Introduction
Colon cancer is a common malignancy of the digestive
tract with an increasing incidence each year (1). At present, 5-fluorouracil (5-Fu)-based
combination chemotherapy is considered the gold standard treatment;
however, it is associated with significant side effects and poor
tolerability (2). Despite the
improved prognosis associated with the use of targeted drugs, the
mortality rate remains high among patients with colon cancer, and
there is an evident clinical requirement for new treatment methods
(3). Gambogic acid (GA), a bioactive
compound extracted from certain Garcinia species, exhibits a
variety of antitumor activities against several types of cancer
(4–6). The limited information available
suggests that GA may inhibit colon cancer by suppressing cellular
activities, including proliferation, apoptosis, invasion and
migration (7).
The phosphoinositide 3-kinase (PI3K)/protein kinase
B (AKT) pathway is an important signaling pathway in cell
development and growth (8–10). Previous studies revealed that
suppressing the PI3K/AKT signaling pathway may inhibit cancer cell
proliferation and increase cancer cell apoptosis by regulating P21
expression (11,12). Other studies confirmed that
inhibition of the PI3K/AKT signaling pathway may suppress cancer
cell invasion and migration, possibly by regulating matrix
metalloprotease (MMP)-2 and −9 expression (13,14). The
current study evaluated the antitumor effect of GA and the
underlying mechanism in a human colon cancer cell line.
Materials and methods
Cells and reagents
Human colon cancer cell line SW620 was purchased
from American Type Culture Collection (Manassas, VA, USA). Fetal
bovine serum (FBS), RPMI-1640 medium and pancreatin were purchased
from Hyclone (GE Healthcare Life Sciences, Logan, UT, USA). GA
(purity, ≥95%) was purchased from Chengdu Pulis Biotech Co., Ltd.
(Chengdu, China). MTT, dimethylsulfoxide (DMSO) and 5-Fu were
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany),
while the Annexin V-fluorescein isothiocyanate (FITC)/propudium
iodide (PI) apoptosis and cell cycle assay kits [Cell cycle and
apoptosis (PI) kit] were purchased from BD Biosciences (San Jose,
CA, USA). Antibodies against PI3K (1:500; cat. no. ab32089), AKT
(1:500; cat. no. ab8805), phosphorylated (p)-AKT (1:500; cat. no.
ab8933), P21 (1:500; cat. no. ab109520), MMP-2 (1:500; cat. no.
ab37150), MMP-9 (1:500; cat. no. ab73734) and GAPDH (1:1,000; cat.
no. ab181602) were purchased from Abcam (Cambridge, MA, USA).
Furthermore, a DM3000 microscope (Leica Microsystems GmbH, Wetzlar,
Germany) and LightCycler® 480 instrument (Roche
Molecular Diagnostics, Pleasanton, CA, USA) were used in the
present study.
Cell culture and experimental
design
SW620 cells were cultured in RPMI-1640 medium with
10% FBS in an incubator at 37°C and 5% CO2 and passaged
upon reaching 90% confluence. The culture was divided into five
groups based on subculturing conditions: i) No-treatment control
(NC) group cultured in RPMI-1640 medium with 10% FBS; ii) NC group
medium + 10 µg/ml GA (low group); iii) NC group medium + 50 µg/ml
GA (medium group); iv) NC group medium + 100 µg/ml GA (high group);
and v) NC group medium + 10 µg/ml 5-Fu (5-Fu group). Cells were
cultured in their respective experimental group media as described
below for each specific assay. Cells were cultured until the
logarithmic growth stage and digested with trypsin prior to seeding
into culture plates for specific experiments.
MTT assay
A 200-µl cell suspension (4×103 cells/ml)
was cultured in 6-well-plates for 5 h to allow time for cell
adherence, after which the medium was replaced with the
group-specific experimental culture media, as described above. Each
group was cultured in triplicate. After 48 h at room temperature,
20 µl MTT (5 g/ml) was added to each well and the plates were
cultured for 30 h at room temperature. DMSO was added to each well
to stop the reaction, and the OD value was measured at 490 nm on
the plate reader to calculate cell proliferation.
Apoptosis assay
SW620 cells (1×106 cells/ml) were seeded
in a 6-well plate (2 ml/well) and cultured for 24 h in
group-specific media at room temperature. Media were removed and
cells collected by resuspending in 300 µl binding buffer, to which
5 µl Annexin V-FITC was added according to the manufacturer's
protocol. Suspensions were incubated at room temperature in the
dark for 15 min, after which 5 µl PI was added. Apoptosis was
detected by flow cytometry (FACSAria; BD Biosciences; wavelength,
488 nm) supplemented with 200 µl 1X binding buffer (diluted in
double distilled water) at room temperature after 1 h. All tests
were performed in triplicate. Data were analyzed using Diva
software (version 8.0.1; FACSAria; BD Biosciences).
Cell cycle assay
SW620 cells in the logarithmic growth phase were
seeded in a 7.5-cm culture flask and cultured for 24 h, then
digested by pancreatin and pelleted by centrifugation (1,000 × g at
4°C for 5 min). Cell cycle status was detected using the
aforementioned cell cycle kit according to the manufacturer's
protocol. Briefly, the cells of each group were treated with their
respective aforementioned conditioned media for 24 h at room
temperature. Afterwards, 5 µl RNase (10 mg/ml) was added to the
PBS-resuspended cell pellet, and the cells were cultured at room
temperature for 1 h. PI (100 µg/ml) was subsequently added and the
cells incubated at room temperature for 30 min. The cell cycle
status was measured in triplicate by flow cytometry (FACSAria; BD
Biosciences) at a wavelength of 488 nm and data were analyzed using
Diva software (version 8.0.1; FACSAria; BD Biosciences).
