Introduction
Cadmium is a toxic heavy metal and an environmental
pollutant. Evidence suggests that cadmium, due to its long
half-life, can accumulate in the body, mostly in the brain, kidney,
and lung (1,2). Importantly, many in vitro and
in vivo studies have demonstrated that acute or chronic
cadmium exposure causes toxic and carcinogenic effects on various
tissues, including brain, liver, kidney, lung, prostate and testis
(3–6). However, the molecular and cellular
mechanisms underlying cadmium-mediated toxic and tumorigenic
effects on tissues still remain elusive.
Cyclooxygenase (COX), also referred to as
prostaglandin (PG) H synthase, is the rate-limiting enzyme in the
biosynthesis of PGs and related eicosanoids from arachidonic acid
metabolism (7). Physiologically,
levels of PGs are involved in the inflammatory response, bone
development, wound healing, and reproductive system. However,
excessive PGs are linked to many inflammatory and neoplastic
diseases. In eukaryotic cells, COX is expressed in two isoforms,
COX-1 and COX-2 (8,9). While COX-1 is constitutively
expressed in most cells and PGs produced by COX-1 are involved in
the maintenance of physiological functions, COX-2 is inducible by
pro-inflammatory cytokines, tumor promoters, mitogenes, and growth
factors in a variety of cell types, including monocytes,
fibroblasts and endothelial cells (8–10).
Evidence that non-steroidal anti-inflammatory drugs or compounds
that inhibit COX-2 lessen major inflammatory symptoms including
fever and pain suggests a role of COX-2 in inflammation (11). Notably, COX-2 expression has been
shown to be increased in glia of rats with traumatic brain injury
(12) and of a mouse model of
sporadic amyotrophic lateral sclerosis (13), suggesting a role of glial COX-2
expression in brain injury or neurodegenerative disease. Moreover,
of further interest, it has been demonstrated that COX-2 is
upregulated in the majority of high-grade human gliomas, and
blockage of COX-2 activity by NS-398, a selective COX-2 inhibitor,
inhibits growth and induces apoptosis of multiple glioma cells
(14). However, the regulation of
COX-2 expression in glioma is not well understood.
Expression of COX-2 is regulated at levels of
transcription, post-transcription and translation. Transcription of
COX-2 is induced in cells following treatment with exogenous
stimuli, which is accompanied by activation of intracellular
signaling pathways that in turn modulate activity of transcription
factors (TFs) and hence stimulate the COX-2 promoter (15). Studies have demonstrated the
presence of multiple cis-acting elements, including nuclear
factor-κB (NF-κB), activation of transcription factor-2 (ATF-2),
and activator protein 1 (AP-1), within the COX-2 promoter, which
are bound by specific TFs, and their importance for COX-2
transcriptional induction (16,17). Evidence also strongly suggests the
post-transcriptional regulation of COX-2 at levels of message
stability and RNA nuclear export and COX-2 translational regulation
by the protein turnover are critical for maximal COX-2 induction
(18–20). It is also documented that COX-2
protein N-glycosylation during translation is critical for the
protein stability, localization, and activity (21–23). Moreover, there is evidence that
activities of a variety of cellular signaling proteins, such as
extracellular signal-regulated protein kinase-1/2 (ERK-1/2), p38
mitogen-activated protein kinase (MAPK), c-Jun N-terminal protein
kinase-1/2 (JNK-1/2), and protein kinase B, are linked to
upregulation of COX-2 expression by influencing levels of COX-2
transcription, post-transcription, and/or translation (24,25). However, little is known concerning
the regulation of COX-2 expression by cadmium in glioma cells.
In this study, we investigated the effect of cadmium
on COX-2 expression in C6 rat glioma cells and determined possible
molecular and cellular mechanisms involved.
Materials and methods
Materials
Dulbecco’s modified Eagle’s medium, fetal bovine
serum (FBS), penicillin, and streptomycin were purchased from
WelGENE (Daegu, Korea). Antibodies of phospho-ERK-1/2 (p-ERK-1/2),
total ERK-1/2 (T-ERK-1/2), phospho-JNK-1/2 (p-JNK-1/2), total
JNK-1/2 (T-JNK-1/2), phospho-p38 MAPK (p-p38 MAPK), total p38 MAPK
(T-p38 MAPK) and phospho-ATF-2 (p-ATF-2) were purchased from Cell
Signaling Technology (Beverly, MA, USA). Anti-COX-2 rabbit
polyclonal antibody was purchased from Cayman Chemical (Ann Arbor,
MI, USA). Anti-rabbit or mouse secondary horseradish peroxidase
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). MG132, N-acetyl cysteine (NAC), SP600125,
and proteinase inhibitor cocktail (100X) were purchased from
Calbiochem (Madison, WI, USA). SB203580, PD98059 and LY294002 were
purchased from BioMol (Plymouth Meeting, PA, USA). RNAzol-B reagent
was purchased from Tel-Test, Inc. (Houston, TX, USA). Enzyme-linked
chemiluminescence (ECL) Western detection reagents were purchased
from Thermo Scientific (Waltham, MA, USA). Nitrocellulose membrane
was purchased from Millipore (Rockford, IL, USA). Bradford reagent
was purchased from Bio-Rad (Hercules, CA, USA). Other reagents,
including anti-actin mouse monoclonal antibody, cadmium
(CdSO4), glucosamine hydrochloride (GS-HCl), tunicamycin
(TN), cycloheximide (CHX), glutathione (GSH), vitamin E (VE),
cobalt chloride (CoCl2) and manganese (MnCl2)
were purchased from Sigma (St. Louis, MO, USA).
Cell culture
C6 rat glioma cells were maintained at 37°C in a
humidified condition of 95% air and 5% CO2 in DMEM
supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin,
and 100 μg/ml streptomycin.
Preparation of whole cell lysates
After treatments at the indicated times or
conditions, C6 cells were washed with ice-cold phosphate-buffered
saline (PBS) supplemented with 1 mM Na3VO4
and 1 mM NaF, and lysed in modified RIPA buffer [50 mM Tris-Cl (pH
7.4), 150 mM NaCl, 0.1% sodium dodecyl sulfate, 0.25% sodium
deoxycholate, 1% Triton X-100, 1% Nonidet P-40, 1 mM EDTA, 1 mM
EGTA, proteinase inhibitor cocktail (1X)]. After centrifugation at
12,000 rpm for 20 min at 4°C, the supernatant was collected and the
protein concentration was determined with Bradford reagent
(Bio-Rad, Mississauga, ON, USA) using bovine serum albumin as the
standard.
