Introduction
Angiogenesis is the formation of new blood vessels
from pre-existing vasculature (1).
It is a complex process that involves degradation of the
extracellular matrix, the proliferation and migration of
endothelial cells and the sprouting of vessels (2). Physiological angiogenesis is of
considerable significance in growth, development, wound healing and
the normal function of several organs. However, the pathological
form of angiogenesis can lead to several diseases, including tumor
formation, retinopathy and rheumatoid arthritis (1,2).
Angiogenic retinopathy, including diabetic retinopathy, retinopathy
of prematurity and retinal vein occlusion, can eventually lead to
blindness (3). Although certain
therapeutic drugs have been shown to disrupt retinal pathological
angiogenesis, several disadvantages have been reported with this
type of treatment, including tumor drug resistance and adverse
effects (4). Therefore, novel
therapeutic targets are required for the treatment of retinal
pathological angiogenesis and for investigating the potential
molecular mechanisms associated with this process.
Cyclin-dependent kinases (CDKs) are a family of
multifunctional enzymes that interact with cyclins to regulate the
cell cycle (5). Several checkpoints
are present in the cell cycle that ensure accurate cell division.
Cyclins bind to CDKs and cause their activation in order to
regulate checkpoint transition (6).
CDK1 serves an important role in the control of the cell cycle by
regulating the centrosome cycle, promoting G2-M transition and
modulating G1 phase progression (7).
It has also been shown that CDK1 is involved in the regulation of
apoptosis (8,9). The cell cycle and apoptosis are well
known biological processes that are involved in the development of
several pathologies, including tumors and angiogenesis (10). CDK1 is overexpressed in several
tumors and contributes to their development (11–14).
However, the expression levels and the role of CDK1 in angiogenesis
have not been investigated in detail.
The present study investigated the expression levels
of CDK1 in pathological angiogenesis using an oxygen-induced
retinopathy (OIR) mouse model. Furthermore, the effects of CDK1
knockdown on angiogenesis were examined using small interfering
(si)RNAs in vitro, and the underlying molecular mechanism
was investigated. The present study highlights the role of CDK1 in
retinal angiogenesis and may provide novel insight for the
development of treatments against pathological
neovascularization.
Materials and methods
Reagents
The antibodies targeting GAPDH (cat. no.
10494-1-AP), CD31 (cat. no. 11265-1-AP), cyclin A1 (cat. no.
13295-1-AP), cyclin B1 (cat. no. 55004-1-AP), cyclin D1 (cat. no.
60186-1-1 g), cyclin E1 (cat. no. 11554-1-AP), CDK1 (cat. no.
19532-1-AP), CDK2 (cat. no. 10122-1-AP), poly (ADP-ribose)
polymerase (PARP)1 (cat. no. 13371-1-AP), p21 (cat. no. 10355-1-AP)
and p53 (cat. no. 10442-1-AP) were purchased from Proteintech
Group, Inc. DMEM, fetal bovine serum (FBS), DAPI,
TRIzol®, Super Signal West Pico chemiluminescent
substrate, and HRP-(cat. no. A-10677), FITC-(cat. no. A-10683) and
Alexa Fluor555-conjugated secondary antibodies (cat. no. A-27017)
were obtained from Thermo Fisher Scientific, Inc. Propidium iodide
(PI) was obtained from Beyotime Institute of Biotechnology. The
PrimeScript RT kit and SYBR Premix were purchased from Takara
Biotechnology Co., Ltd. (Dalian, China). The Annexin V-FITC
apoptosis kit was purchased from eBioscience (San Diego, CA, USA).
Griffonia simplicifolia isolectin B4 was purchased from
Invitrogen (Thermo Fisher Scientific, Inc.). Matrigel Matrix was
purchased from BD Transduction Laboratories (Shanghai, China). The
EdU staining kit was obtained from Guangzhou RiboBio Co., Ltd.
(Guangzhou, China). Bovine serum albumin was obtained from Weiao
Bio (Shanghai, China).
OIR in mice
A total of 32 7-day-old C57BL/6 mice (3.5–3.8 g)
with nursing mothers (SIPPR-BK Experimental Animal Co., Ltd.,
Shanghai, China) were housed under alternate dark-light cycles of
12 h at room temperature with 50% humidity and free access to food
and water. The sex of mice was not considered a factor for the
successful establishment of the OIR model. All surgery was
performed under sodium pentobarbital anesthesia, and all efforts
were made to minimize the suffering of the animals. The OIR model
was used to assess retinal neovascularization, as previously
described (15). The newborn mice
were housed in a hyperoxic (75% oxygen) chamber connected to an
oxygen controller from the first postnatal week (P7-12). At P12,
the mice were returned to normoxia. Following exposure to normoxic
air for 5 days (P12-17), the mice were sacrificed using carbon
dioxide inhalation and their eyes were enucleated and fixed in 4%
paraformaldehyde for 1 h at room temperature. The retinas were cut
four times from the edge to the center and then incubated overnight
at 4°C with isolectin B4 in PBS containing 1 mM CaCl2.
Images were captured using an IX81 microscope (Olympus
Corporation). All animal experiments were performed with the
approval of the Institutional Animal Care and Use Committee of The
Second Military Medical University (Shanghai, China).
Immunohistochemistry
The eyes of the mice were enucleated and fixed in 4%
paraformaldehyde for 1 h. Subsequently, the eyeballs were
post-fixed in perfusion solution overnight, followed by
cryoprotection in 30% sucrose. The frozen sections of retina were
cut to a thickness of 12 µm and mounted on glass slides. The
retinal sections were blocked with 5% bovine serum albumin (BSA)
for 2 h at room temperature. Subsequently, the samples were
incubated with CDK1 and CD31 antibodies and diluted at a ratio of
1:200 in 2% BSA overnight at 4°C. The sections were rinsed with PBS
three times and incubated with secondary antibodies diluted at a
ratio of 1:200 in PBS for 2 h at room temperature. The sections
were subsequently stained with DAPI, washed and mounted with
anti-fading medium. Images were obtained using an IX81 microscope
(Olympus Corporation).
Cell culture
Human umbilical endothelial cells (HUVECs), obtained
from ScienCell Research Laboratories (Carlsbad, CA, USA), were
cultured in DMEM supplemented with 10% FBS. The cells that were
authenticated by cell short tandem repeat genotyping were cultured
at 37°C, in a 5% CO2 cell incubator. Cells were
identified via cell short tandem repeats (STRs) (16). When the cells were grown to ~80%
confluence, they were used for the different assays.