Transwell invasion assay
Matrigel (1:3) was diluted with serum-free medium
(RPMI-1640) and placed in the upper transwell chamber (pore size, 8
µm) at 37°C for 4 h, after which the transwell chambers were placed
in 24-well plates. SW620 cells (1×104 cells/ml; 100
µl/well cell suspension) from each individual experimental group
were added to the upper chamber. Complete culture medium
(RPMI-1640) containing 10% FBS was added to the lower chamber, and
the cells were cultured for 24 h. Subsequently, noninvasive cells
were removed with cotton swabs. The transwell chambers were removed
from the wells, washed with phosphate-buffered saline (PBS) three
times and the remaining cells in the lower chamber were fixed with
4% paraformaldehyde-PBS solution for 10 min at room temperature.
The chambers were subsequently washed with PBS as aforementioned,
inverted and dried. Cells were stained with 0.1% cresyl violet
solution for 20 min at room temperature and washed with PBS three
times. Cells were subsequently counted under an inverted optical
microscope.
Wound healing assay
SW620 cells (5×104/well) from each
experimental group were seeded into a six-well plate and cultured
for 24 h at room temperature. When the cells formed a single,
tightly adherent layer, a scraper with a 2-mm wide tip was used to
scratch the confluent cells. Images of each scratch were captured
under a light microscope at 0 h. Plates were cultured and images
were captured again under the light microscope at 48 h. Image-Pro
Plus software (version 6.0; Media Cybernetics, Inc., Rockville, MD,
USA) was used to measure the scratch width in the 0- and 48-h
images and to calculate the cell migration rate from the
difference.
Western blotting
Cells were incubated for 24 h at room temperature in
the aforementioned conditioned media and collected using lysis
buffer (Beyotime Institute of Biotechnology) and the protein
concentration was measured using a bicinchoninic acid Protein kit
(Beyotime Institute of Biotechnology). Subsequently, cells of the
different groups were washed three times with ice-cold PBS and
lysed with buffer containing 50 mM Tris-HCl (pH 7.6), 150 mM NaCl,
1 mM EDTA, 1% NP-40, 0.5% Na-deoxycholate, 5 µg/ml aprotinin, 5
µg/ml leupeptin and 1 mM phenylmethylsulphonyl fluoride. Cell
lysates were cleared via centrifugation at 12,000 × g at 4°C for 30
min and denatured by boiling in Laemmli buffer (Beyotime Institute
of Biotechnology). Proteins were denatured by incubation in a
boiling water bath for 5 min with 5X SDS gel buffer solution. A
total of 15 µl/lane of each sample was electrophoresed on an 8%
SDS-PAGE (110V; 4 h). The protein samples were transferred to
polyvinylidene fluoride (PVDF) membranes using the semi-dry method.
The PVDF membranes were blocked with 5% skim milk in PBS for 1.5 h
at room temperature. The membranes were subsequently incubated with
the aforementioned primary antibodies overnight at 4°C. Following
this incubation, the membrane was washed three times in PBS and
incubated in goat anti-rabbit second antibody (1:5,000; cat. no.
ab205718; Abcam) solution at room temperature for 2 h. Anti-GAPDH
antibody was used as an internal reference. Specific immune
complexes were detected using the Western Blotting Plus
Chemiluminescence Reagent (Thermo Fisher Scientific, Inc.). Band
intensity was quantified via densitometry analysis using Image-Pro
Plus version 4.5 software (Media Cybernetics, Inc.).
Statistical analysis
Data are presented as the mean ± standard deviation
from three independent experiments. Quantitative results were
analyzed via one-way ANOVA assay followed by a Tukey's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
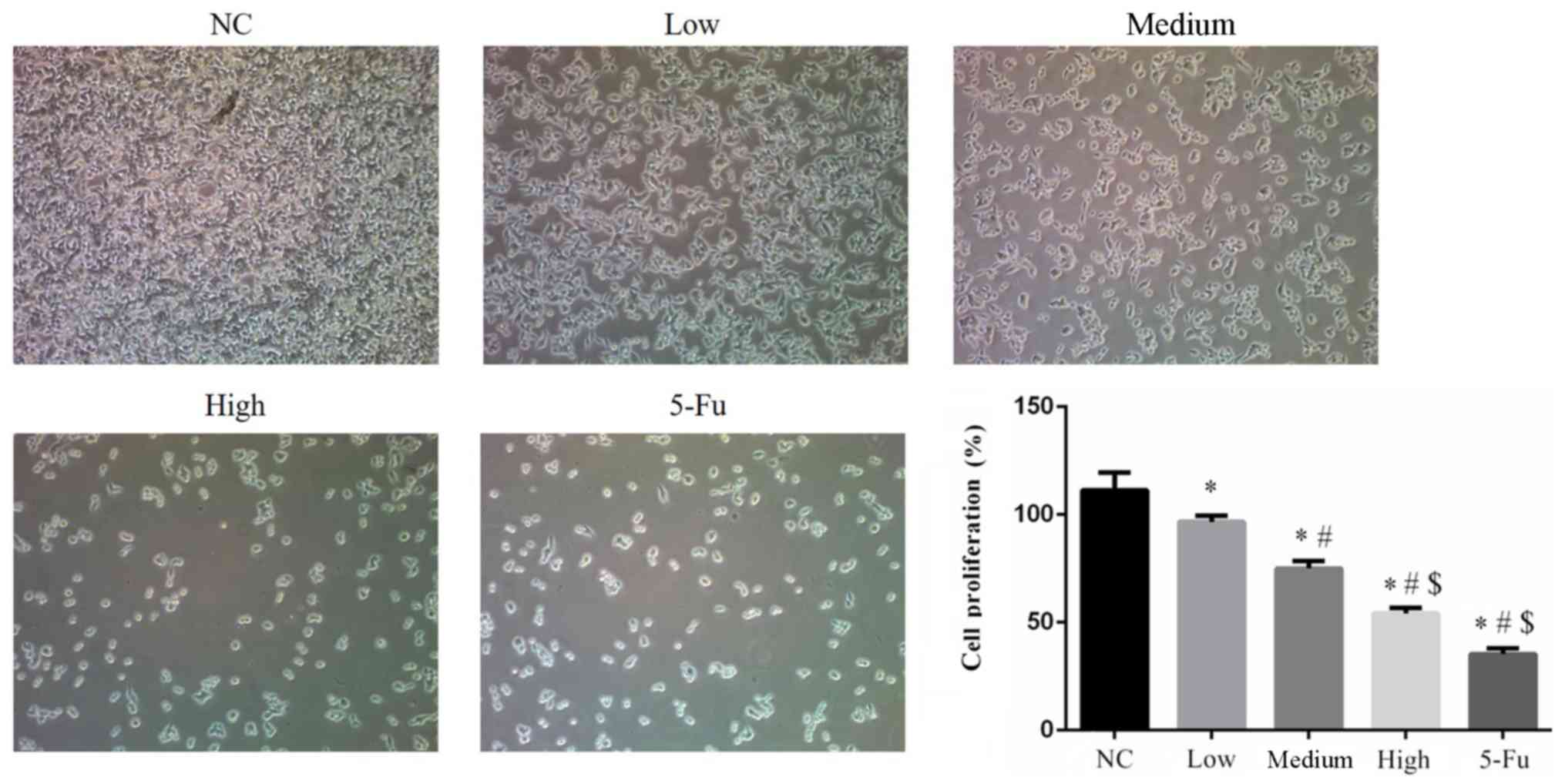
GA suppresses cell proliferation
Cell proliferation rates of the GA-treated groups
were significantly suppressed compared with the NC group (all
P<0.05). The proliferation rate of 5-Fu treated cells was
significantly suppressed when compared with the NC, low and medium
groups (each, P<0.05). Furthermore, significant differences in
proliferation rates were identified between the GA-treated groups
(low, medium and high; all P<0.05; Fig. 1).