Western blot analysis
Equal amounts of protein (40 μg/lane) were
resolved by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto a nitrocellulose membrane. The
membrane was washed with Tris-buffered saline (TBS) (10 mM Tris,
150 mM NaCl) containing 0.05% Tween-20 (TBST) and blocked in TBST
supplemented with 5% non-fat dried milk. The membrane was further
incubated with primary antibodies of COX-2 (1:2,000), p-JNK-1/2
(1:2,000), T-JNK-1/2 (1:2,000), p-ERK-1/2 (1:2,000), T-ERK-1/2
(1:2,000), p-p38 MAPK (1:2,000), T-p38 MAPK (1:2,000), p-ATF-2
(1:2,000) or actin (1:10,000). The membrane was subsequently
incubated with appropriate secondary antibodies coupled to
horseradish peroxidase and developed in ECL Western detection
reagents.
Reverse transcription-polymerase chain
reaction (RT-PCR)
After treatments at the indicated times or
conditions, total cellular RNA was then isolated from the
conditioned cells using RNAzol B reagent according to manual
instructions provided by the manufacturer. Five micrograms of
total-RNA was reverse transcribed using 8 μl of M-MLV RT 5X
buffer, 3 μl of 10 mM dNTPs, 0.45 μl of 10,000 units
RNAse inhibitor, 0.3 μl of 50,000 units M-MLV reverse
transcriptase (Promega, Madison, WI), and 1.5 μl of 50 pM
oligo(dt) (Bioneer, Chungbuk, Korea) in 40 μl volume. Single
stranded cDNA was then amplified by PCR using 4 μl of 5X
Green GoTaq Flexi Buffer, 0.4 μl of 10 mM dNTPs, 0.1
μl of 500 units TaqDNA polymerase, 1.2 μl of 25 mM
MgCl2 (Promega), and 0.4 μl of each 20 pM of
specific sense and antisense primer of COX-2 and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The PCR products
were analyzed on 1.2% agarose gel. The primer sequences used by PCR
were as follows: COX-2, forward, 5′-CTG TAC TAC GCC GAG ATT CCT
GA-3′ and reverse, 5′-GTC CTC GCT TCT GAT CTG TCT TG-3′; GAPDH,
forward, 5′-GGT GAA GGT CGG TGT GAA CG-3′ and reverse, 5′-GGT AGG
AAC ACG GAA GGC CA-3′. The PCR conditions applied were: COX-2, 35
cycles of denaturation at 94°C for 30 sec, annealing at 56°C for 30
sec, and extension at 72°C for 30 sec; GAPDH, 27 cycles of
denaturation at 94°C for 30 sec, annealing at 57°C for 30 sec, and
extension at 72°C for 30 sec, respectively. GAPDH was used as an
internal control to evaluate the relative expression of COX-2.
Measurement of COX-2 protein
stability
C6 cells were then treated with cadmium alone or
cadmium plus MG132 for 2, 4 or 8 h in the absence or presence of
CHX, a translation inhibitor, to block ongoing translation. At each
time, whole cell lysate was prepared and subjected to immunoblot
analysis for COX-2 or actin to determine the amounts of each
protein remaining in the cells.
Results
Cadmium treatment increases not only
expression of COX-2 mRNA but also expression of both N-glycosylated
and unglycosylated COX-2 protein in C6 cells
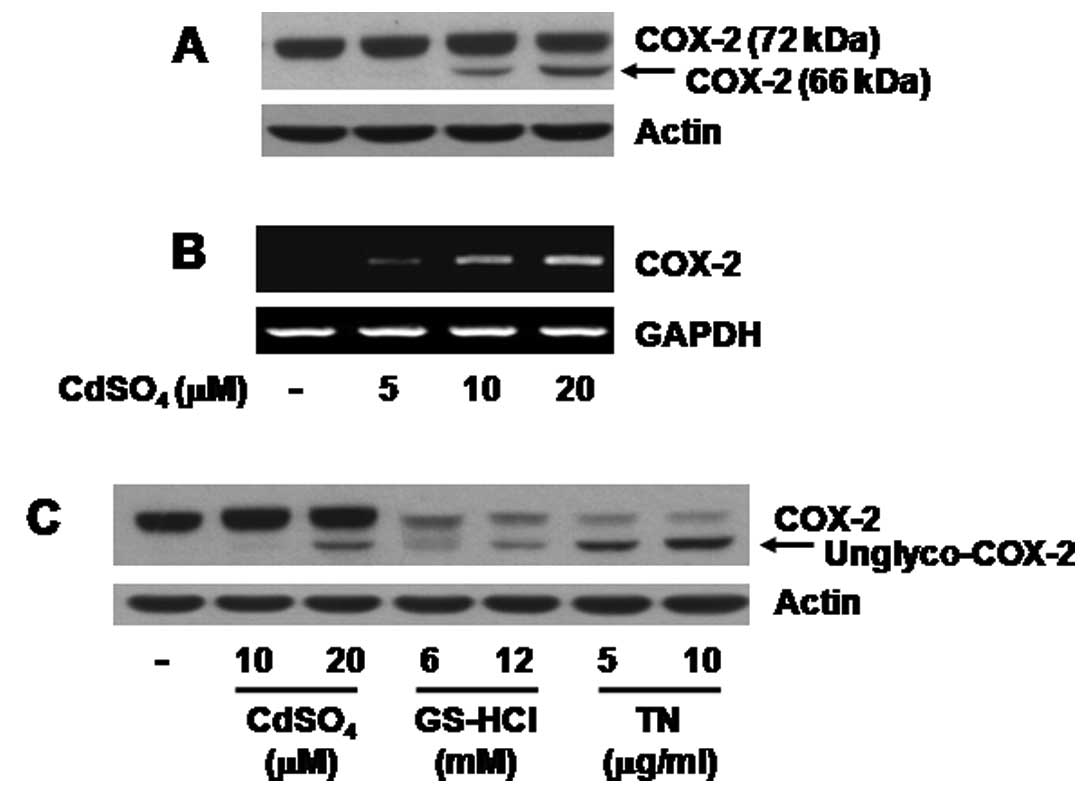
Initially, the effect of different concentrations of
cadmium on expression of COX-2 protein in C6 cells was analyzed by
western blotting. As shown in Fig.
1A (upper panel), treatment with cadmium at 5 μM for 8 h
did not induce expression of COX-2 protein while that with cadmium
at 10 or 20 μM increased expression of COX-2 proteins of 72
and 66 kDa molecular mass. RT-PCR experiments were next carried out
to evaluate the effect of cadmium on expression of COX-2 mRNA. As
shown in Fig. 1B (upper panel),
treatment with cadmium for 8 h increased expression of COX-2 mRNA
in a concentration-dependent manner. We next tested the effect of
cadmium on N-glycosylation of COX-2 in C6 cells. To accomplish
this, TN or GS-HCl that inhibits COX-2 N-glycosylation (23,26), was used as the positive control.