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
TRIzol reagent was used to extract the total RNA
from mouse retinal tissues and from HUVECs. Subsequently,
PrimeScript RT reagent was used to reverse transcribe total RNA to
cDNA. SYBR Premix Ex Taq was subsequently used to assess the
quantity of cDNA. The reactions were performed using a Rotor Gene
3000A (Corbett Research) instrument. The thermocycling conditions
for PCR were as follows: 95°C for 2 min, followed by 40 cycles at
95°C for 10 sec, 55°C for 30 sec and 72°C for 30 sec (17). Each RNA sample was evaluated in
triplicate. mRNA levels were subsequently calculated using the
2−ΔΔCq method (18) and
normalized to the expression of GAPDH. The primer sequences (5′→3′)
used were as follows: TGAGGTAGTAACACTCTGGTA (forward) and
ATGCTAGGCTTCCTGGTT (reverse) for CDK1 (Homo), TGGGCTACACTGAGCACCAG
(forward) and AAGTGGTCGTTGAGGGCAAT (reverse) for GAPDH (Homo),
ACTCCAGGCTGTATCTCAT (forward) and CACTCGTATCGGTATTCCAA (reverse)
for CDK1 (Mus), CAATGAATAGGGCTACAGCA (forward) and
AGGGAGATGCTCAGTGTTGG (reverse) for GAPDH (Mus).
RNA interference
The siRNAs were designed and synthesized by a
commercial company (Shanghai GenePharma Co., Ltd., Shanghai,
China). The target sequences against CDK1-1 were as follows: Sense,
5′-CCUAUGGAGUUGUGUAUAATT-3′ and antisense,
5′-UUAUACACAACUCCAUAGGTT-3′. The target sequences for CDK1-2 were
as follows: Sense, 5′-GGAUGUGCUUAUGCAGGAUTT-3′ and antisense,
5′-AUCCUGCAUAAGCACAUCCTT-3′. The sequences of scramble siRNA were
as follows: Sense, 5′-UUCUCCGAACGUGUCACGUdTdT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAAdTdT. Lipofectamine® 2000 was used
to transfect siRNAs according to the manufacturer's instructions.
Cells were seeded in 24-well plates at a density of
5×103 and transfected with 100 ng/µl siRNA. Samples were
then incubated for 72 h at 37°C with 5% CO2. After 72 h,
the cells were used for subsequent experimentation.
Cell proliferation assay
Cell proliferation was assessed using
5-ethynyl-2′-deoxyuridine (EdU). Following transfection of the
cells with CDK1 siRNAs, they were incubated for 72 h at 37°C and
their medium was removed. EdU (100 µM) was added to the cells and
they were incubated for 2 h at 37°C. Staining was achieved using a
previously described method (17). A
total of 4,000 cells, in a total volume of 100 µl of Endothelial
Cell medium in each well, were incubated overnight at 37°C with 5%
CO2. The grown medium was then replaced with medium
containing CDK1 siRNAs and incubated for 72 h at 37°C. The medium
was then discarded and 100 µl fresh medium containing EdU (100 mM)
was added. Following incubation for 2 h at 37°C, the cells were
stained according to the following protocol: EdU medium was
discarded and the cells were fixed with 4% paraformaldehyde for 30
min at room temperature. The cells were then washed with glycine
for 5 min, followed by two washes with 0.2% Triton X-100 (10 min
each time). The cells were then stained with Apollo fluorescent
azide for 30 min at room temperature, followed by three washes in
0.2% Triton X-100. Furthermore, the cells were stained with Hoechst
for 30 min at room temperature and washed with PBS three times, and
100 µl PBS was added for further analysis. Images were captured and
visualized using an IX81 microscope (Olympus Corporation).
Cell migration assay
A Transwell chamber containing a polycarbonate
filter with an 8-µm pore size was used to evaluate cell migratory
ability. A total of 2×104 cells that were treated with
CDK1 siRNAs for 72 h were seeded in the upper chamber in 0.5% FBS
medium. A total of 700 µl of DMEM containing 1% FBS was added to
the 24-well plate. The cells were allowed to migrate for 12 h, and
the chambers were then fixed with 4% paraformaldehyde for 20 min.
Subsequently, a cotton swab was used to remove the cells from the
upper surface. The chambers were subsequently stained with 0.1%
crystal violet and the migrated cells were counted and analyzed
using an IX81 microscope (Olympus Corporation).
Tube formation assay
The 96-well plate was coated with Matrigel and
incubated at 37°C for 30 min. The cells were treated with CDK1
siRNAs for 72 h and resuspended in fresh medium. A total of
1.5×104 cells were mixed with 100 µl of DMEM and added
to the Matrigel-coated plates. Following 4 h of incubation at 37°C,
images of the capillary-like structures were captured and analyzed
using an IX81 microscope (Olympus Corporation).
Cell cycle assay
The HUVECs were seeded in a 6-well plate and starved
for 16 h. Subsequently, the cells were treated with CDK1 siRNAs for
an additional 72 h, harvested and finally fixed in 70% ethanol at
4°C overnight. The ethanol was removed, and the fixed cells were
washed twice with PBS. Propidium iodide was used to stain the cells
for 30 min at room temperature. The stained cells were assessed
using Cell Lab Quanta SC (Beckman Coulter, Inc.). The data were
finally analyzed using Modfit software (MFLT5.0; Verity Software
House, Inc.).
Cell apoptosis assay
The HUVECs were seeded in a 6-well plate and starved
for 16 h. Subsequently, the cells were treated with CDK1 siRNAs for
an additional 72 h and were then harvested and stained with an
Annexin V-FITC apoptosis kit, according to the manufacturer's
protocol. The stained cells were assessed using Cell Lab Quanta SC
(Beckman Coulter, Inc.). The data were finally analyzed using
FlowJo software (version 7.6.1; FlowJo LLC).
Western blot analysis
Lysis buffer containing protease inhibitor cocktail
was used to extract total protein from mouse retinas and HUVECs.
The protein concentration of each sample was determined using a
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology). Approximately 10 µg of protein was separated in 10%
SDS/PAGE gels and transferred onto polyvinylidene fluoride
membranes. Nonfat milk was used at 5% to block the non-specific
sites. Subsequently, the samples were incubated with GAPDH, CD31,
cyclin A1, cyclin B1, cyclin D1, cyclin E1, CDK1, CDK2, PARP1, p21
and p53 primary antibodies diluted at a ratio of 1:1,000 in
Universal antibody diluent at 4°C overnight, following which the
membranes were washed three times with TBS-T and incubated with
HRP-conjugated secondary antibodies for 2 h at room temperature.
GAPDH was used as a loading control and the protein signals were
finally detected using Super Signal West Pico chemiluminescent
substrate and semi-quantified using the GeneGnome HR Image Capture
system (Syngene Europe).