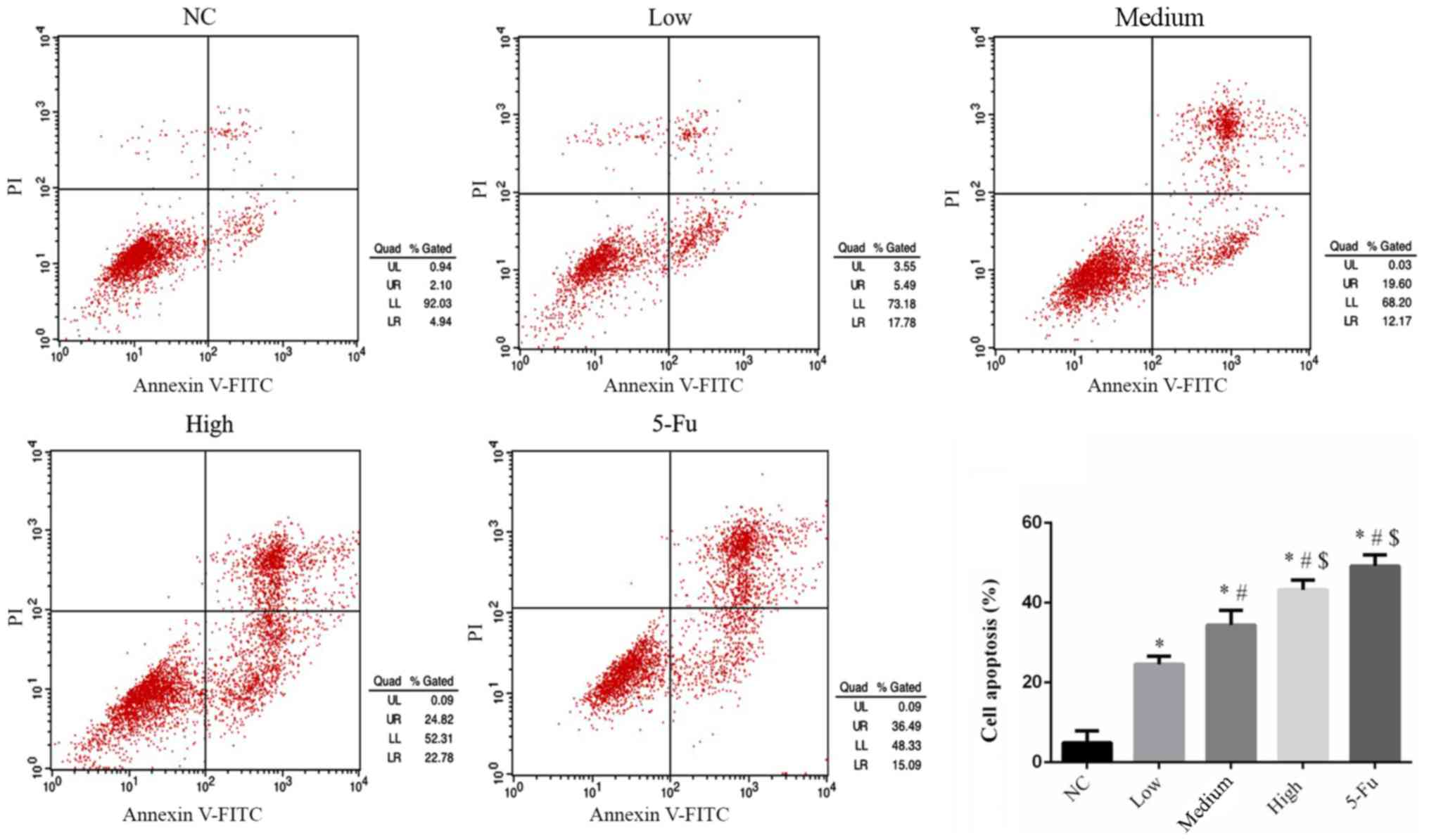
GA influences cell apoptosis
rates
Compared with the NC group, apoptosis rates in all
GA-treated groups were significantly upregulated (P<0.05;
Fig. 2). The rates of apoptosis
increased in a dose-dependent manner. The apoptosis rate of the
5-Fu group was significantly upregulated compared with the NC, low
and medium groups (each, P<0.05; Fig.
2).
