Introduction
The molecular mechanisms underlying the various
‘two-hit models’ of injury have been subject to intensive
investigations in various models of critical illness (1–7).
While a slight degree/shorter period or repeated short periods of
hypoxia can protect cells or tissues from a subsequent, more severe
form of injury (‘ischemic preconditioning’), more severe degrees of
pre-existing injuries can make the organism ultrasensitive to a
second insult in the paradigm of various forms of ‘two-hit
models’.
In the present study, we established an in
vitro model of two-hit injury involving pre-exposure to
hypoxia, followed by a second challenge induced by oxidative stress
in cultured human lung epithelial cells. Due to the role of the
nuclear enzyme poly(ADP-ribose) polymerase-1 (PARP-1) in the
pathogenesis of various diseases associated with oxidative or
nitrative stress (8–10), we also investigated the potential
contribution of the activation of PARP-1 (the major PARP isoform)
to the cell injury associated with the ‘two-hit’ response.
Materials and methods
Reagents
Unless specified otherwise, all the reagents were
purchased from Sigma-Aldrich Co. (St. Louis, MO, USA).
Cell culture
The A549 human lung adenocarcinoma cell line was
grown in RPMI-1640 medium containing 10% fetal bovine serum (FBS;
PAA Laboratories, Dartmouth, MA, USA) 100 U/ml penicillin and 100
μg/ml streptomycin (Invitrogen, Carlsbad, CA, USA) at 37°C, 5%
CO2. Stable gene silencing of PARP-1 with lentiviral
infection (yielding shPARP-1 cells) was performed as previously
described (11), i.e., control
cells were subjected to an identical procedure except that they
were transfected with a non-coding silencing vector (11).
In vitro model of hypoxia
Cell culture plates were placed in gas-tight
incubation chambers (Billups-Rothenberg Inc., Del Mar, CA, USA) and
the chamber atmosphere was replaced by flushing the chamber with
95% N2/5% CO2 mixture at a flow rate of 25
l/min for 5 min. Hypoxia was maintained by clamping and the
chambers were incubated for 24 h at 37°C, as described in a
previous study (12). Following
hypoxia, cells were incubated for the indicated period at 37°C in a
5% CO2 atmosphere in the presence or absence of various
concentrations of hydrogen peroxide.
Measurement of cellular glutathione (GSH)
content
Total cellular GSH content was measured using the
OxiSelect™ Total Glutathione (GSSG/GSH) assay kit (Cell Biolabs,
Inc., San Diego, CA, USA), as previously described (13). This kit provided the enzyme
glutathione reductase, which reduced oxidized glutathione (GSSG) to
reduced GSH in the presence of NADPH. Subsequently, a chromogen
reacted with the thiol group of GSH to produce a colored compound
that was absorbed at 405 nm. The rate of chromophore production was
proportional to the concentration of GSH within the sample. The
rate was determined from the absorbance change over time.
Single-cell electrophoresis (comet
assay)
Broken DNA was allowed to unwind under alkaline
conditions and formed comet-like structures after cell lysis and
electrophoresis. Following trypsinization, single-cell suspension
was treated with hydrogen peroxide for 10 min. Single-stranded DNA
breaks were assayed by comet assay using a CometAssay kit
(Trevigen, Gaithersburg, MA, USA), as previously described
(14). Single cells were exposed
to an electric field in agarose gel and stained with SYBR-Green.
Labeled DNA was then visualized under fluorescence microscopy.
Measurement of intracellular adenosine
triphosphate (ATP) levels
To measure intracellular ATP levels, a CellTiter-Glo
Luminescent Cell Viability assay (Promega, Madison, WI, USA) based
on ATP requiring luciferin-oxyluciferin conversion mediated by a
thermostable luciferase generating a stable ‘glow-type’ luminescent
signal was used. The cells were lysed in 100 μl of CellTiter-Glo
reagent according to the manufacturer’s instructions and the
luminescent signal was recorded for 1 sec on a high sensitivity
luminometer (Synergy 2, Biotek, Winooski, VT, USA) (15). Changes in ATP concentration were
calculated as a percentage of the untreated control.
