Introduction
Colorectal cancer (CRC) is the third most common
cancer, and the fourth leading cause of cancer death worldwide
(1). In recent years, improved
treatment has resulted in a 5-year survival rate of about 50%
(2). However, the survival rate
of advanced stage patients, especially of those with lymph node
metastasis, is still low. A series of genes are involved in the
pathogenesis of CRC. Investigation of these genes is important to
clarify the tumorigenic pathway and is helpful for developing new
biomarkers for early detection and accurate diagnosis.
Ubiquitin C-terminal hydrolase-L1 (UCHL1) is a
member of the deubiquitinating enzymes (DUBs), widely expressed in
neuronal tissues, and functions to maintain ubiquitin balance by
associating and releasing ubiquitin (3). Previous studies showed that UCHL1
played an important role in neurodegenerative diseases including
Parkinson's disease and Alzheimer's disease (4). Recently, growing evidence indicates
that UCHL1 also plays a role in tumorigenesis and tumor
progression. Interestingly, the roles of UCHL1 are conflicting in
different cancer types. UCHL1 acts as a tumor promoter in
pancreatic cancer, prostate cancer, and lung cancer (5–7).
However, it acts as a tumor suppressor in ovarian cancer,
hepatocellular cancer, and nasopharyngeal carcinoma (8–10).
In esophageal cancer, breast cancer and CRC, its function is
ambiguous (10,11–15). The role of UCHL1 in cancer cells
is still largely unexplored.
In our previous study, we found that the expression
of UCHL1 was increased in the lymph node metastasis (LNM) CRC group
compared with the non-LNM CRC group. The re-expression of UCHL1
through gene transfection increased the invasive ability of HCT8
cells (16). In this study, we
further demonstrated that UCHL1 re-expression promoted the
proliferation, migration, and metastatic capabilities of HCT8 cells
both in vitro and in vivo. We also found that UCHL1
could decelerate β-catenin degradation depending on its
deubiquitinating activity. Accumulated β-catenin consequently
activated the β-catenin/T cell factor (TCF) pathway and induced the
expression of cyclin D1 and uPA.
Materials and methods
Cell culture and transfection
HCT8 cells were maintained in Roswell Park Memorial
Institute (RPMI)-1640 (Invitrogen, USA) supplemented with 10%
heat-inactivated fetal bovine serum (FBS) at 37°C in a humidified
5% CO2 atmosphere. DNA transfection was performed using
Lipofectamine (Invitrogen) according to the manufacturer's
protocol. For stable transfection, cells were selected using full
growth medium containing G418 (600 μg/ml) for 14 days, and the
positive cells were detected by western blot analysis.
Expression plasmids
Wild-type pcDNA3.1/UCHL1 expression plasmid against
UCHL1 was a gift from Dr Julia Shackelford. The pcDNA3.1/UCHL1-C90S
mutant was generated by inserting a specific mutation at the Cys 90
site in wild-type pcDNA3.1/UCHL1 plasmid by overlap PCR.
Cell growth assay by MTT
Cells were seeded in triplicate into 96-well plates
with 3×103 cells/well. Cells were maintained for 5 days,
and then 20 μl of MTT (Sigma, USA) solution (5 mg/ml) was added and
cells were further incubated at 37°C for 4 h. Absorbance at 492 nm
was determined with a microplate reader. All MTT assays were
repeated three times.
In vitro cell invasion and migration
assays
Cell invasion and migration assays were carried out
in a chemotaxis chamber (Millipore). Cells (1×105)
suspended in 200 μl of serum-free medium were loaded into the upper
chamber and allowed to pass through an 8-μm-pore polyethylene
terephthalate filter. The lower chamber was filled with complete
medium. The chamber was either pre-coated with Matrigel (1:8; BD
Biosciences, Bedford, MA, USA) for invasion assays or left uncoated
for migration assays. Cells that failed to pass through the filters
were removed by scrubbing with cotton swabs after 20 h (migration
assay) or 24 h (invasion assay). Cells on the undersurface were
fixed in methanol and stained with 0.5% crystal violet, then
photographed and quantified in 5 random fields per membrane. Each
sample was assayed in triplicate.
Tumor growth and metastasis in nude
mice
For the tumor growth assay, cells (40 μl,
4×106 cells) were collected and inoculated
subcutaneously into the nail pad of 4-week-old male nude mice.
Experimental and control groups had 5 mice each. Mice were
sacrificed 38 days later. Tumor growth curves were calculated.
After tumor excision, the tissues were fixed in 10% buffered
formalin. All formalin-fixed and paraffin-embedded samples were
carefully examined after staining with hematoxylin and eosin
(H&E) and UCHL1 antibody.
For the tumor metastasis assay, cells (50 μl
2.5×106 cells) were collected and injected into the tail
vein of 4-week-old male nude mice. Each group included 6 mice. Mice
were sacrificed 4 weeks later and examined for metastasis
development. Lungs and visibly observable lymph nodes were excised.
All lungs were fixed with Bouin's fluid. All lymph nodes were
formalin-fixed and paraffin-embedded, then carefully examined after
staining with H&E and UCHL1 antibody.
All animal experiments were conducted according to
the institutional and national guidelines for animal
experiments.
Immunohistochemical staining
Immunohistochemical (IHC) staining was performed as
previously described (16).
Specimens were incubated with anti-UCHL1 (1:500, Abcam) overnight
at 4°C, and then incubated with a biotinylated second antibody
(goat anti-rabbit/IgG 1:100 dilution) for 30 min at 37°C.
Half-life analysis
Cells were treated with cyclohexamide (CHX, 100
μg/ml; Sigma) for 0, 3 or 6 h to inhibit further protein synthesis.
Then cells were harvested and analyzed by western blot
analysis.