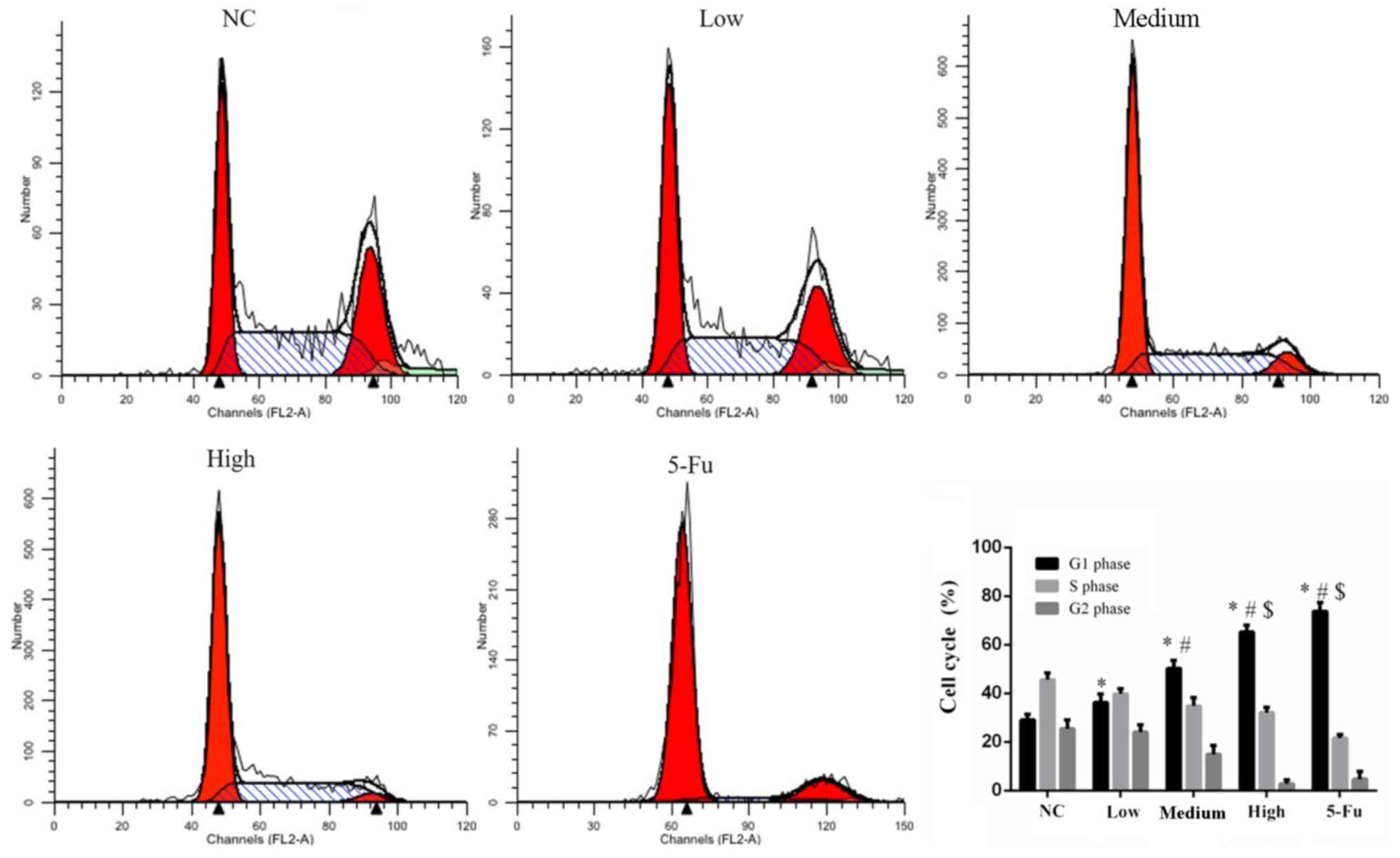
GA affects the cell cycle
A significantly increased number of G1-phase cells
was observed among GA treated groups compared with the NC group
(all P<0.05; Fig. 3). The number
of 5-Fu treated cells in the G1 phase were significantly increased
compared with the NC, low and medium groups (each, P<0.05;
Fig. 3). The increase in the number
of G1-phase cells was dose dependent and the differences between
GA-treated groups were statistically significant (P<0.05).