Western blot analysis
Western blot analysis for poly(ADP-ribose) (PAR)
polymers was conducted as previously described (11). Cells were washed once with
phosphate buffered-saline (PBS) and collected by scraping into 200
μl ice-cold lysis buffer containing 62.5 mM Tris-HCl (pH 6.8), 2%
SDS, 10% glycerol, 1 mM PMSF, and protease inhibitors. The extracts
were sonicated, and the supernatants were collected following
centrifugation. The protein concentration was determined by BCA
Protein assay (Thermo Scientific, Rockford, IL, USA). Prior to
boiling, samples were digested with 50 mM dithiothreitol (DTT) and
0.1% bromophenol blue. Protein (20 μg) was loaded onto 8%
polyacrylamide gels. Proteins were separated by electrophoresis and
then transferred to nitrocellulose membrane. For immunoblotting,
membranes were blocked with 5% non-fat milk in Tris-buffered saline
(TBS) for 60 min. Primary antibody against poly(ADP-ribose)
polymers (Trevigen, Gaithersburg, MA, USA) or against PARP-1 (Cell
Signaling Technology, Inc., Danvers, MA, USA) were applied at
1,000-fold of dilution in blocking buffer, overnight at 4°C. After
washing 3 times in TBS containing 0.2% Tween-20 (TBST), secondary
antibody (peroxidase-conjugated goat anti-rabbit) and
peroxidase-conjugated anti-actin (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) were applied at 4,000-fold of dilution in
blocking buffer for 1 h at room temperature. Blots were washed 3
times in TBST, once in TBS and were then incubated in enhanced
chemiluminescence substrate and Supersignal West Pico
Chemiluminescent substrate (Thermo Scientific), and exposed to
photographic film.
MTT mitochondrial activity assay
The MTT assay was performed as previously described
(16). Briefly, cells were
treated after normoxia or hypoxia with hydrogen peroxide in 96-well
plates. Then, 24 h later
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT, Calbiochem, EMD BioSciences, San Diego, CA, USA) was added to
the cells (0.5 mg/ml) for an additional hour. The medium was then
aspirated and the formazan crystals were dissolved by the addition
of 100 μl isopropanol. Optical density was detected on a Synergy 2
reader (BioTek Instruments, Inc., Winooski, VT, USA) at 570 nm with
background measurement at 690 nm. Results are shown as percentage
compared to the untreated control.
Lactate dehydrogenase (LDH) assay
LDH release was measured as previously described
(17). Briefly, cell culture
supernatant (30 μl) was mixed with 100 μl freshly prepared LDH
assay reagent and the changes in absorbance were read kinetically.
LDH release values are shown as Vmax (mOD/min).
Measurement of plasma membrane
integrity
Plasma membrane integrity was measured by propidium
iodide (PI) uptake as described in a previous study (18). Briefly, the cells were stained
with 5 μg/ml PI for 15 min. Detached and trypsinized cells were
then collected, washed once with PBS and analyzed by the Guava
Easycyte Plus flow cytometry system (Millipore, Billerica, MA,
USA).
Experimental protocols
Control cells and cells with stably silenced PARP-1
(shPARP-1) were exposed to 24 h of hypoxia followed by the
determination of cellular GSH levels, as well as quantification of
PARP-1 expression (by western blotting). At 24 h, the cells were
subjected to oxidative stress with hydrogen peroxide. Ten minutes
later, DNA damage was assessed by the comet assay and PARP
activation was assessed by western blotting for PAR polymers. At 24
h after the H2O2 challenge, cell viability
was detected by the MTT and LDH assays, and by PI uptake, then
cellular ATP levels were quantified (Fig. 1A).
Statistical analysis
Data were shown as the means ± SEM. One-way ANOVA
was applied for statistical analysis, while the Tukey’s post hoc
test was used for the determination of significance. P<0.0.5 was
considered statistically significant. Statistical calculations were
performed using Graphpad Prism 5 analysis software. Experiments
were performed at least 3 times on different days.