Protein preparation and western blot
analysis
For whole-cell protein extraction, cells were lysed
with lysis buffer [50 mmol/l Tris-HCl, pH 7.4; 150 mmol/l NaCl;
1.5% NP-40; 0.1% SDS; 50 μg/ml phenylmethylsulfony fluoride (PMSF);
with freshly added proteinase inhibitor cocktail] for 40 min on
ice, followed by centrifugation at 14,000 × g for 15 min at 4°C.
The supernatants were collected as total protein.
For preparation of cytoplasmic and nuclear protein,
cells were collected and rinsed with cold phosphate-buffered saline
(PBS) twice, then centrifuged at 3000 × g for 5 min at 4°C. The
pellets were suspended in buffer A (10 mM HEPES, pH 7.9; 1.5 mM
MgCl2; 10 mM KCl; 0.5 mM DTT; 0.2% NP-40; 50 μg/ml PMSF;
with freshly added proteinase inhibitor cocktail) and kept for 15
min at 4°C, then centrifuged at 5,000 × g for 5 min. The
supernatants (cytoplasmic protein) were harvested. The insoluble
pellets were then centrifuged at 14,000 × g for 1 min, again after
being washed with cold buffer A. The pellets were lysed in lysis
buffer and kept for 40 min at 4°C, then centrifuged at 14,000 × g
for 10 min. The supernatants were harvested as nuclear protein.
Western blot analysis was performed using conventional protocols as
described previously (16).
Statistical analysis
All statistical analyses were performed using SPSS
13.0 software (SPSS Inc., Chicago, IL, USA). Data were expressed as
means ± SD. The Student's t-test was used for statistical
comparison. P-values <0.05 indicated statistical
significance.
Results
UCHL1 promotes proliferation, migration
and invasion capabilities of HCT8 cells in vitro
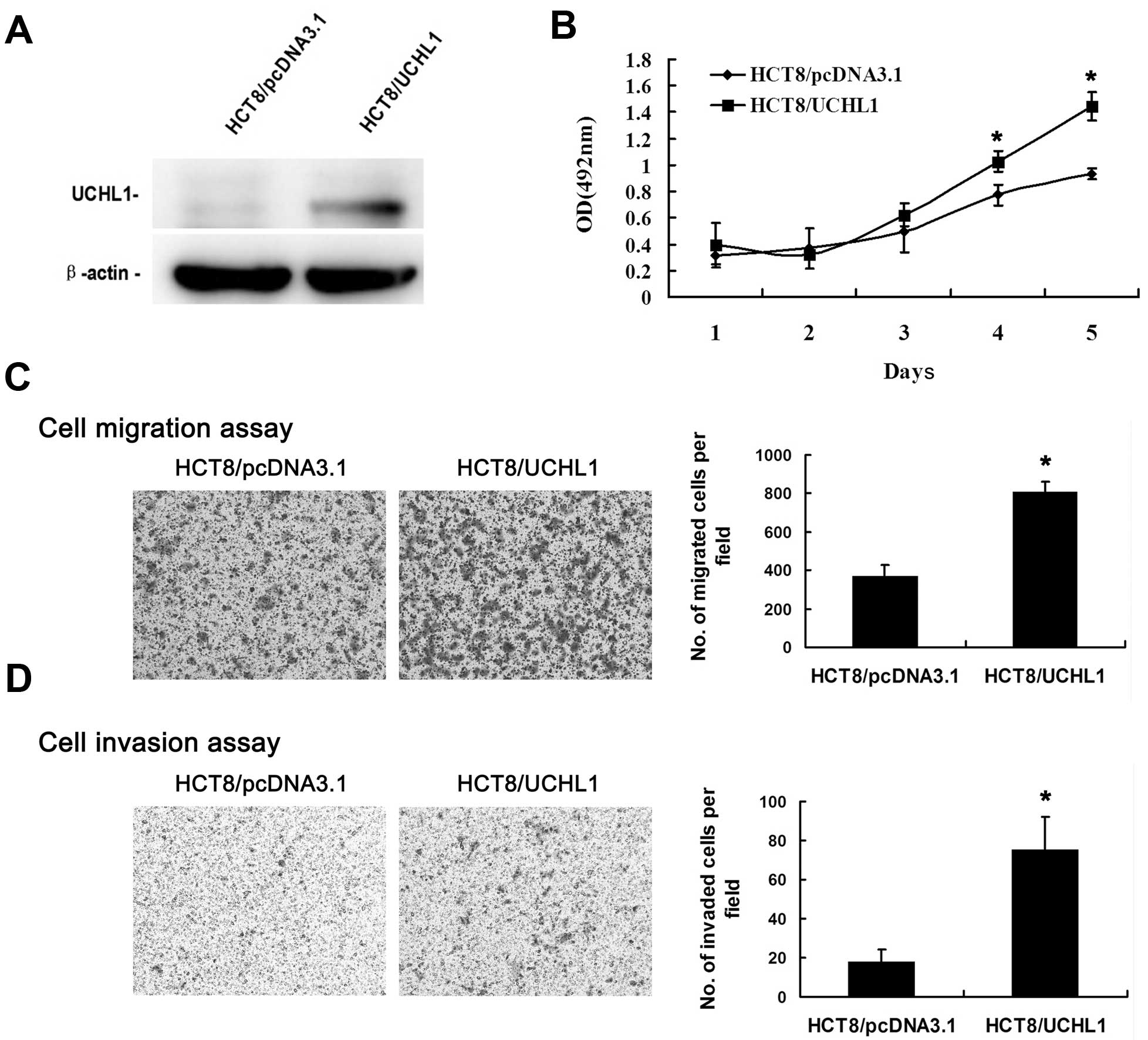
As we previously reported (16), HCT8 did not express endogenous
UCHL1. Thus, we used HCT8 to generate the UCHL1 stable cell line.
The HCT8/UCHL1 stable cells expressed high exogenous UCHL1, and the
HCT8/pcDNA3.1 cells were used as empty vector control (Fig. 1A). HCT8/UCHL1 cells showed
increased cell proliferation, migration, and invasion capabilities
in vitro compared with control cells (P<0.05) (Fig. 1B–D). These results implied that
UCHL1 promoted the proliferation, migration, and invasion
capabilities of HCT8 cells in vitro.
