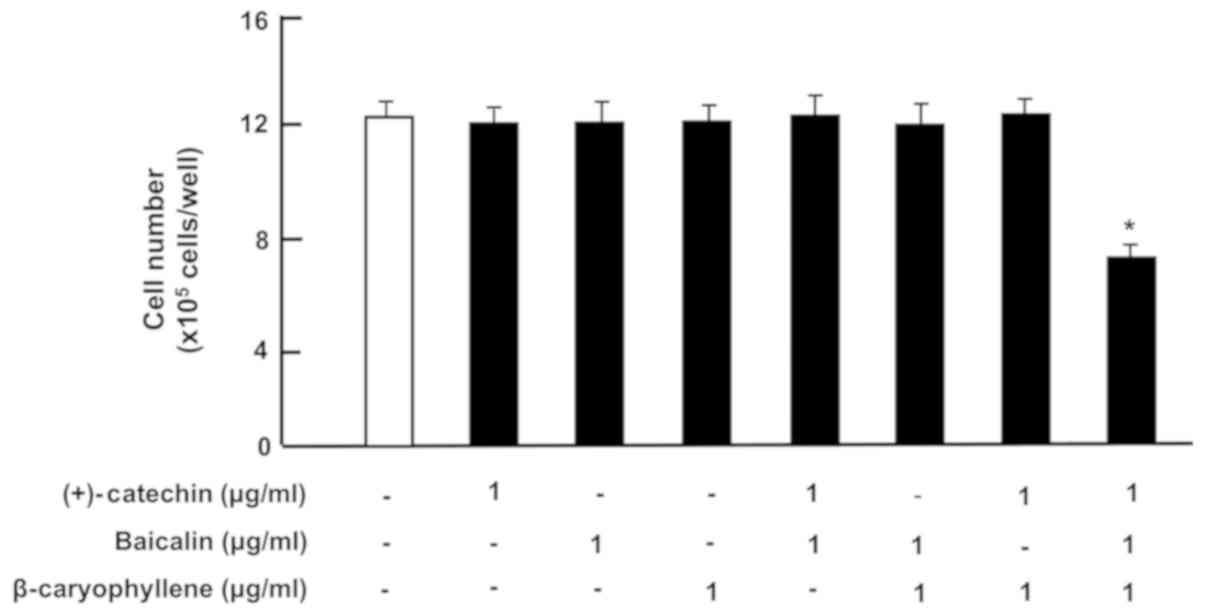 | Figure 1.Effects of (+)-catechin, baicaline,
and β-caryophylline on the proliferation of RAW264.7 cells in
vitro. RAW264.7 cells (1×105/ml per well) were
cultured for 3 days in DMEM containing either vehicle (1% DMSO),
(+)-catechin (1 µg/ml of medium), baicalin (1 µg/ml),
β-caryophyllene (1 µg/ml), (+)-catechin (1 µg/ml of medium) plus
baicaline (1 µg/ml), or all three combined factors (each 1 µg/ml).
After culture, the number of attached cells on dish was counted.
Data are presented as mean ± SD obtained from 8 wells of 2
replicate wells per data set using different dishes and cell
preparations. *P<0.001 vs. control (white bar). One-way ANOVA,
Tukey-Kramer post hoc test. DMEM, Dulbecco's modification of
Eagle's medium; DMSO, dissolved in dimethylsulfoxide; SD, standard
deviation. |
Next, to determine the effects of (+)-catechin,
baicalin or β-caryophyllene on the death of RAW264.7 cells, the
cells were cultured for 3 days in the absence of botanical factors,
and then the cells were cultured for an additional 24 h in the
presence of either vehicle (1% DMSO as a final concentration),
(+)-catechin (1 µg/ml), baicalin (1 µg/ml), or β-caryophyllene (1
µg/ml). The numbers of cells were not changed by the addition of
each factor (Fig. 2). Of note, the
combination of (+)-catechin, baicalin and β-caryophyllene exhibited
a potential-stimulatory effect on cell death (including necrotic
cell death and apoptotic cell death) (Fig. 2). Further experiment may be used to
determine a specific apoptotic cell death. Result showed the
reduction of cell number approximately 60% with the three combined
drugs as compared with that of control. Thus, the combination of
(+)-catechin, baicalin and β-caryophyllene was demonstrated to
reveal potential effects on the depression of proliferation and the
stimulation of death in mouse macrophage RAW264.7 cells, leading to
the reduction of macrophage related to inflammation.
