Introduction
Chronic pain that is severe and difficult to manage
can degrade the quality of life of patients. The two most common
forms of chronic pain are inflammatory pain and neuropathic pain.
These two conditions show similar clinical symptoms; however, their
underlying mechanisms and responses are dissimilar. In general,
chronic pain involves changes in the expression of messenger RNAs
(mRNAs) or proteins that regulate cytokines, receptors and
neurotransmitters in the dorsal root ganglia and the brain
(1,2). Previous studies have suggested that
the differential expression of multiple pain-associated genes plays
a key role in the development and maintenance of chronic pain
(3–5).
The hippocampus, a brain region that participates in
learning and memory formation, plays a fundamental role in pain
perception (6,7). The association between the
hippocampus and pain has been the focus of a number of recent
studies. In particular, recent studies have indicated that
peripheral neuropathy alters hippocampal gene expression, which
suggests that the hippocampus may contribute to neuropathic pain
symptoms (8–10).
There are a few studies, however, that directly
examine the association between the hippocampus and inflammatory or
neuropathic pain. Furthermore, few studies have examined the role
that hippocampal microRNAs (miRNAs) may play in chronic pain,
particularly for the two different pain models. We hypothesized
that hippocampal miRNA expression may differ between inflammatory
and neuropathic pain models. An improved understanding of the
underlying mechanisms for chronic pain may lead to more effective
treatment strategies.
In this study, we used TaqMan®
low-density array (TLDA) and quantitative real-time PCR (qRT-PCR)
of miRNAs to examine miRNA expression in the hippocampus in two rat
models of chronic pain. The model of neuropathic pain comprised
rats with chronic constriction injury (CCI), whereas the model of
inflammatory pain comprised rats injected with complete Freund’s
adjuvant (CFA), as previously described (11,12). In addition, we examined
differences in gene expression between the hippocampus bilaterally,
as bilateral effects have been observed with unilateral chronic
pain models induced by inflammation or by ischemic monomelic
neuropathy (13).
Materials and methods
All experimental procedures were approved by the
Institutional Committee on Laboratory Animals of Nippon Medical
School, Tokyo, Japan (approval no. 19-091) and were performed under
the guidelines of the International Association for the Study of
Pain.
Male Sprague-Dawley rats (6–7 weeks of age and
200–250 g in weight; Experimental Animal Center, Saitama, Japan)
were used. The rats were housed in clear plastic cages with sawdust
bedding at standard room temperature under a 12-h light/dark cycle.
All rats received food and water ad libitum. We divided the
rats into 4 experimental groups as follows: i) a group of rats with
CCI that were sacrificed 7 days after the procedure (CCI group,
n=6); ii) a group of sham-CCI operated rats (sham-CCI group, n=6)
[as previously described (11)];
iii) a group of rats injected with CFA that were sacrificed 7 days
after the procedure (CFA group, n=6); and iv) a group of normal
saline-injected rats (sham-CFA group, n=6) [as previously described
(12)]. All surgical procedures
were performed under deep anesthesia with sodium pentobarbital [50
mg/kg intraperitoneal (i.p.) injection].
Rat model of neuropathic pain induced by
CCI
The CCI model was established according to the
methods of Bennett and Xie (11).
The left (ipsilateral) common sciatic nerve was exposed and loosely
ligated with 4-0 silk thread at 4 regions that were spaced at
approximately 1-mm intervals. The sciatic nerve in the
sham-operated group was similarly exposed but not ligated.
CFA injection for the induction of
inflammatory pain
The CFA model was established by injecting CFA (50%
in saline, 20 μl) into the plantar surface of the left hindpaw. In
the sham-CFA group, normal saline was similarly injected into the
left hindpaw as previously described (12).
Behavioral assessment
The Plantar Test apparatus (Ugo Basile, Comerio,
Italy) was used to examine thermal hyperalgesia, and the von Frey
test (Muromachi Kikai, Tokyo, Japan) was used to examine mechanical
allodynia. These tests were performed on the day before surgery
(day 0), and on post-operative days 1, 3, 5, 7, 9, 11, 13 and 15,
as previously described (14).