UCHL1 promotes tumorigenesis and
metastasis of HCT8 cells in vivo
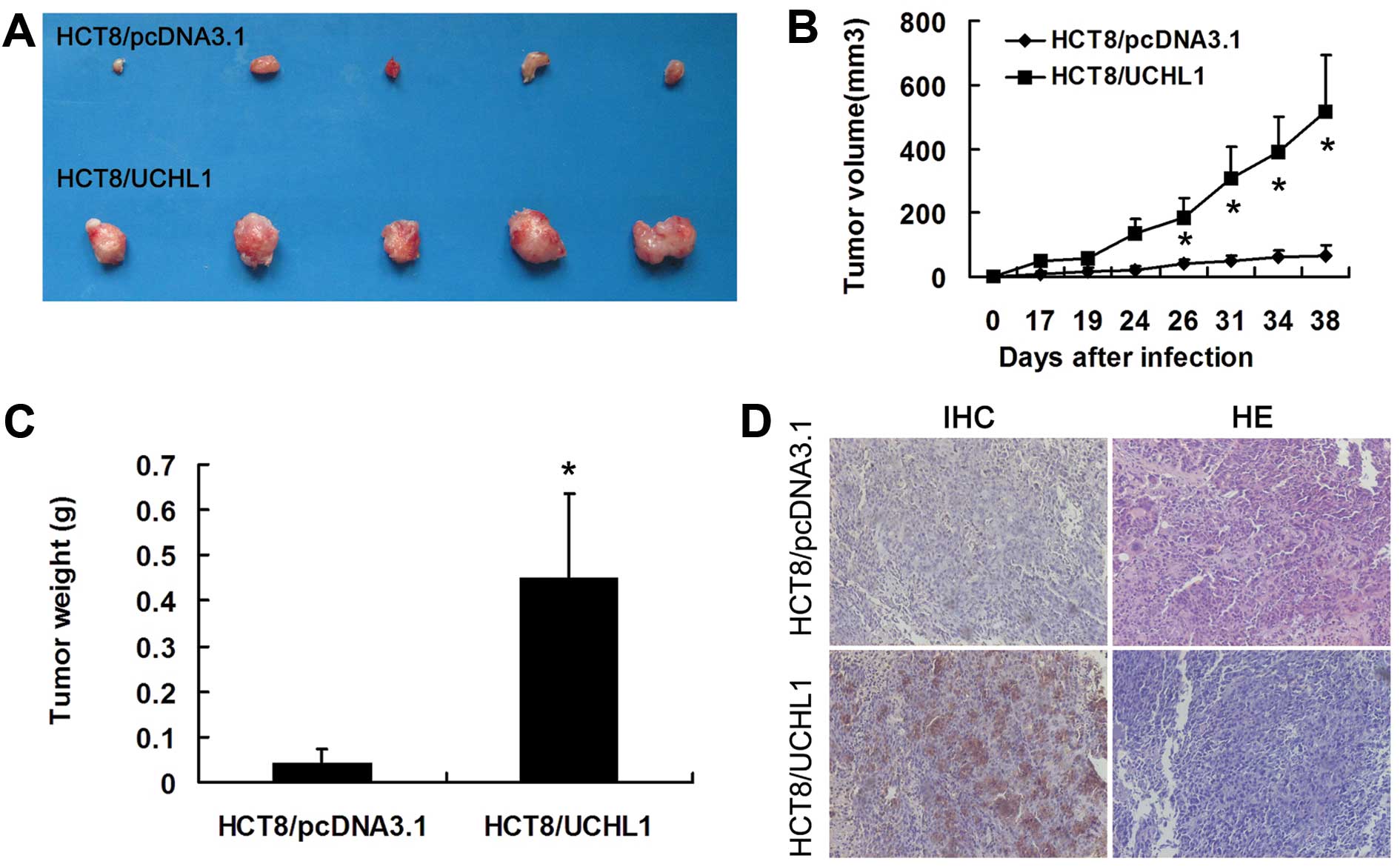
To investigate UCHL1 functions in vivo, we
injected HCT8/pcDNA3.1 and HCT8/UCHL1 cells into nude mice through
the nail pad. Mice were humanely sacrificed and dissected for
tumors 38 days later. Tumor nodules resected from the HCT8/UCHL1
group showed positive UCHL1 staining (Fig. 2D). In addition, the nodules from
this group were larger and heavier than those from the control
group (P<0.05) (Fig. 2A–C).
However, we did not find lymph node metastasis or lung metastasis
in either group in this model.