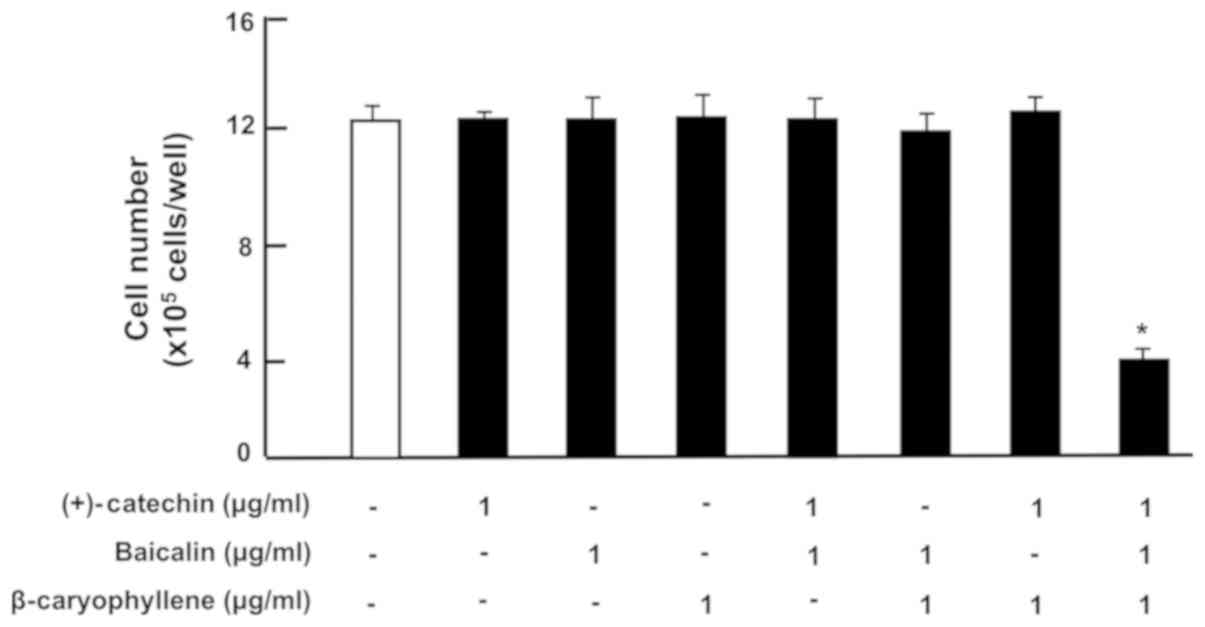 | Figure 2.Effects of (+)-catechin, baicaline,
and β-caryophylline on the death of RAW264.7 cells in vitro.
RAW264.7 cells (1×105/ml per well) were cultured for 3
days in reaching upon subconfluence without botanical factors, and
then were further cultured for 24 h in DMEM containing either
vehicle, (+)-catechin (1 µg/ml), baicalin (1 µg/ml),
β-caryophyllene (1 µg/ml), (+)-catechin (1 µg/ml) plus baicalin (1
µg/ml), baicalin (1 µg/ml) plus β-caryophyline (1 µg/ml), and
catechin (1 µg/ml) plus β-caryophyline (1 µg/ml), or all three
combined factors (each 1 µg/ml). After culture, the number of
attached cells on dish was counted. Data are presented as mean ± SD
obtained from 8 wells of 2 replicate wells per data set using
different dishes and cell preparations. *P<0.001 vs. control
(white bar). One-way ANOVA, Tukey-Kramer post hoc test. DMEM,
Dulbecco's modification of Eagle's medium; SD, standard
deviation. |
Moreover, we investigated the effects of
(+)-catechin, baicalin or β-caryophyllene on the production of
inflammatory cytokines, including TNF-α, IL-6 and IL-1β, in
RAW264.7 cells. In the previous studies (15), we investigated the effect of
increasing dosages of the used three botanical factors in the range
of 0.1 to 100 µg/ml of medium in RAW264.7 cells in vitro.
Potential-suppressive effect on the number of cells was found in
over 1 µg/ml of each factor, which were based on our previous
finding. Cytokine production by cells was resulted from the number
of living cell on the dish. Therefore, it was important to estimate
cytokine production with the use of chemical dosage, which did not
have any effects on cell number. In this experiment, the cells were
cultured for 3 days without botanical factors, and the cells, upon
reaching subconfluency, were cultured for an additional 5 h in the
presence of either vehicle, (+)-catechin (1 µg/ml), baicalin (1
µg/ml), β-caryophyllene (1 µg/ml), or three combined factors with
or without LPS (100 ng/ml). The number of RAW264.7 cells was not
changed by the culture with (+)-catechin, baicalin,
β-caryophyllene, or a combination of the three factors in the
presence (Fig. 3A) or absence
(Fig. 3B) of LPS.
