Introduction
Osteoporosis (OP) is the most common type of bone
disorder (1). OP is a public health
issue with gradually increasing incidence (2). In China, the incidence of OP was 14.94%
in 2008, and reached 27.96% between 2012 and 2015 (3,4). In
Europe, ~27.5 million people were affected by OP in 2010 (5). As a bone disease, the main
characteristics of OP include low bone density, easy bone damage,
decreasing bone mass and increased risk of fracture (6). Hip fractures are frequently observed in
patients with OP (7). Diagnosis of
OP is very difficult until a fracture occurs (8). With an increasing number of elderly
people in China, it is important to investigate methods for the
prevention and treatment of osteoporosis. Elderly patients can
suffer from motor dysfunction and impaired body functions, which
can have adverse effects on mental health. Such mental health
issues can lead to complications with the treatment and care of
patients with OP. Therefore, elderly patients with OP require
nursing interventions to aid their physical and mental health in
order to promote quick recovery and improve their life quality
(9). There is an urgent need to
understand the pathogenesis and molecular mechanisms of OP to
identify potential biomarkers and therapeutic targets.
MicroRNAs (miRNAs) are small non-coding regulatory
RNAs (~23 nucleotides in length) that play a crucial role in the
post-transcriptional regulation of gene expression by suppressing
target mRNA translation by binding to its 3′untranslated region
(UTR) region (10–12). Therefore, miRNAs can regulate
multiple mRNAs to create a complex regulatory network of signaling
pathways including oncogene pathways and tumor suppressor pathways,
such as the Wnt and PI3K pathways (13–16).
Altering the expression of these miRNAs alters the function of
their target genes, some of which regulate crucial cellular
mechanisms such as growth, proliferation, migration, invasion and
metastasis (17–19). Previous studies have reported that
changes in circulating miRNAs are associated with osteoporotic
fractures. Seeliger et al (20) reported that five miRNAs were
upregulated both in serum and bone tissue of patients with OP. Long
non-coding RNA maternally expressed 3 (MEG3) inhibited osteogenic
differentiation by decreasing miR-133a-3p expression, and the
expression of MEG3 was found to be upregulated in bone marrow stem
cells in ovariectomized mice and in patients with OP (21). An increasing number of miRNAs have
been shown to play an important role in osteoblastogenesis and
osteoporosis (22–24). However, the function of miR-135b-5p
in osteoporosis remains unclear.
Therefore, the aims of the present study were to
determine the expression of miR-135b-5p in bone tissue fragments of
patients with OP, to investigate the role of miR-135b-5p in
osteogenic differentiation and osteoblast growth, and to examine
the underlying mechanism of miR-135b-5p.
Materials and methods
Cell culture and tissue samples
The mouse preosteoblast cell line MC3T3-E1 was
purchased from American Type Culture Collection (cat. no.
CRL-2594). The cells were cultured for 24 h in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in a humidified incubator with 5%
CO2.
For osteogenic differentiation, MC3T3-E1 cells were
cultured at 37°C in osteogenic induction medium (25) containing 10% FBS, 100 nM
dexamethasone, 5 mM L-glycerophosphate and 50 mg/ml ascorbic acid
(Sigma-Aldrich; Merck KGaA) for 14 days.
Bone fragments (bone tissues, 100 mg) were collected
from 30 patients with OP [female, 15; male, 15; age, 52–73 years;
body mass index (BMI), 23.6±5.4 kg/m2; lumbar spine bone
mineral density, 0.79±0.10 g/cm2; femoral neck bone
mineral density, 0.62±0.10 g/cm2; total hip bone mineral
density, 0.65±0.07 g/cm2] and 30 patients with
osteoarthritis (control; female, 15; male, 15; 51–72 years; BMI,
25.1±3.7 kg/m2; lumbar spine bone mineral density,
1.00±0.15 g/cm2; femoral neck bone mineral density,
0.85±0.13 g/cm2; total hip bone mineral density,
0.87±0.14 g/cm2) at the Third Affiliated Hospital of Sun
Yat-sen University, between May 2015 and May 2017. Bone fragments
extracted from the transcervical region of the femoral neck, were
dissected into smaller fragments, washed three times in PBS and
stored at −80°C until further use. The present study was approved
by the Ethics Committee of the Third Affiliated Hospital of Sun
Yat-sen University and every patient provided written informed
consent.
Cell transfection
MC3T3-E1 cells were seeded in 6-well plates at a
density of 1×106 cells/well and cultured at 37°C for 24
h. Then, 100 nM mimic control (5′-UAUAUCGUGUUAUUAGCGUUCCU-3′;
Shanghai GenePharma Co., Ltd.), 100 nM miR-135b-5p mimic
(5′-UAUGGCUUUUCAUUCCUAUGUGA-3′; Shanghai GenePharma Co., Ltd.), 2
µl runt-related transcription factor 2 (RUNX2)-plasmid (cat. no.
sc-400183-ACT; Santa Cruz Biotechnology, Inc.), 2 µl
control-plasmid (cat. no. sc-108083; Santa Cruz Biotechnology,
Inc.) or 100 nM miR-135b-5p mimic + 2 µl RUNX2-plasmid were
transfected into MC3T3-E1 cells using Lipofectamine®
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) as per
the manufacturer's protocol. Cells without any treatment were used
the control group, and 48 h after cell transfection, transfection
efficiency was detected using reverse transcription-quantitative
PCR (RT-qPCR).
Alkaline phosphatase assay
MC3T3-E1 cells were seeded in 96-well plates and
then transfected with mimic control, miR-135b-5p mimic or
miR-135b-5p mimic+RUNX2-plasmid for 48 h. Then, the ALP assay kit
(cat. no. P0321; Beyotime Institute of Biotechnology) was used to
determine the extracellular ALP activity in MC3T3-E1 cells
following the manufacturer's protocol.
RT-qPCR
Total RNA was extracted with the TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) from tissue samples or
cells according to the manufacturer's instructions. Then,
complementary DNA was synthesized from total RNA by using the
TaqMan MicroRNA Reverse Transcription kit (Invitrogen; Thermo
Fisher Scientific, Inc.). The temperature protocol for the reverse
transcription reaction consisted of primer annealing at 25°C for 5
min, cDNA synthesis at 42°C for 60 min and termination at 80°C for
2 min. RT-qPCR assay was performed using SYBR Premix Ex TaqTM II
(TliRNaseH Plus) kit (cat. no. RR820a; Takara Bio, Inc.). The
reaction volume was 20 µl. The thermocycling conditions were as
follows: Initial denaturation at 94°C for 30 sec; 40 cycles of
denaturation at 94°C for 5 sec, annealing at 60°C for 20 sec; and a
final extension at 72°C for 30 sec. GAPDH or U6 were used as an
internal reference. Primer sequences were as following: U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; GAPDH forward,
5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3′; miR-135b-5p forward,
5′-GGTATGGCTTTTCATTCCT-3′ and reverse, 5′-CAGTGCGTGTCGTGGAGT3′;
RUNX2 forward, 5′-GATGATGACACTGCCACCTCT-3′ and reverse,
5′-AGGGCCCAGTTCTGAAGC-3′; ALP forward,
5′-CCGAATTCATGTTGGCCTGTTCAACT-3′ and reverse,
5′-ATGTCGACTTAGTTATTTTCATAATACCAAATTCC-3′; OC forward,
5′-CAGACCTAGCAGACACCATGA-3′ and reverse,
5′-CTGCCAGAGTTTGGCTTTAGG-3′; Osterix forward,
5′-AGAGATCTGAGCTGGGTAG-3′ and reverse, 5′-AAGAGAGCCTGGCAAGAGG-3′.