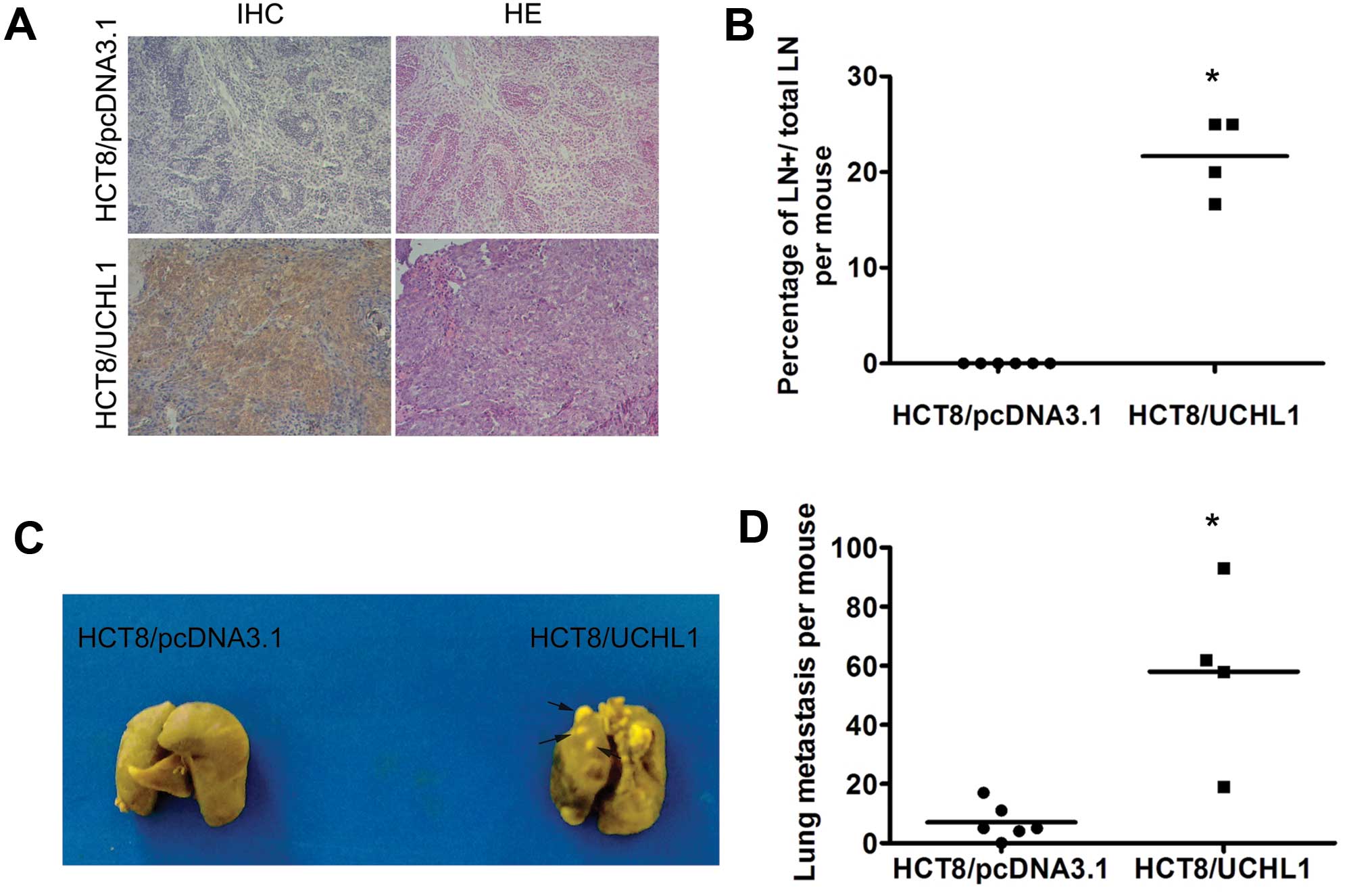
To investigate whether UCHL1 promotes tumor
metastasis in vivo, we injected HCT8/pcDNA3.1 and HCT8/UCHL1
cells into nude mice via tail veins. Mice were sacrificed 4 weeks
later. In the HCT8/UCHL1 group, 2 mice died before the end of the
experiment. In the remaining 4 mice, all of them developed lymph
node metastases, accompanied with positive UCHL1 staining (Fig. 3A). However, no lymph node
metastasis was found in HCT8/pcDNA3.1 group. The percentage of
metastatic lymph nodes with respect to the total visible lymph
nodes per mouse was significantly higher in the HCT8/UCHL1 group
(22%) than in the control group (0%; P<0.05) (Fig. 3B). Furthermore, UCHL1
re-expression dramatically promoted lung metastasis of HCT8 cells.
The metastatic loci in the HCT8/UCHL1 group (58±30) were
significantly more than those in the control group (7±6; P<0.05)
(Fig. 3C–D). These results
indicated that UCHL1 promoted tumorigenesis and metastasis of HCT8
cells in vivo.