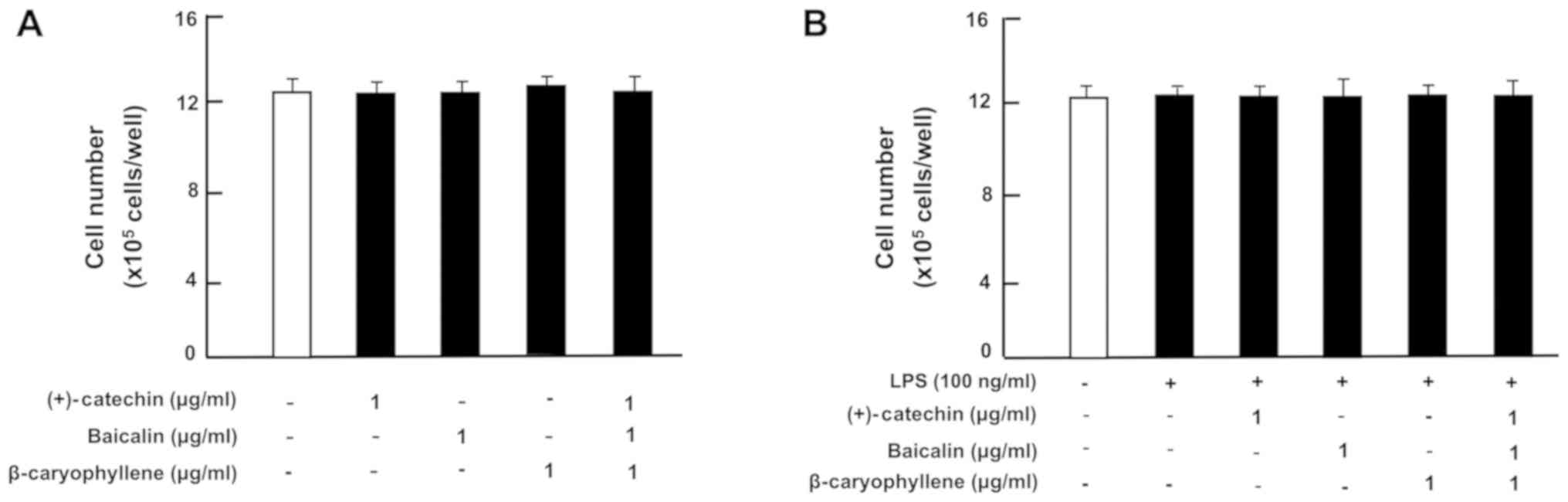 | Figure 3.Effects of (+)-catechin, baicaline,
and β-caryophylline on the number of RAW264.7 cells with or without
LPS stimulation. RAW264.7 cells (1×105/ml per well) were
cultured in the absence of botanical factors for 3 days in reaching
upon subconfluence, and then the cells were further incubated for 5
h after the addition of either vehicle (1% DMSO), (+)-catechin (1
µg/ml), baicalin (1 µg/ml), β-caryophyllene (1 µg/ml), or all three
combined factors (each 1 µg/ml) without (A) or with (B) LPS (100
ng/ml). After incubation, the number of attached cells on dish was
counted. Data are presented as mean ± SD obtained from 8 wells of 2
replicate wells per data set using different dishes and cell
preparations. One-way ANOVA, Tukey-Kramer post hoc test. LPS,
lipopolysaccharide; DMSO, dissolved in dimethylsulfoxide; SD,
standard deviation. |
Then, under culture conditions that did not alter
the cell number, we determined the effects of (+)-catechin,
baicalin, β-caryophyllene, or three combined factors on the
production of TNF-α, IL-6, and IL-1β in RAW264.7 cells incubated
with or without LPS (100 ng/ml). In the absence of LPS, the
productions of TNF-α (Fig. 4A), IL-6
(Fig. 5A) and IL-1β (Fig. 6A) was not significantly altered by
the addition of (+)-catechin (1 µg/ml), baicalin (1 µg/ml),
β-caryophyllene (1 µg/ml), or their combination in RAW264.7 cells.
Treatment with LPS caused a marked production of TNF-α (Fig. 4B), IL-6 (Fig. 5B), or IL-1β (Fig. 6B). These elevations were suppressed
by the addition of (+)-catechin (1 µg/ml), baicalin (1 µg/ml), or
β-caryophyllene (1 µg/ml). Interestingly, the combination of all
three factors caused a potential-suppressive effect on the
production of TNF-α (Fig. 4B), IL-6
(Fig. 5B), and IL-1β (Fig. 6B).