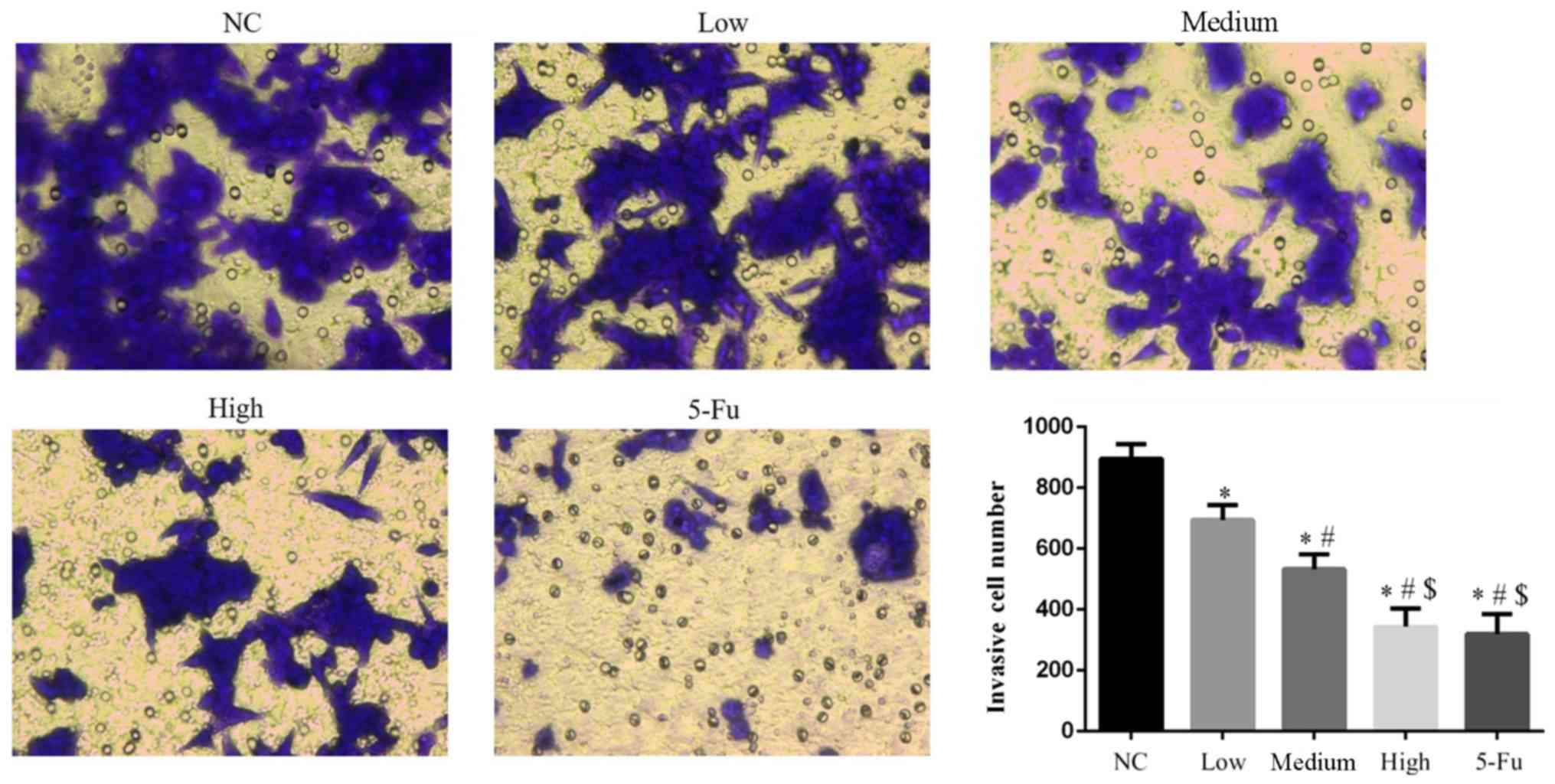
GA suppresses cell invasion
Counts of invasive cells were significantly lower in
all GA groups compared with the NC group (all P<0.05; Fig. 4). The number of invasive cells in the
5-Fu group were significantly reduced when compared with the NC,
low and medium groups (each, P<0.05). Significant differences
were also observed between GA-treated groups (P<0.05).
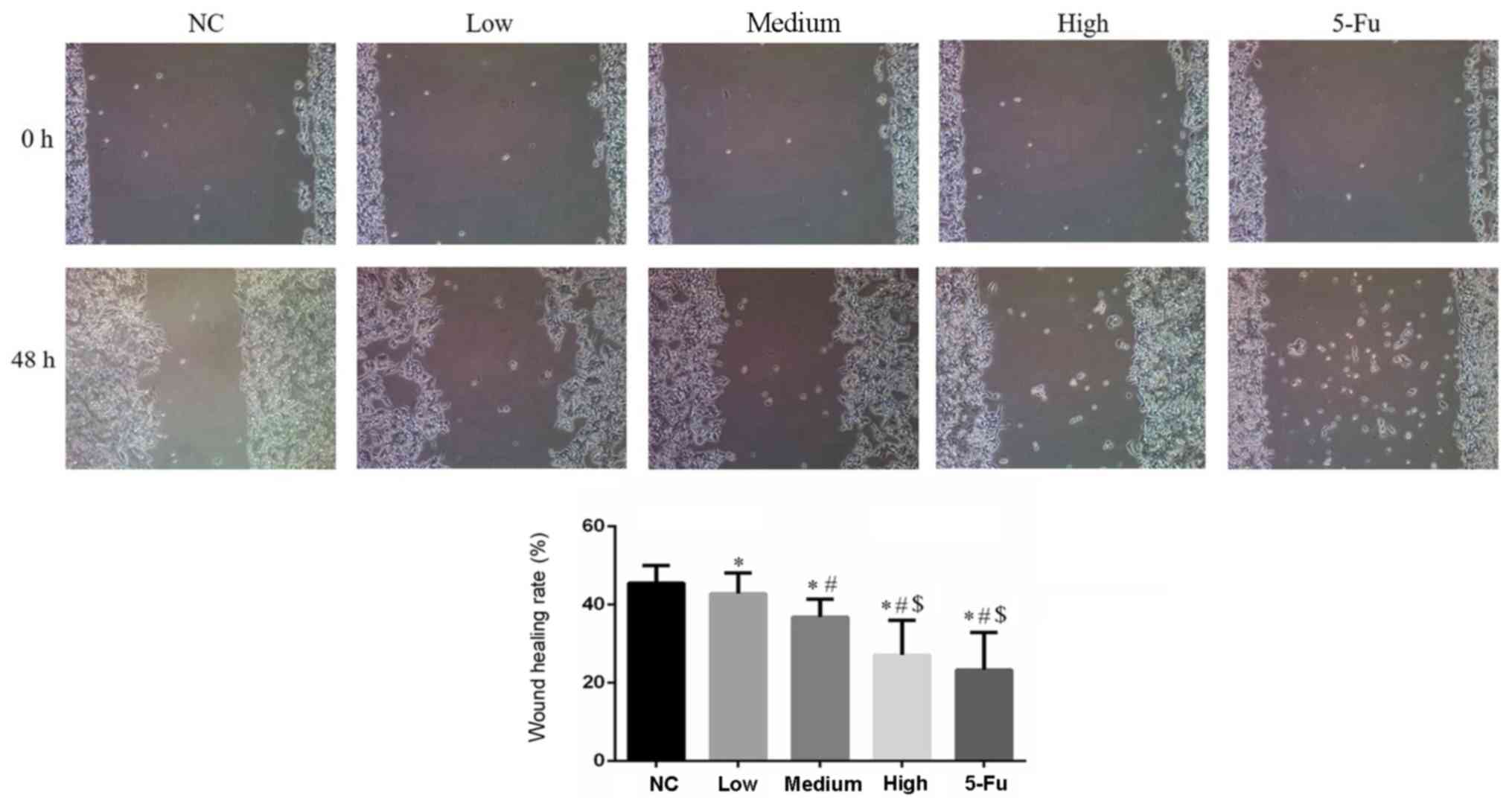
GA affects cell migration
The wound healing rates of GA-treated groups were
significantly inhibited compared with the NC group (P<0.05;
Fig. 5). The wound healing rate of
the 5-Fu group was significantly reduced compared with the NC, low
and medium groups. This effect was dose-dependent.
