Introduction
microRNAs (miRNAs) are non-coding, single-stranded
RNAs that repress gene expression by interacting with messenger RNA
(mRNA) by inhibiting mRNA translation or by inducing mRNA cleavage
(1–3). To date, more than 600 human miRNAs
have been identified and up to 30% of all protein-coding genes are
estimated to be regulated by miRNAs (4,5).
Although the precise function of individual miRNAs has not been
characterized, biochemical and genetic studies have revealed that
miRNAs regulate a variety of biological processes, such as cell
proliferation, apoptosis, development and differentiation. In
addition, it has been reported that certain miRNAs are aberrantly
expressed in human cancers, suggesting that miRNAs may function as
oncogenes or tumor-suppressor genes depending on the targets that
are regulated (6–9).
A subgroup of miRNAs control promoter methylation
and chromatin structure by regulating DNA methyltransferases and
histone modifiers. Enhancer of zeste homolog 2 (EZH2) is a member
of the polycomb repressive complex 2, which is involved in the
methylation of histone H3 at lysine 27 (H3K27). EZH2 contributes to
the epigenetic silencing of target genes and regulates the biology
of cancer cells. Overexpression of miR-101 inhibits EZH2 and has a
strong influence on the phenotype of various cancers, including
prostate, breast cancer and bladder transitional cell carcinoma. In
addition, EZH2 has been identified as a target of miR-101 by the
luciferase reporter assay (10,11).
In the present study, the differential expression of
miR-101 and EZH2 in lung cancer and normal lung tissues was
determined. We hypothesized that miR-101 has a role in lung cancer
invasion. To test this hypothesis, a cell invasion assay was
performed, and the expression of genes related to cancer invasion
was determined.
Materials and methods
Cell lines
Lung cancer cells (A549, H522 and HCC1438) were
maintained in RPMI-1640 medium (Gibco BRL, Rockville, MD, USA) with
10% FBS and antibiotics (100 U/ml of penicillin and 100 mg/ml of
streptomycin).
Tissues
Tumor and corresponding normal lung tissue specimens
were obtained from Korean patients with non-small cell lung cancer
who underwent curative resection at the Konyang University
Hospital. None of the patients had received chemotherapy or
radiotherapy prior to surgery. Informed consent was obtained from
each patient before surgery. This study was approved by the
Bioethics Committee of Konyang University Hospital. All of the
tumor and macroscopically normal lung tissue samples were obtained
at the time of surgery, and were rapidly frozen in liquid nitrogen
and stored at −80°C until analysis. Tissue samples were
histologically confirmed by H&E staining.
Immunohistochemistry (IHC) of the global
methylation of H3K27 (H3K27me)
Sections (4 μm) of formalin-fixed paraffin-embedded
tissues were deparaffinized. The antigen was retrieved with 0.01 M
citrate buffer (pH 6.0) by heating the sample in an autoclave at a
controlled final temperature of 121°C for 30 min. Endogenous
peroxidase activity was blocked using 0.3% hydrogen peroxide. For
immunohistochemical detection of H3K27me, specimens were incubated
for 1 h at room temperature with the histone H3 (trimethyl K27)
antibody (1:100; Abcam, Cambridge, MA, USA). The sections were
visualized with 3-3′-diaminobenzidine (DAB), and tissue sections
were counterstained with Mayer's hematoxylin. IHC of H3K27me was
scored as follows: 0, no staining; 1, weak or focal staining of the
cancer cell nuclei; 2, moderate nuclear staining in most cancer
cells; and 3, strong nuclear staining in most cancer cells. When
the score was ≥1, the section was considered to have positive
H3K27me expression.
miRNA precursor transfection
Cells were plated in 6-well plates at a density of
1.5×105 cells/well. The next day, cells were transfected
with 50 nM Pre-miR™ miRNA Precursor (Ambion Inc., Austin, TX, USA)
and Pre-miR™ miRNA Precursor-Negative Control #1 (Ambion Inc.) with
Oligofectamine (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer's instructions.