For the plantar test, each rat was placed on a glass plate with
radiant heat equipment (a 50-W halogen reflector bulb) underneath.
After the acclimation period, radiation heat was applied to either
the contralateral or ipsilateral hindpaw pad. The latency of paw
withdrawal from the thermal stimulus was measured 3 times at 5-min
intervals, and the average value of these 3 measurements was used
as the response latency. For the von Frey test, each rat was placed
on a metallic mesh floor, covered with a plastic box, and a von
Frey filament with bending forces from 2.0 to 32.0 g was applied
from under the mesh floor to the plantar surface of either the
contralateral or ipsilateral hindpaw. In individual trials, each
paw was stimulated with each filament 5 times at 10-sec intervals.
The weakest force (g) that induced the withdrawal of the stimulated
paw at least 3 times in each trial was considered the paw
withdrawal threshold.
miRNA profiling
On post-operative day 7, the rats were deeply
anesthetized with pentobarbital (300 mg/100 g body weight, i.p.)
and decapitated immediately. The brains were rapidly removed, and
the hippocampi were dissected as previously described (15). The unilateral hippocampi were
divided into left and right sections. Each sample was placed in RNA
later® (Applied Biosystems, LLC, Foster City, CA, USA)
and stored at −80°C until use. Total RNA was isolated using a
mirVana™ miRNA isolation kit® (Applied Biosystems, LLC)
according to the manufacturer’s instructions. RNA quantity and
quality were assessed using the NanoDrop ND-1000 spectrophotometer
(Thermo Fisher Scientific, Waltham, MA). All RNA samples with an
A260/280 nm reading of 1.8 were used for quantitative analysis.
Total RNA samples containing miRNAs were used for qRT-PCR.
The miRNA expression profiles were analyzed using
TLDA Rodent MicroRNA cards v.3 A and B (Applied Biosystems, LLC).
Each card contains 373 pre-loaded rodent miRNA targets, which are
all catalogued in the miRBase database (16) and 3 endogenous controls. The
procedures were performed as previously described (17,18). Briefly, TLDAs were performed using
a two-step process. First, 800 ng total RNA per sample was reverse
transcribed using Megaplex RT primer pool A and B, which contains
up to 381 stem-looped primers per pool, and a TaqMan MicroRNA
Reverse Transcription kit (Applied Biosystems, LLC). Second, the
resulting complementary DNA was diluted, mixed with TaqMan
Universal PCR master mix (Applied Biosystems, LLC), and deionized
in distilled water (Wako, Tokyo, Japan) and loaded into 1 of the 8
fill ports on the TLDA microfluidic cards. The cards were briefly
centrifuged for 1 min at 1,600 × g to distribute samples to the
multiple wells connected to the fill ports, and then sealed to
prevent well-to-well contamination. Finally, the cards were
processed and analyzed using a 7900 HT Real-Time PCR System
(Applied Biosystems, LLC).
Data analysis was performed using DataAssist
software v2.0 (Applied Biosystems, LLC). The data were represented
as the threshold cycle (Ct) values, where Ct represents a unitless
value defined as the fractional cycle number at which the sample
fluorescence signal passes a fixed threshold above baseline. For
each miRNA, the expression level was calculated using the
comparative Ct method (ΔΔCt) and was further analyzed by comparing
the fold change to the basal levels in the control samples. ΔCt was
the difference in the Ct values derived from the experimental
samples and the control, and ΔΔCt represented the difference
between paired samples, as calculated by the following formula:
ΔΔCt = ΔCt of sample after surgical procedure −ΔCt of the control.
The expression ratio shows the relative quantity of the target gene
(Xtarget) to the control gene (Xcontrol). The fold change was
computed by the formula Xtarget/Xcontrol = 2−ΔΔCt. For
miRNAs, graphic displays were visualized as heat map results of
hierarchical clustering. Distances between the samples and assays
were calculated for hierarchical clustering based on ΔCt values
using the Euclidean distance.