Results
Hypoxia reduced the cellular GSH content
without affecting the expression of PARP-1 protein
Hypoxia significantly reduced the intracellular GSH
levels to 37±2% of normoxic values in wild-type cells and to a
smaller degree (to 46±2% of normoxic values) in the shPARP-1 cells
(Fig. 1B). The expression level
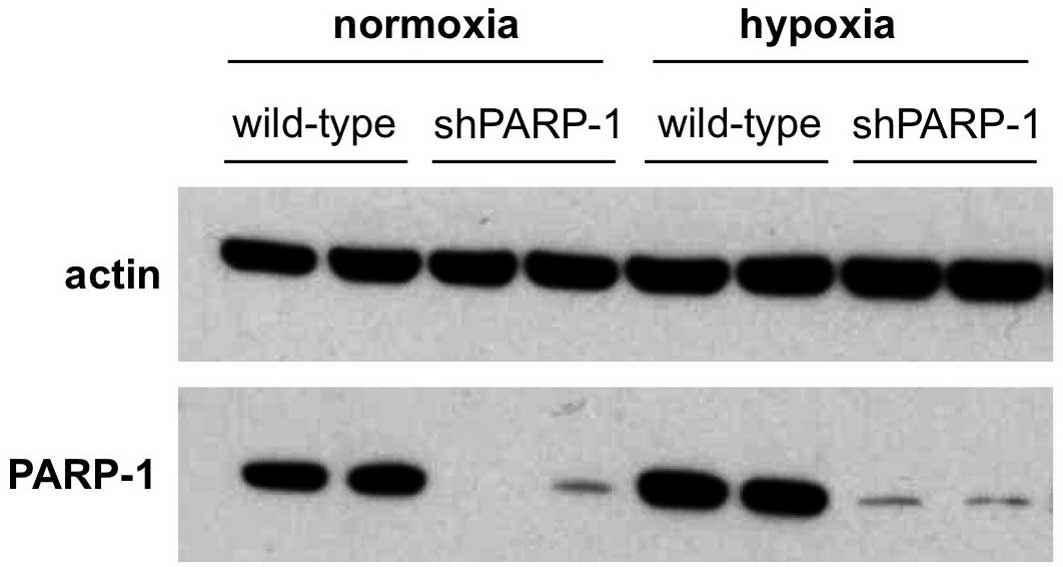
of PARP-1 remained stable after hypoxia. As expected, shPARP-1 cell
lines exhibited an efficient reduction of the corresponding
protein, with a small amount of residual PARP-1 enzyme remaining to
be identified (Fig. 2). Hypoxia,
by itself, did not induce an increase in DNA strand breakage
(Fig. 3) and did not activate
PARP-1 (Fig. 4). Although these
findings suggest that the current protocol of hypoxia used, by
itself, does not result in a significant degree of PARP-1
activation (or PARP-mediated decreases in cell viability), cell
viability was reduced by hypoxia in a partially PARP-1-dependent
manner. For instance, when measured at 48 h after the start of the
hypoxia (i.e., 24 h of hypoxia and 24 h of reoxygenation), ATP
levels were reduced to 58±2% of control in the wild-type cells,
while these levels were maintained at 71±3% of control in the
shPARP-1 cells (Fig. 5).
Mitochondrial activity, as evaluated by the MTT assay, showed a
similar pattern (60±7% of control in the wild-type cells, and
71±10% of control in the shPARP-1 cells) (Fig. 6). However, hypoxia alone did not
create a significant loss of cell membrane integrity as there were
no increases in LDH levels in the supernatant, nor did hypoxia
induce any detectable increase in the percentage of PI-positive
cells (Fig. 7).
Pre-exposure to hypoxia increased
sensitivity of cells to subsequent, oxidative stress-induced DNA
damage
H2O2 (50 or 100 μM) induced a
concentration-dependent increase in DNA breakage, as assessed by
single-cell electrophoresis (comet assay) (Fig. 3A). Cells that were pre-exposed to
hypoxia exhibited higher sensitivity to the subsequent oxidative
stress challenge, as evidenced by the significantly higher tail
moment developing in response to the lower dose of
H2O2 used (Fig.
3B). In accordance with the fact that DNA strand breakage is a
canonical trigger of PARP activation (8–10),
H2O2 induced a concentration-dependent
increase in the poly(ADP-ribose) polymer (PAR) level in all cell
groups. This was evidenced as a smear of 115–150 kDa, indicating
auto-modified PARP-1 (11). While
in cells that previously underwent hypoxia, baseline DNA strand
breakage or PARP activation remained unchanged,
H2O2 treatment triggered a substantially more
pronounced PARP-1 activation response, i.e., cells contained more
positive bands with the most immunopositivity identified in the
region of 75–150 kDa (Fig. 4). In
shPARP-1 cells H2O2 challenge only resulted
in a slight elevation of cellular PAR polymer content. However, the
H2O2-induced PARylation response was more
elevated in cells that were pre-exposed to hypoxia (Fig. 4). Despite the fact that shPARP-1
cells contained substantially lower amounts of PARP-1 compared to
wild-type cells, when these cells were previously exposed to
hypoxia, the H2O2-induced PARylation response
was higher than the corresponding response in wild-type cells that
were previously not exposed to hypoxia.