Quantitative real-time PCR assay
Total RNA was isolated with TRIzol (Gibco-BRL,
Glasgow, UK) according to the protocols of the manufacturer, and
the first strand of cDNA was synthesized using the oligo (dT)
primer system (SuperScript III First-Strand Synthesis System;
Invitrogen). Aliquots of the reaction mixture were used for the
qPCR amplification with the IQ5 system (Bio-Rad Laboratories,
Hercules, CA, USA) using iQ SYBR Green Supermix (Bio-Rad). The PCR
was run for 40 cycles of denaturation at 95°C for 15 sec, annealing
at 55°C for 15 sec and elongation at 72°C for 15 sec. Gene
expression was quantified by the comparative CT method, by
normalizing CT values to the housekeeping gene β-actin. After
amplification, melting curve analysis was performed to ensure the
specificity of the products.
TaqMan miRNA expression assay
qRT-PCR analysis for miRNAs was performed in
duplicate with a TaqMan MicroRNA assay kit (Applied Biosystems,
Foster City, CA, USA) according to the manufacturer's instructions,
and RNU6B was used for normalization.
Western blot analysis
Cells were lysed in Pro-Prep protein extraction
solution (INtRON Biotechnology, Gyeonnggi-do, Korea) 48 h after
transfection. An equal amount of proteins was resolved by 8%
SDS-PAGE gels (Laemmli, 1970). The primary antibodies used for the
analysis were mouse anti-EZH2 (1:1,000; BD Biosciences, San Jose,
CA, USA) and mouse anti-β-actin antibodies (1:2,000; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA).
Chromatin immunoprecipitation assay
(ChIP)
ChIP was carried out using a kit purchased from
Upstate Biotechnology (Charlottesville, VA, USA) according to the
protocol recommended by the manufacturer. The trimethyl H3 (Lys-27)
chip grade antibody was purchased from Abcam. The CDH1 gene was
amplified by PCR from the immunoprecipitated chromatin using the
following primers: sense, 5′-TAGAGGGTCACCGCGTCTAT-3′ and antisense,
5′-TCACAGGTGCTTTGCAGTTC-3′. The PCR product was resolved on a 2%
agarose gel and visualized using ethidium bromide staining and UV
illumination.
Invasion assay
Invasion assays were performed in triplicate using
48-well microchemotaxis chambers with 8-μm pore membranes
(Millipore, Bedford, MA, USA) pre-coated with 10 μg/ml Matrigel (BD
Bioscience). The cells were plated in 6-well plates and treated
with 5 μM 5′-azacytidine for 48 h. The bottom chamber was filled
with 1% penicillin/RPMI-1640, and the cells were then placed in the
upper chamber. A549 cells containing 5 μM 5′-azacytidine were
incubated at 37°C for 24 h. The membranes were fixed and stained
with Diff-Quik reagents (Dade Behring, Inc., Deerfield, IL,
USA).
Statistical analysis
Statistical differences in the expression of miRNAs
in cancer and normal lung tissues were analyzed using a
non-parametric test (Mann-Whitney U test). Statistical differences
in the expression of miRNAs in cell lines relative to the control
were analyzed using the Student's t-test. Statistical significance
of correlations between the global methylation of H3K27 and lung
cancer invasion were analyzed by the Fisher extraction test.
Statistical analysis was performed using SPSS 12.0 computer
software (SPSS Inc., Chicago, IL, USA). A p-value <0.05 was
considered to be statistically significant.