Statistical analyses
Values are expressed as the means ± standard
deviation. A two-tailed paired t-test was used to compare latencies
or threshold values in behavioral tests between the ipsilateral and
contralateral sides (CCI group; n=6). Dunnett’s test for multiple
comparisons was used to compare latencies and threshold values
obtained in the behavioral tests performed pre-operatively (day 0,
n=6) with those obtained post-operatively (days 1, 3, 5, 7, 9, 11,
13 and 15; n=6). ANOVA followed by Tukey’s test were performed
using KyPlot 5.0 software (KyensLab Inc., Tokyo, Japan). Using
Pearson’s correlation, we assessed whether the hippocampal miRNAs
that showed expression changes in the rat models of chronic pain
were differentially expressed between the left and right
hippocampus.
Results
Behavioral assessment
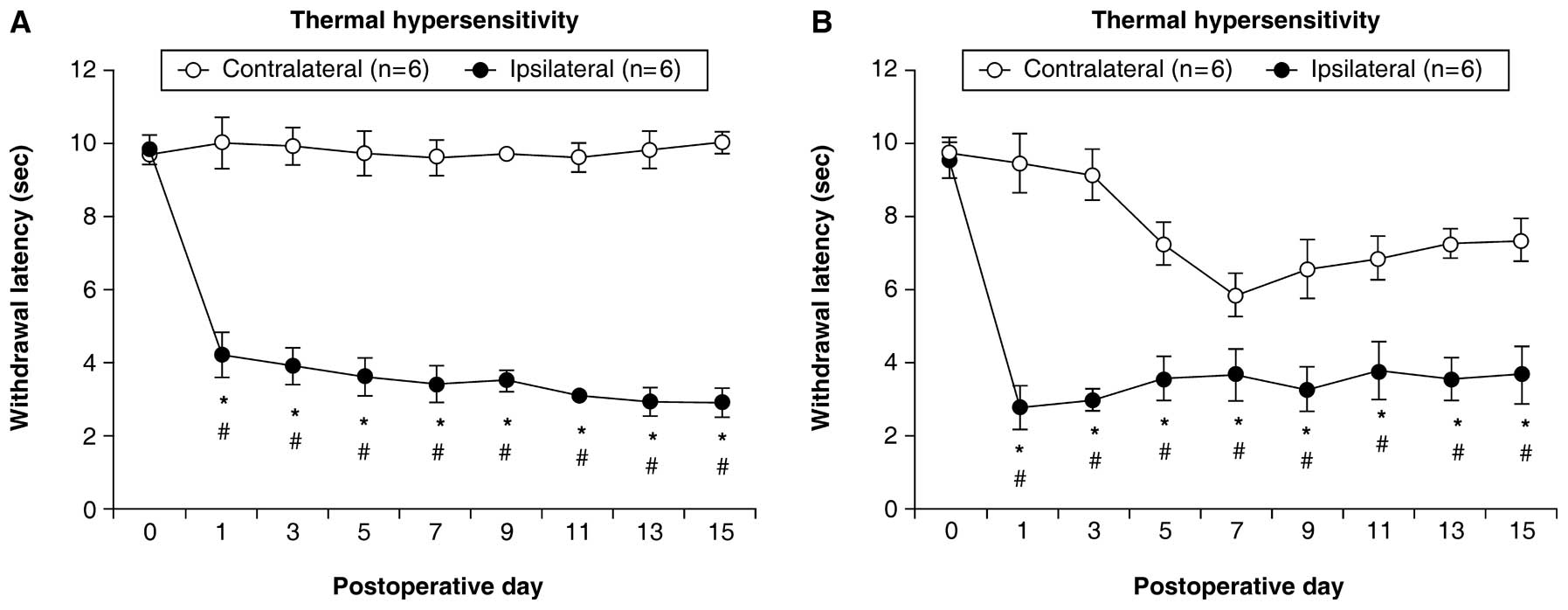
The rats with CCI showed a significant decrease in
the latency of paw withdrawal in the plantar test and threshold
values in the von Frey test for the ipsilateral side on
post-operative days (1, 3, 5, 7, 9, 11, 13 and 15) compared with
day 0 (P<0.05). On the contralateral side, significant decreases
were not observed (Figs. 1A and
2A).