The relative expression levels of the genes were analyzed using
2−ΔΔCq method (26). All
experiments were performed in triplicate.
Western blot assay
The proteins were extracted from cells or tissues
samples using RIPA lysis buffer (cat. no. P0013E; Beyotime
Institute of Biotechnology). The protein concentration was detected
using the bicinchoninic acid assay kit (cat. no. BCA1-1KT;
Sigma-Aldrich; Merck KGaA). Equal amount of proteins (30 µg per
lane) was separated by 12% SDS-PAGE for 40 min. The protein was
transferred to a PVDF membrane (EMD Millipore). The membrane was
blocked with 5% skimmed milk for 1 h at room temperature. The
membrane was washed three times with 1X PBS-0.1% Tween-20 (PBST).
The membranes were then incubated with primary antibodies: Runx2
(1:1,000; cat. no. 12556; Cell Signaling Technology, Inc.), ALP
(1:1,000; cat. no. sc-365765; Santa Cruz Biotechnology, Inc.), OC
(1:1,000; cat. no. ab93876; Abcam), Osterix (1:1,000; cat. no.
sc-393060; Santa Cruz Biotechnology, Inc.), and β-actin (1:1,000;
cat. no. 4970; Cell Signaling Technology, Inc.) overnight at 4°C.
Subsequently, the protein was incubated with a horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G secondary
antibody (1:1,000; cat. no. 7074; Cell Signaling Technology, Inc.)
overnight at 4°C. Finally, protein blots were visualized and
analyzed using a chemiluminescence system (Beyotime Institute of
Biotechnology). β-actin was used as an internal control.
MTT assay
Cell viability was evaluated using an MTT assay. For
the MTT detection, MC3T3-E1 cells were seeded in 96-well plates and
then transfected with mimic control, miR-135b-5p mimic or
miR-135b-5p mimic+RUNX2-plasmid for 48 h. Subsequently, 20 µl MTT
(5 mg/ml; Sigma-Aldrich; Merck KGaA) was added into each well and
cultured at 37°C for 4 h. To assess cell viability, absorbance was
measured at 490 nm using a microplate spectrophotometer (Bio-Rad
Laboratories, Inc.).
Flow cytometry analysis
Cells were centrifuged (500 × g, 5 min, 4°C), and
re-suspended in 100 µl of FITC-binding buffer. Then, ~5 µl of
ready-to-use Annexin V-FITC and 5 µl propidium iodide (PI) (cat.
no. 70-AP101-100; MultiSciences Biotech Co., Ltd.) were added to
the mixture. Cells were incubated for 30 min in the dark at room
temperature. Annexin V-FITC and PI fluorescence were analyzed using
a BD FACSCalibur flow cytometer (Becton, Dickinson and Company),
Apoptosis in cultured MC3T3-E1 cells was determined by flow
cytometry according to the manufacturer's instructions. Data were
analyzed by Cell Quest software (version 5.1; BD Biosciences).
Luciferase reporter gene assay
TargetScan 7.2 (http://www.targetscan.org/vert_72/) was used to
predict the potential targets of miR-135b-5p, and the binding sites
between RUNX2 and miR-135b-5p were observed. To confirm the binding
sites between RUNX2 and miR-135b-5p, luciferase reporter gene
assays were performed. Luciferase reporter plasmids (psi-CHECK2)
containing the wild-type 3′UTRs of RUNX2 (WT-RUNX2), as well as
mutant 3′UTRs of RUNX2 (MUT-RUNX2) were manufactured by TsingKe
Biotech Co., Ltd. Lipofectamine 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to cotransfect MC3T3-E1 cells with the
wild-type or mutant 3′UTR luciferase reporter plasmids, and the
miR-135b-5p mimic or mimic control respectively. Cells were
harvested after transfection for 48 h, and the luciferase
activities were measured using the Dual-Luciferase®
Reporter Assay Kit (Promega Corporation) following the
manufacturer's instructions. Firefly luciferase activity was
normalized to Renilla luciferase activity.
Statistical analysis
Data are presented as mean ± SD. SPSS 22.0 software
(IBM Corp.) was used for statistical analysis. Differences between
groups were determined using a one-way ANOVA with Tukey's post hoc
test or Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
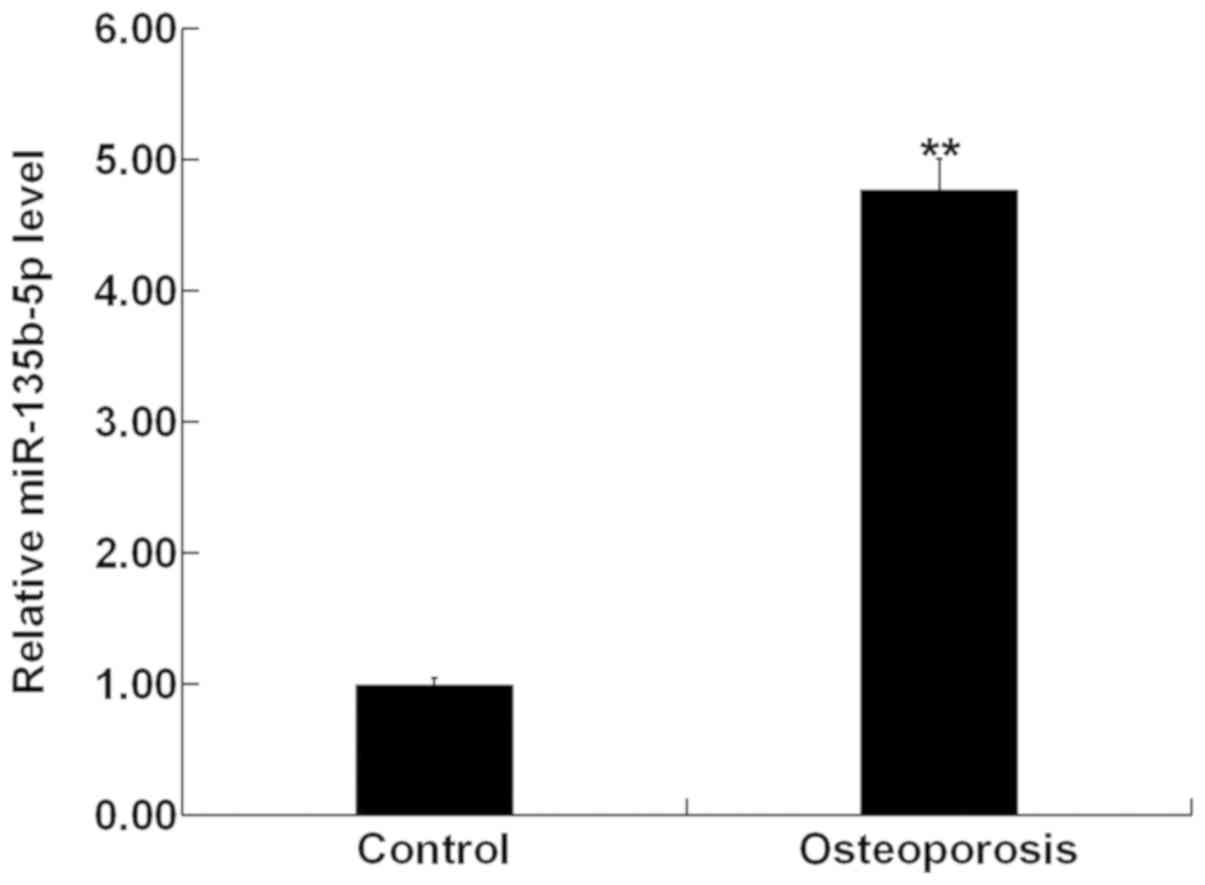
Expression of miR-135b-5p in bone
tissue fragments of patients with OP
The present study used RT-qPCR to investigate the
expression level of miR-135b-5p in bone tissue fragments from 30
patients with OP and 30 patients with osteoarthritis (control
group). The present results showed that compared with the control
group, the expression level of miR-135b-5p was significantly
increased in bone tissue fragments of patients with OP (Fig. 1).