As shown in Fig. 1C (upper
panel), the size (66 kDa) of COX-2 induced by cadmium was nearly
identical to that of COX-2 induced by GS-HCl or TN in the C6 cells.
Expression of control actin protein or GAPDH mRNA remained
unchanged under these experimental conditions (Fig. 1A–C, lower panels). Due to the
strongest effect on expression of COX-2 proteins (72 and 66 kDa)
and COX-2 mRNA, we selected 20 μM of cadmium for further
study.
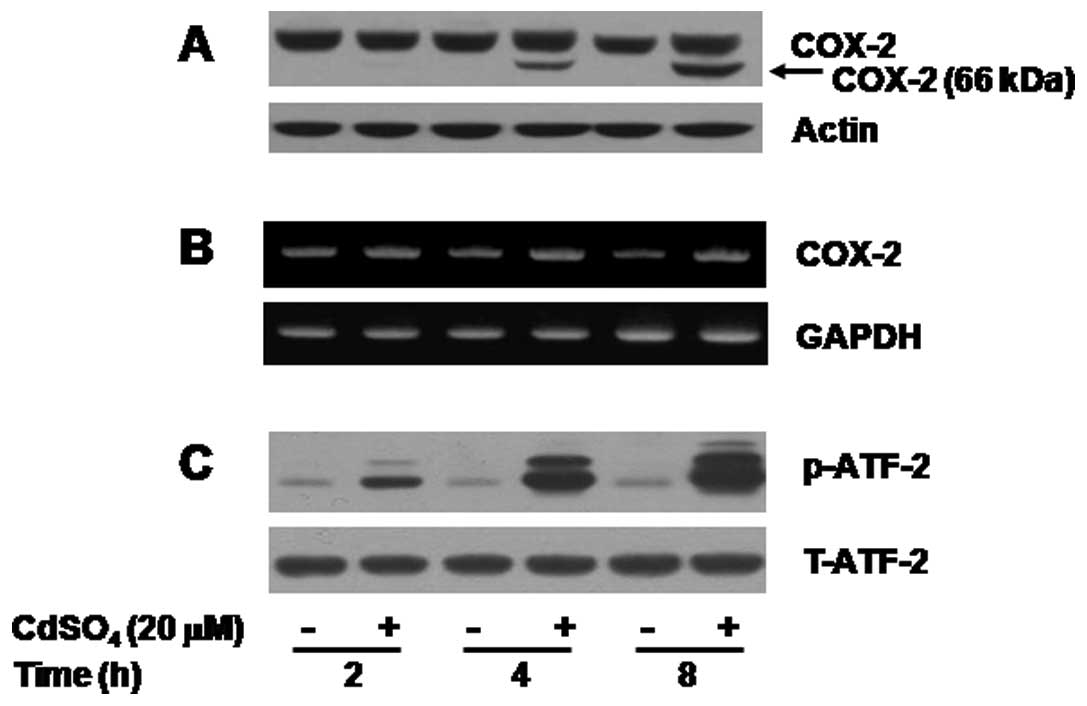
Kinetics of the expression and/or
activation of COX-2 and ATF-2 in the cadmium-treated C6 cells
Kinetic studies were next performed to determine the
time of induction of COX-2 proteins and mRNA by cadmium. Cadmium
treatment at 2 h did not modulate expression of COX-2 protein while
there was a time-dependent induction of both N-glycosylated and
unglycosylated COX-2 proteins at the cadmium treatment of 4 or 8 h
(Fig. 2A, upper panel). In case
of COX-2 mRNA, treatment with cadmium at 2 h slightly increased
expression of COX-2 mRNA (Fig.
2B, upper panel). There was a further enhancement of COX-2 mRNA
expression at 4 or 8 h. Expression of actin protein or GAPDH mRNA
remained unchanged during these experimental conditions (Fig. 2A and B, lower panels). The effect
of cadmium on activation of TFs, herein ATF-2, in C6 cells was next
investigated. Cadmium treatment led to a strong time-dependent
activation of ATF-2 in C6 cells (Fig.
2C, upper panel). Expression of total ATF-2 protein remained
unchanged during these experimental conditions (Fig. 2C, lower panel), suggesting that
cadmium treatment increases phosphorylation of the preexisting
ATF-2 protein without de novo protein synthesis.
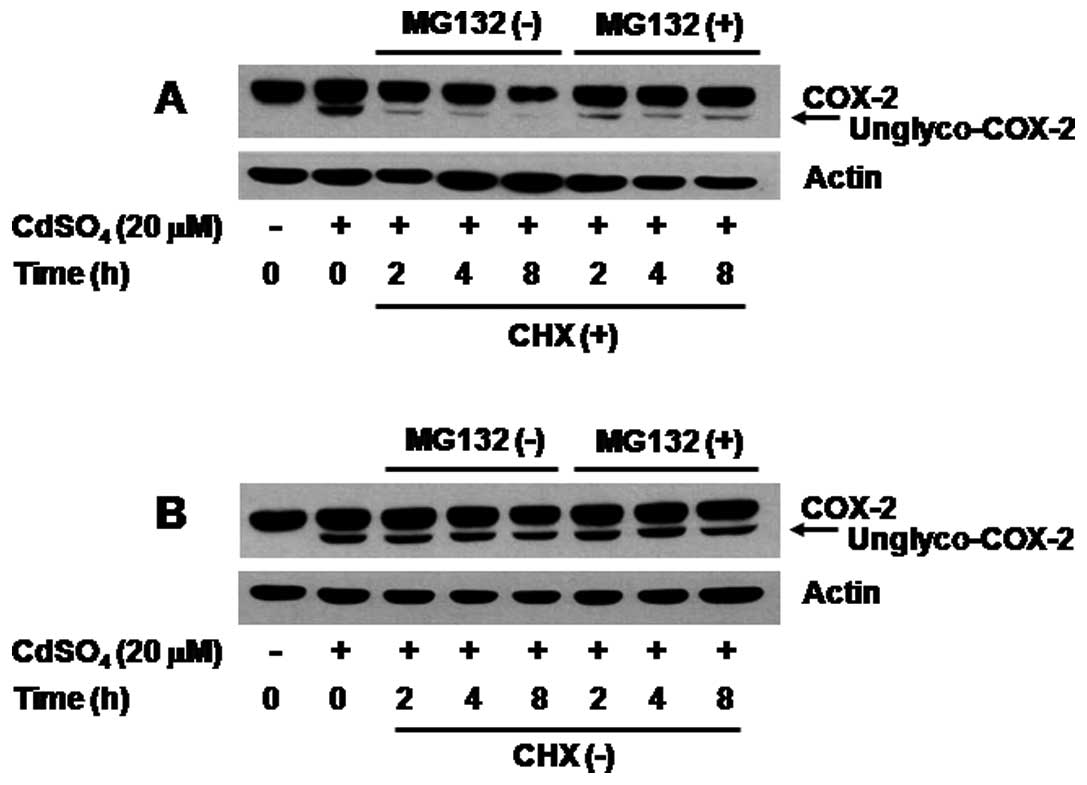
Stability of both N-glycosylated and
unglycosylated COX-2 proteins induced by cadmium in C6 cells is 26S
proteasome-dependent and the cadmium-induced expression of both
forms of COX-2 occurs in a translation-dependent manner
We next determined the stability of both
N-glycosylated and unglycosylated COX-2 proteins induced by cadmium
in C6 cells using protein stability assay with CHX, a protein
synthesis inhibitor. When translation was blocked in the presence
of CHX, the amount of both N-glycosylated and unglycosylated COX-2
proteins induced by cadmium was sharply decreased in a
time-dependent manner, suggesting that when translation is
inhibited, both forms of COX-2 induced by cadmium are rapidly
degraded (Fig. 3A, upper panel).