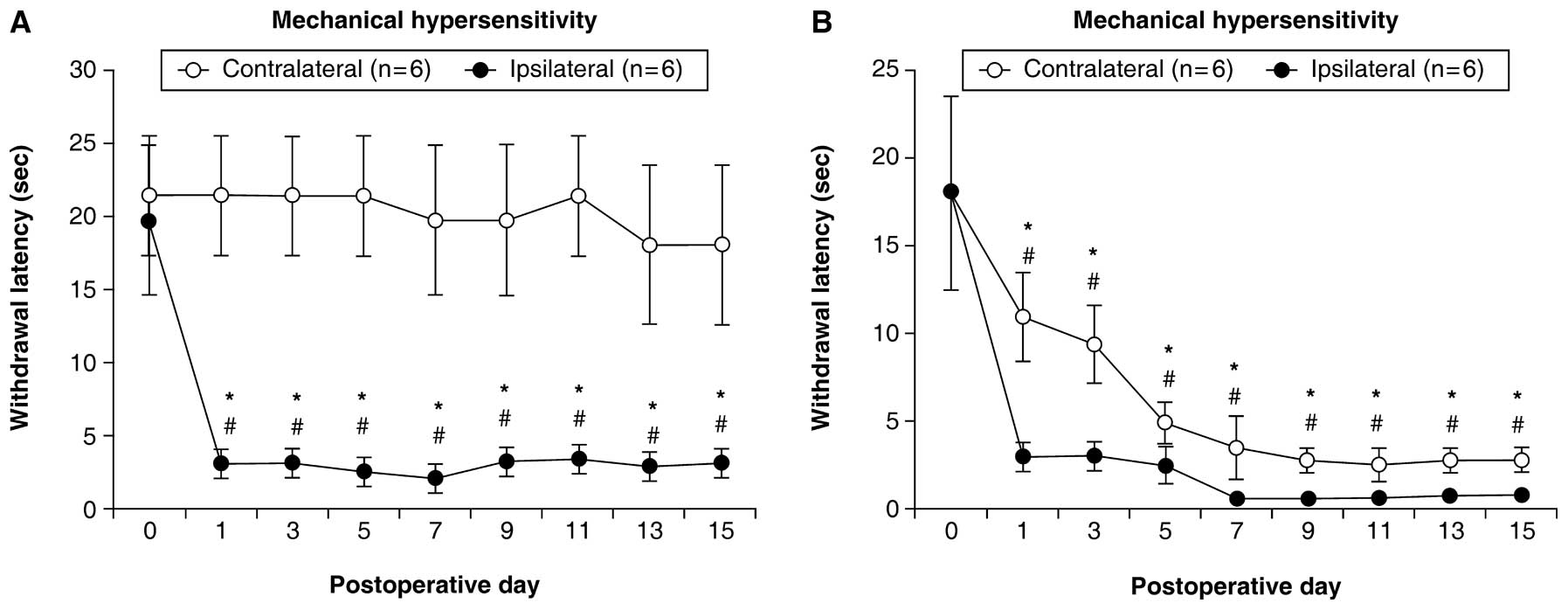
Even though the CFA injection was administered to
the left hindpaw, the rats with CFA showed a significant decrease
in the latency of paw withdrawal in the plantar test, and a
significant decrease in threshold values in the von Frey test for
the hindpaws bilaterally. These differences were observed on the
post-operative days examined compared with day 0 (P<0.05)
(Figs. 1B and 2B).