Deubiquitinase activity is crucial for
UCHL1 to promote the malignant transformation of HCT8 cells
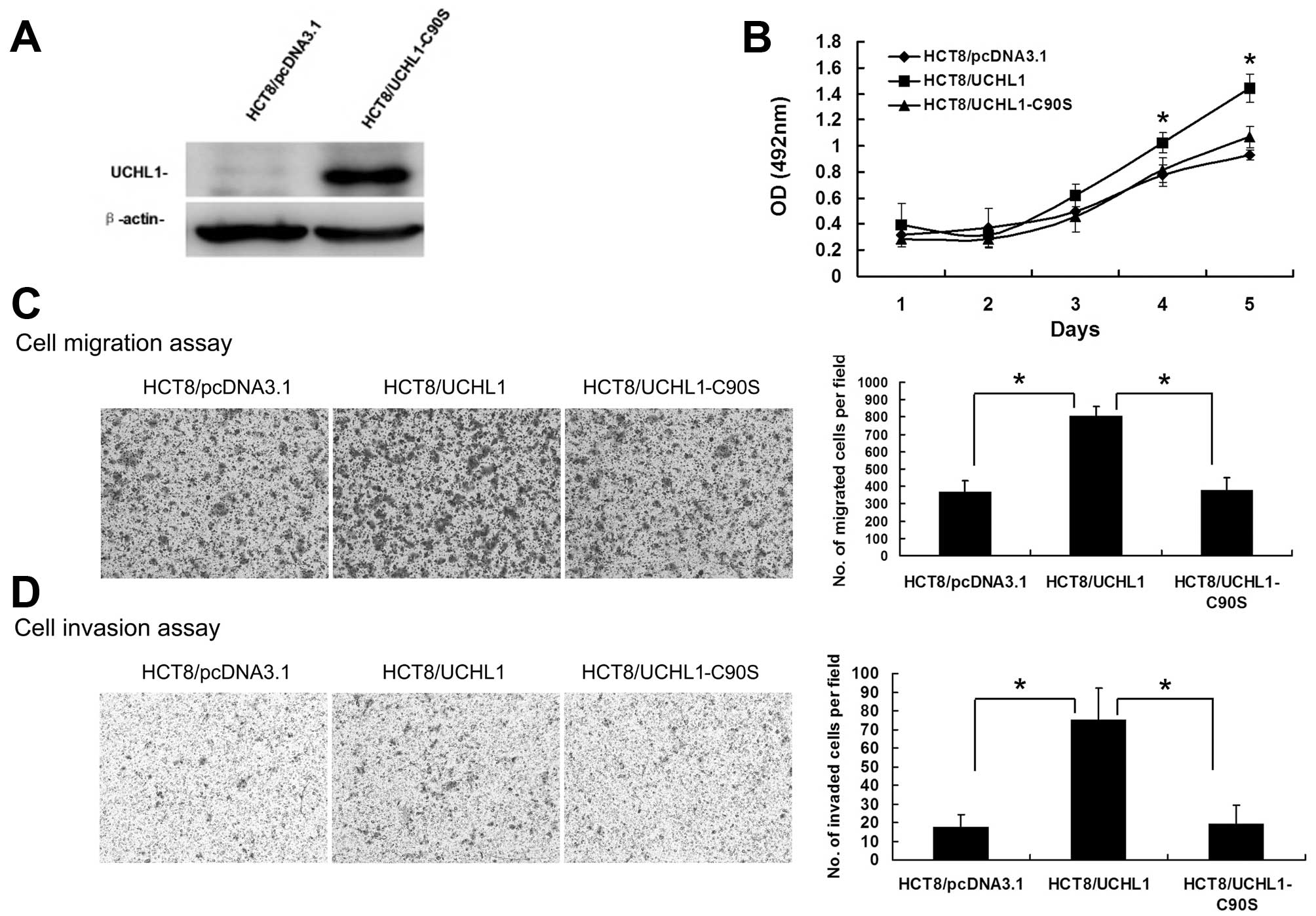
To investigate if the deubiquitinating capability is
essential for UCHL1 function, we constructed a C90S mutant (with
cysteine 90 converted to serine), with impaired deubiquitinase
activity (17). Then the
HCT8/UCHL1-C90S stable cell line was generated (Fig. 4A). The proliferation, migration,
and invasion capabilities of HCT8/UCHL1-C90S cells were similar to
those of HCT8/pcDNA3.1 cells (Fig.
4B–D). These results indicated that the malignant
transformation promoting function of UCHL1 was dependent on its
deubiquitinating activity.
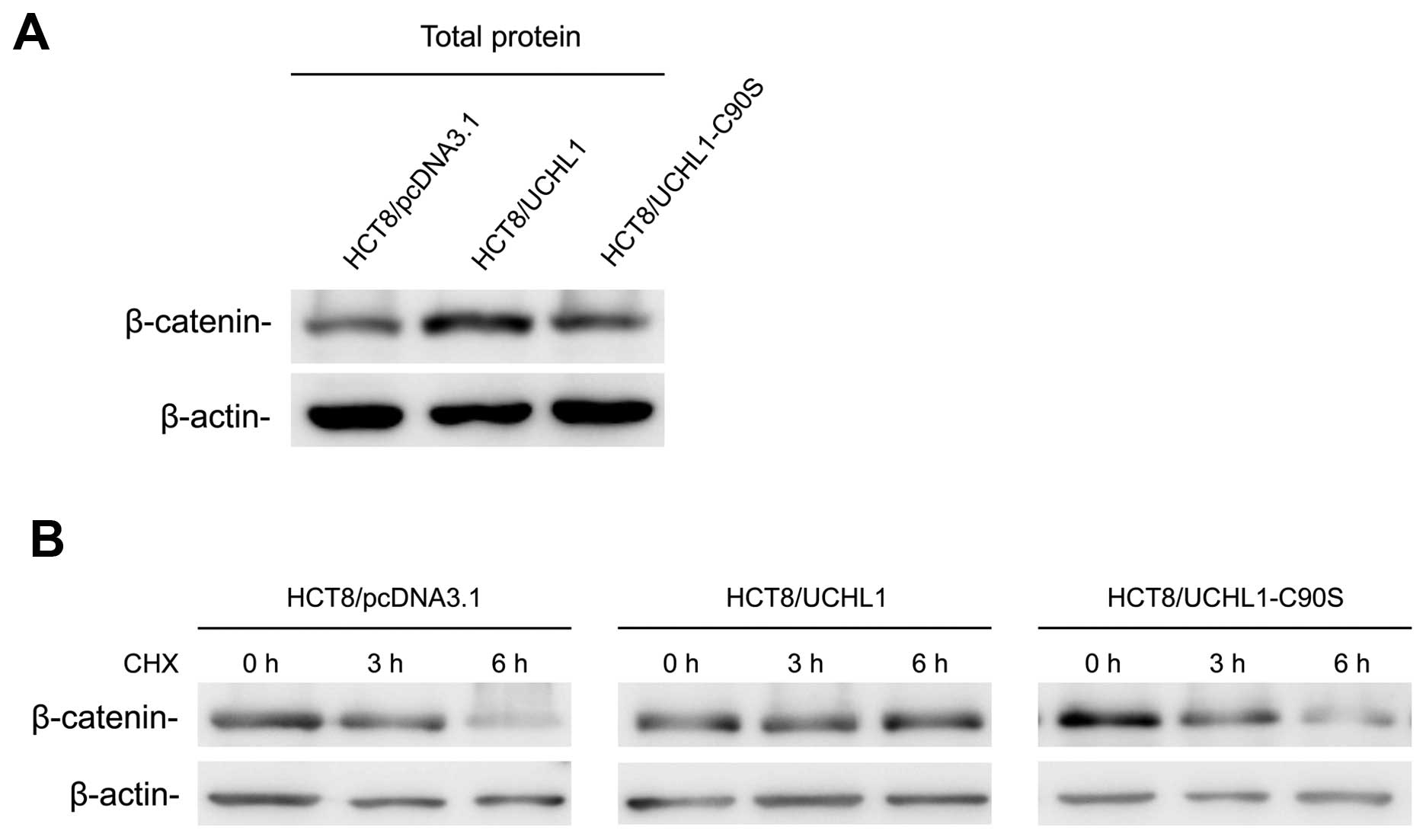
UCHL1 upregulates β-catenin by enhancing
its stability
To see whether UCHL1 could affect β-catenin
expression in HCT8 cells, we detected β-catenin in three stable
cell lines. Our results confirmed that β-catenin expression was
upregulated by UCHL1, but could not be upregulated by the
UCHL1-C90S mutant (Fig. 5A).