Statistical analysis
The data were analyzed and presented using Prism
software (GraphPad 5.0; GraphPad Software, Inc.). All data are
presented as the mean ± SD and the differences between the groups
were evaluated using the unpaired t-test or one-way ANOVA.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CDK1 is expressed at a high level in
pathological retinas
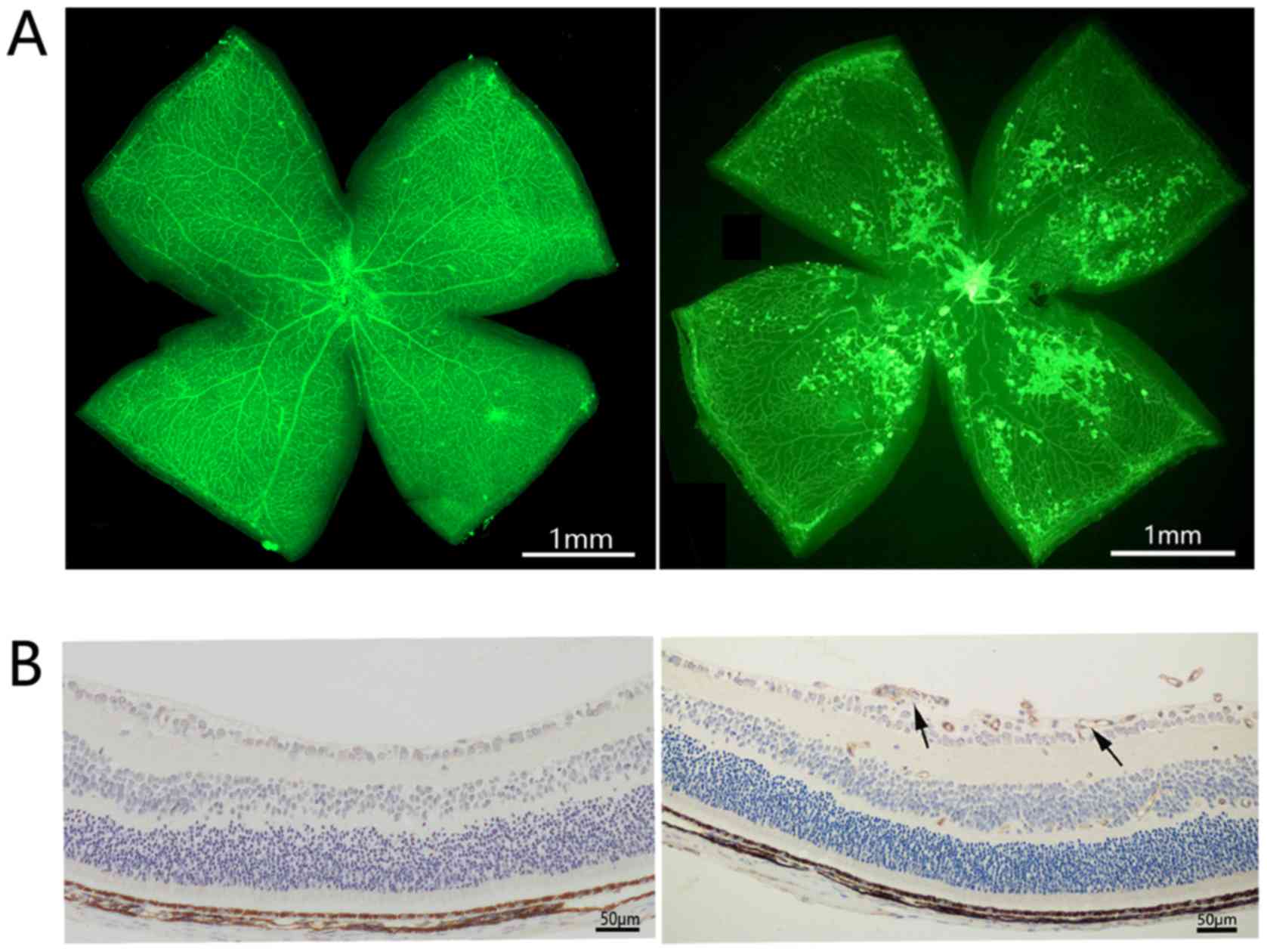
The OIR mouse model is a well-recognized model for
the evaluation of retinal angiogenesis. Newborn mice were exposed
to hyperoxia between P7 and P12. On P12, the mice were grown under
normoxic conditions. On P17, retinal neovascularization was
determined in OIR mice by isolectin-B4 staining and
immunohistochemical staining of CD31. In order to investigate the
role of CDK1 in retinal angiogenesis, the expression of CDK1 was
initially examined in OIR mouse retinas using immunofluorescence
staining, RT-qPCR analysis and western blotting. OIR mice exhibited
high retinal neovascularization and the immunohistochemical data
indicated that CD31 was overexpressed in the retinas of OIR mice
(Fig. 1A and B). Immunofluorescence
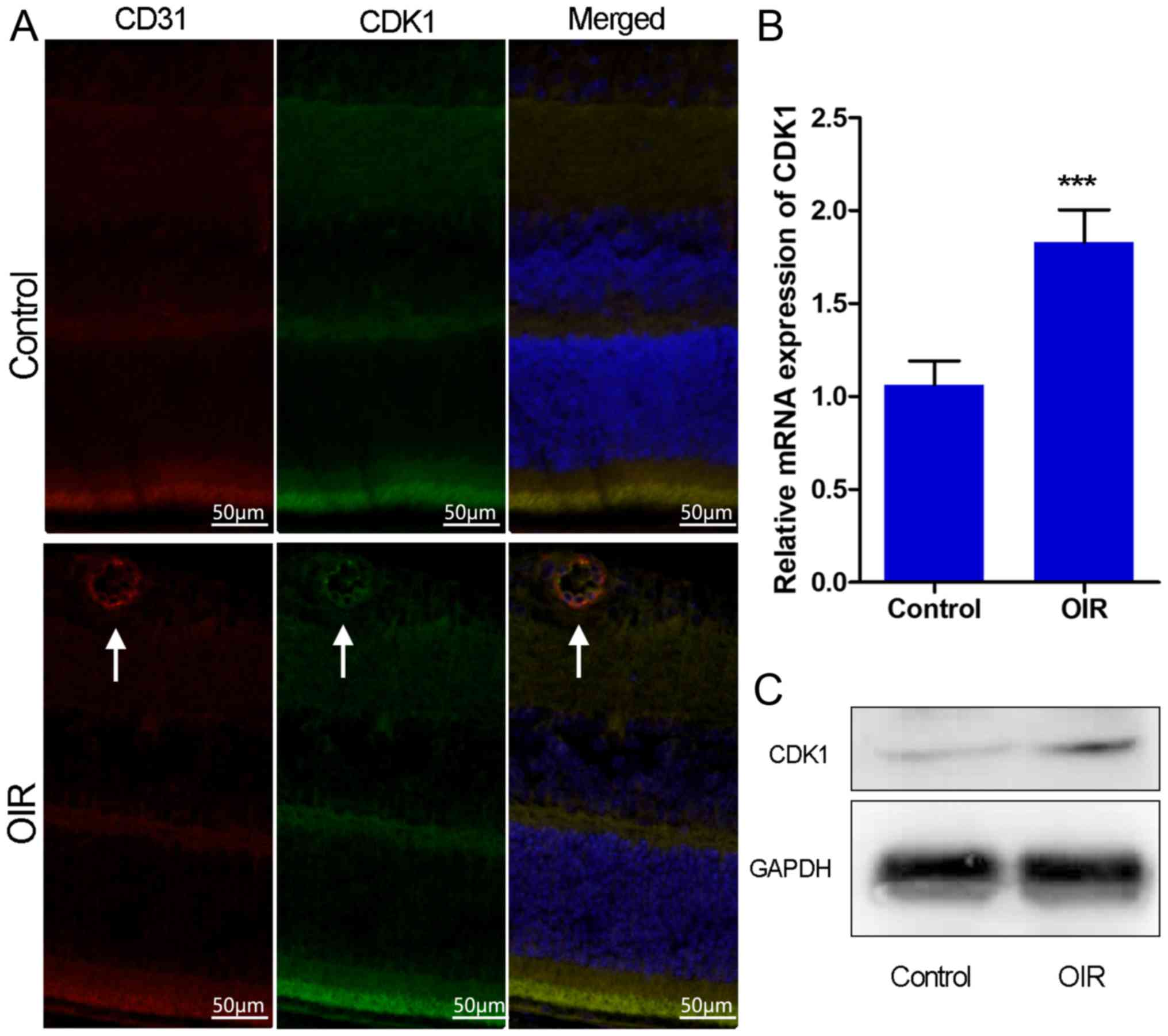
microscopy demonstrated that the overexpression of CDK1 colocalized
with neovascularization in the retinas of OIR mice (Fig. 2A). RT-qPCR and western blot assays
demonstrated that the expression levels of CDK1 were significantly
higher in the OIR mouse retinas compared with those in the control
group (Fig. 2B and C). The data
indicated that CDK1 was overexpressed in pathological retinas and,
notably, in pathological retinal angiogenesis.