TLDA
Of the 373 rat miRNAs, 237 (63.5%) were detected in
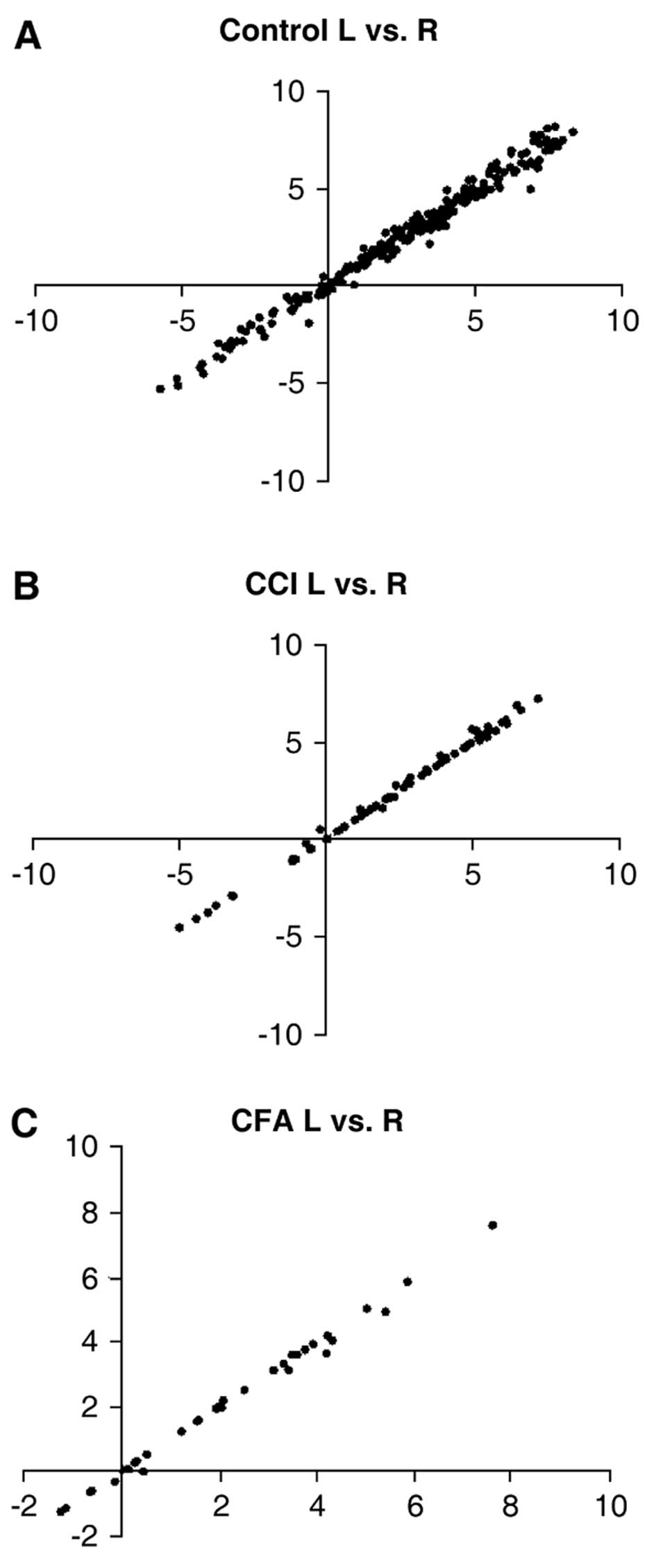
the rat hippocampus. In the control rats, we observed no difference
in miRNA expression between the left and right hippocampus
(correlation value, 0.99; P<0.0001). Compared with the
sham-operated rats, TLDA in the rats with CCI identified 54 miRNAs
(22.7%) that were differentially expressed, including 7 miRNAs that
were downregulated (Table I).
 | Table IThe 54 miRNAs that were differentially
expressed in the CCI group compared with the sham-operated
group. |
Table I
The 54 miRNAs that were differentially
expressed in the CCI group compared with the sham-operated
group.