To determine whether UCHL1 upregulates β-catenin
expression by enhancing its stability, a half-life assay was
performed. The data showed that UCHL1 re-expression significantly
extended the half-life of β-catenin. In contrast, we failed to
observe this function in the UCHL1-C90S mutant (Fig. 5B). These results indicated that
UCHL1 upregulated β-catenin by enhancing its stability, depending
on its deubiquitinating activity.
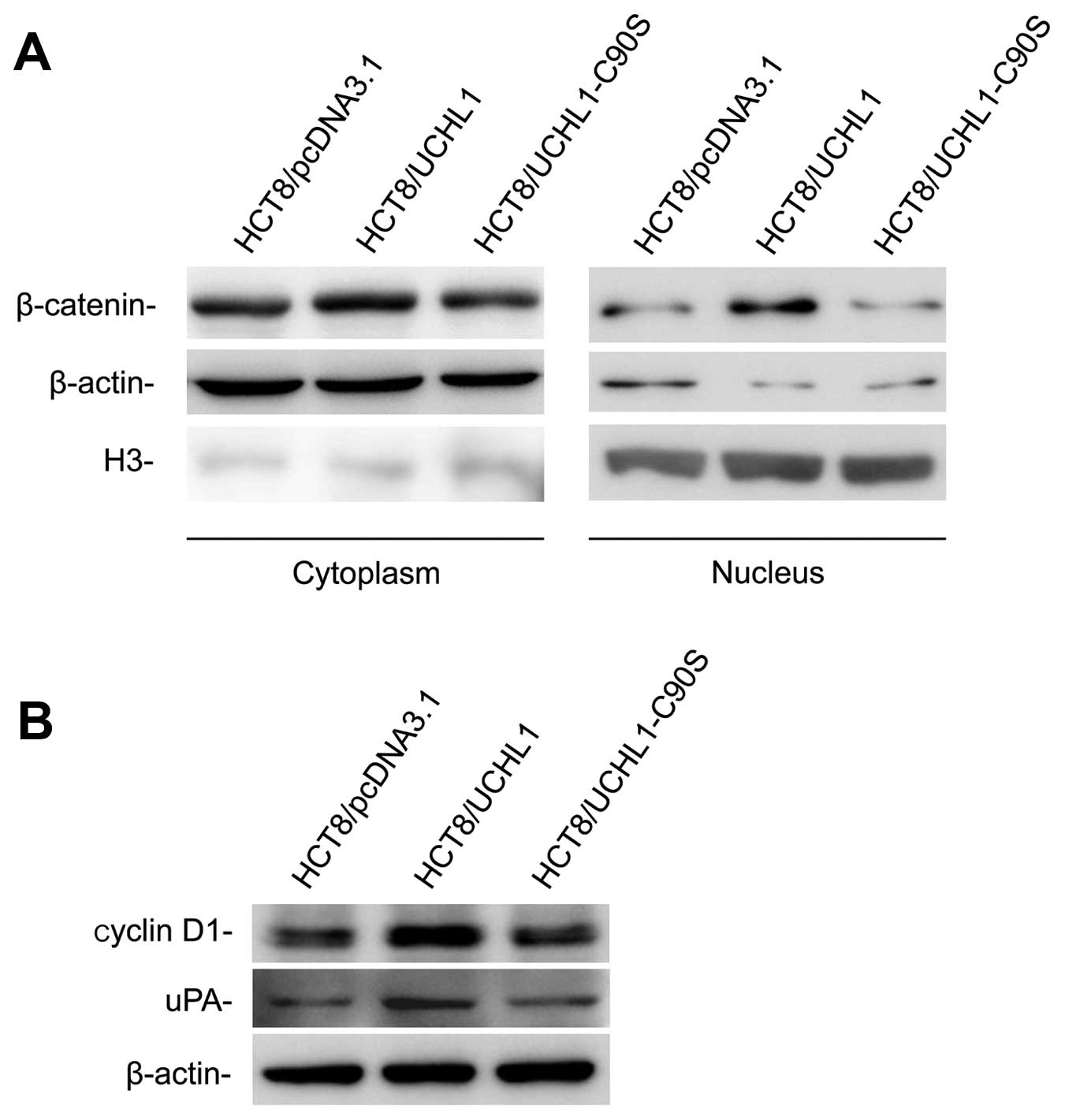
UCHL1 induces modification of
proliferation, migration and invasion-related genes
To further confirm the effect of UCHL1 on
β-catenin/TCF signaling, the immunoblotting analyses were carried
out using the protein isolated from the cell cytoplasm and nucleus
in three stable cell lines. Our results demonstrated that UCHL1
re-expression increased the accumulation of β-catenin in cytoplasm,
which resulted in increased β-catenin translocation into cell
nucleus (Fig. 6A). However,
β-catenin remained unchanged in HCT8/UCHL1-C90S cells compared with
HCT8/pcDNA 3.1 cells.
Cyclin D1 and serine protease urokinase plasminogen
activator (uPA) are transcriptionally induced by β-catenin in the
nucleus (18,19). As shown in Fig. 6B, both molecules were upregulated
in HCT8/UCHL1 cells, but not in HCT8/pcDNA 3.1 and HCT8/UCHL1-C90S
cells. These data suggested that UCHL1 activated the β-catenin/TCF
signaling, depending on its deubiquitinating activity.