Silencing of CDK1 inhibits HUVEC
growth
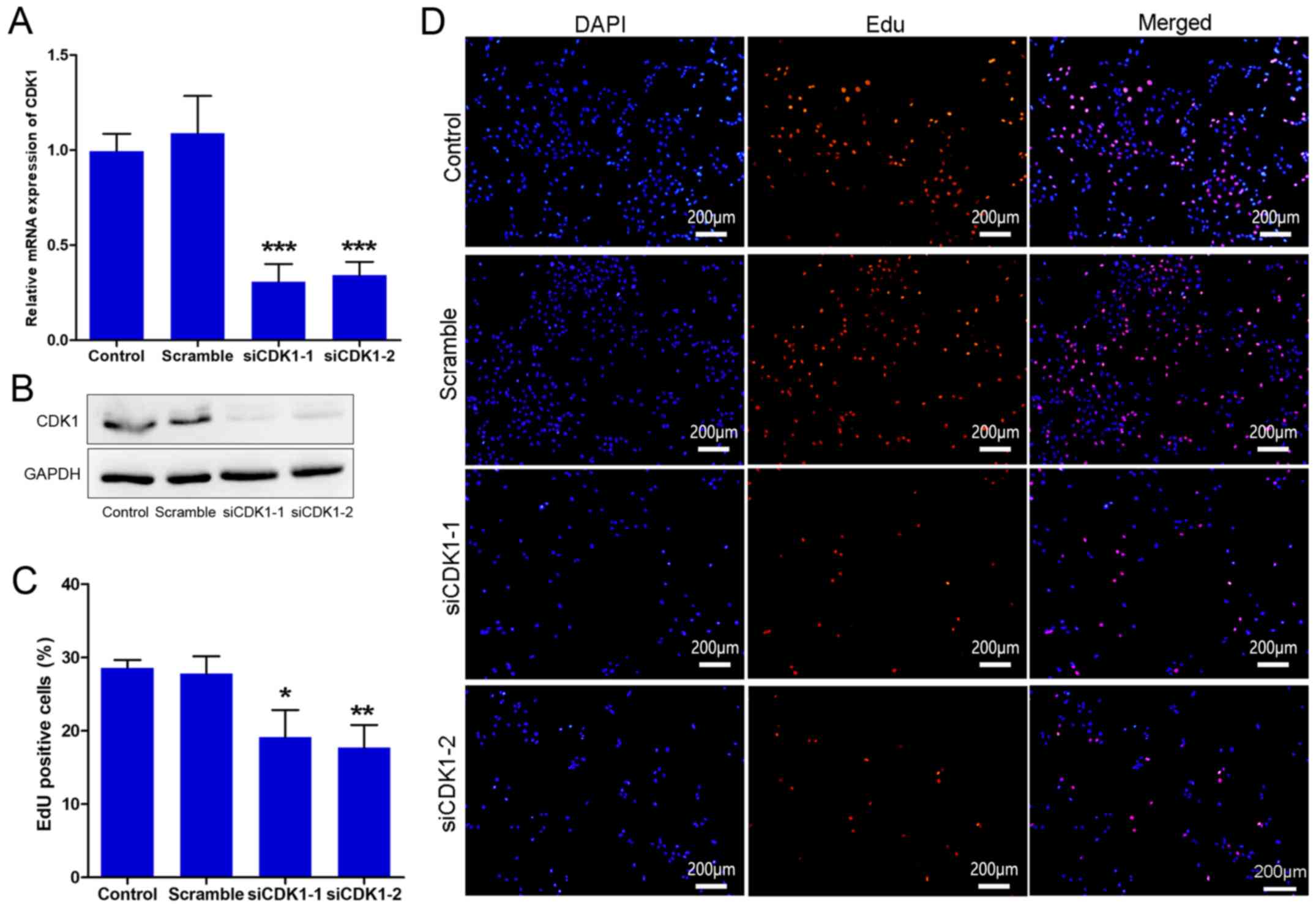
To further investigate the role of CDK1 in
angiogenesis, CDK1 siRNAs were synthesized and their knockdown
efficiency was assessed. The results indicated that the CDK1 siRNAs
significantly inhibited the expression of CDK1 at the mRNA and
protein levels (Fig. 3A and B). The
effect of CDK1 siRNAs on cell proliferation was subsequently
investigated by EdU. The data indicated that the cells treated with
CDK1 siRNAs proliferated more slowly than those of the control and
scramble groups (Fig. 3C and D). The
data indicated that the transfection was efficient and that the
knockdown of CDK1 may exert anti-angiogenic activity, partially
through inhibition of the growth of vascular endothelial cells.