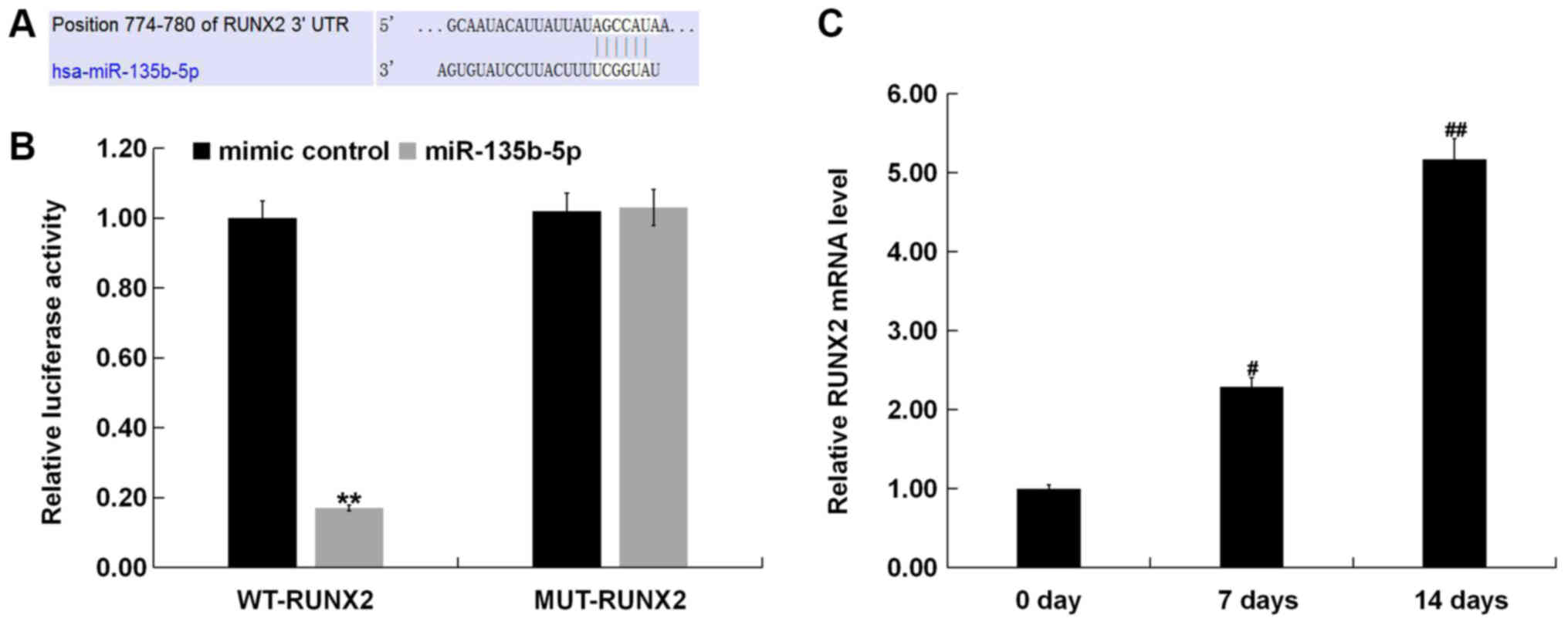
Expression of miR-135b-5p during
osteogenic differentiation
To determine whether osteogenesis could be induced
in MC3T3-E1 cells, RT-qPCR and western blot assay were performed to
detect the mRNA and protein expression levels of osteogenic markers
such as OC, Osterix and ALP. A specific kit was used to examine the
activity of ALP. RT-qPCR results showed that mRNA expression levels
of OC, Osterix and ALP in MC3T3-E1 cells were significantly
increased the 7 and 14th day after induction, compared with the
control group (Fig. 2A-C). Protein
expression levels of OC, Osterix and ALP in MC3T3-E1 cells were
also increased on the 7 and 14th day after induction, compared with
the control group (Fig. 2D). The ALP
assay indicated that ALP activity was significantly increased
(Fig. 2E). To investigate the role
of miR-135b-5p in osteogenic differentiation, the present study
examined the expression of miR-135b-5p by RT-qPCR. The present
results indicated that, compared with the control group, the
expression level of miR-135b-5p was significantly decreased in in
MC3T3-E1 cells on days 7 and 14 after induction (Fig. 2F). The present results suggested that
miR-135b-5p may be involved in the development and progression of
OP by affecting osteogenic differentiation.
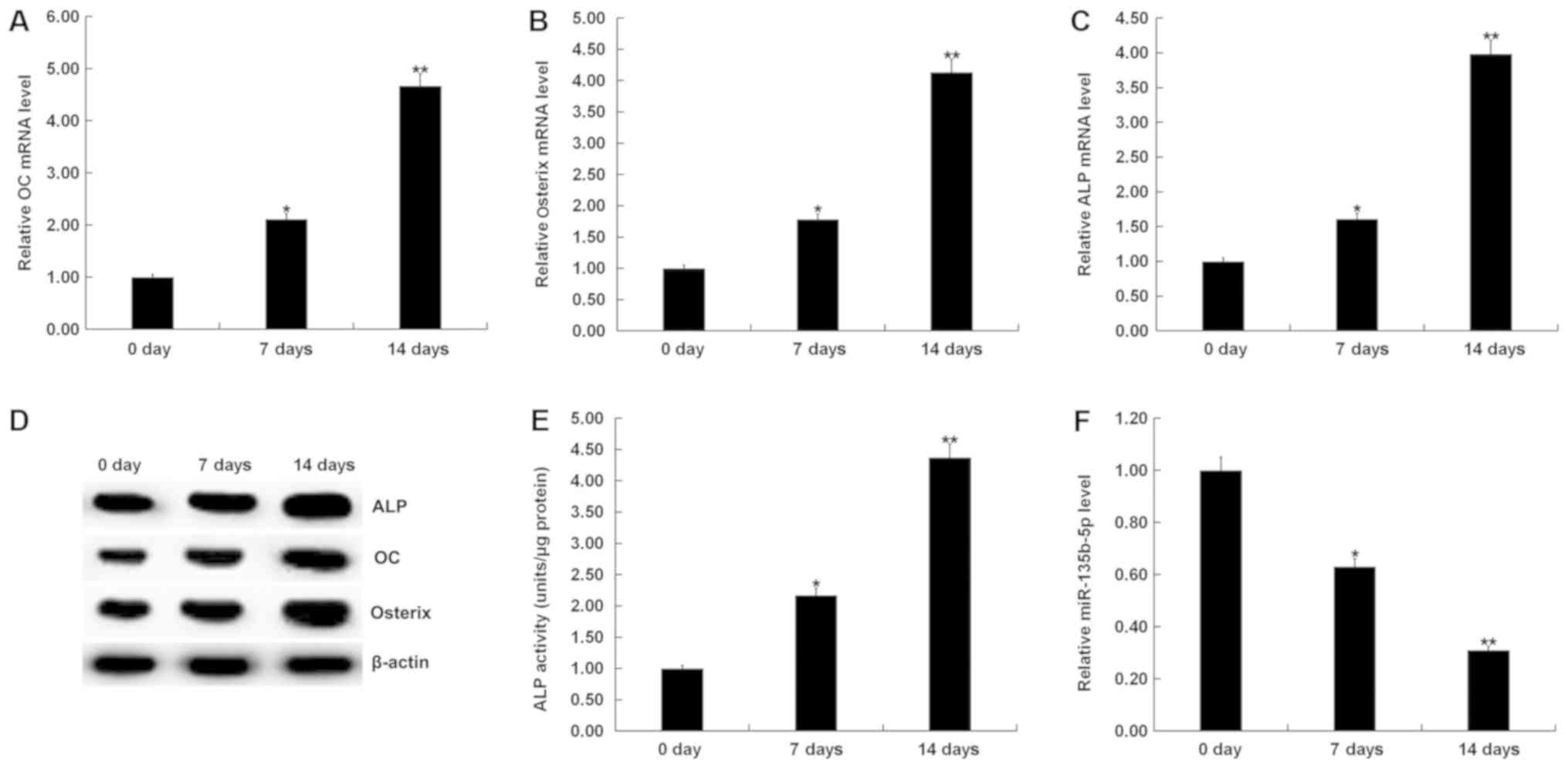 | Figure 2.Expression of miR-135b-5p is
gradually decreased with the prolongation of osteogenic
differentiation induction time. (A) RT-qPCR analysis of OC
expression level on 0, 7 and 14 days after osteogenic
differentiation induction of MC3T3-E1 cells. (B) RT-qPCR analysis
of Osterix expression level on 0, 7 and 14 days after induction of
MC3T3-E1 cells. (C) RT-qPCR analysis of ALP on 0, 7 and 14 days
after induction of MC3T3-E1 cells. (D) Western blot analysis of OC,
Osterix and ALP protein expression levels on 0, 7 and 14 days after
induction of MC3T3-E1 cells. (E) ALP assay was used to detect ALP
activity on 0, 7 and 14 days after induction of MC3T3-E1 cells. (F)
RT-qPCR analysis of miR-135b-5p on 0, 7 and 14 days after induction
of MC3T3-E1 cells. *P<0.05, **P<0.01 vs. 0 day. miR-135b-5p,
microRNA-135b-5p; RT-qPCR, reverse transcription-quantitative PCR;
OC, osteocalcin; ALP, alkaline phosphatase. |
RUNX2 is a direct target of
miR-135b-5p
To reveal the molecular mechanism of the role of
miR-135-5p in OP, TargetScan was used to predict potential targets
of miR-135b-5p, and RUNX2 was identified as a potential target of