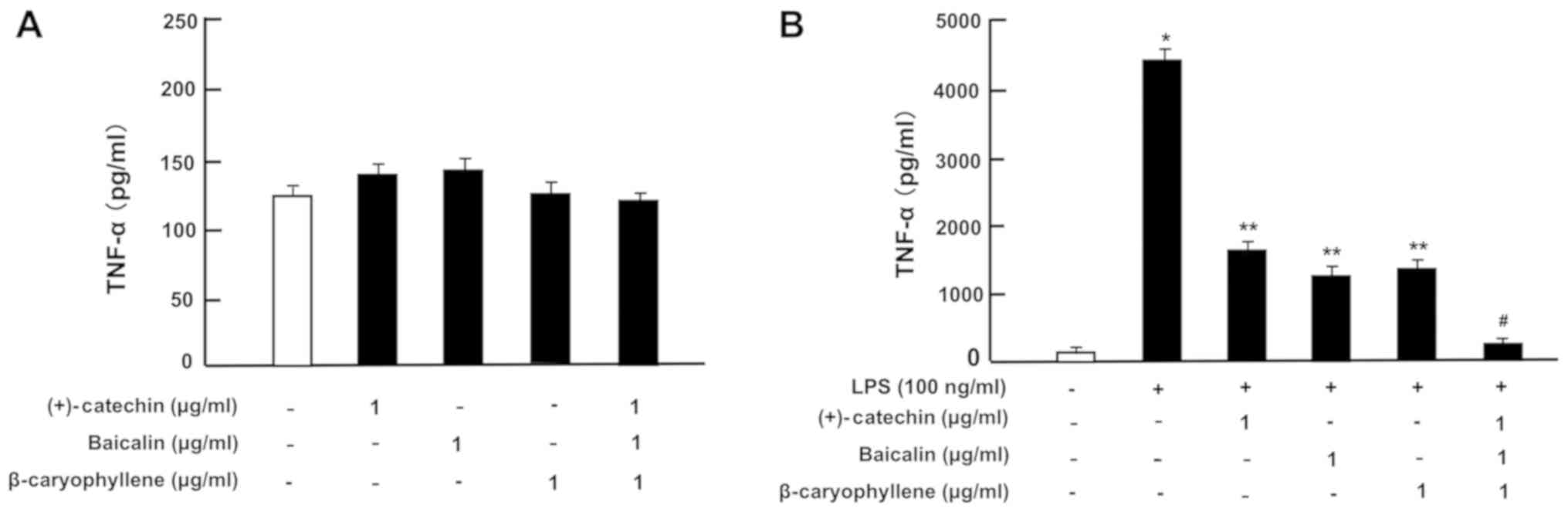 | Figure 4.Effects of (+)-catechin, baicaline,
and β-caryophylline on the production of TNF-α with LPS stimulation
in RAW264.7 cells. Cells (1×105/ml per well) were
cultured in the absence of botanical factors for 3 days in reaching
on subconfluence, and then were further incubated for 5 h after the
addition of either vehicle (1% DMSO), (+)-catechin (1 µg/ml),
baicalin (1 µg/ml), β-caryophyllene (1 µg/ml), or all three
combined factors (each 1 µg/ml) without (A) or with (B) LPS (100
ng/ml). After incubation, the medium was collected, and TNF-α
concentration in the mediun was determined. Data are presented as
mean ± SD obtained from 8 wells of 2 replicate wells per data set
using different dishes and cell preparations. *P<0.001 vs.
control (white bar); **P<0.001 vs. LPS (black bar);
#P<0.001 vs. LPS plus either (+)-catechin, baicalin,
or β-caryophyllene. One-way ANOVA, Tukey-Kramer post hoc test. LPS,
lipopolysaccharide; DMSO, dissolved in dimethylsulfoxide; SD,
standard deviation. |
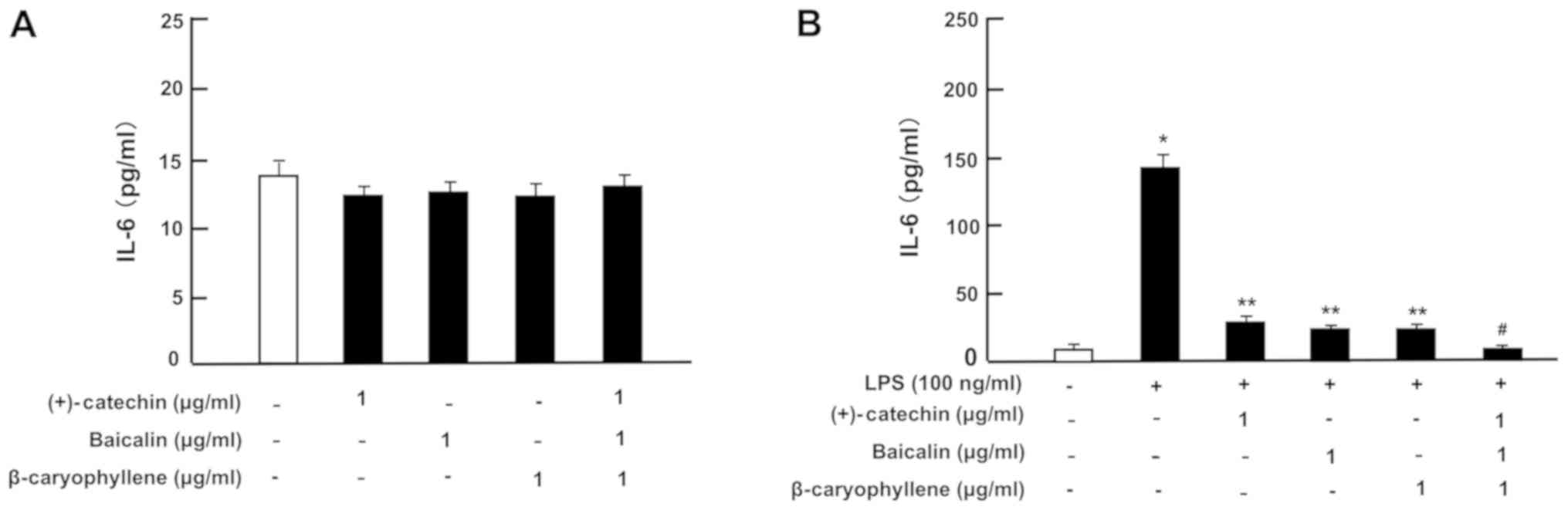 | Figure 5.Effects of (+)-catechin, baicaline,
and β-caryophylline on the production of IL-6 with LPS stimulation
in RAW264.7 cells. Cells (1×105/ml per well) were
cultured in the absence of botanical factors for 3 days in reaching
on subconfluence, and then were further incubated for 5 h after the
addition of either vehicle (1% DMSO), (+)-catechin (1 µg/ml),