Results
miR-101 and EZH2 are inversely expressed
in lung cancer tissue samples
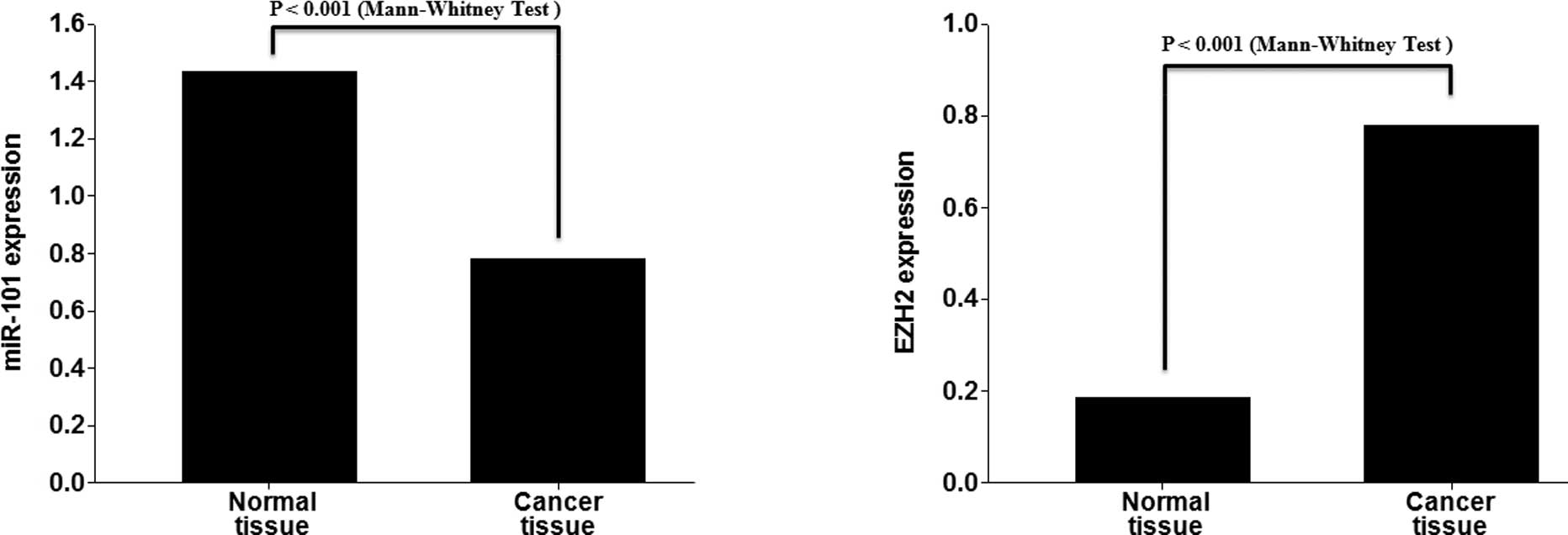
To analyze the expression of miR-101 and EZH2 in
lung cancer, qRT-PCR analysis for miRNAs was performed with 17
matched normal lung and lung cancer tissues. miR-101 exhibited
decreased expression in the lung cancer tissues compared to the
normal lung tissues (p<0.001). In contrast, EZH2 exhibited
increased expression in the lung cancer tissues compared to the
normal lung tissues (p<0.001; Fig.
1A). All cases of the 17 matched normal lung and lung cancer
tissues exhibited a negative correlation (Fig. 1B).
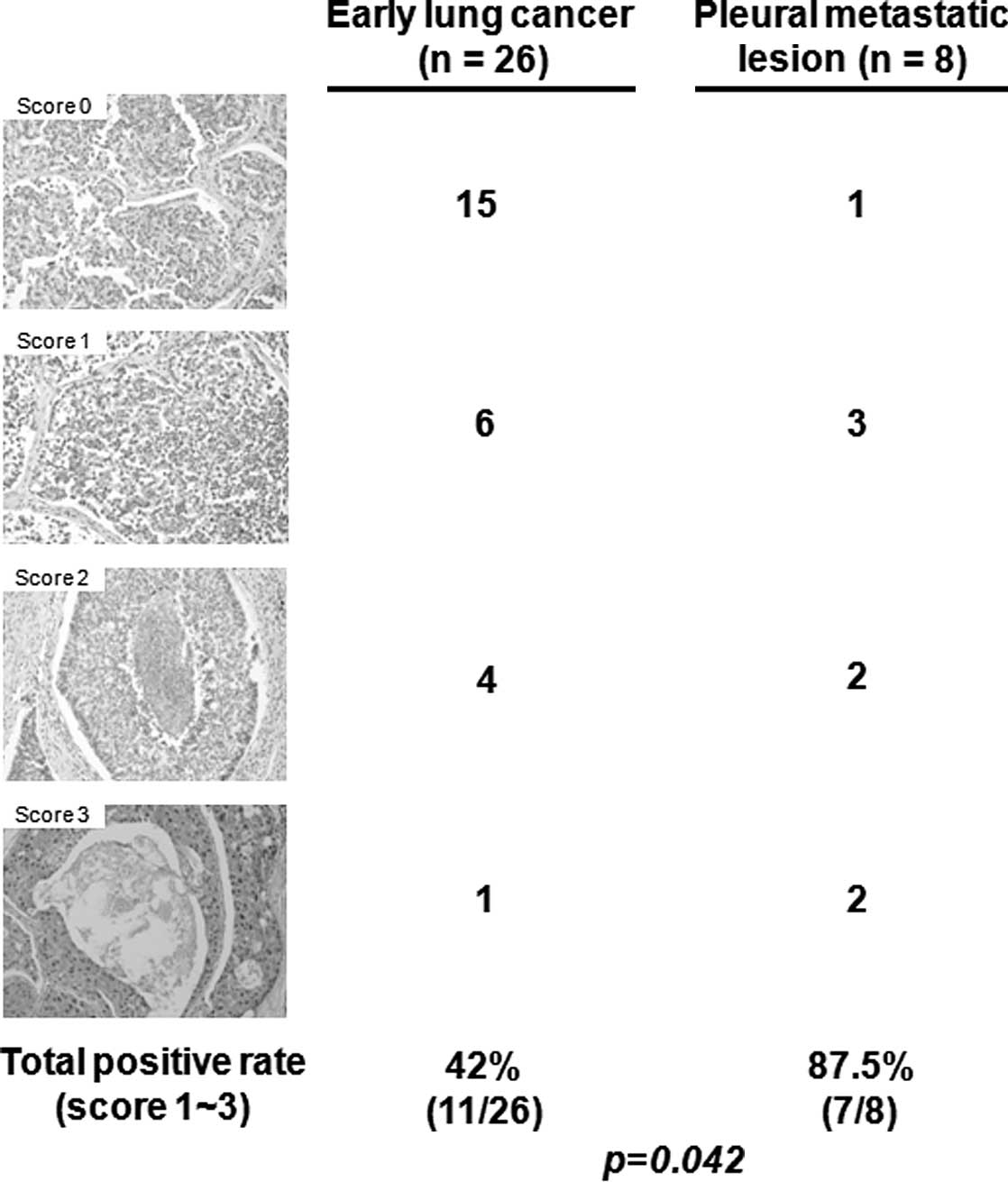
Global methylation of H3K27 was highly