GA affects the PI3K/AKT pathway
protein expression
Compared with the NC group, protein expression
levels of PI3K, AKT, p-AKT, MMP-2 and −9 significantly decreased,
and the expression levels of P21 significantly increased following
treatment with GA at low, medium and high doses (P<0.05;
Fig. 6). Furthermore, the p-AKT/AKT
ratio decreased in all GA-treated groups compared with the NC group
and this effect was dose-dependent (P<0.05).
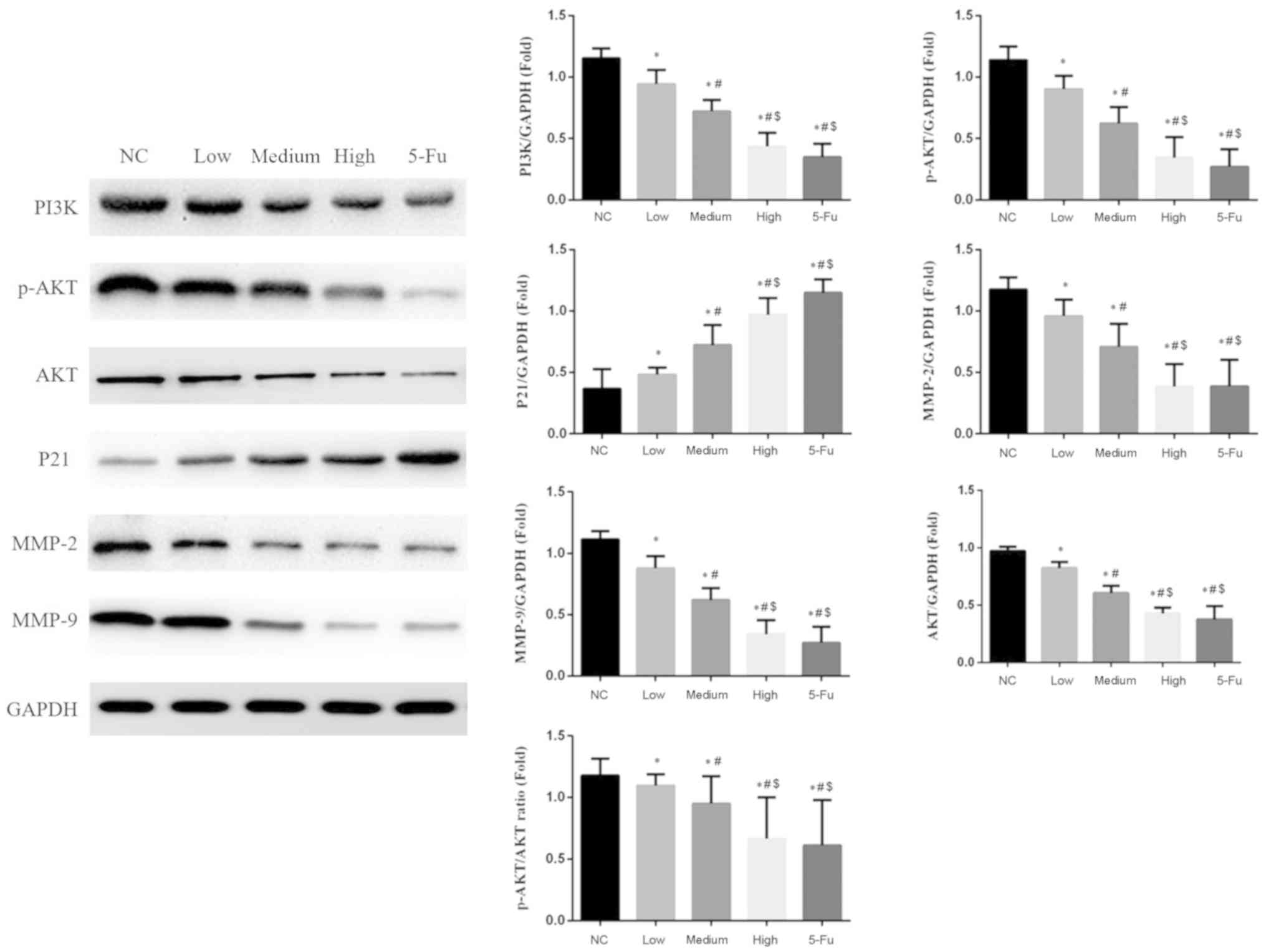 | Figure 6.Relative protein expression levels in
different groups. *P<0.05 vs. the NC group;
#P<0.05 vs. the low group; $P<0.05 vs.
the medium group. NC, no-treatment control group; low, group
treated with 10 µg/ml GA; medium, group treated with 50 µg/ml GA;
high, group treated with 100 µg/ml GA; 5-Fu, group treated with 10
µg/ml 5-fluorouracil; GA, gambogic acid; PI3K, phosphoinositide
3-kinase; AKT, protein kinase B; p-, phosphorylated; MMP, matrix
metalloprotease. |
Discussion
GA is the main active ingredient in gamboge. In
Traditional Chinese Medicine, GA is used to treat ulcers, swelling
and poisoning (15,16). Previous studies have revealed
antitumor effects of GA, including induction of apoptosis, and
inhibition of proliferation and pro-neoplastic protein expression
(17–20). GA was determined to selectively kill
cancer cells without affecting normal hematopoietic cells (21). Previous studies revealed that GA
exhibited antitumor effects in colon cancer, however, the
underlying mechanism of action remains unclear (22,23). In
the present study, GA exhibited antitumor effects by inhibiting
cell proliferation, invasion and migration and increasing
apoptosis. Furthermore, GA arrested cells in the G1 phase of the
cell cycle. At present, 5-Fu is the preferred drug for the clinical
treatment of tumors. Therefore, 5-Fu was used as a positive control
in the present study.