Role of PARP-1 in cell death induced
H2O2 with or without pre-exposure to
hypoxia
H2O2 caused a similar degree
of concentration-dependent decrease in the ATP content of both the
control and shPARP-1 cells (Fig.
5). When the cells were pre-exposed to hypoxia, this
H2O2-induced drop in ATP content was
amplified, and PARP-1 deletion provided significant protection
against this amplification (Fig.
5). Similar patterns were observed with respect to various
indices of cell viability: the H2O2-induced
decreases in MTT activity, LDH release into the supernatant, and
increases in the proportion of PI-positive cells, were all more
pronounced in the cells that were previously subjected to hypoxia,
than the corresponding control cells, and this hypersensitivity was
attenuated in the shPARP-1 cells, compared to wild-type cells
(Figs. 6 and 7). The control cells (not pre-exposed to
hypoxia) tolerated the H2O2 exposure
relatively well: even at the highest concentration (800 μM) of
H2O2 used, there was only a <50% decrease
in cellular ATP content and mitochondrial MTT conversion, with the
two responses being ameliorated by PARP-1 deficiency (Figs. 5 and 6). Moreover, the control cells (not
pre-exposed to hypoxia), did not exhibit any sign of membrane
integrity breakdown after H2O2 exposure, with
the exception of the 800 μM concentration, where a slight,
PARP-1-dependent increase in PI positivity and LDH release were
observed (Fig. 7).
Discussion
The principal finding of the current study is that a
24 h pre-exposure of human lung epithelial cells to hypoxia renders
them hypersensitive to subsequent oxidative stress in a partially
PARP-1-dependent manner. The cells that were not pre-exposed to
hypoxia tolerated the H2O2 exposure
relatively well across the entire concentration range used, with a
decrease in ATP and a slight increase in PI positivity noted at the
highest concentration of H2O2 (800 μM).
Similarly, hypoxic exposure by itself did not
produce cytotoxicity (as evidenced by lack of LDH release or lack
of change in PI positivity), although it created a partial
energetic imbalance (evidenced by a reduced degree of
mitochondria-dependent MTT conversion), and a decrease in the
intracellular ATP levels. The hypoxia-preexposed cells responded to
a markedly more pronounced cell death response following exposure
to H2O2 compared to the control cells (i.e.,
cells not pre-exposed to hypoxia). For instance, at 200 μM
H2O2, ATP levels and mitochondrial MTT
conversion were reduced by 75–80% of baseline values in the cells
that were pre-exposed to hypoxia, while the same parameters were
reduced by only 20–25% in cells that were not subjected to hypoxia.
Thus, it can be determined that decreases in mitochondrial MTT
conversion and cellular ATP content that are <50% (e.g., in
response to 400–800 μM H2O2 in the
non-hypoxic control group, or in response to 50–100 μM
H2O2 in the cells that were pre-exposed to
hypoxia) do not result in any detectable changes in membrane
permeability (i.e., lack of increase in LDH release or PI
positivity), while insults that reduce the ATP levels <50% are
generally associated with a breakdown of cell membrane integrity in
the current experimental system.