Silencing of CDK1 inhibits the
migration of HUVECs
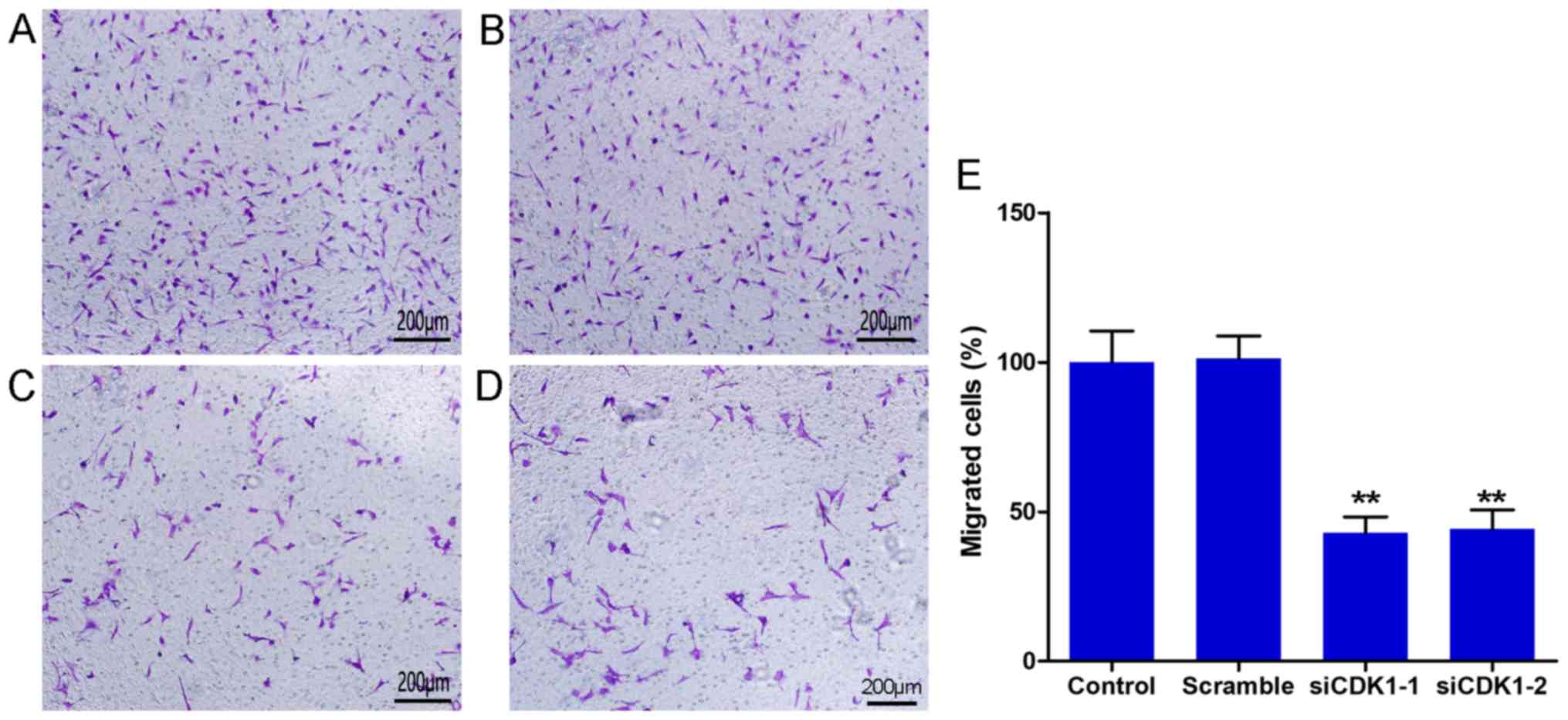
The Transwell chamber assay was used to determine
cell migration. The results indicated that CDK1 siRNAs inhibited
the number of migrated cells significantly compared with that in
the control and the scramble groups (Fig. 4A-D). No differences were observed
between the control and the scramble groups (Fig. 4E). The number of migrated cells in
the siCDK1-1 group was decreased to 57.1% compared with that of the
control group, whereas the number of migrated cells in the siCDK1-2
group was decreased to 55.7% compared with that of the control
group (Fig. 4E). The data suggested
that silencing of CDK1 inhibited vascular endothelial cell
migration, which is considered a vital process for
angiogenesis.
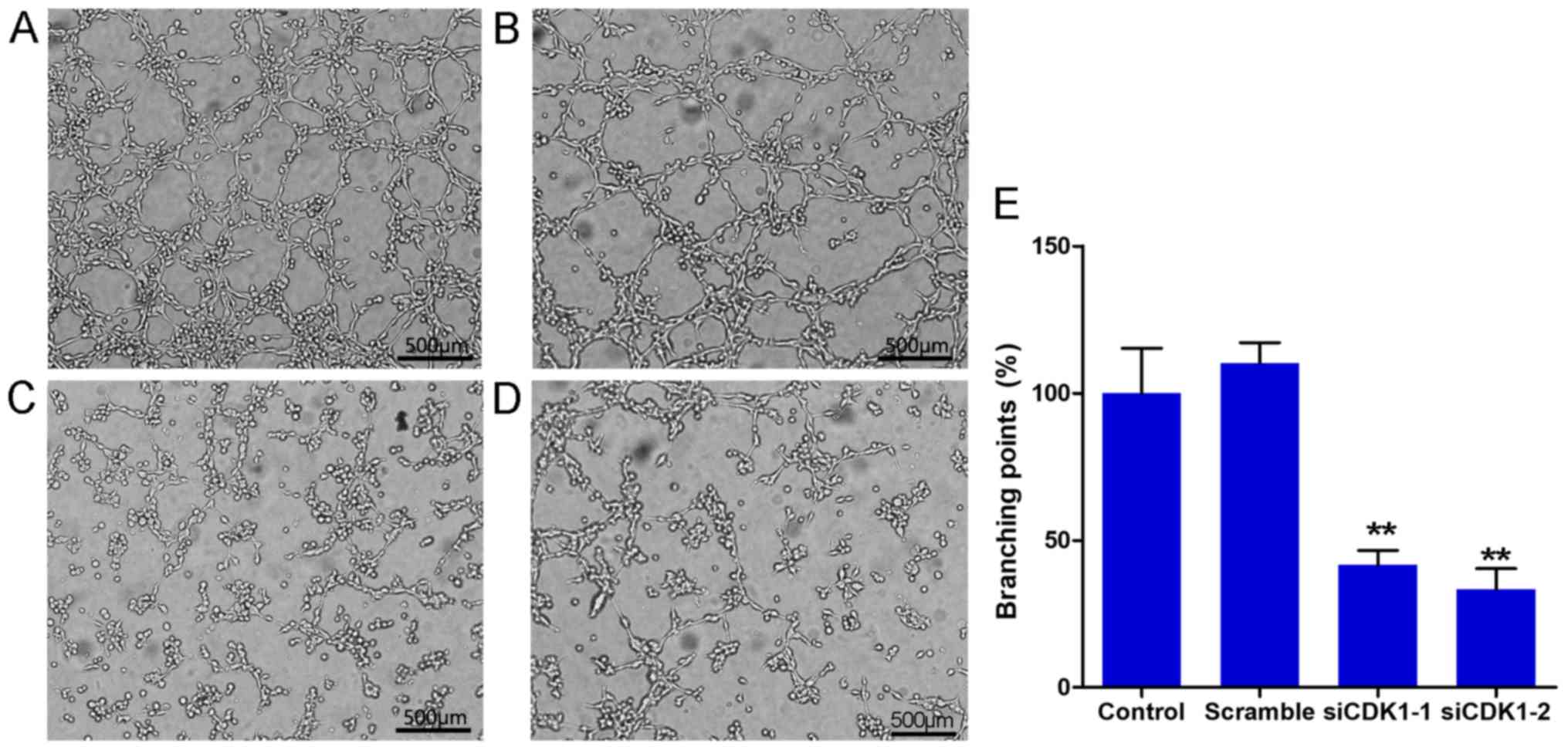
Silencing of CDK1 inhibits the tube
formation of HUVECs
The tube formation assay is a well-known method to
evaluate the angiogenic activity of endothelial cells in
vitro. In the present study, tube formation assays were
performed using Matrigel. The results indicated that CDK1 siRNAs
significantly inhibited tube formation of HUVECs compared with that
noted in the control and scramble groups (Fig. 5A-D). No significant difference was
noted with regard to tube formation between the control and
scramble groups (Fig. 5E). The
percentage of branching points in the siCDK1-1 group decreased to
58.3% compared with that of the control group, and the percentage
of migrated cells in the siCDK1-2 group decreased to 66.7% compared
with that of the control group (Fig.