baicalin (1 µg/ml), β-caryophyllene (1 µg/ml), or all three
combined factors (each 1 µg/ml) without (A) or with (B) LPS (100
ng/ml). After incubation, the medium was collected, and IL-6
concentration in the mediun was determined. Data are presented as
mean ± SD obtained from 8 wells of 2 replicate wells per data set
using different dishes and cell preparations. *P<0.001 vs.
control (white bar); **P<0.001 vs. LPS (black bar);
#P<0.001 vs. LPS plus either (+)-catechin, baicalin,
or β-caryophyllene. One-way ANOVA, Tukey-Kramer post hoc test. LPS,
lipopolysaccharide; DMSO, dissolved in dimethylsulfoxide; SD,
standard deviation. |
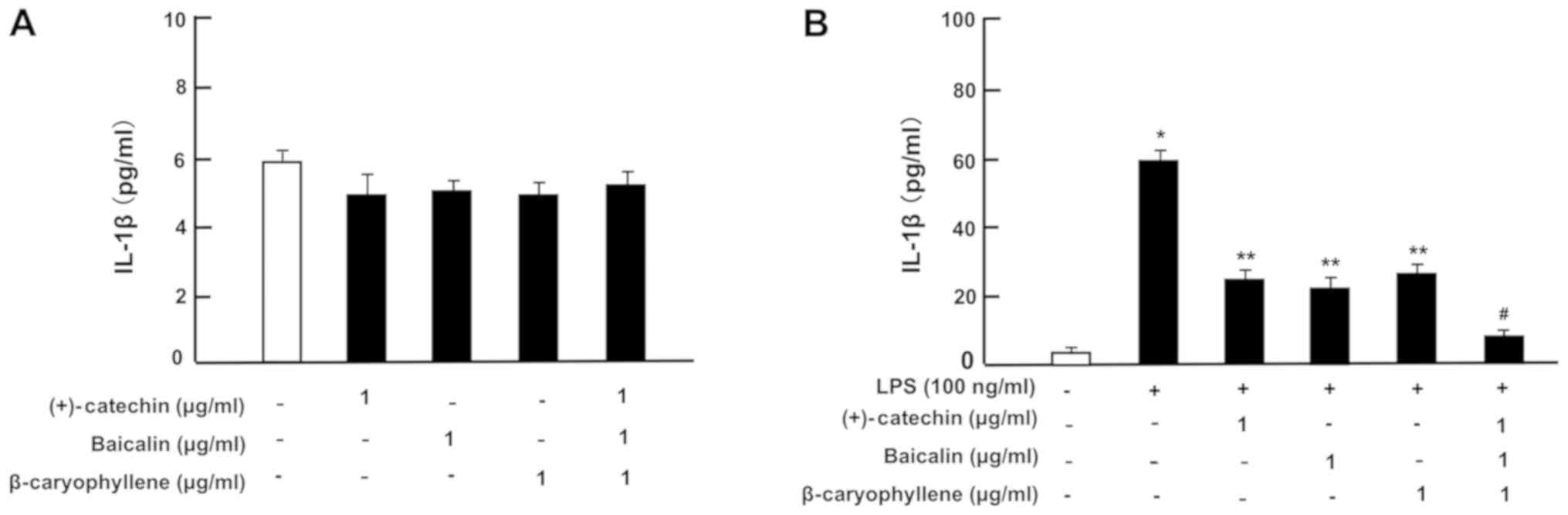 | Figure 6.Effects of (+)-catechin, baicaline,
and β-caryophylline on the production of IL-1β with LPS stimulation
in RAW264.7 cells. Cells (1×105/ml per well) were
cultured in the absence of botanical factors for 3 days in reaching
on subconfluence, and then were further incubated for 5 h after the
addition of either vehicle (1% DMSO), (+)-catechin (1 µg/ml),
baicalin (1 µg/ml), β-caryophyllene (1 µg/ml), or all three
combined factors (each 1 µg/ml) without (A) or with (B) LPS (100
ng/ml). After incubation, the medium was collected, and IL-1β
concentration in the mediun was determined. Data are presented as
mean ± SD obtained from 8 wells of 2 replicate wells per data set
using different dishes and cell preparations. *P<0.001 vs.