miR-135b-5p (Fig. 3A). To further
examine the bioinformatic prediction, a dual-luciferase reporter
assay was performed to detect the relationship between RUNX2 and
miR-135b-5p. The present results showed that miR-135b-5p mimic
significantly suppressed luciferase activity when MC3T3-E1 cells
were cotransfected with a reporter plasmid containing the WT 3′-UTR
and miR-135b-5p mimic (Fig. 3B).
However, the luciferase activity of the MUT 3′-UTR did not change.
RT-qPCR was used to detect the expression of RUNX2, and the results
showed that the expression of RUNX2 was significantly increased in
MC3T3-E1 cells 7 and 14 days after induction compared with the
control group (Fig. 3C).
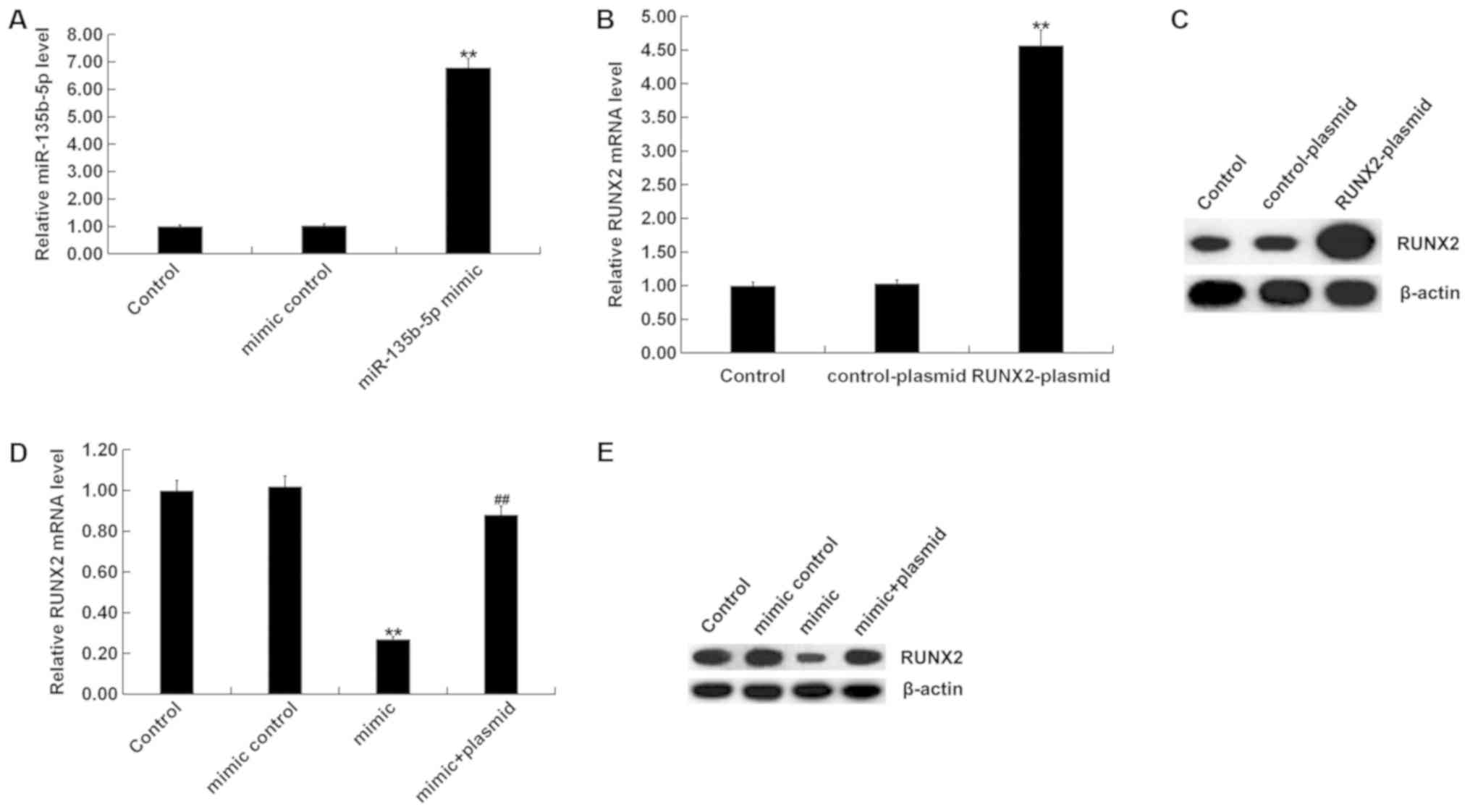
Role of miR-135b-5p in osteogenic
differentiation
To investigate the role of miR-135b-5p in osteogenic
differentiation, MC3T3-E1 cells were transfected with mimic
control, miR-135b-5p mimic, RUNX2-plasmid, control-plasmid or
miR-135b-5p mimic+RUNX2-plasmid for 48 h. RT-qPCR assay and western
blot assay were performed to detect transfection efficiency. The
miR-135b-5p mimic significantly increased the expression level of
miR-135b-5p in MC3T3-E1 cells compared with the control group
(Fig. 4A). RUNX2-plasmid
significantly increased the expression level of RUNX2 in MC3T3-E1
cells (Fig. 4B and C). The
miR-135b-5p mimic significantly reduced the expression level of
RUNX2 in MC3T3-E1 cells, which was reversed by RUNX2
overexpression, compared with the control group (Fig. 4D and E). Osteogenic differentiation
was induced in MC3T3-E1 cells after transfection. The levels of
mRNA and protein expression of the osteogenic markers OC, Osterix
and ALP were detected by RT-qPCR and western blot assay. In
addition, levels of ALP activity were analyzed using a commercially
available kit. RT-qPCR assay and western blot assay results showed
that compared with the control group, the mRNA and protein
expression levels of OC, Osterix and ALP in the induction group
were markedly increased (Fig. 5A-D),
and ALP activity was significantly increased (Fig. 5E). Compared with the induction group,
miR-135b-5p mimic significantly reduced the mRNA and protein
expression levels of OC, Osterix and ALP, and decreased ALP
activity; these reductions were significantly reversed by RUNX2
overexpression. The present results suggested that miR-135b-5p may
be involved in the development and progression of OP by affecting
osteogenic differentiation.