| Assay | Fold change ± SD | P-value |
|---|
| hsa-miR-324-3p | 1.64±0.40 | <0.001 |
| mmu-miR-125a-5p | 2.10±0.24 | <0.001 |
| mmu-miR-132 | 2.11±0.46 | <0.001 |
| mmu-miR-151-3p | 3.51±0.18 | <0.001 |
| mmu-miR-17 | 1.77±0.13 | <0.001 |
| mmu-miR-181c | 0.51±0.21 | <0.001 |
| mmu-miR-191 | 3.06±1.20 | <0.001 |
| mmu-miR-222 | 2.38±0.44 | <0.001 |
| mmu-miR-29c | 0.37±0.29 | <0.001 |
| mmu-miR-31 | 2.04±0.43 | <0.001 |
| mmu-miR-320 | 1.76±0.54 | <0.001 |
| mmu-miR-434-3p | 3.35±0.78 | <0.001 |
| mmu-miR-539 | 2.24±0.68 | <0.001 |
| rno-miR-345-3p | 2.48±0.37 | <0.001 |
| rno-miR-381 | 0.25±0.31 | <0.001 |
| hsa-miR-30a-3p | 1.83±0.51 | <0.01 |
| hsa-miR-30e-3p | 1.55±0.40 | <0.01 |
| mmu-miR-126-3p | 1.96±0.28 | <0.01 |
| mmu-miR-133a | 2.48±1.55 | <0.01 |
| mmu-miR-140 | 1.65±0.25 | <0.01 |
| mmu-miR-150 | 2.04±0.98 | <0.01 |
| mmu-miR-212 | 2.14±0.74 | <0.01 |
| mmu-miR-30a | 1.42±0.18 | <0.01 |
| mmu-miR-323-3p | 1.96±0.58 | <0.01 |
| mmu-miR-331-3p | 1.42±0.14 | <0.01 |
| mmu-miR-383 | 2.02±1.07 | <0.01 |
| mmu-miR-431 | 1.63±0.46 | <0.01 |
| mmu-miR-487b | 1.45±0.36 | <0.01 |
| mmu-miR-770-5p | 1.82±0.64 | <0.01 |
| mmu-miR-872# | 1.92±0.50 | <0.01 |
| rno-miR-125b# | 2.09±0.61 | <0.01 |
| rno-miR-146B | 1.74±0.41 | <0.01 |
| rno-miR-339-3p | 3.21±1.57 | <0.01 |
| rno-miR-409-3P | 1.75±0.51 | <0.01 |
| rno-miR-504 | 1.76±0.56 | <0.01 |
| rno-miR-632 | 0.58±0.23 | <0.01 |
| rno-miR-664 | 1.70±0.50 | <0.01 |
| rno-miR-7a# | 1.85±0.63 | <0.01 |
| hsa-miR-28-3p | 2.17±0.43 | <0.05 |
| hsa-miR-423-3P | 1.65±0.41 | <0.05 |
| mmu-miR-128a | 1.61±0.23 | <0.05 |
| mmu-miR-134 | 1.43±0.31 | <0.05 |
| mmu-miR-138 | 2.07±0.95 | <0.05 |
| mmu-miR-186 | 2.54±2.32 | <0.05 |
| mmu-miR-193b | 1.98±1.10 | <0.05 |
| mmu-miR-204 | 1.81±0.84 | <0.05 |
| mmu-miR-23b | 1.53±0.19 | <0.05 |
| mmu-miR-24 | 1.57±0.40 | <0.05 |
| mmu-miR-30d | 0.63±0.16 | <0.05 |
| mmu-miR-325 | 0.58±0.44 | <0.05 |
| mmu-miR-376b | 0.58±0.26 | <0.05 |
| mmu-miR-380-5p | 1.45±0.41 | <0.05 |
| mmu-miR-384-5p | 1.62±0.56 | <0.05 |
| rno-miR-351 | 2.24±0.54 | <0.05 |
Compared with the normal saline-injected rats, the
CFA-injected rats had 40 miRNAs (16.8%) that were differentially
expressed, including 25 miRNAs that were downregulated (Table II).