Importantly, the degradation was effectively blocked by treatment
with MG132, a 26S proteasomal inhibitor. However, when translation
was ongoing (no CHX), there was expression of both N-glycosylated
and unglycosylated COX-2 proteins induced by cadmium in C6 cells
that were grown in the absence or presence of MG132 (Fig. 3B, upper panel). Expression of
actin protein remained unchanged under these experimental
conditions (Fig. 3, lower
panels).
Cadmium-induced COX-2 transcriptional
upregulation in C6 cells is closely related to oxidative stress and
activation of the family of MAPKs
Previously, studies have shown the cadmium-mediated
expression of COX-2 through oxidative stress (27,28). This promptly led us to investigate
the role of oxidative stress in the cadmium-induced COX-2
expression in C6 cells using sulfhydryl group-containing reducing
agents (NAC, GSH and an antioxidant VE). Pretreatment with NAC or
GSH, but not with VE, strongly suppressed the cadmium-induced
expression of both N-glycosylated and unglycosylated COX-2 proteins
(Fig. 4A, upper panel). We
further determined whether intracellular signaling proteins
participate in the cadmium-induced COX-2 expression in C6 cells by
use of the specific pharmacological inhibitor of ERK-1/2 (PD98059,
PD), JNK-1/2 (SP600125, SP), p38 MAPK (SB203580, SB) or PI3K
(LY294002, LY). Pretreatment with PD98059, SP600125 or SB203580
effectively blocked the cadmium-induced expression of both forms of
COX-2 while that with LY294002 had little effect (Fig. 4C, upper panel). Results of RT-PCR
analyses, demonstrated that pretreatment with NAC, GSH, PD98059,
SP600125 or SB203580 suppressed the cadmium-induced COX-2 mRNA
expression (Fig. 4B and D, upper
panels). Expression of actin protein or GAPDH mRNA remained
unchanged under these experimental conditions (Fig. 4, lower panels).
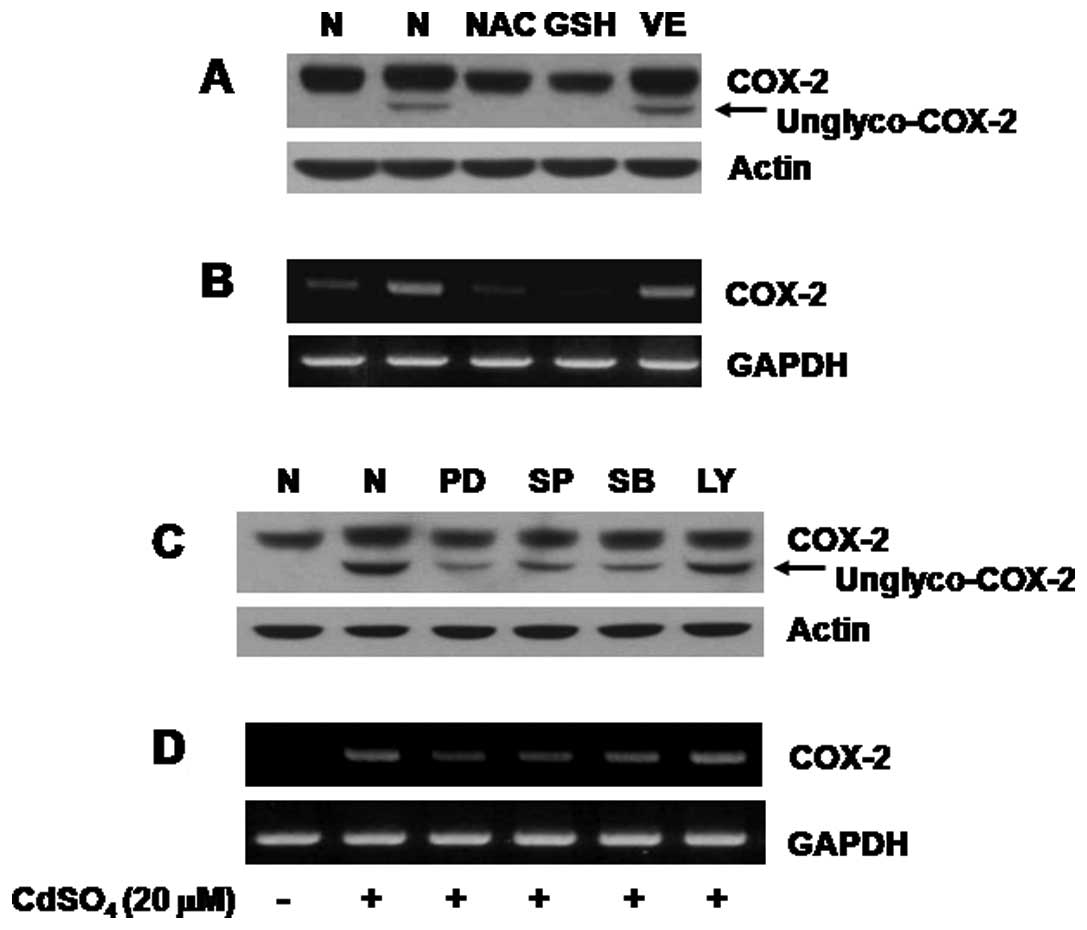 | Figure 4Effect of NAC, GSH or VE on
expression of COX-2 protein and mRNA in the cadmium-treated C6
cells. (A and B) C6 cells were pretreated without or with
N-acetyl-cysteine (NAC, 10 mM), glutathione (GSH, 10 mM) or vitamin
E (VE, 100 μM) for 1 h and then treated without or with
cadmium (20 μM) in the absence or presence of each agent for
an additional 8 h. Whole cell lysates were prepared and analyzed by
(A) western blot analysis and (B) RT-PCR, respectively. (C and D)
C6 cells were pretreated without or with PD98059 (PD, 50
μM), SP600125 (SP, 25 μM), SB203580 (SB, 25
μM) or LY294002 (LY, 25 μM) for 1 h and then treated
without or with cadmium (20 μM) in the absence or presence
of each inhibitor for an additional 8 h. Whole cell lysates were
prepared and analyzed by (C) western blot analysis and (D) RT-PCR,
respectively. Data are representative of three independent
experiments. |
Oxidative stress lies upstream of
activation of the family of MAPKs in the cadmium-treated C6
cells
We next investigated any link between oxidative
stress and activation of the family of MAPKs in the cadmium-treated
C6 cells. Cadmium treatment for 4 h increased phosphorylation of
ERK-1/2, p38 MAPK and JNK-1/2 in C6 cells (Fig. 5, upper panels). However, cadmium
treatment did not affect total expression of each member of the
MAPKs in C6 cells, suggesting that cadmium treatment also enhances
phosphorylation of the preexisting MAPKs in C6 cells without de
novo protein synthesis (Fig.