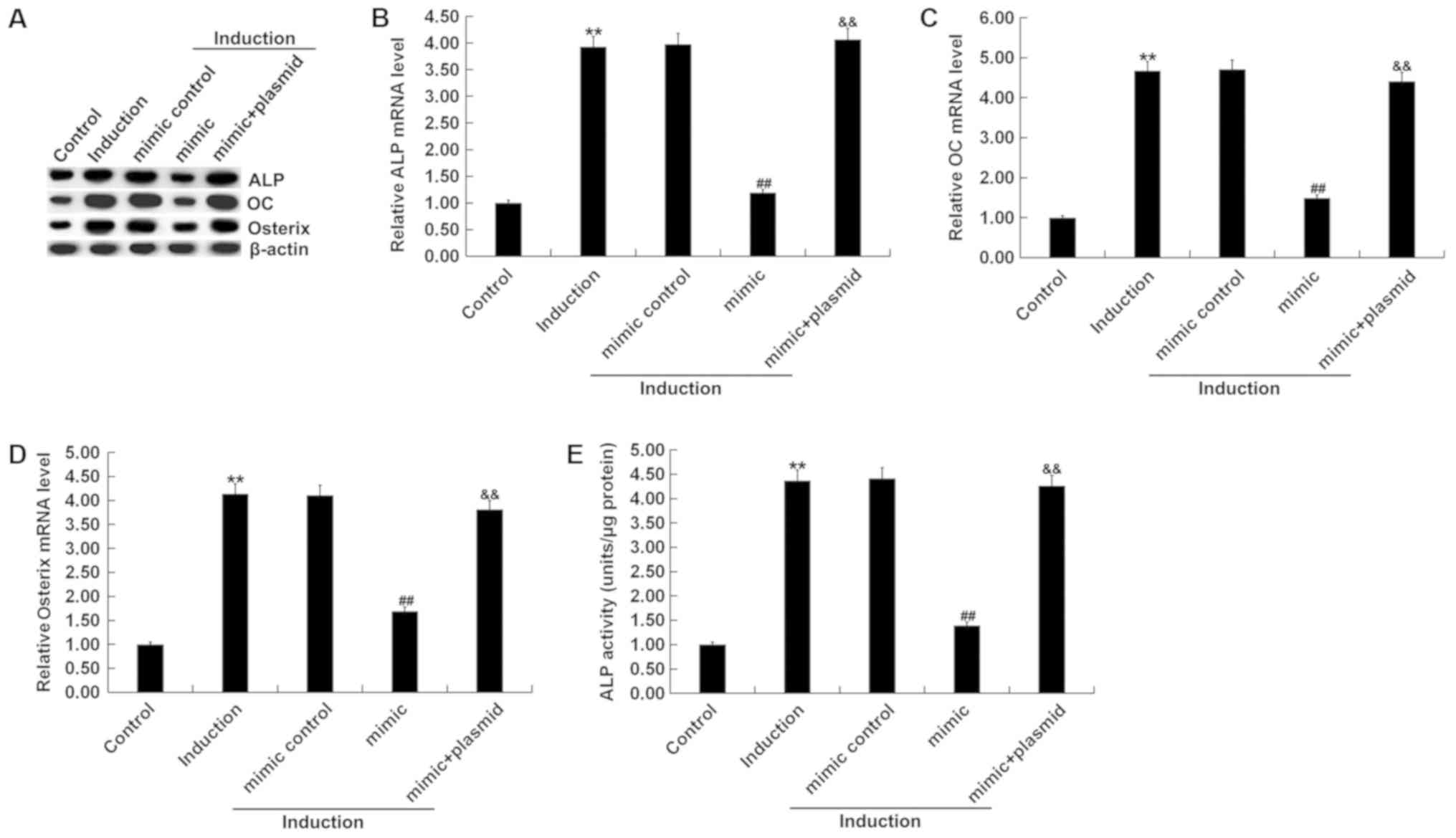 | Figure 5.Role of miR-135b-5p in osteogenic
differentiation. (A) Western blot assay results of the protein
expression level of ALP, OC and Osterix. Reverse
transcription-quantitative PCR detected the mRNA expression levels
of (B) ALP, (C) OC and (D) Osterix. (E) ALP assay was used to
measure ALP activity. Control, MC3T3-E1 cells without any
treatment. Induction, MC3T3-E1 cells subjected to osteogenic
differentiation induction. Mimic control, MC3T3-E1 cells were
transfected with mimic control for 48 h, then subjected to
osteogenic differentiation induction. Mimic, MC3T3-E1 cells were
transfected with miR-135b-5p mimic for 48 h, then subjected to
osteogenic differentiation induction. Mimic + plasmid, MC3T3-E1
cells were transfected with miR-135b-5p mimic + RUNX2-plasmid for
48 h, then subjected to osteogenic differentiation induction.