control (white bar); **P<0.001 vs. LPS (black bar);
#P<0.001 vs. LPS plus either (+)-catechin, baicalin,
or β-caryophyllene. One-way ANOVA, Tukey-Kramer post hoc test. LPS,
lipopolysaccharide; DMSO, dissolved in dimethylsulfoxide; SD,
standard deviation. |
Discussion
Current study was investigated to determine the
effects of the combination of (+)-catechin, baicalin and
β-caryophyllene, which reveal anti-inflammatory effects, on the
proliferation and death and the production of inflammatory
cytokines in mouse macrophage RAW264.7 cells in vitro. The
growth of cells was not altered in the presence of (+)-catechin,
baicalin, and β-caryophyllene. Of note, the three combined factors
were demonstrated to exhibit a potential-suppressive effect on the
proliferation and a potential -stimulatory effect on the death of
RAW264.7 cells in vitro. Current results confirmed our
previous findings that the three combination of (+)-catechin,
baicalin and β-caryophyllene reveals synergistic effects on the
reduction of the number of mouse macrophage RAW264.7 cells
(15), leading to the elimination of
macrophage related to inflammation.
Moreover, we investigated whether the combination of
(+)-catechin, baicalin and β-caryophyllene potently suppressed the
production of inflammatory cytokines, including TNF-α, IL-1β and
IL-6, under short-culture conditions, which did not reveal
suppressive effects on cell number with each factor alone or three
combined molecules (Fig. 2). Of
note, the combination of (+)-catechin, baicalin and β-caryophyllene
was found to exhibit an additive-depressive effect on the
production of cytokines as compared with that of each factor alone.
This result suggests that three combined factors have a
potential-suppressive effect on the production of cytokines
involved in inflammation. It is assumed that the three combined
factors suppress the production of inflammatory cytokines in a
short culture period, and this leads to a reduction in the number
of macrophages with longer culture in vitro.
In a previous study, in which RAW264.7 cells were
cultured with the combination of (+)-catechin, baicalin and
β-caryophyllen for 3 days, we demonstrated that the combined three
factors induced G2/M phase cell cycle arrest in the proliferation
of RAW264.7 cells using various inhibitors related to cell-cycle
arrest in vitro (15). The
suppressive effects on cell proliferation of three combined
botanical factors were suggested to link to inhibiting manifold
intracellular signaling pathways, which are related to signaling
pathways of PI3/Akt, ERK/MAPK, and Ca2+ in RAW264.7
cells, using various inhibitors (8,22–25).
Moreover, three combined factors were demonstrated to decrease the
protein levels of Akt and MAPKs (p44/42 and p38) in RAW264.7 cells.
The stimulatory effects of three combined botanical molecules on
cell death were related to the activation of caspase-3 (15), which activates nuclear DNA
fragmentation and induces apoptotic cell death (26).
In the current study, RAW264.7 cells, upon reaching
subconfluency, were cultured for 5 h after the treatment with the
combination of (+)-catechin, baicalin and β-caryophyllen, the
number of cells was not altered with these combinations. Of note,
the productions of cytokines with LPS activation was demonstrated
to potentially be suppressed by the treatment with three factors in
combination in RAW264.7 cells. The combination of (+)-catechin,
baicalin and β-caryophyllene was showed to decrease COX-1 and COX-2
in RAW264.7 cells in vitro (15). Inflammation-inducing factors are
reported to increase COX-2 and NF-κB p65, which is associated with
the mitogen-activated protein kinase (MAPK)-signaling pathways
(27–30). In addition, TNF-α-enhanced COX-2 and
NF-κB p65 levels were suppressed by culture with these three
botanical molecules in combination (15). LPS treatment has been reported to
enhance the production of TNF-α, IL-6 and IL-1β due to stimulating
signaling pathways linked to MAPK and NF-κB p65 in macrophage
RAW264.7 cells (31–33). It is possible that the combination of
(+)-catechin, baicalin and β-caryophyllene may exhibit potent
suppressive effects on the LPS-enhanced production of inflammatory
cytokines by inhibiting signaling pathways involved in MAPK and
NF-κB p65 in the activated macrophages. However, this remains to be
elucidated.