Discussion
Although much evidence indicates that UCHL1 is
related to CRC, the mechanism is still ambiguous. Some
investigations have shown that UCHL1 methylation was more frequent
in CRC tissues than in normal colorectal tissues (9,10,20), whereas others have indicated that
high UCHL1 expression was related to colorectal tumor progression,
invasion, lymph node metastasis, and poor clinical outcome
(15,21–23). In this study, we confirmed our
previous results by establishing that UCHL1 played an oncogenic
role by positively regulating the proliferation, invasion, and
metastasis of HCT8 in vitro and in vivo. Nail pad
injection of stable HCT8/UCHL1 cells promoted tumor formation in
nude mice. These data are consistent with that UCHL1 expression is
related to tumor size and tumor stage in CRC (15,16). In addition, tail vein injection of
stable HCT8/UCHL1 cells promoted both lymph node and lung
metastases in nude mice. These data are consistent with that UCHL1
expression is related to lymph node metastases in clinical
colorectal patients (16,23).
The Wnt/β-catenin pathway is implicated in over 90%
of colon cancers. The activation of β-catenin results in
constitutive activity of β-catenin TCF transcriptional complex,
which drives tumor formation (24). While exploring the mechanism by
which UCHL1 promoted the malignant transformation of HCT8, we found
that UCHL1 upregulated the expression of β-catenin by decelerating
its degradation depending on its deubiquitinating activity. In
agreement with our data, a recent study has also demonstrated that
UCHL1 upregulated β-catenin signaling in 293 and NIH 3T3 cells
(25). It is well known that the
β-catenin subcellular distribution is greatly related to its
oncogenic transcriptional regulation. The stabilized form of
β-catenin accumulates in cytoplasm and is then translocated into
the cell nucleus, where it carries out a pivotal role in
tumorigenesis by activating the Wnt signaling target genes. In our
investigation, UCHL1 induction increased the accumulation of
β-catenin in the cytoplasm, which resulted in increased β-catenin
translocation into the cell nucleus. Then upregulation of cyclin D1
and uPA were detected. Both cyclin D1 and uPA are target genes of
β-catenin (18,19). Cyclin D1 is a major regulator of
the progression of cells into the proliferative stage of the cell
cycle (26). The increased
expression of cyclin D1 can stimulate CDK activity and promote the
cell cycle progression. uPA is a protease involved in degradation
of the extracellular matrix. Upregulation of uPA promotes tumor
progression, particularly tumor invasion and metastasis (27,28). These results agree with the cell
behaviors that we have demonstrated in this study. Thus, we believe
that UCHL1 promotes the malignant transformation of HCT8 by
activating β-catenin signaling.
Several studies have demonstrated the importance of
the deubiquitinating activity of UCHL1 in cancer progression and
metastasis (5,13,25). In our study, C90S, a mutant of
UCHL1 with impaired deubiquitinating activity, did not induce the
activation of β-catenin signaling and it did not promote the
proliferation, migration, and invasion capabilities of HCT8 cells.
However, identification of other target genes of UCHL1
deubiquitinating activity will provide more insight into the
mechanisms of UCHL1 in oncogenesis in CRC.
The conflicting study results of UCHL1 function in
CRC may be because of the inadequate sample and the exact subtype
of CRC. Besides, UCHL1, which contains two opposing enzymatic
activities, deubiquitinating enzyme activity and ubiquitin ligase
enzyme activity, may function differently under certain
circumstances. Thus, further study is needed to elaborate the exact
role of UCHL1, to clarify the function of UCHL1 in the tumorigenic
pathway and to assist in the development of novel biomarkers for
early detection and accurate diagnosis.
In summary, we provide both in vitro and
in vivo evidence that UCHL1 functions as an oncogene by
increasing cell proliferation, migration, and invasion capabilities
in HCT8 cells through the β-catenin/TCF pathway depending on its
deubiquitinating activity.
Acknowledgements
This research was supported by the
National Key Basic Research Program of China (2007CB914700). We are
grateful to Dr Julia Shackelford (Lineberger Comprehensive Cancer
Center, University of North Carolina at Chapel Hill, Chapel Hill,
North Carolina, USA) for providing us with wild-type pcDNA3.1/UCHL1
plasmid.
References
|
1.
|
DM ParkinF BrayJ FerlayP PisaniGlobal
cancer statistics, 2002CA Cancer J
Clin5574108200510.3322/canjclin.55.2.74
|
|
2.
|
B LevinP BoyleWorld cancer report
2008International Agency for Research on CancerLyon3743792008
|
|
3.
|
H OsakaYL WangK TakadaS TakizawaR SetsuieH
LiY SatoK NishikawaYJ SunM SakuraiUbiquitin carboxy-terminal
hydrolase L1 binds to and stabilizes monoubiquitin in neuronHum Mol
Genet1219451958200310.1093/hmg/ddg21112913066
|
|
4.
|
R SetsuieK WadaThe functions of UCH-L1 and
its relation to neurodegenerative diseasesNeurochem
Int51105111200710.1016/j.neuint.2007.05.00717586089
|
|
5.
|
HJ KimYM KimS LimYK NamJ JeongHJ KimKJ
LeeUbiquitin C-terminal hydrolase-L1 is a key regulator of tumor
cell invasion and
metastasisOncogene28117127200910.1038/onc.2008.36418820707
|
|
6.
|
A LeiblichSS CrossJW CattoG PesceFC HamdyI
RehmanHuman prostate cancer cells express neuroendocrine cell
markers PGP 9.5 and chromogranin
AProstate6717611769200710.1002/pros.2065417929277
|
|
7.
|
E TezelK HibiT NagasakaA NakaoPGP9.5 as a
prognostic factor in pancreatic cancerClin Cancer
Res6476447672000
|
|
8.