The functional importance of PARP-1 in the cytotoxic
response in our experimental model is supported by the fact that
shPARP-1 cells tolerated the hypoxic/oxidative challenge better
compared to the wild-type cells, as evidenced by an improved
maintenance of ATP, and a lower degree of suppression of
mitochondrial activity, coupled with a lower degree of LDH release
and lower proportion of PI-positive cells. The rank order of the
degree of PARP activation also mirrored the degree of energetic and
functional alterations across the four experimental groups studied:
the most pronounced response was noted in the wild-type group
subjected to hypoxia/H2O2, followed by the
shPARP-1 group subjected to hypoxia/H2O2,
followed by the wild-type group subjected to
H2O2 without hypoxia, and finally (the least
degree of PARP activation and the least degree of energetic and
functional changes) the shPARP-1 group subjected to
H2O2 without hypoxia (Figs. 5–7). The protective effect of PARP-1
deficiency, as well as the identical rank order for PARP-1
activation and the functional changes are all in strong support of
the hypothesis that activation of PARP-1 plays a direct causal role
in this process. According to the ‘canonical’ pathway of PARP-1
activation/cytotoxicity, DNA strand breakage activates PARP-1,
which, in turn, triggers a marked depletion of its substrate,
cellular NAD+, and, secondarily, cellular ATP, leading
to mitochondrial dysfunction and cell death via the necrotic path
(19–21). The cell responses elicited by
H2O2 in this study are in concordance with
those of that model. However, during the hypoxic period, it is
likely that mechanisms other than activation of PARP-1 play a key
role in the process of ATP depletion, because hypoxia alone did not
induce a PARylation response. Of note, the current experimental
design did not measure the full time-course of cellular ATP during
the 24 h of hypoxia and the subsequent reoxygenation. However, in
other studies utilizing 24 h hypoxia in epithelial cells, ATP
levels were measured in more detail (15,22). Based on these studies, we
hypothesized that in the current experiments hypoxia induces a
marked decrease in cellular ATP levels by 24 h, while the levels
measured at 48 h after the initiation of hypoxia (i.e., at 24 h
hypoxia + 24 h subsequent to normoxia) result from a partial
recovery of cellular ATP. Thus, at the time point of the
H2O2 exposure in the current protocol
(immediately after the end of the 24 h of hypoxia), the cells were
subjected to oxidative stress at a time when intracellular ATP
levels were the most severely depleted. In this respect, the
current in vitro model represents a form of the ‘two-hit’
injury model where the second ‘hit’ (oxidative stress) follows, in
short succession, the end of the first ‘hit’ (hypoxia). This type
of response, is relatively common, because revascularization of
ischemic organs in vivo is typically associated with an
oxidative (and nitrative) stress ‘burst’ response.
Although the functionality of the epithelial cell
monolayers was not assessed in the current experimental model,
previous studies have demonstrated that PARP-1 activation is an
important regulator of epithelial cell permeability both in
vitro and in vivo in various models of critical illness
(23–26). Therefore, it is likely that
PARP-1-dependent alterations in epithelial cell energetics also
translate to functional alterations in epithelial barrier function.
Moreover, in addition to being a regulator of cellular
bioenergetics (8,11,15,18–22), PARP-1, also affects
pro-inflammatory gene transcription and inflammatory mediator
production (27–32). Additional studies are needed to
investigate whether the well-known propensity of the ‘two-hit’
models of critical illness to an enhanced inflammatory mediator
production can also be reproduced in the current in vitro
system, and whether PARP activation is important in these
responses.
The precise early-stage trigger of PARP-1
overactivation (and subsequent cellular dysfunction) in the current
experimental model was investigated. Based on our findings showing
that H2O2 induces a more pronounced degree of
DNA strand breaks in the cells pre-exposed to hypoxia than in the
control cells (Fig. 3), we
hypothesize that hypoxia renders the nuclear DNA more sensitive to
oxidative stress, either by permitting the oxidants to penetrate
the cells better, and/or by causing a more pronounced degree of
secondary oxidant production (33), for instance by the promotion of
mitochondrial dysfunction and mitochondrial oxidant production. In
general, oxidative stress reflects an imbalance between the
systematic occurrence of reactive oxygen species and the ability of
the biological system to detoxify the harmful intermediates or to
repair the resulting damage.