5, lower panels). Interestingly, pretreatment with NAC or GSH,
but not VE, strongly interfered with the cadmium-mediated
phosphorylation of ERK-1/2, p38 MAPK and JNK-1/2 in C6 cells.
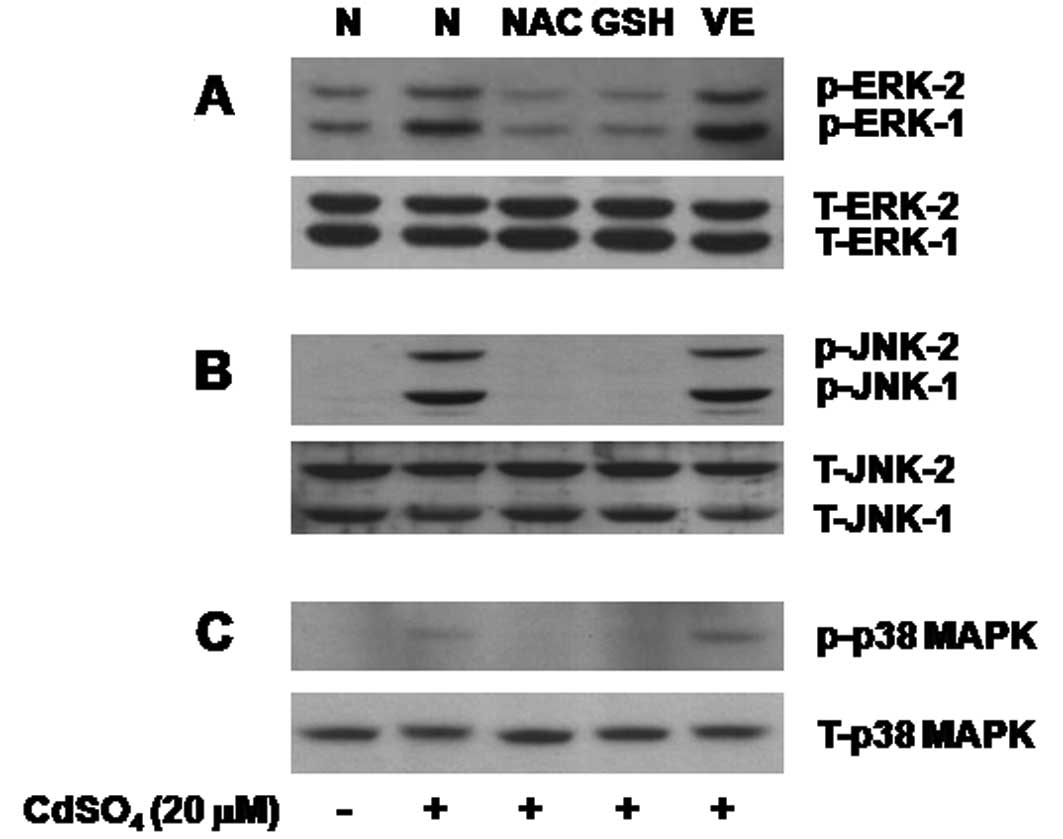 | Figure 5Effect of NAC, GSH, or VE on
activation of ERK-1/2, JNK-1/2, and p38 MAPK in the cadmium-treated
C6 cells. (A–C) C6 cells were pretreated without or with
N-acetyl-cysteine (NAC, 10 mM), glutathione (GSH, 10 mM) or vitamin
E (VE, 100 μM) for 1 h and then treated without or with
cadmium (20 μM) in the absence or presence of each agent for
an additional 8 h. Whole cell lysates were prepared and analyzed by
western blot analysis. Data are representative of three independent
experiments. p-ERK-1/2, phosphorylated ERK-1/2; T-ERK-1/2, total
ERK-1/2; p-JNK-1/2, phosphorylated JNK-1/2; T-JNK-1/2, total
JNK-1/2; p-p38 MAPK, phosphorylated p38 MAPK; T-p38 MAPK, total p38
MAPK. |
Cadmium specifically induces expression
of both N-glycosylated and unglycosylated forms of COX-2 in C6
cells
To evaluate the specificity, C6 cells were treated
with cadmium or different heavy metals (cobalt or manganese) and
expression of COX-2 in the conditioned cells was measured.
Treatment with cadmium (10 or 20 μM) induced expression of
both N-glycosylated and unglycosylated COX-2 proteins, while there
was no induction of COX-2 by treatment with cobalt (5, 10 or 20
μM) or manganese (250 or 500 μM) in C6 cells
(Fig. 6, upper panel). Expression
of actin protein remained unchanged under these experimental
conditions (Fig. 6, lower
panel).
Discussion
Cadmium is a toxic heavy metal and an environmental
hazard to humans. Deregulated expression of COX-2 is linked to many
neuronal diseases, including inflammation and cancer (29,30). In this study, we investigated the
effect of cadmium on expression of COX-2 in C6 rat glioma
cells.