expressed in the metastatic lesions
To determine the association of trimethylation of
H3K27 (H3K27me3) with lung invasion, H3K27me3 was examined by IHC
in 26 early lung cancer tissues without lymph node metastasis and 8
pleural metastatic tissues. H3K27me3 expression was significantly
higher in the pleural metastatic lesions (87.5%) than in the early
lung cancer lesions (42%) (p=0.042; Fig. 2).
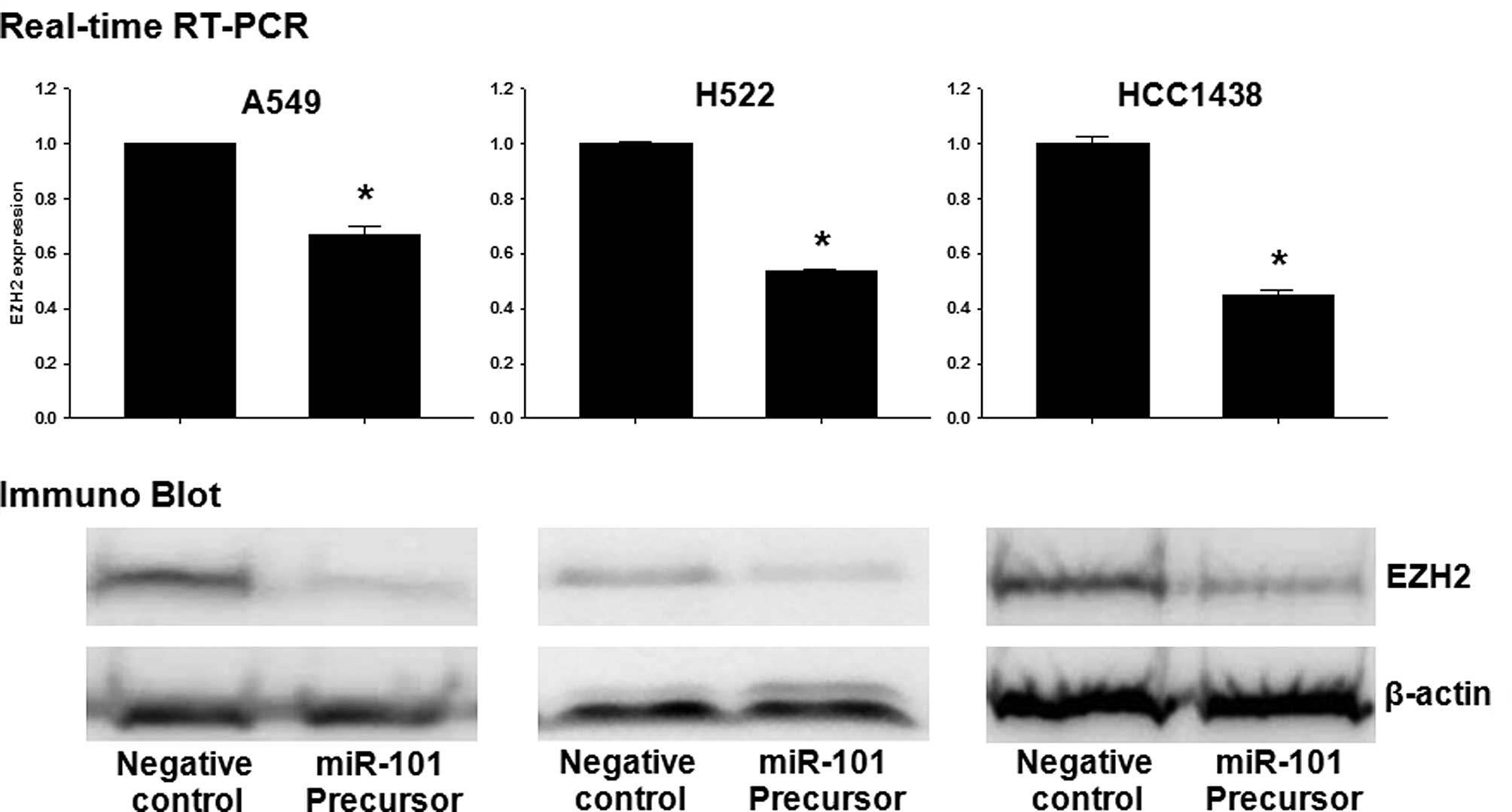
EZH2, CDH1 and MMP-2 are regulated by
miR-101 in lung cancer cells
To determine whether or not overexpression of
miR-101 induces down-regulation of EZH2, qRT-PCR and Western
blotting were performed in A549, H522 and H1438 cells transfected
with the miR (negative control) or miR-101 precursor.
Overexpression of miR-101 induced a marked reduction in EZH2 mRNA
and EZH2 protein (Fig. 3A). To
explore the possible regulation of invasion-related genes by
miR-101 in lung cancer cells, the effect of miR-101 on the
expression of CDH1 and MMP-2 in H1438 cells was analyzed. The
over-expression of miR-101 had an inhibitory effect on the
expression of the MMP-2 gene. In contrast, the expression of CDH1
was increased by overexpression of miR-101 (Fig. 3B). Next, the trimethylation of
H3K27 (H3K27me3) of the CDH promoter was examined using a ChIP
assay. A reduction was found in the trimethyl H3K27 histone mark at
the promoter of CDH1 in the cells transfected with the miR-101
precursor (Fig. 3C).