5E). The data suggested that CDK1 served an important role in
angiogenesis, and that knockdown of CDK1 inhibited angiogenesis
in vitro.
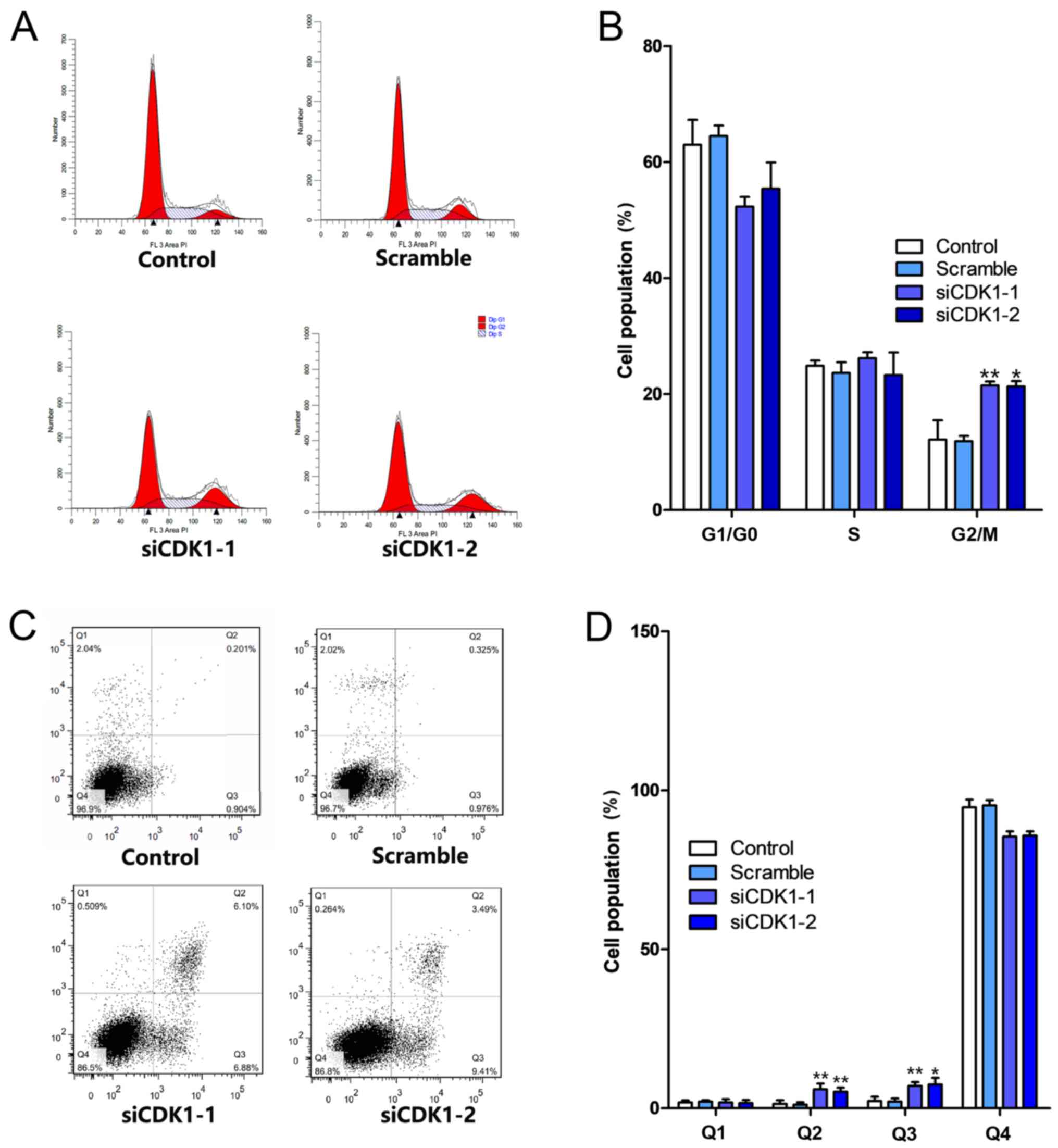
Silencing of CDK1 inhibits the cell
cycle and apoptosis
The effects of CDK1 knockdown on cell cycle
progression and the induction of apoptosis were investigated. The
knockdown of CDK1 induced cell cycle arrest at the G2/M phase
(Fig. 6A). The percentage of the
cell population in the G2/M phase increased by ~10% in both the
siCDK1-1 and siCDK1-2 groups compared with that noted in the
control group (Fig. 6B).