Previously, the cadmium-mediated upregulation of
COX-2 protein expression was shown (27,31). COX-2 is an N-glycoprotein and has
four glycosylation sites (asparagine) within the structure
(21,22). In general, protein N-glycosylation
occurs during translation and is important for protein stability,
activity, and/or cellular localization (32). Based on this fact, it was
previously shown that exposure to GS-HCl or TN, inhibitors of
protein N-glycosylation, prevents COX-2 N-glycosylation leading to
generation of the unglycosylated COX-2 with decreased molecular
mass and enzymatic activity (23,26). In this study, cadmium induced
expression not only of the normally expressed COX-2 of 72 kDa
(N-glycosylated COX-2) but also the unglycosylated COX-2 of 66 kDa
in C6 cells (Fig. 1A), as
assessed by the unglycosylated COX-2 induced by GS-HCl or TN
(Fig. 1C). However, considering
the magnitude on expression of the unglycosylated COX-2 induced by
cadmium and the known inhibitor of COX-2 N-glycosylation, it seems
that cadmium only partially interferes with N-glycosylation of
COX-2 in C6 cells. Moreover, taking into account a previous study
that cadmium perturbs COX-2 N-glycosylation in H4 neuronal cells
(27), it is unlikely that the
cadmium-mediated expression of the unglycosylated COX-2 is limited
to C6 cells. The regulation of COX-2 turnover is an important
contributor to COX-2 expression. The t1/2 of COX-2
protein in cells is suggested to be within hours. There is evidence
suggesting a 26S proteasome-dependent COX-2 protein turnover
(33). Notably, data of CHX-based
protein stability assays herein demonstrated that both
N-glycosylated and unglycosylated COX-2 proteins induced by cadmium
are rapidly degraded while their degradation is effectively blocked
by MG132 in the presence of CHX (Fig.
3A), which suggests that both forms of COX-2 induced by cadmium
are labile and the proteasomal pathway is involved in their
turnover. In addition, the present study further demonstrated that
cadmium can induce and maintain expression of the unglycosylated
COX-2 protein in C6 cells in the absence of CHX (Fig. 3B), implying that cadmium induces
expression of the unglycosylated COX-2 protein by targeting
N-glycosylation of nascent COX-2 polypeptide. To the best of our
knowledge, this is the first report of the proteasome- and
translation-dependent regulation of the expression of the
N-glycosylated and unglycosylated COX-2 proteins by cadmium.
Our data also indicate cadmium-induced COX-2
transcriptional upregulation in C6 cells (Figs. 1B and 2B). COX-2 transcriptional induction was
found to be largely affected by activities of TFs (7,10,16,17) and
signaling proteins (34,35). Previous results from our
laboratory demonstrated that cadmium does not induce activation of
NF-κB but triggers activation of ERK-1/2, p38 MAPK, and JNK-1/2 in
C6 cells (36). In this study, we
demonstrated for the first time that cadmium has the ability to
rapidly induce activation of ATF-2 in C6 cells (Fig. 2C). In a recent study, the
activation of p38 MAPK, but not JNK-1/2, was crucial for the
cadmium-induced COX-2 expression in H4 cells (27). However, the present study showed
that cadmium-induced COX-2 expression in C6 cells was dependent on
the activation not only of p38 MAPK but also ERK-1/2 and JNK-1/2
(Fig. 4C and D). There is
evidence suggesting that p38 MAPK and/or JNK-1/2 regulate
activation of ATF-2 (37,38) and ERK-1/2 and/or JNK-1/2 are
involved in AP-1 activation (39,40). A recent finding revealed the
effect of cadmium on the rapid upregulation of c-fos and c-jun, two
key components of the AP-1 transcription factor, in HPT human
proximal tubule cells of the kidney (41). Although speculative, it is thus
likely that the cadmium-induced COX-2 transcriptional upregulation
in C6 cells is in part mediated through the p38 MAPK/JNK-1/2-ATF-2
and ERK-1/2/JNK-1/2-AP-1 pathways.
Early studies indicated a link between oxidative
stress and COX-2 expression (42,43). Oxidative stress often occurs due
to an imbalance in intracellular levels of oxidants (e.g., ROS) and
reducing agents (e.g., GSH). Previous studies demonstrated a
cadmium-mediated oxidative stress by decreasing cellular GSH and/or
increasing cellular ROS (44,45). Supporting these findings, we
previously demonstrated that cadmium rapidly lowers cellular GSH
and the GSH reduction is important for the metal-mediated
expression of MAPK phosphatase-1 in C6 cells (36). In this study, we further showed
that oxidative stress probably linked to a reduction in cellular
GSH is critical for COX-2 transcriptional upregulation by cadmium
in C6 cells (Fig. 4A and B). It
is known that oxidative stress lies upstream of the activation of
TFs and/or signaling proteins in cadmium-induced COX-2 expression
and apoptotic death of neuronal cells (28). The present study also suggests
crosstalk between oxidative stress and the activation of the family
of MAPKs in cadmium-treated C6 cells (Fig. 5), in which cadmium treatment
primarily lowers cellular GSH, which subsequently leads to
activation of p38 MAPK, ERK-1/2 and JNK-1/2.
The specificity of cadmium to induce expression of
both N-glycosylated and unglycosylated forms of COX-2 in C6 cells
was also noted. This notion is based on the absence of induction of
COX-2 protein in C6 cells exposed to other heavy metals, such as
cobalt or manganese (Fig. 6).
However, previous reports have demonstrated cobalt- or
manganese-mediated upregulation of COX-2 expression in various cell
types, in which expression of COX-2 protein was increased in vector
or phospholipase D cDNA-transfected U87 MG human astroglioma cells
treated with cobalt (200 μM, 20 h) (46) or in PC3 human prostate cancer
cells exposed to cobalt (100 μM, 12 h) (47). Strong COX-2 protein expression was
also observed in cultured astrocytes treated with manganese (100
μM, 16 h) (48) and in
A549 human airway cells exposed to manganese (100 μM, 16 h)
(25). It is plausible that the
differential effect on COX-2 expression by cobalt or manganese in
the present study and previous reports may be due to the use of
differential experimental systems, including cDNA transfection,
treatment concentration and time of each metal and/or different
type of cells used.
It is now obvious that cadmium induces expression of
both N-glycosylated and unglycosylated COX-2 in C6 cells. However,
the biological role (or relevance) of the cadmium-mediated
expression of both forms of COX-2 in C6 cells remains unclear at
this moment. However, assuming that COX-2 expression (and activity)
contributes to the growth of glioma (14) and is linked to neuroinflammation
(30), it is conceivable that the
N-glycosylated COX-2 induced by cadmium may contribute to C6 cell
growth and/or induce a pro-inflammatory response. It is well
documented that inhibition of protein N-glycosylation leads to
production of misfolded and/or nonfunctional proteins (32) and cellular accumulation of such
damaged proteins elicits ER stress (49,50). Thus, it is suggested that
accumulation of the unglycosylated proteins (herein COX-2) induced
by cadmium may contribute to ER stress in C6 cells.