**P<0.01 vs. control group. ##P<0.01 vs. induction
group. &&P<0.01 vs. mimic group. miR-135b-5p,
microRNA-135b-5p; OC, osteocalcin; ALP, alkaline phosphatase;
RUNX2, runt-related transcription factor 2. |
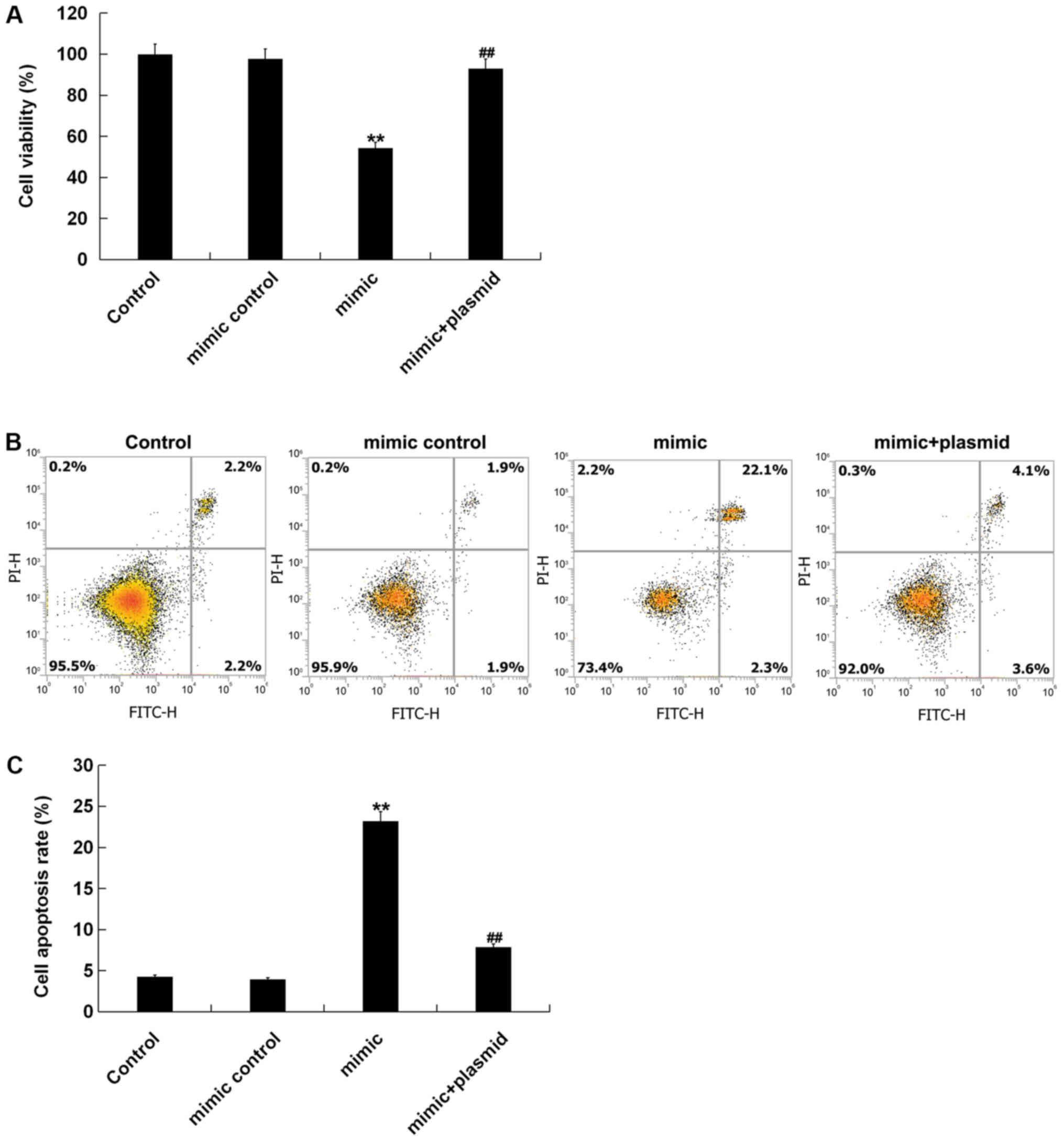
Effect of miR-135b-5p on cell
viability and apoptosis of MC3T3-E1 cells
The present study investigated whether miR-135-5p
had an effect on the viability and apoptosis of MC3T3-E1 cells.
MC3T3-E1 cells were transfected with mimic control, miR-135b-5p
mimic or miR-135b-5p mimic+RUNX2-plasmid for 48 h. Then, cell
viability was detected by MTT. In addition, cell apoptosis was
measured by flow cytometry. The present results showed that
compared with the control group, miR-135b-5p mimic significantly
reduced cell viability of MC3T3-E1 cells and induced cell
apoptosis. These effects, however, were significantly reversed by
RUNX2 overexpression (Fig. 6).
Discussion
The present study investigated the expression of
miR-135b-5p in the bone tissues of patients with OP and explored
its mechanism of action. The present results suggested that
miR-135b-5p was highly expressed in the bone tissues of patients
with OP compared with patients with osteoarthritis. Osteoblasts are
responsible for the formation and mineralization of the skeleton
(27), and play important roles in
OP; therefore, research on osteoblasts has become an important
topic for OP in in vitro research (28,29). The
present study focused on the effects of miR-135b-5p on osteogenic
differentiation and osteoblast proliferation. Therefore, the mouse
pre-osteoblast cell line MC3T3-E1, which is widely used in
osteogenic differentiation investigation (30–32), was
selected for this study. Osteogenic differentiation was induced in
MC3T3-E1 cells. The present results showed that on 7 and 14th day
after induction, the mRNA expression levels of OC, Osterix and ALP
were significantly increased. Protein expression levels of OC,
Osterix and ALP were also increased in MC3T3-E1 cells 7 and 14 days
after osteogenic differentiation induction. ALP activity was
increased and the expression of miR-135b-5p was significantly
decreased in MC3T3-E1 cells on 7 and 14th day after osteogenic
induction. Therefore, the present results suggested that
miR-135b-5p may be involved in the occurrence and development of
osteoporosis by affecting osteogenic differentiation. In addition,
bioinformatics analysis predicted that RUNX2 was a potential target
of miR-135b-5p. Luciferase reporter gene assay was performed to
investigate this interaction. The present results suggested that
the expression level of RUNX2 was increased in MC3T3-E1 cells on 7
and 14th day after osteogenic differentiation induction. The
present results indicated that miR-135b-5p mimic significantly
reduced cell viability and induced cell apoptosis, and that these
effects were significantly reversed by RUNX2-plasmid.
OP is a common and complex disease that has a higher
rate of incidence in the aged population (33,34). OP
is a multifactorial bone disease with microstructural deterioration
and impaired bone strength, making bones susceptible to fragility
and fracture (35). OP is the most
frequent geriatric disease, especially in postmenopausal women
(36). OP occurs due to an imbalance
between osteoclastic bone resorption and osteoblastic bone
formation (37). Restoring and
maintaining the balance between bone formation and bone resorption
is an effective way to treat OP (38,39).
Physical exercise is an important stimulus for prevention and
treatment of osteoporosis (40,41). Due
to the high prevalence and severity of OP, it is important to
understand its pathogenicity and molecular mechanisms for drug
development and treatment.
Previous studies have reported that miRNAs play an
important role in regulating osteoblast differentiation and bone
formation (42,43). miR-135b-5p is a specific miRNA that
has been well studied in cancer and has been identified as an
oncogene or tumor suppressor (44,45). Li
et al (44) showed that the
expression of miR-135b-5p was downregulated in human osteoblastoma
tissues. miR-135b-5p inhibited lipopolysaccharide-induced tumor
necrosis factor-α secretion by suppressing reactive oxygen species
production and NF-κB activation (46). miR-135b-5p was previously found to be
highly expressed in pancreatic cancer (45). However, to the best of our knowledge,
there is little research on the effect and mechanism of miR-135b-5p
in OP. The present results suggested that the expression level of
miR-135b-5p was upregulated in bone tissue fragments of patients
with OP. However, the present study did not analyzed the
correlation of miR-135b-5p expression and the characteristics of
patients with OP (age, sex, BMI and Bone Mineral Density). This is
limitation of the present study, and will need to be investigated
in future studies.
To further examine the specific mechanism in which
miR-135b-5p may regulate MC3T3-E1 cells, the bioinformatics
software TargetScan was used to predict the target gene of
miR-135b-5p. Dual luciferase reporter gene assay results indicated
that RUNX2 was a direct target of miR-135b-5p. A previous study
indicated that miR-135b inhibited osteoblastoma cells by mediating
the expression of protein phosphatase Mg2+/Mn2+ dependent 1E
(47). RUNX2 is a key regulator of
bone osteogenic differentiation (48–50).