Furthermore, the knockdown of CDK1 induced apoptosis compared with
that in the control and scramble groups (Fig. 6C). The percentages of early and late
apoptotic cells in the CDK1 knockdown groups increased by ~9%
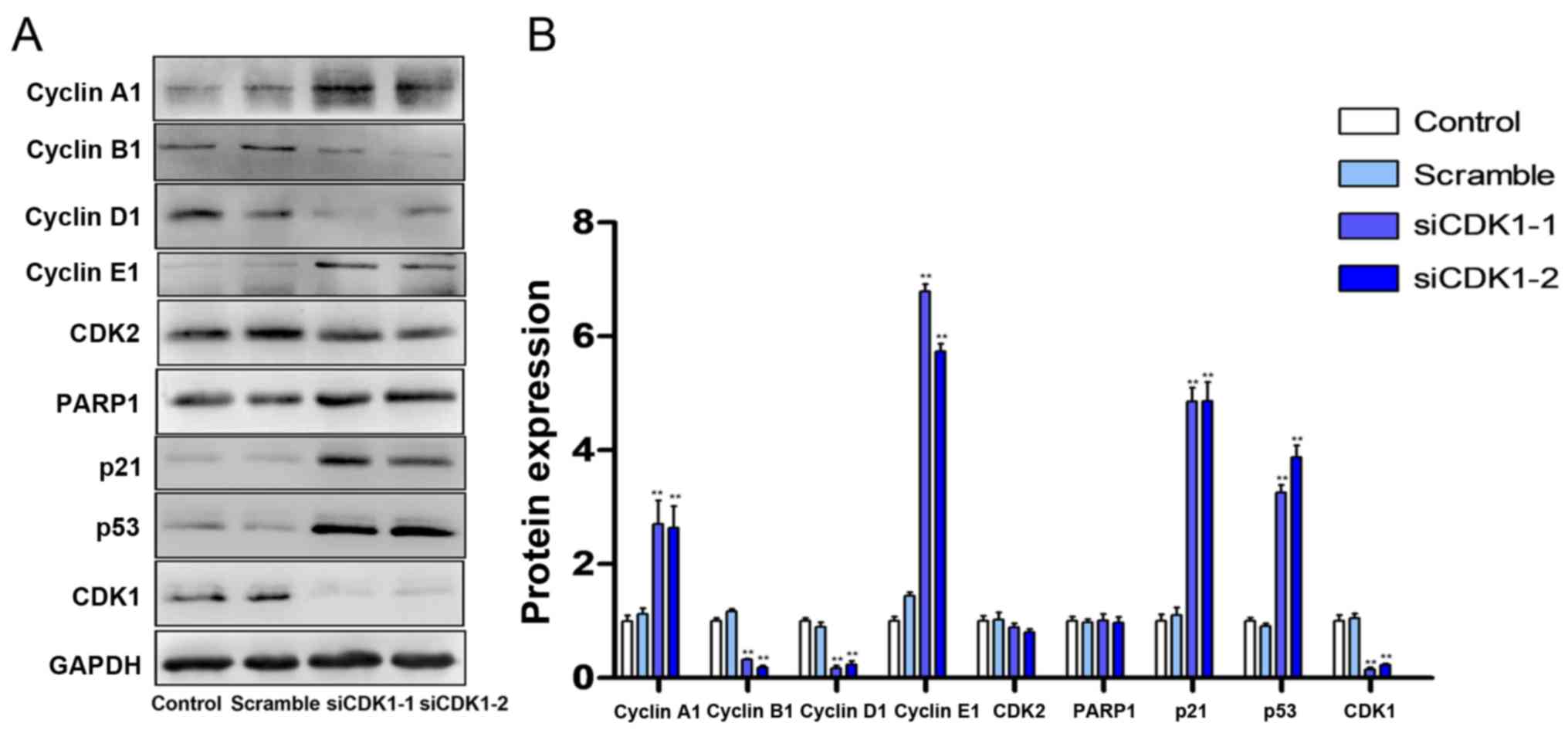
compared with those of the control group (Fig. 6D). Subsequently, the expression
levels of cell cycle- and apoptosis-related proteins were
investigated using western blot analysis. The results indicated
that silencing of CDK1 inhibited the expression levels of cyclin B,
cyclin D and CDK2, and increased the expression levels of cyclin A,
cyclin E, PARP1, p21 and p53 (Fig. 7A
and B).
Discussion
Previous studies have revealed that CDK1 was
expressed at a high level in several types of tumor tissues and
that it served an important role in the incidence and development
of tumors (12,14,19).
However, the role of CDK1 in angiogenesis has not been fully
investigated. The formation of pathological angiogenesis resembles
the formation of tumor angiogenesis, and both of these phenomena
are characterized by the uncontrolled growth of multiple cells
(20). In addition, tumor growth is
usually accompanied by angiogenesis, which provides oxygen and
nutrients to the tumor tissue (20).
It has been reported that CDK1 can stabilize hypoxia-inducible
factor-1α (HIF-1α) in order to promote tumor angiogenesis and that
HIF-1α is an important molecule that initiates the angiogenic
cascade (14). These findings
suggest that CDK1 may be involved in the formation of pathological
angiogenesis.
Pathological retinal angiogenesis is a severe
disorder, which can result in retinal detachment, optic neuropathy
and glaucoma (21,22). These diseases can cause vision loss
and even blindness. The vascular endothelial growth factor (VEGF)
antibody has been used clinically as an inhibitor of retinal
angiogenesis. However, subsequent studies have demonstrated that
VEGF may cause cardiovascular adverse effects and that certain
patients can develop drug resistance (23). Therefore, the identification of novel
therapeutic targets is required to treat retinopathy in combination
with the VEGF antibody and/or independently. In the present study,
it was demonstrated for the first time, to the best of our
knowledge, that CDK1 was overexpressed in pathological retinal
tissues. The overexpression of CDK1 occurred concomitantly with
neovascularization, as demonstrated by the colocalization of
protein expression. This finding indicated that CDK1 served an
important role in the formation of pathological angiogenesis. The
cell proliferative, migratory and tube formation assays further
suggested that targeting of CDK1 may be an important strategy to
improve retinal neovascularization.
Although CDK1 is known to regulate the cell cycle
process, it serves a multitude of roles in different tumor
microenvironments. The silencing of CDK1 induced HCC827 cell cycle
arrest at the G1 phase, whereas the silencing of CDK1 induced HeLa
cell cycle arrest at the G2/M phase (12,24).
These two cell lines are of human non-small cell lung cancer and
human cervical cancer origin, respectively. The present study
indicated that silencing of CDK1 induced endothelial cell cycle
arrest at the G2/M phase. The CDK1-cyclin B1 complex, also known as
M-phase-promoting factor is particularly relevant with regard to
mitotic progression (25–27). The data obtained in the present study
indicated that the expression of cyclin B1 was inhibited in the
CDK1 siRNA group. In addition, the expression levels of the cell
cycle related genes, namely cyclin A1, cyclin D1, cyclin E1 and
CDK2, were significantly altered. These may partially account for
the cell cycle arrest induced by CDK1 siRNAs. Apoptosis is an
additional biological process that requires the involvement of CDK1
(28,29). The results of the present study
indicated that the silencing of CDK1 induced apoptosis and
increased the expression of PARP1. These data were consistent with
previous findings demonstrating that CDK1 protected mitotic cells
against the induction of apoptosis (8). Furthermore, the present study showed
that the silencing of CDK1 increased the expression levels of p53
and p21. p53 and p21 are two key genes that regulate the cell cycle
and the apoptotic process (30–32).
These data indicate that the silencing of CDK1 may activate p21 and
p53 in order to induce endothelial cell cycle arrest and
apoptosis.
In the present study, the expression of CDK1 in OIR,
its role in angiogenesis in vitro and the possible mechanism
underlying this process were investigated for the first time. The
data indicated that the expression of CDK1 was high in pathological
retinal angiogenesis and that CDK1 siRNAs inhibited endothelial
cell proliferation, migration and tube formation. Furthermore, CDK1
siRNAs induced cell cycle arrest and apoptosis. The possible
mechanism was associated with inhibition of the expression of cell
cycle- and apoptosis-related proteins mediated by increasing the
protein expression levels of the p21 and p53. Collectively, the
results highlight the role of CDK1 in retinal angiogenesis and
suggest its potential application for pathological
neovascularization therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant. no. 81700839), the
Shanghai Municipal Planning Commission of Science and Research Fund
(grant. no. 20170054), the Research Foundation for Youth of Second
Military Medical University (grant. no. 2016QN13) and the Shanghai
Pujiang Program (grant. no. 17PJD041).
Availability of data and materials
All data generated or analyzed during the current
study are included in this published article.
Authors' contributions
XG, YZ and RZ conceived and designed the
experiments, and wrote the manuscript. HZ, ZZ and JW acquired,
analyzed and interpreted the data. WS and MZ conceived the current
study and revised the manuscript for intellectual content. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed with the
approval of the Institutional Animal Care and Use Committee of The
Second Military Medical University (Shanghai, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Potente M and Carmeliet P: The link
between angiogenesis and endothelial metabolism. Annu Rev Physiol.