In conclusion, the present study demonstrates that
the LPS-enhanced production of inflammatory cytokines in mouse
macrophage RAW264.7 cells is potentially suppressed by the
treatment with the combination of (+)-catechin, baicalin and
β-caryophyllene with comparatively low levels, which did not show
any effects on the number of RAW264.7 cells. This combination may
be a useful anti-inflammatory therapeutic tool.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
MY conceived and designed the study, and performed
the experiment. MY and RL discussed the findings and interpreted
the results. MY wrote the manuscript, and RL reviewed and edited
the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the study are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Villanueva-Romero R, Gutiérrez-Cañas I,
Carrión M, Pérez García S, Seoane IV, Martínez C, Gomariz RP and
Juarranz Y: The anti-inflammatory mediator, vasoactive intestinal
peptide, modulates the differentiation and function of Th subsets
in rheumatoid arthritis. J Immunol Res. 2018:60437102018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kalaiselvi P, Rajashree K, Bharathi Priya
L and Padma VV: Cytoprotective effect of epigallocatechin-3-gallate
against deoxynivalenol-induced toxicity through anti-oxidative and
anti-inflammatory mechanisms in HT-29 cells. Food Chem Toxicol.
56:110–118. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morrison M, van der Heijden R, Heeringa P,
Kaijzel E, Verschuren L, Blomhoff R, Kooistra T and Kleemann R:
Epicatechin attenuates atherosclerosis and exerts anti-inflammatory
effects on diet-induced human-CRP and NFκB in vivo.
Atherosclerosis. 233:149–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qian Y, Chen Y, Wang L and Tou J: Effects
of baicalin on inflammatory reaction, oxidative stress and PKDl and
NF-kB protein expressions in rats with severe acute pancreatitis1.
Acta Cir Bras. 33:556–564. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bitto A, Squadrito F, Irrera N, Pizzino G,
Pallio G, Mecchio A, Galfo F and Altavilla D: Flavocoxid, a
nutraceutical approach to blunt inflammatory conditions. Mediators
Inflamm. 2014:7908512014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sharma C, Al Kaabi JM, Nurulain SM, Goyal
SN, Kamal MA and Ojha S: Polypharmacological properties and
therapeutic potential of β-caryophyllene: a dietary
phytocannabinoid of pharmaceutical promise. Curr Pharm Des.
22:3237–3264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klauke AL, Racz I, Pradier B, Markert A,
Zimmer AM, Gertsch J and Zimmer A: The cannabinoid CB2
receptor-selective phytocannabinoid beta-caryophyllene exerts
analgesic effects in mouse models of inflammatory and neuropathic
pain. Eur Neuropsychopharmacol. 24:608–620. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gertsch J, Leonti M, Raduner S, Racz I,
Chen JZ, Xie XQ, Altmann KH, Karsak M and Zimmer A and Zimmer A:
Beta-caryophyllene is a dietary cannabinoid. Proc Natl Acad Sci
USA. 105:9099–9104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ormeño E, Baldy V, Ballini C and Fernandez
C: Production and diversity of volatile terpenes from plants on
calcareous and siliceous soils: effect of soil nutrients. J Chem
Ecol. 34:1219–1229. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Katsuyama S, Mizoguchi H, Kuwahata H,
Komatsu T, Nagaoka K, Nakamura H, Bagetta G, Sakurada T and
Sakurada S: Involvement of peripheral cannabinoid and opioid
receptors in β-caryophyllene-induced antinociception. Eur J Pain.
17:664–675. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Paula-Freire LI, Andersen ML, Gama VS,
Molska GR and Carlini EL: The oral administration of
trans-caryophyllene attenuates acute and chronic pain in mice.
Phytomedicine. 21:356–362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chavan MJ, Wakte PS and Shinde DB:
Analgesic and anti-inflammatory activity of Caryophyllene oxide
from Annona squamosa L. bark. Phytomedicine. 17:149–151. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ghelardini C, Galeotti N, Di Cesare
Mannelli L, Mazzanti G and Bartolini A: Local anaesthetic activity
of beta-caryophyllene. Farmaco. 56:387–389. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martinez RM, Zarpelon AC, Cardoso RD,
Vicentini FT, Georgetti SR, Baracat MM, Andrei CC, Moreira IC,
Verri WA Jr and Casagrande R: Tephrosia sinapou ethyl acetate
extract inhibits inflammatory pain in mice: Opioid receptor
dependent inhibition of TNFα and IL-1β production. Pharm Biol.