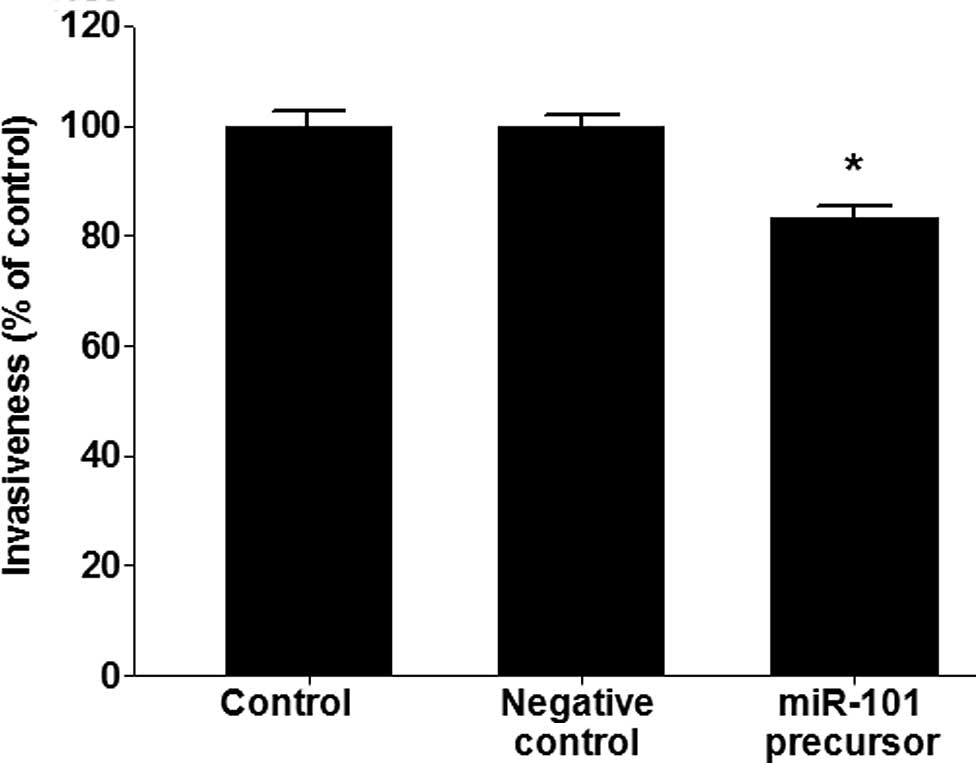
miR-101 functions as an inhibitor of
invasion in lung cancer
To test whether or not miR-101 causes inhibition of
invasion, a cell invasion assay was performed in A549 lung cancer
cells. miR-101 inhibited the invasiveness of A549 cells
(p<0.005; Fig. 4).
Discussion
Epigenetic alteration of genes is one of the central
mechanisms involved in the development and progression of cancer,
and includes hypermethylation of the CpG island on promoters and
epigenetic modification states of histone. The global histone
modification level predicts prognosis in various types of cancers
(12–14). EZH2, which plays a part in the
trimethylation of H3K27, leads to the silencing of tumor-suppressor
genes. EZH2 is frequently overexpressed in various cancers,
including prostate and breast cancer, and is associated with
aggressive forms of these cancers (15,16).
In the present study, expression of H3K27me3 in the metastatic lung
cancer tissues was higher than in early lung cancer. This finding
suggests that EZH2 plays a role in the progression of lung
cancer.
Invasion is the most lethal feature of cancer. The
initial stages of tumor invasion are characterized by the
disruption of cell-cell adhesion. CDH1 (E-cadherin) is involved in
epithelial cell-cell communication and functions as a metastasis
suppressor molecule (17,18). The cytoplasmic tails of cadherins
bind to several cytoplasmic proteins, most notably β-catenin, a
transcriptional cofactor in Wnt signaling (19). Reduced E-cadherin expression is
inversely correlated with lymph node stage and tumor
differentiation in lung cancer (20). Moreover, reduced expression of
E-cadherin is associated with the prognosis of patients with
non-small cell lung cancer (21,22).
The process of invasion also requires a dynamic process, such as
proteolysis. The degradation of extracellular membranes, which
includes the basement membrane, is an early event in proteolysis. A
subgroup of proteases, the matrix metalloproteinases (MMPs), a
family of zinc- and calcium-dependent proteolytic enzymes, are
essential for the breakdown of the basement membrane for metastatic
spread (23,24). In particular, MMP-2 interacts with
integrin αVβ3 and increases VEGF expression through PI3K/AKT
signaling in A549 lung cancer cells (25). Increased expression of MMP-2
promotes cell invasion and is a predictive biomarker for poor
survival in non-small cell lung cancer (26–28).