The PI3K/AKT signaling pathway is activated in the
majority of tumors (24,25), which further enhances
tumor-associated cell activity, including proliferation, invasion
and migration (25). In the current
study, GA inhibited the expression of PI3K, AKT and p-AKT in human
colon cancer SW620 cells. Furthermore, the ratio of p-AKT to AKT
decreased significantly following treatment with GA indicating that
inhibition of the AKT-associated signaling may mediate the
antitumor activity of GA. P21 is an important protein, which is
downstream of AKT, that serves as a suppressor of tumor development
(26) and can prevent cells from
exiting the G1 phase (27–29). In the current study, following
treatment with GA, the protein expression of P21 increased along
with a dose-dependent increase in the number of cells in the G1
phase of the cell cycle.
Migration of cancer cells from the primary tumor and
subsequent invasion through the tissue barrier is key to metastasis
(30,31). Extracellular matrix and basement
membranes are a natural barrier in the process of tumor
infiltration and diffusion, and MMPs that degrade the matrix are
important for promoting tumor invasion and metastasis (32,33).
Previous studies indicated that MMP-2 and −9 increase cancer cell
invasion and migration and are the downstream gene targets in the
PI3K/AKT pathway (34,35). The present results suggest that GA
suppressed SW620 cell invasion and migration, and regulated the
MMP-2 and −9 protein expression via downregulation of the PI3K/AKT
signaling pathway. GA appeared to suppress SW620 cancer cell
pro-neoplastic and pro-metastatic activities, including
proliferation, invasion and migration, by altering the PI3K/AKT/P21
signaling and MMP-2 and −9 activity in vitro.
The current study was limited by the use of a single
cell line and lack of in vivo experiments. In future
studies, the effects of GA should be investigated in different
colon cancer cell lines and in vivo.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZZ designed the current study, acquired/analyzed the
data and drafted the manuscript. JM designed the current study,
acquired the data and revised the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bernadach M, Lapeyre M, Dillies AF, Miroir
J, Moreau J, Kwiatkowski F, Pham-Dang N, Saroul N, Durando X and
Biau J: Toxicity of docetaxel, platine, 5-fluorouracil-based
induction chemotherapy for locally advanced head and neck cancer:
The importance of nutritional status. Cancer Radiother. 23:273–280.
2019.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zu M, Ma L, Zhang X, Xie D, Kang Y and
Xiao B: Chondroitin sulfate-functionalized polymeric nanoparticles
for colon cancer-targeted chemotherapy. Colloids Surf B
Biointerfaces. 177:399–406. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ishaq M, Khan MA, Sharma K, Sharma G,
Dutta RK and Majumdar S: Gambogic acid induced oxidative stress
dependent caspase activation regulates both apoptosis and autophagy
by targeting various key molecules (NF-κB, Beclin-1, p62 and NBR1)
in human bladder cancer cells. Biochim Biophys Acta.
1840:3374–3384. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang J and Yuan Z: Gambogic acid
sensitizes ovarian cancer cells to doxorubicin through ROS-mediated
apoptosis. Cell Biochem Biophys. 67:199–206. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang S, Wang L, Chen M and Wang Y:
Gambogic acid sensitizes resistant breast cancer cells to
doxorubicin through inhibiting P-glycoprotein and suppressing
survivin expression. Chem Biol Interact. 235:76–84. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang W, Wang X, Shi C, Guo D, Xu G, Wang
L, Bodman A and Luo J: Fine-tuning vitamin E-containing
telodendrimers for efficient delivery of gambogic acid in colon
cancer treatment. Mol Pharm. 12:1216–1229. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Danielsen SA, Eide PW, Nesbakken A, Guren
T, Leithe E and Lothe RA: Portrait of the PI3K/AKT pathway in
colorectal cancer. Biochim Biophys Acta. 1855:104–121.
2015.PubMed/NCBI
|
|
9
|
Polivka J Jr and Janku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Faes S and Dormond O: PI3K and AKT:
Unfaithful partners in cancer. Int J Mol Sci. 16:21138–21152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chang YL, Zhou PJ, Wei L, Li W, Ji Z, Fang
YX and Gao WQ: MicroRNA-7 inhibits the stemness of prostate cancer
stem-like cells and tumorigenesis by repressing KLF4/PI3K/Akt/p21
pathway. Oncotarget. 6:24017–24031. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zheng L, Zhang Y, Liu Y, Zhou M, Lu Y,
Yuan L, Zhang C, Hong M, Wang S and Li X: MiR-106b induces cell
radioresistance via the PTEN/PI3K/AKT pathways and p21 in
colorectal cancer. J Transl Med. 13:2522015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou R, Xu L, Ye M, Liao M, Du H and Chen
H: Formononetin inhibits migration and invasion of MDA-MB-231 and
4T1 breast cancer cells by suppressing MMP-2 and MMP-9 through
PI3K/AKT signaling pathways. Horm Metab Res. 46:753–760. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li W, Liu Z, Zhao C and Zhai L: Binding of
MMP-9-degraded fibronectin to β6 integrin promotes invasion via the
FAK-Src-related Erk1/2 and PI3K/Akt/Smad-1/5/8 pathways in breast
cancer. Oncol Rep. 34:1345–1352. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fu Q, Li C and Yu L: Gambogic acid
inhibits spinal cord injury and inflammation through suppressing
the p38 and Akt signaling pathways. Mol Med Rep. 17:2026–2032.