51:1262–1271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamaguchi M and Levy RM: The combination
of β-caryophyllene, baicalin and catechin synergistically
suppresses the proliferation and promotes the death of RAW267.4
macrophages in vitro. Int J Mol Med. 38:1940–1946. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pomari E, Stefanon B and Colitti M: Effect
of plant extracts on H2O2-induced inflammatory gene expression in
macrophages. J Inflamm Res. 7:103–112. 2014.PubMed/NCBI
|
|
17
|
Yamaguchi M, Vikulina T, Arbiser JL and
Weitzmann MN: Suppression of NF-κB activation by gentian violet
promotes osteoblastogenesis and suppresses osteoclastogenesis. Curr
Mol Med. 14:783–792. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamaguchi M and Weitzmann MN: The bone
anabolic carotenoid p-hydroxycinnamic acid promotes osteoblast
mineralization and suppresses osteoclast differentiation by
antagonizing NF-κB activation. Int J Mol Med. 30:708–712. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamaguchi M and Daimon Y: Overexpression
of regucalcin suppresses cell proliferation in cloned rat hepatoma
H4-II-E cells: involvement of intracellular signaling factors and
cell cycle-related genes. J Cell Biochem. 95:1169–1177. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Izumi T and Yamaguchi M: Overexpression of
regucalcin suppresses cell death in cloned rat hepatoma H4-II-E
cells induced by tumor necrosis factor-alpha or thapsigargin. J
Cell Biochem. 92:296–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Okuno T, Gijón MA, Zarini S, Martin SA,
Barkley RM, Johnson CA, Ohba M, Yokomizo T and Murphy RC: Altered
eicosanoid production and phospholipid remodeling during cell
culture. J Lipid Res. 59:542–549. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Meijer L, Borgne A, Mulner O, Chong JP,
Blow JJ, Inagaki N, Inagaki M, Delcros JG and Moulinoux JP:
Biochemical and cellular effects of roscovitine, a potent and
selective inhibitor of the cyclin-dependent kinases cdc2, cdk2 and
cdk5. Eur J Biochem. 243:527–536. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Singh SV, Herman-Antosiewicz A, Singh AV,
Lew KL, Srivastava SK, Kamath R, Brown KD, Zhang L and Baskaran R:
Sulforaphane-induced G2/M phase cell cycle arrest involves
checkpoint kinase 2-mediated phosphorylation of cell division cycle
25C. J Biol Chem. 279:25813–25822. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Serrano-Nascimento C, da Silva Teixeira S,
Nicola JP, Nachbar RT, Masini-Repiso AM and Nunes MT: The acute
inhibitory effect of iodide excess on sodium/iodide symporter
expression and activity involves the PI3K/Akt signaling pathway.
Endocrinology. 155:1145–1156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen S, Wang Y, Ruan W, Wang X and Pan C:
Reversing multidrug resistance in hepatocellular carcinoma cells by
inhibiting extracellular signal-regulated kinase/mitogen-activated
protein kinase signaling pathway activity. Oncol Lett. 8:2333–2339.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao Y, Jing Z, Li Y and Mao W: Berberine
in combination with cisplatin suppresses breast cancer cell growth
through induction of DNA breaks and caspase-3-dependent apoptosis.
Oncol Rep. 36:567–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Echizen K, Hirose O, Maeda Y and Oshima M:
Inflammation in gastric cancer: interplay of the
COX-2/prostaglandin E2 and Toll-like receptor/MyD88 pathways.
Cancer Sci. 107:391–397. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin CC, Chan CM, Huang YP, Hsu SH, Huang
CL and Tsai SJ: Methylglyoxal activates NF-κB nuclear translocation
and induces COX-2 expression via a p38-dependent pathway in
synovial cells. Life Sci. 149:25–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li N, Liu BW, Ren WZ, Liu JX, Li SN, Fu
SP, Zeng YL, Xu SY, Yan X, Gao YJ, et al: GLP-2 attenuates
LPS-induced inflammation in BV-2 cells by inhibiting ERK1/2, JNK1/2
and NF-κB signaling pathway. Int J Mol Sci. 17:1902016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cha SM, Cha JD, Jang EJ, Kim GU and Lee
KY: Sophoraflavanone G prevents Streptococcus mutans surface
antigen I/II-induced production of NO and PGE2 by inhibiting
MAPK-mediated pathways in RAW 264.7 macrophages. Arch Oral Biol.
68:97–104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu X, Yin P, Wan C, Chong X, Liu M, Cheng
P, Chen J, Liu F and Xu J: Punicalagin inhibits inflammation in
LPS-induced RAW264.7 macrophages via the suppression of
TLR4-mediated MAPKs and NF-κB activation. Inflammation. 37:956–965.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cho BO, So Y, Jin CH, Nam BM, Yee ST and
Jeong IY: 3-deoxysilybin exerts anti-inflammatory effects by
suppressing NF-κB activation in lipopolysaccharide-stimulated
RAW264.7 macrophages. Biosci Biotechnol Biochem. 78:2051–2058.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao G, Zhang T, Ma X, Jiang K, Wu H, Qiu
C, Guo M and Deng G: Oridonin attenuates the release of
pro-inflammatory cytokines in lipopolysaccharide-induced RAW264.7
cells and acute lung injury. Oncotarget. 8:68153–68164.
2017.PubMed/NCBI
|