In conclusion, we demonstrate for the first time
that cadmium induces expression of COX-2 through both
transcriptional and co-translational (N-glycosylation) regulation
in C6 cells in which the cadmium-induced COX-2 transcriptional
upregulation is closely related to oxidative stress-dependent
activation of the family of MAPKs and the cadmium-induced
expression of both N-glycosylated and unglycosylated COX-2 proteins
occurs in the proteasome- and/or translation-dependent control.
Acknowledgements
The present study was supported by the
Bisa Research Grant of Keimyung University in 2010.
References
|
1.
|
L GerhardssonV EnglystNG LundströmCadmium,
copper and zinc in tissues of deceased copper smelter workersJ
Trace Elem Med
Biol16261266200210.1016/S0946-672X(02)80055-412530590
|
|
2.
|
S SatarugSH GarrettMA SensDA SensCadmium,
environmental exposure, and health outcomesEnviron Health
Perspect118182190201010.1289/ehp.0901234
|
|
3.
|
G BertinD AverbeckCadmium: cellular
effects, modifications of biomolecules, modulation of DNA repair
and genotoxic consequences
(Review)Biochimie8815491559200610.1016/j.biochi.2006.10.00117070979
|
|
4.
|
JY ImSG PaikPL HanCadmium-induced
astroglial death proceeds via glutathione depletionJ Neurosci
Res83301308200610.1002/jnr.2072216385582
|
|
5.
|
J LiuW QuMB KadiiskaRole of oxidative
stress in cadmium toxicity and carcinogenesisToxicol Appl
Pharmacol238209214200910.1016/j.taap.2009.01.02919236887
|
|
6.
|
JM MatésJA SeguraFJ AlonsoJ MárquezRoles
of dioxins and heavy metals in cancer and neurological diseases
using ROS-mediated mechanismsFree Radic Biol
Med4913281341201020696237
|
|
7.
|
WL SmithDL DeWittRM
GaravitoCyclooxygenases: structural, cellular, and molecular
biologyAnnu Rev
Biochem69145182200010.1146/annurev.biochem.69.1.14510966456
|
|
8.
|
WL SmithDL DewittProstaglandin
endoperoxide H synthases-1 and -2Adv
Immunol62167215199610.1016/S0065-2776(08)60430-7
|
|
9.
|
JR VaneYS BakhleRM BottingCyclooxygenases
1 and 2Annu Rev Pharmacol
Toxicol3897120199810.1146/annurev.pharmtox.38.1.97
|
|
10.
|
WL SmithRM GaravitoDL DeWittProstaglandin
endoperoxide H synthases (cyclooxygenases)-1 and -2J Biol
Chem2713315733160199610.1074/jbc.271.52.331578969167
|
|
11.
|
GW WilliamsAn update on nonsteroidal
anti-inflammatory drugs and cyclooxygenase-2 inhibitorsCurr Pain
Headache Rep9377389200510.1007/s11916-005-0017-416282038
|
|
12.
|
KI StraussMF BarbeRM MarshallProlonged
cyclooxygenase-2 induction in neurons and glia following traumatic
brain injury in the ratJ
Neurotrauma17695711200010.1089/08977150041543610972245
|
|
13.
|
C MaihöfnerS Probst-CousinM
BergmannExpression and localization of cyclooxygenase-1 and -2 in
human sporadic amyotrophic lateral sclerosisEur J
Neurosci1815271534200314511332
|
|
14.
|
T JokiO HeeseDC NikasExpression of
cyclooxygenase 2 (COX-2) in human glioma and in vitro inhibition by
a specific COX-2 inhibitor, NS-398Cancer
Res6049264931200010987308
|
|
15.
|
HR HerschmanST ReddyW XieFunction and
regulation of prostaglandin synthase-2Adv Exp Med
Biol4076166199710.1007/978-1-4899-1813-0_99321932
|
|
16.
|
R NewtonLM KuitertM BergmannEvidence for
involvement of NF-kappaB in the transcriptional control of COX-2
gene expression by IL-1betaBiochem Biophys Res
Commun2372832199710.1006/bbrc.1997.70649266823
|
|
17.
|
H InoueC YokoyamaS HaraTranscriptional
regulation of human prostaglandin-endoperoxide synthase-2 gene by
lipopolysaccharide and phorbol ester in vascular endothelial cells.
Involvement of both nuclear factor for interleukin-6 expression
site and cAMP response elementJ Biol
Chem2702496524971199510.1074/jbc.270.42.24965
|
|
18.
|
A RistimäkiS GarfinkelJ
WessendorfInduction of cyclooxygenase-2 by interleukin-1 alpha.
Evidence for post-transcriptional regulationJ Biol
Chem269117691177519948163473
|
|
19.
|
SK SrivastavaT TetsukaD Daphna-IkenAR
MorrisonIL-1 beta stabilizes COX II mRNA in renal mesangial cells:
role of 3′-untranslated regionAm J
Physiol267F504F50819948092265
|
|
20.
|
BC JangU Muñoz-NajarJH PaikLeptomycin B,
an inhibitor of the nuclear export receptor CRM1, inhibits COX-2
expressionJ Biol
Chem27827732776200310.1074/jbc.C20062020012468543
|
|
21.
|
JF NemethGP Hochgesang JrLJ MarnettRM
CaprioliCharacterization of the glycosylation sites in
cyclooxygenase-2 using mass
spectrometryBiochemistry4031093116200110.1021/bi002313c11258925
|
|
22.
|
T HlaK NeilsonHuman cyclooxygenase-2
cDNAProc Natl Acad Sci
USA8973847388199210.1073/pnas.89.16.73841380156
|
|
23.
|
JC OttoDL DeWittWL SmithN-glycosylation of
prostaglandin endoperoxide synthases-1 and -2 and their
orientations in the endoplasmic reticulumJ Biol
Chem268182341824219938349699
|
|
24.
|
WH TungHL HsiehCM YangEnterovirus 71
induces COX-2 expression via MAPKs, NF-kappaB, and AP-1 in SK-N-SH
cells: Role of PGE(2) in viral replicationCell
Signal22234246201010.1016/j.cellsig.2009.09.01819800403
|
|
25.
|
BC JangInduction of COX-2 in human airway
cells by manganese: role of PI3K/PKB, p38 MAPK, PKCs, Src, and
glutathione depletionToxicol In
Vitro23120126200910.1016/j.tiv.2008.11.00519084589
|
|
26.
|
BC JangSH SungJG ParkGlucosamine
hydrochloride specifically inhibits COX-2 by preventing COX-2
N-glycosylation and by increasing COX-2 protein turnover in a
proteasome-dependent mannerJ Biol
Chem2822762227632200710.1074/jbc.M610778200
|
|
27.