79:43–66. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Usui Y, Westenskow PD, Murinello S,
Dorrell MI, Scheppke L, Bucher F, Sakimoto S, Paris LP, Aguilar E
and Friedlander M: Angiogenesis and eye disease. Annu Rev Vis Sci.
1:155–184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rubio RG and Adamis AP: Ocular
angiogenesis: Vascular endothelial growth factor and other factors.
Dev Ophthalmol. 55:28–37. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tah V, Orlans HO, Hyer J, Casswell E, Din
N, Sri Shanmuganathan V, Ramskold L and Pasu S: Anti-VEGF therapy
and the retina: An update. J Ophthalmol. 2015:6276742015.PubMed/NCBI
|
|
5
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Asghar U, Witkiewicz AK, Turner NC and
Knudsen ES: The history and future of targeting cyclin-dependent
kinases in cancer therapy. Nat Rev Drug Discov. 14:130–146. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Allan LA and Clarke PR: Phosphorylation of
caspase-9 by CDK1/cyclin B1 protects mitotic cells against
apoptosis. Mol Cell. 26:301–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matthess Y, Raab M, Knecht R, Becker S and
Strebhardt K: Sequential Cdk1 and Plk1 phosphorylation of caspase-8
triggers apoptotic cell death during mitosis. Mol Oncol. 8:596–608.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pietenpol JA and Stewart ZA: Cell cycle
checkpoint signaling: Cell cycle arrest versus apoptosis.
Toxicology. 181:475–481. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeng Y, Stauffer S, Zhou J, Chen X, Chen Y
and Dong J: Cyclin-dependent kinase 1 (CDK1)-mediated mitotic
phosphorylation of the transcriptional co-repressor Vgll4 inhibits
its tumor-suppressing activity. J Biol Chem. 292:15028–15038. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pu S, Zhao Y, Zhou G, Zhu H, Gong L, Zhang
W, Huang G, Wang D and Liu D: Effect of CDK1 shRNA on
proliferation, migration, cell cycle and apoptosis in non-small
cell lung cancer: Effect of CDK1 shRNA on NSCLC cells. J Cell
Physiol. 233:75142017.
|
|
13
|
Manchado E, Guillamot M, de Cárcer G,
Eguren M, Trickey M, García-Higuera I, Moreno S, Yamano H, Cañamero
M and Malumbres M: Targeting mitotic exit leads to tumor regression
in vivo: Modulation by Cdk1, Mastl, and the PP2A/B55α, δ
phosphatase. Cancer Cell. 18:641–654. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Warfel NA, Dolloff NG, Dicker DT, Malysz J
and El-Deiry WS: CDK1 stabilizes HIF-1α via direct phosphorylation
of Ser668 to promote tumor growth. Cell Cycle. 12:3689–3701. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Connor KM, Krah NM, Dennison RJ, Aderman
CM, Chen J, Guerin KI, Sapieha P, Stahl A, Willett KL and Smith LE:
Quantification of oxygen-induced retinopathy in the mouse: A model
of vessel loss, vessel regrowth and pathological angiogenesis. Nat
Protoc. 4:1565–1573. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo T, Song H, Zhao Z, Qi Z and Zhao S:
Overexpression of Annexin A2 Receptor inhibits neovascularization
via the promotion of Krüppel-Like transcription factor 2. Cell
Physiol Biochem. 46:1617–1627. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song H, Wang W, Zhao P, Qi Z and Zhao S:
Cuprous oxide nanoparticles inhibit angiogenesis via down
regulation of VEGFR2 expression. Nanoscale. 6:3206–3216. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sung WW, Lin YM, Wu PR, Yen HH, Lai HW, Su
TC, Huang RH, Wen CK, Chen CY, Chen CJ and Yeh KT: High
nuclear/cytoplasmic ratio of Cdk1 expression predicts poor
prognosis in colorectal cancer patients. BMC Cancer. 14:9512014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ronca R, Benkheil M, Mitola S, Struyf S
and Liekens S: Tumor angiogenesis revisited: Regulators and
clinical implications. Med Res Rev. 37:1231–1274. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Campochiaro PA: Molecular pathogenesis of
retinal and choroidal vascular diseases. Prog Retin Eye Res.
49:67–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Puro DG, Kohmoto R, Fujita Y, Gardner TW
and Padovani-Claudio DA: Bioelectric impact of pathological
angiogenesis on vascular function. Proc Natl Acad Sci USA.
113:9934–9939. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Simó R and Hernandez C: Intravitreous
anti-VEGF for diabetic retinopathy: Hopes and fears for a new
therapeutic strategy. Diabetologia. 51:15742008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiao H, Tian M, Ge J, Wei X, Li Z, Li X,
Tao D, Hu J and Gong J: The role of CDK1 siRNA interference in cell
cycle and cell apoptosis. Front Med China. 3:3842009. View Article : Google Scholar
|
|
25
|
Rattani A, Vinod PK, Godwin J,
Tachibana-Konwalski K, Wolna M, Malumbres M, Novák B and Nasmyth K:
Dependency of the spindle assembly checkpoint on Cdk1 renders the
anaphase transition irreversible. Curr Biol. 24:630–637. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jang SH, Kim AR, Park NH, Park JW and Han
IS: DRG2 regulates G2/M progression via the cyclin B1-Cdk1 complex.
Mol Cells. 39:699–704. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z, Fan M, Candas D, Zhang TQ, Qin L,
Eldridge A, Wachsmann-Hogiu S, Ahmed KM, Chromy BA, Nantajit D, et
al: Cyclin B1/Cdk1 coordinates mitochondrial respiration for
cell-cycle G2/M progression. Dev Cell. 29:217–232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang P, Kawakami H, Liu W, Zeng X,
Strebhardt K, Tao K, Huang S and Sinicrope FA: Targeting CDK1 and
MEK/ERK overcomes apoptotic resistance in BRAF-mutant human
colorectal cancer. Mol Cancer Res. 16:378–389. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang R, Shi H, Ren F, Zhang M, Ji P, Wang
W and Liu C: The aberrant upstream pathway regulations of CDK1
protein were implicated in the proliferation and apoptosis of
ovarian cancer cells. J Ovarian Res. 10:602017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Karimian A, Ahmadi Y and Yousefi B:
Multiple functions of p21 in cell cycle, apoptosis and
transcriptional regulation after DNA damage. DNA Repair (Amst).
42:63–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qi LW, Zhang Z, Zhang CF, Anderson S, Liu
Q, Yuan CS and Wang CZ: Anti-colon cancer effects of 6-shogaol
through G2/M cell cycle arrest by p53/p21-cdc2/cdc25A crosstalk. Am
J Chin Med. 43:743–756. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen J: The cell-cycle arrest and
apoptotic functions of p53 in tumor initiation and progression.
Cold Spring Harb Perspect Med. 6:a0261042016. View Article : Google Scholar : PubMed/NCBI
|