2018.PubMed/NCBI
|
|
16
|
Wu X, Long L, Liu J, Zhang J, Wu T, Chen
X, Zhou B and Lv TZ: Gambogic acid suppresses inflammation in
rheumatoid arthritis rats via PI3K/Akt/mTOR signaling pathway. Mol
Med Rep. 16:7112–7118. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Duan D, Zhang B, Yao J, Liu Y, Sun J, Ge
C, Peng S and Fang J: Gambogic acid induces apoptosis in
hepatocellular carcinoma SMMC-7721 cells by targeting cytosolic
thioredoxin reductase. Free Radic Biol Med. 69:15–25. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi X, Chen X, Li X, Lan X, Zhao C, Liu S,
Huang H, Liu N, Liao S, Song W, et al: Gambogic acid induces
apoptosis in imatinib-resistant chronic myeloid leukemia cells via
inducing proteasome inhibition and caspase-dependent Bcr-Abl
downregulation. Clin Cancer Res. 20:151–163. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang LH, Yang JY, Yang SN, Li Y, Ping GF,
Hou Y, Cui W, Wang ZZ, Xiao W and Wu CF: Suppression of NF-κB
signaling and P-glycoproteins function by gambogic acid
synergistically potentiates adriamycin-induced apoptosis in lung
cancer. Curr Cancer Drug Targets. 14:91–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chi Y, Zhan XK, Yu H, Xie GR, Wang ZZ,
Xiao W, Wang YG, Xiong FX, Hu JF, Yang L, et al: An open-labeled,
randomized, multicenter phase II a study of gambogic acid injection
for advanced malignant tumors. Chin Med J (Engl). 126:1642–1646.
2013.PubMed/NCBI
|
|
21
|
Wang X and Chen W: Gambogic acid is a
novel anti-cancer agent that inhibits cell proliferation,
angiogenesis and metastasis. Anticancer Agents Med Chem.
12:994–1000. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei F, Zhang T, Yang Z, Wei JC, Shen HF,
Xiao D, Wang Q, Yang P, Chen HC, Hu H, et al: Gambogic acid
efficiently kills stem-like colorectal cancer cells by upregulating
ZFP36 expression. Cell Physiol Biochem. 46:829–846. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao G, Bian Y, Qian H, Yang M, Hu J, Li L,
Yu L, Liu B and Qian X: Gambogic acid regulates the migration and
invasion of colorectal cancer via microRNA-21-mediated activation
of phosphatase and tensin homolog. Exp Ther Med. 16:1758–1765.
2018.PubMed/NCBI
|
|
24
|
Toren P and Zoubeidi A: Targeting the
PI3K/Akt pathway in prostate cancer: Challenges and opportunities
(review). Int J Oncol. 45:1793–1801. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen J, Shao R, Li F, Monteiro M, Liu JP,
Xu ZP and Gu W: PI3K/Akt/mTOR pathway dual inhibitor BEZ235
suppresses the stemness of colon cancer stem cells. Clin Exp
Pharmacol Physiol. 42:1317–1326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu S, Ren C, Liu Y and Epner DE: PI3K-Akt
signaling is involved in the regulation of p21(WAF/CIP) expression
and androgen-independent growth in prostate cancer cells. Int J
Oncol. 28:245–251. 2006.PubMed/NCBI
|
|
27
|
Yeo D, He H, Baldwin GS and Nikfarjam M:
The role of p21-activated kinases in pancreatic cancer. Pancreas.
44:363–369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei CY, Tan QX, Zhu X, Qin QH, Zhu FB, Mo
QG and Yang WP: Expression of CDKN1A/p21 and TGFBR2 in breast
cancer and their prognostic significance. Int J Clin Exp Pathol.
8:14619–14629. 2015.PubMed/NCBI
|
|
29
|
Li J, Li Z, Kan Q, Sun S, Li Y and Wang S:
Association of p21 3′UTR gene polymorphism with cancer risk:
Evidence from a meta-analysis. Sci Rep. 5:131892015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nakayama H, Ohuchida K, Yonenaga A, Sagara
A, Ando Y, Kibe S, Takesue S, Abe T, Endo S, Koikawa K, et al:
S100P regulates the collective invasion of pancreatic cancer cells
into the lymphatic endothelial monolayer. Int J Oncol. 55:211–222.
2019.PubMed/NCBI
|
|
31
|
Kapali AS, George NA, Iype EM, Thomas S,
Varghese BT, Balagopal PG and Sebastian P: Retrospective outcome
analysis of Buccal mucosal and lower alveolar squamous cell
carcinoma from a high-volume tertiary cancer Centre. Indian J Surg
Oncol. 10:286–291. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Iochmann S, Bléchet C, Chabot V, Saulnier
A, Amini A, Gaud G, Gruel Y and Reverdiau P: Transient RNA
silencing of tissue factor pathway inhibitor-2 modulates lung
cancer cell invasion. Clin Exp Metastasis. 26:457–467. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Safranek J, Pesta M, Holubec L, Kulda V,
Dreslerova J, Vrzalova J, Topolcan O, Pesek M, Finek J and Treska
V: Expression of MMP-7, MMP-9, TIMP-1 and TIMP-2 mRNA in lung
tissue of patients with non-small cell lung cancer (NSCLC) and
benign pulmonary disease. Anticancer Res. 29:2513–2518.
2009.PubMed/NCBI
|
|
34
|
Yuan H, Yang P, Zhou D, Gao W, Qiu Z, Fang
F, Ding S and Xiao W: Knockdown of sphingosine kinase 1 inhibits
the migration and invasion of human rheumatoid arthritis
fibroblast-like synoviocytes by down-regulating the PI3K/AKT
activation and MMP-2/9 production in vitro. Mol Biol Rep.
41:5157–5165. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fan Z, Duan X, Cai H, Wang L, Li M, Qu J,
Li W, Wang Y and Wang J: Curcumin inhibits the invasion of lung
cancer cells by modulating the PKCα/Nox-2/ROS/ATF-2/MMP-9 signaling
pathway. Oncol Rep. 34:691–698. 2015. View Article : Google Scholar : PubMed/NCBI
|