RUNX2 promotes bone formation and inhibits bone resorption by
regulating specific extracellular matrix protein expression levels
and the cell cycle of osteoblasts (50). Previous studies have reported that
miR-338-3p can suppress tumor cell proliferation and metastasis in
osteosarcoma cells by targeting RUNX2 (51). Li et al (52) demonstrated that miR-129-1-3p
regulated circulating stretch-induced differentiation of
endothelial progenitor cells by targeting RUNX2. The present
results suggested that osteogenic differentiation inhibition caused
by miR-135b-5p was suppressed by RUNX2 overexpression. In addition,
the present results suggested that miR-135b-5p mimic significantly
reduced cell viability and induced cell apoptosis in MC3T3-E1
cells, and that all these effects were significantly reversed by
RUNX2 overexpression.
In conclusion, the present results suggested that
miR-135-5p may be involved in the occurrence and development of OP
via the inhibition of osteogenic differentiation and osteoblast
proliferation by targeting RUNX2. However, the present study was a
preliminary study of the role of miR-135b-5p in OP. Therefore,
further research is required to understand the role of miR-135b-5p,
such as the analysis of the relationship between miR-135b-5p
expression and the characteristics of patients with OP. The
mechanism of miR-135b-5p in human osteoblast should also be
investigated. In addition, in vivo studies should be
performed, and will be addressed in future experiments.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BC and WY contributed to study design, data
collection, statistical analysis, data interpretation and
manuscript preparation. HZ and KL contributed to data collection,
statistical analysis and manuscript preparation. AD, GZ and KP
contributed to data collection and data interpretation.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Third Affiliated Hospital of Sun Yat-sen
University and every patient provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cosman F, de Beur SJ, LeBoff MS, Lewiecki
EM, Tanner B, Randall S and Lindsay R: Erratum to: Clinician's
guide to prevention and treatment of osteoporosis. Osteoporos Int.
26:2045–2047. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sotorník I: Osteoporosis-epidemiology and
pathogenesis. Vnitr Lek. 62 (Suppl 6):S84–S87. 2016.(In Czech).
|
|
3
|
Body JJ, Bergmann P, Boonen S, Boutsen Y,
Devogelaer JP, Goemaere S, Kaufman JM, Rozenberg S and Reginster
JY: Evidence-based guidelines for the pharmacological treatment of
postmenopausal osteoporosis: A consensus document by the Belgian
Bone Club. Osteoporos Int. 21:1657–1680. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen P, Li Z and Hu Y: Prevalence of
osteoporosis in China: A meta-analysis and systematic review. BMC
Public Health. 16:10392016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Svedbom A, Hernlund E, Ivergård M,
Compston J, Cooper C, Stenmark J, McCloskey EV, Jönsson B and Kanis
JA; EU Review Panel of IOF, : Osteoporosis in the European Union: A
compendium of country-specifc reports. Arch Osteoporos. 8:1372013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ge DW, Wang WW, Chen HT, Yang L and Cao
XJ: Functions of microRNAs in osteoporosis. Eur Rev Med Pharmacol
Sci. 21:4784–4789. 2017.PubMed/NCBI
|
|
7
|
Dadra A, Aggarwal S, Kumar P, Kumar V,
Dibar DP and Bhadada SK: High prevalence of vitamin D deficiency
and osteoporosis in patients with fragility fractures of hip: A
pilot study. J Clin Orthop Trauma. 10:1097–1100. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leutner M, Matzhold C, Bellach L,
Deischinger C, Harreiter J, Thurner S, Klimek P and Kautzky-Willer
A: Diagnosis of osteoporosis in statin-treated patients is
dose-dependent. Ann Rheum Dis. 78:1706–1711. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Loh FE, Stuart B, Sturpe D, Davidoff A,
Onukwugha E and Hochberg M: Osteoporosis medication use: A
comparison of elderly females living in long-term care facilities
versus community Dwellers. Sr Care Pharm. 34:109–126.
2019.PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–97. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Darnet S, Moreira FC, Hamoy IG, Burbano R,
Khayat A, Cruz A, Magalhães L, Silva A, Santos S, Demachki S, et
al: High-throughput sequencing of miRNAs reveals a tissue signature
in gastric cancer and suggests novel potential biomarkers.
Bioinform Biol Insights. 9 (Suppl 1):S1–S8. 2015.
|
|
12
|
Tseng CW, Lin CC, Chen CN, Huang HC and
Juan HF: Integrative network analysis reveals active microRNAs and
their functions in gastric cancer. BMC Syst Biol. 5:992011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cui H, Wang L, Gong P, Zhao C, Zhang S,
Zhang K, Zhou R, Zhao Z and Fan H: Deregulation between miR-29b/c
and DNMT3A is associated with epigenetic silencing of the CDH1
gene, affecting cell migration and invasion in gastric cancer. PLoS
One. 10:e01239262015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He Y, Wang J, Wang J, Yung VY, Hsu E, Li
A, Kang Q, Ma J, Han Q, Jin P, et al: MicroRNA-135b regulates
apoptosis and chemoresistance in colorectal cancer by targeting
large tumor suppressor kinase 2. Am J Cancer Res. 5:1382–1395.
2015.PubMed/NCBI
|
|
15
|
Imam JS, Plyler JR, Bansal H, Prajapati S,
Bansal S, Rebeles J, Chen HI, Chang YF, Panneerdoss S, Zoghi B, et
al: Genomic loss of tumor suppressor miRNA-204 promotes cancer cell
migration and invasion by activating AKT/mTOR/Rac1 signaling and
actin reorganization. PLoS One. 7:e523972012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Valeri N, Braconi C, Gasparini P, Murgia
C, Lampis A, Paulus-Hock V, Hart JR, Ueno L, Grivennikov SI, Lovat
F, et al: MicroRNA-135b promotes cancer progression by acting as a
downstream effector of oncogenic pathways in colon cancer. Cancer
Cell. 25:469–483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Y, Xu D, Bao C, Zhang Y, Chen D, Zhao
F, Ding J, Liang L, Wang Q, Liu L, et al: MicroRNA-135b, a HSF1
target, promotes tumor invasion and metastasis by regulating RECK
and EVI5 in hepatocellular carcinoma. Oncotarget. 6:2421–2433.
2015.PubMed/NCBI
|
|
18
|
Lu Y, Hu J, Sun W, Li S, Deng S and Li M:
MiR-29c inhibits cell growth, invasion, and migration of pancreatic
cancer by targeting ITGB1. Onco Targets Ther. 9:99–109.
2016.PubMed/NCBI
|
|
19
|
Wang H, Zhu Y, Zhao M, Wu C, Zhang P, Tang
L, Zhang H, Chen X, Yang Y and Liu G: miRNA-29c suppresses lung
cancer cell adhesion to extracellular matrix and metastasis by
targeting integrin β1 and matrix metalloproteinase2 (MMP2). PLoS
One. 8:e701922013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seeliger C, Karpinski K, Haug AT, Vester
H, Schmitt A, Bauer JS and van Griensven M: Five freely circulating
miRNAs and bone tissue miRNAs are associated with osteoporotic
fractures. J Bone Miner Res. 29:1718–1728. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Q, Li Y and Zhang Y, Ma L, Lin L,
Meng J, Jiang L, Wang L, Zhou P and Zhang Y: LncRNA MEG3 inhibited
osteogenic differentiation of bone marrow mesenchymal stem cells
from postmenopausal osteoporosis by targeting miR-133a-3p. Biomed
Pharmacother. 89:1178–1186. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li H, Xie H, Liu W, Hu R, Huang B, Tan YF,
Xu K, Sheng ZF, Zhou HD, Wu XP and Luo XH: A novel microRNA
targeting HDAC5 regulates osteoblast differentiation in mice and
contributes to primary osteoporosis in humans. J Clin Invest.