EZH2 is known to play a role in the regulation of
CDH1 and MMP-2. Overexpression of EZH2 induces silencing of CDH1
via trimethylation of H3K27 in the CDH1 promoter. Notably, a
previous study showed that the expression of CDH1 and MMP-2 is
inversely correlated (29) and
CDH1 regulates the expression of MMP-2 (30,31).
The transition from a non-invasive phenotype to an invasive
phenotype requires reduced CDH1 and overexpression of MMPs
(29,31). Our data suggest that EZH2 functions
as the invasion mediator in lung cancer. Currently, miR-101 is
considered a tumor-suppressive miRNA and has been reported to
suppress cell proliferative and colony forming ability through EZH2
regulation in endometrial, bladder, liver, breast, prostate and
gastric cancer (10,32–34).
miR-101 has multiple predicted targets, such as Cox-2, Mcl-1 and
FOS. These genes have been confirmed as targets of miR-101 by the
luciferase reporter assay (32,33,35).
This evidence suggests that miR-101 modulates multiple steps, such
as proliferation, adhesion and migration of tumor cells, in lung
cancer.
In the present study, miR-101 was underexpressed and
EZH2 was overexpressed in lung cancer. Substantial EZH2 suppression
by miR-101 was observed at the mRNA and protein levels in lung
cancer tissue samples. Moreover, overexpression of miR-101 in a
lung cancer cell line altered the expression of CDH1 and MMP-2.
Targeting tumor invasion and metastasis is an attractive strategy
for the treatment of lung cancer. Therefore, these findings serve
as an incentive for the further investigation of miR-101 as a
therapeutic tool for lung cancer.
Acknowledgements
This study was supported by the
National Research Foundation of Korea Grant funded by the Korean
Government (NRF-2009-0072810).
References
|
1.
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Kim VN and Nam JW: Genomics of microRNA.
Trends Genet. 22:165–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Lim LP, Lau NC, Garrett-Engele P, et al:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Bentwich I, Avniel A, Karov Y, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Berezikov E, Guryev V, van de Belt J,
Wienholds E, Plasterk RHA and Cuppen E: Phylogenetic shadowing and
computational identification of human microRNA genes. Cell.
120:21–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Cheng AM, Byrom MW, Shelton J and Ford LP:
Antisense inhibition of human miRNAs and indications for an
involvement of miRNA in cell growth and apoptosis. Nucleic Acids
Res. 33:1290–1297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Xu P, Guo M and Hay BA: MicroRNAs and the
regulation of cell death. Trends Genet. 20:617–624. 2004.
View Article : Google Scholar
|
|
8.
|
Karp X and Ambros V: Encountering
microRNAs in cell fate signaling. Science. 310:1288–1289. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Chen C-Z, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Varambally S, Cao Q, Mani R-S, et al:
Genomic loss of microRNA-101 leads to overexpression of histone
methyltransferase EZH2 in cancer. Science. 322:1695–1699. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Friedman JM, Liang G, Liu C-C, et al: The
putative tumor suppressor microRNA-101 modulates the cancer
epigenome by repressing the polycomb group protein EZH2. Cancer
Res. 69:2623–2629. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Anglim PP, Alonzo TA and Laird-Offringa
IA: DNA methylation-based biomarkers for early detection of
non-small cell lung cancer: an update. Mol Cancer. 7:812008.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Barlesi F, Giaccone G, Gallegos-Ruiz MI,
et al: Global histone modifications predict prognosis of resected
non small-cell lung cancer. J Clin Oncol. 25:4358–4364. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Ellinger J, Kahl P, von der Gathen J, et
al: Global levels of histone modifications predict prostate cancer
recurrence. Prostate. 70:61–69. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Varambally S, Dhanasekaran SM, Zhou M, et
al: The polycomb group protein EZH2 is involved in progression of
prostate cancer. Nature. 419:624–629. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Collett K, Eide GE, Arnes J, et al:
Expression of enhancer of zeste homologue 2 is significantly
associated with increased tumor cell proliferation and is a marker
of aggressive breast cancer. Clin Cancer Res. 12:1168–1174. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Vleminckx K, Vakaet L, Mareel M, Fiers W
and van Roy F: Genetic manipulation of E-cadherin expression by
epithelial tumor cells reveals an invasion suppressor role. Cell.
66:107–119. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Semb H and Christofori G: The
tumor-suppressor function of E-cadherin. Am J Hum Genet.