One of the key intracellular antioxidants is GSH, a
key intracellular tripeptide thiol composed of glutamic acid,
cysteine and glycine. GSH functions as an antioxidant, preventing
damage to cell components caused by reactive oxygen species
(34). It exists in reduced GSH
and oxidized GSSG form. In the reduced state, the thiol group of
the cysteine provides reduction equivalents to other unstable
molecules, such as reactive oxygen species. After donating the
electron, GSH becomes reactive, but reacts readily with another
oxidized glutathione to form glutathione disulfide (GSSG). Under
normal circumstances >90% of the total glutathione pool is in
the reduced state (GSH), while the remaining glutathione pool is in
the disulfide form (GSSG). The higher concentration of GSH is due
to the glutathione reductase enzyme, which is constitutively active
and inducible following oxidative stress (34). Earlier studies demonstrated that
pharmacological depletion of intracellular GSH with
L-buthionine-(S,R)-sulfoximine (an inhibitor of γ-glutamylcysteine
synthetase) renders endothelial cells hypersensitive to oxidative
and nitrative stress and PARP activation (35,36). Therefore, GSH levels were measured
in our experiments and hypoxia was observed to result in a
substantial decrease in its levels (Fig. 1). In accordance with previous
studies using pharmacological PARP inhibitors such as
3-aminobenzamide (37), the
hypoxia-mediated GSH depletion was, in the current experiments, at
least partially dependent on the presence of PARP-1. GSH depletion
has been previously shown to exacerbate the degree of
oxidant-mediated DNA damage in a variety of experimental systems
in vitro(38–40). Therefore, the above findings are
consistent with the hypothesis that GSH depletion is a causative
step in the process of hypoxia-associated, PARP-1-mediated
enhancement of oxidative stress-induced cytotoxicity in the
current, ‘two-hit’ in vitro experimental model. The present
study also supports the view that restoration of endogenous
antioxidants, such as GSH and/or pharmacological inhibition of
PARP-1, can be of benefit in various models of critical illness,
including ‘two-hit models’ and other pathophysiological conditions
where hypoxia is followed by a subsequent oxidative stress-mediated
insult.
Acknowledgements
This study was supported by the US National
Institutes of Health 2P50 GM060338 to C.S.
Abbreviations:
|
ATP
|
adenosine triphosphate
|
|
GSH
|
glutathione
|
|
H2O2
|
hydrogen peroxide
|
|
MTT
|
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium
bromide
|
|
LDH
|
lactate dehydrogenase
|
|
PI
|
propidium iodide
|
|
PAR
|
poly(ADP-ribose) polymer
|
|
PARP
|
poly(ADP-ribose) polymerase
|
References
|
1
|
Saadia R and Schein M: Multiple organ
failure. How valid is the ‘two hit’ model? J Accid Emerg Med.
16:163–166. 1999.
|
|
2
|
Rotstein OD: Modeling the two-hit
hypothesis for evaluating strategies to prevent organ injury after
shock/resuscitation. J Trauma. 54:S203–S206. 2003.PubMed/NCBI
|
|
3
|
West MA, Li MH, Seatter SC and Bubrick MP:
Pre-exposure to hypoxia or septic stimuli differentially regulates
endotoxin release of tumor necrosis factor, interleukin-6,
interleukin-1, prostaglandin E2, nitric oxide, and superoxide by
macrophages. J Trauma. 37:82–89. 1994. View Article : Google Scholar
|
|
4
|
Kim SY, Choi YJ, Joung SM, Lee BH, Jung YS
and Lee JY: Hypoxic stress up-regulates the expression of Toll-like
receptor 4 in macrophages via hypoxia-inducible factor. Immunology.
129:516–524. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brehmer F, Bendix I, Prager S, van de
Looij Y, Reinboth BS, Zimmermanns J, Schlager GW, Brait D,
Sifringer M, Endesfelder S, Sizonenko S, Mallard C, Bührer C,
Felderhoff-Mueser U and Gerstner B: Interaction of inflammation and
hyperoxia in a rat model of neonatal white matter damage. PLoS One.
7:e490232012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng XQ, Song LJ, Li H, Di H, Zhang YY
and Chen DF: Beneficial effect of the polysaccharides from
Bupleurum smithii var. parvifolium on ‘two-hit’ acute lung injury
in rats. Inflammation. 35:1715–1722. 2012.
|
|
7
|
Kox M, Vaneker M, van der Hoeven JG,
Scheffer GJ, Hoedemaekers CW and Pickkers P: Effects of vagus nerve
stimulation and vagotomy on systemic and pulmonary inflammation in
a two-hit model in rats. PLoS One. 7:e344312012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Szabo C: Role of poly(ADP-ribose)
synthetase in inflammation. Eur J Pharmacol. 350:1–19. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pacher P and Szabo C: Role of
poly(ADP-ribose) polymerase 1 (PARP-1) in cardiovascular diseases:
the therapeutic potential of PARP inhibitors. Cardiovasc Drug Rev.