|
L LiQ TaoH JinA van HasseltFF PoonX WangMS
ZengWH JiaYX ZengAT ChanY CaoThe tumor suppressor UCHL1 forms a
complex with p53/MDM2/ARF to promote p53 signaling and is
frequently silenced in nasopharyngeal carcinomaClin Cancer
Res1629492958201010.1158/1078-0432.CCR-09-317820395212
|
|
9.
|
E Okochi-TakadaK NakazawaM WakabayashiA
MoriS IchimuraT YasugiT UshijimaSilencing of the UCHL1 gene in
human colorectal and ovarian cancersInt J
Cancer11913381344200610.1002/ijc.2202516642472
|
|
10.
|
J YuQ TaoKF CheungH JinFF PoonX WangH LiYY
ChengC RöckenMP EbertEpigenetic identification of ubiquitin
carboxyl-terminal hydrolase L1 as a functional tumor suppressor and
biomarker for hepatocellular carcinoma and other digestive
tumorsHepatology48508518200810.1002/hep.22343
|
|
11.
|
DL MandelkerK YamashitaY TokumaruK
MimoriDL HowardY TanakaAL CarvalhoWW JiangHL ParkMS KimPGP9.5
promoter methylation is an independent prognostic factor for
esophageal squamous cell carcinomaCancer
Res6549634968200510.1158/0008-5472.CAN-04-392315930319
|
|
12.
|
Y MiyoshiS NakayamaY TorikoshiS TanakaH
IshiharaT TaguchiY TamakiS NoguchiHigh expression of ubiquitin
carboxy-terminal hydrolase-L1 and -L3 mRNA predicts early
recurrence in patients with invasive breast cancerCancer
Sci97523529200610.1111/j.1349-7006.2006.00202.x16734731
|
|
13.
|
T TakaseK HibiT YamazakiH NakayamaM
TaguchiY KasaiK ItoS AkiyamaT NagasakaA NakaoPGP9.5 overexpression
in esophageal squamous cell
carcinomaHepatogastroenterology5012781280200314571718
|
|
14.
|
WJ WangQQ LiJD XuXX CaoHX LiF TangQ ChenJM
YangZD XuXP LiuOverexpression of ubiquitin carboxy terminal
hydrolase-L1 induces apoptosis in breast cancercellsInt J
Oncol3310371045200818949367
|
|
15.
|
T YamazakiK HibiT TakaseE TezelH NakayamaY
KasaiK ItoS AkiyamaT NagasakaA NakaoPGP9.5 as a marker for invasive
colorectal cancerClin Cancer Res8192195200211801558
|
|
16.
|
Y MaM ZhaoJ ZhongL ShiQ LuoJ LiuJ WangX
YuanC HuangProteomic profiling of proteins associated with lymph
node metastasis in colorectal cancerJ Cell
Biochem11015121519201010.1002/jcb.2272620524204
|
|
17.
|
CN LarsenJS PriceKD WilkinsonSubstrate
binding and catalysis by ubiquitin C-terminal hydrolases:
identification of two active
siteresiduesBiochemistry3567356744199610.1021/bi960099f8639624
|
|
18.
|
E HiendlmeyerS RegusS WassermannF HlubekA
HaynlA DimmlerC KochC KnollM van BeestU ReuningBeta-catenin
upregulates the expression of the urokinase plasminogen activator
in human colorectal tumorsCancer
Res6412091214200410.1158/0008-5472.CAN-3627-214973118
|
|
19.
|
M ShtutmanJ ZhurinskyI SimchaC AlbaneseM
D'AmicoR PestellA Ben-Ze'evThe cyclin D1 gene is a target of the
β-catenin/LEF-1 pathwayProc Natl Acad Sci USA96552255271999
|
|
20.
|
S FukutomiN SekiK KodaM
MiyazakiIdentification of methylation-silenced genes in colorectal
cancer cell lines genomic screening using oligonucleotide
arraysScand J
Gastroenterol4214861494200710.1080/00365520701491173
|
|
21.
|
Y Akishima-FukasawaY InoY NakanishiA
MiuraY MoriyaT KondoY KanaiS HirohashiSignificance of PGP9.5
expression in cancer-associated fibroblasts for prognosis of
colorectal carcinomaAm J Clin
Pathol1347179201010.1309/AJCPRJP39MIDSGBH20551269
|
|
22.
|
Y HongKS HoKW EuPY CheahA susceptibility
gene set for early onset colorectal cancer that integrates diverse
signaling pathway: implication for tumorigenesisClin Cancer
Res1311071114200710.1158/1078-0432.CCR-06-163317317818
|
|
23.
|
H MizukamiA ShirahataT GotoM SakataM
SaitoK IshibashiG KigawaH NemotoY SanadaK HibiPGP9.5 methylation as
a marker for metastatic colorectal cancerAnticancer
Res2826972700200819035297
|
|
24.
|
M BienzH CleversLinking colorectal cancer
to Wnt
signalingCell103311320200010.1016/S0092-8674(00)00122-711057903
|
|
25.
|
A BhedaW YueA GullapalliC WhitehurstR
LiuJS PaganoJ ShackelfordPositive reciprocal regulation of
ubiquitin C-terminal hydrolase L1 and beta-catenin/TCF
signalingPLoS One4e5955200910.1371/journal.pone.000595519536331
|
|
26.
|
CJ SherrCancer cell
cyclesScience27416721677199610.1126/science.274.5293.16728939849
|
|
27.
|
S MazaudR GuyotCJ GuigonN CoudouelB Le
Magueresse-BattistoniS MagreBasal membrane remodeling during
follicle histogenesis in the rat ovary: contribution of proteinases
of the MMP and PA familiesDev
Biol277403416200510.1016/j.ydbio.2004.10.00115617683
|
|
28.
|
H MorganPA HillHuman breast cancer
cell-mediated bone collagen degradation requires plasminogen
activation and matrix metalloproteinase activityCancer Cell
Int51200510.1186/1475-2867-5-1
|