19:3666–3677. 2009. View
Article : Google Scholar
|
|
23
|
Li CJ, Cheng P, Liang MK, Chen YS, Lu Q,
Wang JY, Xia ZY, Zhou HD, Cao X, Xie H, et al: MicroRNA-188
regulates age-related switch between osteoblast and adipocyte
differentiation. J Clin Invest. 125:1509–1522. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Guo B, Li Q, Peng J, Yang Z, Wang
A, Li D, Hou Z, Lv K, Kan G, et al: miR-214 targets ATF4 to inhibit
bone formation. Nat Med. 19:93–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng F, Wang F and Xu Z: MicroRNA-98-5p
prevents bone regeneration by targeting high mobility group AT-Hook
2. Exp Ther Med. 18:2660–2666. 2019.PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using realtime quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zou W, Greenblatt MB, Brady N, Lotinun S,
Zhai B, de Rivera H, Singh A, Sun J, Gygi SP, Baron R, et al: The
microtubule-associated protein DCAMKL1 regulates osteoblast
function via repression of Runx2. J Exp Med. 210:1793–1806. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu B, Xue F, Zhang C and Li G: Ginkgolide
B promotes osteoblast differentiation via activation of canonical
Wnt signalling and alleviates osteoporosis through a bone anabolic
way. J Cell Mol Med. 23:5782–5793. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao W, Zhang WL, Yang B, Sun J and Yang
MW: NIPA2 regulates osteoblast function via its effect on apoptosis
pathways in type 2 diabetes osteoporosis. Biochem Biophys Res
Commun. 513:883–890. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yuan M, Wang Y and Qin YX: SPIO-Au
core-shell nanoparticles for promoting osteogenic differentiation
of MC3T3-E1 cells: Concentration-dependence study. J Biomed Mater
Res A. 105:3350–3359. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhai F, Song N, Ma J, Gong W, Tian H, Li
X, Jiang C and Wang H: FGF18 inhibits MC3T3-E1 cell osteogenic
differentiation via the ERK signaling pathway. Mol Med Rep.
16:4127–4132. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Y, Xu F, Pei HX, Zhu X, Lin X, Song
CY, Liang QH, Liao EY and Yuan LQ: Vaspin regulates the osteogenic
differentiation of MC3T3-E1 through the PI3K-Akt/miR-34c loop. Sci
Rep. 6:255782016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schuiling KD, Robinia K and Nye R:
Osteoporosis update. J Midwifery Womens Health. 56:615–627. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tucker KL: Dietary intake and bone status
with aging. Curr Pharm Des. 9:2687–2704. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Caillet P: Consensus development
conference: Diagnosis, prophylaxis, and treatment of osteoporosis.
Osteoporosis International. 295:914–915. 1987.
|
|
36
|
Wade SW, Strader C, Fitzpatrick LA,
Anthony MS and O'Malley CD: Estimating prevalence of osteoporosis:
Examples from industrialized countries. Arch Osteoporos. 9:1822014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Raisz LG: Pathogenesis of osteoporosis:
Concepts, conflicts, and prospects. J Clin Invest. 115:3318–3325.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Marie PJ and Kassem M: Osteoblasts in
osteoporosis: Past, emerging, and future anabolic targets. Eur J
Endocrinol. 165:1–10. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ruiz-Gaspà S, Blanch-Rubió J,
Ciria-Recasens M, Monfort J, Tío L, Garcia-Giralt N, Nogués X,
Monllau JC, Carbonell-Abelló J and Pérez-Edo L: Reduced
proliferation and osteocalcin expression in osteoblasts of male
idiopathic osteoporosis. Calcif Tissue Int. 86:220–226. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kerschan-Schindl K: Prevention and
rehabilitation of osteoporosis. Wien Med Wochenschr. 166:22–27.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moreira LD, Oliveira ML, Lirani-Galvão AP,
Marin-Mio RV, Santos RN and Lazaretti-Castro M: Physical exercise
and osteoporosis: Effects of different types of exercises on bone
and physical function of postmenopausal women. Arq Bras Endocrinol
Metabol. 58:514–522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lian JB, Stein GS, van Wijnen AJ, Stein
JL, Hassan MQ, Gaur T and Zhang Y: MicroRNA control of bone
formation and homeostasis. Nat Rev Endocrinol. 8:212–227. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Valenti MT, Dalle Carbonare L and Mottes
M: Role of microRNAs in progenitor cell commitment and osteogenic
differentiation in health and disease (Review). Int J Mol Med.
41:2441–2449. 2018.PubMed/NCBI
|
|
44
|
Li ZW, Zhu YR, Zhou XZ, Zhuo BB and Wang
XD: microRNA-135b expression silences Ppm1e to provoke AMPK
activation and inhibit osteoblastoma cell proliferation.
Oncotarget. 8:26424–26433. 2017.PubMed/NCBI
|
|
45
|
Li P, Fan JB, Gao Y, Zhang M, Zhang L,
Yang N and Zhao X: miR-135b-5p inhibits LPS-induced TNFα production
via silencing AMPK phosphatase Ppm1e. Oncotarget. 7:77978–77986.
2016.PubMed/NCBI
|
|
46
|
Han X, Saiyin H, Zhao J, Fang Y, Rong Y,
Shi C, Lou W and Kuang T: Overexpression of miR-135b-5p promotes
unfavorable clinical characteristics and poor prognosis via the
repression of SFRP4 in pancreatic cancer. Oncotarget.
8:62195–62207. 2017.PubMed/NCBI
|
|
47
|
Stein GS, Lian JB, van Wijnen AJ, Stein
JL, Montecino M, Javed A, Zaidi SK, Young DW, Choi JY and Pockwinse
SM: Runx2 control of organization, assembly and activity of the
regulatory machinery for skeletal gene expression. Oncogene.
23:4315–4329. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xiao ZS, Hjelmeland AB and Quarles LD:
Selective deficiency of the ‘bone-related’ Runx2-II unexpectedly
preserves osteoblast-mediated skeletogenesis. J Biol Chem.
279:20307–20313. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang S, Wei D, Wang D, Phimphilai M,
Krebsbach PH and Franceschi RT: In vitro and in vivo synergistic
interactions between the Runx2/Cbfa1 transcription factor and bone
morphogenetic protein-2 in stimulating osteoblast differentiation.
J Bone Miner Res. 18:705–715. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Komori T: Regulation of osteoblast
differentiation by Runx2. Adv Exp Med Biol. 658:43–9. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jia F, Zhang Z and Zhang X:
MicroRNA-338-3p inhibits tumor growth and metastasis in
osteosarcoma cells by targeting RUNX2/CDK4 and inhibition of MAPK
pathway. J Cell Biochem. 120:6420–6430. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li N, Wang WB, Bao H, Shi Q, Jiang ZL, Qi
YX and Han Y: MicroRNA-129-1-3p regulates cyclic stretch-induced
endothelial progenitor cell differentiation by targeting Runx2. J
Cell Biochem. 120:5256–5267. 2018. View Article : Google Scholar : PubMed/NCBI
|