25:235–260. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Graziani G and Szabo C: Clinical
perspectives of PARP inhibitors. Pharmacol Res. 52:109–118. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Erdélyi K, Bai P, Kovács I, Szabó E,
Mocsár G, Kakuk A, Szabo C, Gergely P and Virág L: Dual role of
poly(ADP-ribose) glycohydrolase in the regulation of cell death in
oxidatively stressed A549 cells. FASEB J. 23:3553–3563.
2009.PubMed/NCBI
|
|
12
|
Salzman A, Denenberg AG, Ueta I, O’Connor
M, Linn SC and Szabo C: Induction and activity of nitric oxide
synthase in cultured human intestinal epithelial monolayers. Am J
Physiol. 270:G565–G573. 1996.PubMed/NCBI
|
|
13
|
Jakovljevic B, Novakov-Mikic A, Brkic S,
Bogavac MA, Tomic S and Miler V: Lipid peroxidation in the first
trimester of pregnancy. J Matern Fetal Neonatal Med. 25:1316–1318.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Suzuki K, Olah G, Modis K, Coletta C, Kulp
G, Gerö D, Szoleczky P, Chang T, Zhou Z, Wu L, Wang R,
Papapetropoulos A and Szabo C: Hydrogen sulfide replacement therapy
protects the vascular endothelium in hyperglycemia by preserving
mitochondrial function. Proc Natl Acad Sci USA. 108:13829–13834.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Módis K, Gero D, Erdélyi K, Szoleczky P,
DeWitt D and Szabo C: Cellular bioenergetics is regulated by PARP1
under resting conditions and during oxidative stress. Biochem
Pharmacol. 83:633–643. 2012.PubMed/NCBI
|
|
16
|
Southan GJ, Zingarelli B, O’Connor M,
Salzman AL and Szabo C: Spontaneous rearrangement of
aminoalkylisothioureas into mercaptoalkylguanidines, a novel class
of nitric oxide synthase inhibitors with selectivity towards the
inducible isoform. Br J Pharmacol. 117:619–632. 1996. View Article : Google Scholar
|
|
17
|
Gerö D, Módis K, Nagy N, Szoleczky P, Tóth
ZD, Dormán G and Szabo C: Oxidant-induced cardiomyocyte injury:
identification of the cytoprotective effect of a dopamine 1
receptor agonist using a cell-based high-throughput assay. Int J
Mol Med. 20:749–761. 2007.PubMed/NCBI
|
|
18
|
Virág L and Szabo C: Purines inhibit
poly(ADP-ribose) polymerase activation and modulate oxidant-induced
cell death. FASEB J. 15:99–107. 2011.PubMed/NCBI
|
|
19
|
Virág L, Scott GS, Cuzzocrea S, Marmer D,
Salzman AL and Szabo C: Peroxynitrite-induced thymocyte apoptosis:
the role of caspases and poly(ADP-ribose) synthetase (PARS)
activation. Immunology. 94:345–355. 1998.PubMed/NCBI
|
|
20
|
Virág L, Salzman AL and Szabo C:
Poly(ADP-ribose) synthetase activation mediates mitochondrial
injury during oxidant-induced cell death. J Immunol. 161:3753–3759.
1998.PubMed/NCBI
|
|
21
|
Ha HC and Snyder SH: Poly(ADP-ribose)
polymerase is a mediator of necrotic cell death by ATP depletion.
Proc Natl Acad Sci USA. 96:13978–13982. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Szoleczky P, Módis K, Nagy N, Dóri Tóth Z,
DeWitt D, Szabo C and Gero D: Identification of agents that reduce
renal hypoxia-reoxygenation injury using cell-based screening:
purine nucleosides are alternative energy sources in LLC-PK1 cells
during hypoxia. Arch Biochem Biophys. 517:53–70. 2012.
|
|
23
|
Szabo C, Saunders C, O’Connor M and
Salzman AL: Peroxynitrite causes energy depletion and increases
permeability via activation of poly(ADP-ribose) synthetase in
pulmonary epithelial cells. Am J Respir Cell Mol Biol. 16:105–109.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kennedy M, Denenberg AG, Szabo C and
Salzman AL: Poly(ADP-ribose) synthetase activation mediates
increased permeability induced by peroxynitrite in Caco-2BBe cells.
Gastroenterology. 114:510–518. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liaudet L, Szabó A, Soriano FG, Zingarelli
B, Szabo C and Salzman AL: Poly(ADP-ribose) synthetase mediates
intestinal mucosal barrier dysfunction after mesenteric ischemia.