63:1588–1593. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Halbleib JM and Nelson WJ: Cadherins in
development: cell adhesion, sorting, and tissue morphogenesis.
Genes Dev. 20:3199–3214. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Sulzer MA, Leers MP, van Noord JA, Bollen
EC and Theunissen PH: Reduced E-cadherin expression is associated
with increased lymph node metastasis and unfavorable prognosis in
non-small cell lung cancer. Am J Respir Crit Care Med.
157:1319–1323. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Liu D, Huang C, Kameyama K, et al:
E-cadherin expression associated with differentiation and prognosis
in patients with non-small cell lung cancer. Ann Thorac Surg.
71:949–955. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Huang C, Liu D, Masuya D, et al: Clinical
application of biological markers for treatments of resectable
non-small cell lung cancers. Br J Cancer. 92:1231–1239. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Liotta LA, Tryggvason K, Garbisa S, Hart
I, Foltz CM and Shafie S: Metastatic potential correlates with
enzymatic degradation of basement membrane collagen. Nature.
284:67–68. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Nelson AR, Fingleton B, Rothenberg ML and
Matrisian LM: Matrix metalloproteinases: biologic activity and
clinical implications. J Clin Oncol. 18:11352000.PubMed/NCBI
|
|
25.
|
Chetty C, Lakka SS, Bhoopathi P and Rao
JS: MMP-2 alters VEGF expression via αVβ3 integrin-mediated
PI3K/AKT signaling in A549 lung cancer cells. Int J Cancer.
127:1081–1095. 2010.
|
|
26.
|
Leinonen T, Pirinen R, Bohm J, Johansson R
and Kosma VM: Increased expression of matrix metalloproteinase-2
(MMP-2) predicts tumour recurrence and unfavourable outcome in
non-small cell lung cancer. Histol Histopathol. 23:693–700.
2008.PubMed/NCBI
|
|
27.
|
Kim S-H, Choi HY, Lee J, et al: Elevated
activities of MMP-2 in the non-tumorous lung tissues of curatively
resected stage I NSCLC patients are associated with tumor
recurrence and a poor survival. J Surg Oncol. 95:337–346. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Hung W-C, Tseng W-L, Shiea J and Chang
H-C: Skp2 overexpression increases the expression of MMP-2 and
MMP-9 and invasion of lung cancer cells. Cancer Lett. 288:156–161.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Polette M, Gilles C, de Bentzmann S,
Gruenert D, Tournier J-M and Birembaut P: Association of
fibroblastoid features with the invasive phenotype in human
bronchial cancer cell lines. Clin Exp Metastasis. 16:105–112. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Miyaki M, Tanaka K, Kikuchi-Yanoshita R,
Muraoka M, Konishi M and Takeichi M: Increased cell-substratum
adhesion, and decreased gelatinase secretion and cell growth,
induced by E-cadherin transfection of human colon carcinoma cells.
Oncogene. 11:2547–2552. 1995.
|
|
31.
|
Nawrocki-Raby B, Gilles C, Polette M, et
al: E-cadherin mediates MMP down-regulation in highly invasive
bronchial tumor cells. Am J Pathol. 163:653–661. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Su H, Yang J-R, Xu T, et al: MicroRNA-101,
down-regulated in hepatocellular carcinoma, promotes apoptosis and
suppresses tumorigenicity. Cancer Res. 69:1135–1142. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Li S, Fu H, Wang Y, et al: MicroRNA-101
regulates expression of the v-fos FBJ murine osteosarcoma viral
oncogene homolog (FOS) oncogene in human hepatocellular carcinoma.
Hepatology. 49:1194–1202. 2009. View Article : Google Scholar
|
|
34.
|
Wang H-J, Ruan H-J, He X-J, et al:
MicroRNA-101 is down-regulated in gastric cancer and involved in
cell migration and invasion. Eur J Cancer. 46:2295–2303. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Strillacci A, Griffoni C, Sansone P, et
al: MiR-101 downregulation is involved in cyclooxygenase-2
overexpression in human colon cancer cells. Exp Cell Res.
315:1439–1447. 2009. View Article : Google Scholar : PubMed/NCBI
|