|
ME Figueiredo-PereiraZ LiM JansenP
RockwellN-acetylcysteine and celecoxib lessen cadmium cytotoxicity
which is associated with cyclooxygenase-2 upregulation in mouse
neuronal cellsJ Biol
Chem2772528325289200210.1074/jbc.M10914520011997384
|
|
28.
|
P RockwellJ MartinezL PapaE GomesRedox
regulates COX-2 upregulation and cell death in the neuronal
response to cadmiumCell
Signal16343353200410.1016/j.cellsig.2003.08.00614687664
|
|
29.
|
A HaraI OkayasuCyclooxygenase-2 and
inducible nitric oxide synthase expression in human astrocytic
gliomas: correlation with angiogenesis and prognostic
significanceActa
Neuropathol1084348200410.1007/s00401-004-0860-015088099
|
|
30.
|
SH ChoiS AidF BosettiThe distinct roles of
cyclooxygenase-1 and -2 in neuroinflammation: implications for
translational researchTrends Pharmacol
Sci30174181200910.1016/j.tips.2009.01.00219269697
|
|
31.
|
CS ParkOS KimSM YunPresenilin
1/gamma-secretase is associated with cadmium-induced E-cadherin
cleavage and COX-2 gene expression in T47D breast cancer
cellsToxicol Sci106413422200810.1093/toxsci/kfn19718791180
|
|
32.
|
J RothC ZuberS ParkProtein
N-glycosylation, protein folding, and protein quality controlMol
Cells30497506201010.1007/s10059-010-0159-z21340671
|
|
33.
|
M SinghP ChaudhryS ParentE
AsselinUbiquitinproteasomal degradation of COX-2 in TGF-β
stimulated human endometrial cells is mediated through endoplasmic
reticulum mannosidase IEndocrinology153426437201222109885
|
|
34.
|
H PhamR VincentiLW SliceCOX-2 promoter
activation by AT1R-Gq-PAK-p38beta signaling in intestinal
epithelial cellsBiochim Biophys
Acta1779408413200810.1016/j.bbagrm.2008.05.00418515110
|
|
35.
|
ZR HealyF ZhuJD StullK
KonstantopoulosElucidation of the signaling network of COX-2
induction in sheared chondrocytes: COX-2 is induced via a
Rac/MEKK1/MKK7/JNK2/c-Jun-C/EBPbeta-dependent pathwayAm J Physiol
Cell Physiol294C1146C1157200810.1152/ajpcell.00542.200718367585
|
|
36.
|
SM KimJG ParkWK BaekCadmium specifically
induces MKP-1 expression via the glutathione depletion-mediated p38
MAPK activation in C6 glioma cellsNeurosci
Lett440289293200810.1016/j.neulet.2008.05.06418573614
|
|
37.
|
KC ChenYL ChiouLS ChangJNK1/c-Jun and p38
alpha MAPK/ATF-2 pathways are responsible for upregulation of
Fas/FasL in human chronic myeloid leukemia K562 cells upon exposure
to Taiwan cobra phospholipase A2J Cell
Biochem108612620200910.1002/jcb.2229319670268
|
|
38.
|
N TindbergM Porsmyr-PalmertzA
SimiContribution of MAP kinase pathways to the activation of ATF-2
in human neuroblastoma cellsNeurochem
Res25527531200010.1023/A:100752031145710823586
|
|
39.
|
BY HoYM WuKJ ChangTM PanDimerumic acid
inhibits SW620 cell invasion by attenuating
H2O2-mediated MMP-7 expression via JNK/C-Jun
and ERK/C-Fos activation in an AP-1-dependent mannerInt J Biol
Sci7869880201121814482
|
|
40.
|
J DingY HuangB NingTNF-alpha induction by
nickel compounds is specific through ERKs/AP-1-dependent pathway in
human bronchial epithelial cellsCurr Cancer Drug
Targets918190200910.2174/15680090978731399519200052
|
|
41.
|
SH GarrettV PhillipsS SomjiTransient
induction of metallothionein isoform 3 (MT-3), c-fos, c-jun and
c-myc in human proximal tubule cells exposed to cadmiumToxicol
Lett1266980200210.1016/S0378-4274(01)00448-911738272
|
|
42.
|
C YangH LingM ZhangOxidative stress
mediates chemical hypoxia-induced injury and inflammation by
activating NF-κb-COX-2 pathway in HaCaT cellsMol
Cells31531538201121533553
|
|
43.
|
JH YoonTG LimKM LeeTangeretin reduces
ultra-violet B (UVB)-induced cyclooxygenase-2 expression in mouse
epidermal cells by blocking mitogen-activated protein kinase (MAPK)
activation and reactive oxygen species (ROS) generationJ Agric Food
Chem59222228201110.1021/jf103204x
|
|
44.
|
MS YangD LiT LinIncrease in intracellular
free/bound NAD[P]H as a cause of Cd-induced oxidative stress in the
HepG(2) cellsToxicology2476102008
|
|
45.
|
N PathakS KhandelwalInfluence of cadmium
on murine thymocytes: potentiation of apoptosis and oxidative
stressToxicol
Lett165121132200610.1016/j.toxlet.2006.02.00416563667
|
|
46.
|
BH AhnMH ParkYH LeeUpregulation of
cyclooxygenase-2 by cobalt chloride-induced hypoxia is mediated by
phospholipase D isozymes in human astroglioma cellsBiochim Biophys
Acta177317211731200710.1016/j.bbamcr.2007.06.00117640750
|
|
47.
|
XH LiuA KirschenbaumS YaoUpregulation of
vascular endothelial growth factor by cobalt chloride-simulated
hypoxia is mediated by persistent induction of cyclooxygenase-2 in
a metastatic human prostate cancer cell lineClin Exp
Metastasis17687694199910.1023/A:1006728119549
|
|
48.
|
SL LiaoYC OuSY ChenInduction of
cyclooxygenase-2 expression by manganese in cultured
astrocytesNeurochem
Int50905915200710.1016/j.neuint.2006.09.01617084486
|
|
49.
|
M HongS LuoP BaumeisterUnderglycosylation
of ATF6 as a novel sensing mechanism for activation of the unfolded
protein responseJ Biol
Chem2791135411363200410.1074/jbc.M30980420014699159
|
|
50.
|
SM YuSJ KimEndoplasmic reticulum stress
(ER-stress) by 2-deoxy-D-glucose (2DG) reduces cyclooxygenase-2
(COX-2) expression and N-glycosylation and induces a loss of COX-2
activity via a Src kinase-dependent pathway in rabbit articular
chondrocytesExp Mol Med42777786201010.3858/emm.2010.42.11.079
|