Shock. 14:134–141. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shimoda K, Murakami K, Enkhbaatar P,
Traber LD, Cox RA, Hawkins HK, Schmalstieg FC, Komjati K, Mabley
JG, Szabo C, Salzman AL and Traber DL: Effect of poly(ADP ribose)
synthetase inhibition on burn and smoke inhalation injury in sheep.
Am J Physiol Lung Cell Mol Physiol. 285:L240–L249. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Szabo C, Lim LH, Cuzzocrea S, Getting SJ,
Zingarelli B, Flower RJ, Salzman AL and Perretti M: Inhibition of
poly(ADP-ribose) synthetase attenuates neutrophil recruitment and
exerts antiinflammatory effects. J Exp Med. 186:1041–1049. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cuzzocrea S, Zingarelli B, Gilad E, Hake
P, Salzman AL and Szabo C: Protective effects of 3-aminobenzamide,
an inhibitor of poly(ADP-ribose) synthase in a carrageenan-induced
model of local inflammation. Eur J Pharmacol. 342:67–76. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Veres B, Gallyas F Jr, Varbiro G, Berente
Z, Osz E, Szekeres G, Szabo C and Sumegi B: Decrease of the
inflammatory response and induction of the Akt/protein kinase B
pathway by poly(ADP-ribose) polymerase 1 inhibitor in
endotoxin-induced septic shock. Biochem Pharmacol. 65:1373–1382.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goldfarb RD, Marton A, Szabó E, Virág L,
Salzman AL, Glock D, Akhter I, McCarthy R, Parrillo JE and Szabo C:
Protective effect of a novel, potent inhibitor of poly(adenosine
5′-diphosphate-ribose) synthetase in a porcine model of severe
bacterial sepsis. Crit Care Med. 30:974–980. 2002.
|
|
31
|
Soriano FG, Liaudet L, Szabó E, Virág L,
Mabley JG, Pacher P and Szabo C: Resistance to acute septic
peritonitis in poly(ADP-ribose) polymerase-1-deficient mice. Shock.
17:286–292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bai P and Virág L: Role of
poly(ADP-ribose) polymerases in the regulation of inflammatory
processes. FEBS Lett. 586:3771–3777. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cantoni O and Guidarelli A: Peroxynitrite
damages U937 cell DNA via the intermediate formation of
mitochondrial oxidants. IUBMB Life. 60:753–756. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jean JC, Liu Y and Joyce-Brady M: The
importance of gamma-glutamyl transferase in lung glutathione
homeostasis and antioxidant defense. Biofactors. 17:161–173. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cuzzocrea S, Zingarelli B, O’Connor M,
Salzman AL and Szabo C: Effect of L-buthionine-(S,R)-sulphoximine,
an inhibitor of gamma-glutamylcysteine synthetase on peroxynitrite-
and endotoxic shock-induced vascular failure. Br J Pharmacol.
123:525–537. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kiss L, Chen M, Gero D, Módis K, Lacza Z
and Szabo C: Effects of 7-ketocholesterol on the activity of
endothelial poly(ADP-ribose) polymerase and on
endothelium-dependent relaxant function. Int J Mol Med.
18:1113–1117. 2006.PubMed/NCBI
|
|
37
|
Mizumoto K, Glascott PA Jr and Farber JL:
Roles for oxidative stress and poly(ADP-ribosyl)ation in the
killing of cultured hepatocytes by methyl methanesulfonate. Biochem
Pharmacol. 46:1811–1818. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bump EA and Brown JM: Role of glutathione
in the radiation response of mammalian cells in vitro and in vivo.
Pharmacol Ther. 47:117–136. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zucker B, Hanusch J and Bauer G:
Glutathione depletion in fibroblasts is the basis for
apoptosis-induction by endogenous reactive oxygen species. Cell
Death Differ. 4:388–395. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Puhakka A, Ollikainen T, Soini Y, Kahlos
K, Säily M, Koistinen P, Pääkkö P, Linnainmaa K and Kinnula VL:
Modulation of DNA single-strand breaks by intracellular glutathione
in human lung cells exposed to asbestos fibers. Mutat Res.
514:7–17. 2002. View Article : Google Scholar : PubMed/NCBI
|