| Table IIThe 40 miRNAs that were
differentially expressed in the CFA group compared with the normal
saline group. |
Table II
The 40 miRNAs that were
differentially expressed in the CFA group compared with the normal
saline group.
| Assay | Fold change ±
SD | P-value |
|---|
| hsa-miR-324-3p | 1.53±0.30 | <0.001 |
| mmu-miR-15b | 1.09±0.26 | <0.001 |
| mmu-miR-191 | 3.06±0.90 | <0.001 |
| mmu-miR-326 | 0.52±0.50 | <0.001 |
| mmu-miR-770-5p | 1.85±0.46 | <0.001 |
| rno-miR-409-3P | 1.63±0.50 | <0.001 |
| hsa-miR-223 | 2.83±2.23 | <0.01 |
| hsa-miR-30a-3p | 1.86±1.08 | <0.01 |
|
mmu-miR-125a-5p | 2.10±0.50 | <0.01 |
| mmu-miR-126-3p | 1.96±0.35 | <0.01 |
| mmu-miR-133a | 2.48±0.92 | <0.01 |
| mmu-miR-134 | 1.43±0.38 | <0.01 |
| mmu-miR-138# | 1.98±0.80 | <0.01 |
| mmu-miR-139-5p | 1.96±0.51 | <0.01 |
| mmu-miR-148b | 1.03±0.41 | <0.01 |
| mmu-miR-212 | 2.06±1.20 | <0.01 |
| mmu-miR-296-5p | 0.61±0.19 | <0.01 |
| mmu-miR-652 | 1.13±0.16 | <0.01 |
| mmu-miR-708 | 1.48±0.23 | <0.01 |
| mmu-miR-872 | 1.00±0.13 | <0.01 |
| rno-miR-146B | 1.77±0.62 | <0.01 |
|
rno-miR-219-2-3p | 0.83±0.15 | <0.01 |
| rno-miR-664 | 1.45±0.55 | <0.01 |
| rno-miR-7a# | 1.49±0.58 | <0.01 |
| hsa-miR-140-3p | 1.54±0.45 | <0.05 |
| hsa-miR-30e-3p | 1.37±0.44 | <0.05 |
| mmu-miR-142-3p | 0.70±0.06 | <0.05 |
| mmu-miR-146a | 2.27±0.88 | <0.05 |
| mmu-miR-150 | 2.04±0.68 | <0.05 |
| mmu-miR-219 | 0.87±0.23 | <0.05 |
| mmu-miR-27a | 0.98±0.16 | <0.05 |
| mmu-miR-324-5p | 0.90±0.08 | <0.05 |
| mmu-miR-375 | 0.78±0.37 | <0.05 |
| mmu-miR-383 | 2.02±0.57 | <0.05 |
| mmu-miR-539 | 2.24±0.64 | <0.05 |
| mmu-miR-7a | 1.03±0.21 | <0.05 |
| mmu-miR-872# | 2.40±1.48 | <0.05 |
| rno-miR-125b# | 2.05±1.02 | <0.05 |
| rno-miR-204# | 2.52±0.79 | <0.05 |
| rno-miR-504 | 1.30±0.18 | <0.05 |
Twenty miRNAs were expressed in both the rats with
CCI and the CFA-injected rats, whereas 34 miRNAs were expressed in
only the rats with CCI, and 22 were expressed in only the
CFA-injected rats. There were no significant differences observed
in the comparisons between the sham-operated rats and the normal
saline-injected rats.
We observed no difference between the hippocampi
bilaterally for either the CCI group (correlation value, 0.99;
P<0.0001) or the CFA group (correlation value, 0.98;
P<0.0001). Furthermore, there was no difference bilaterally
among the control rats, the rats with CCI or the CFA-injected rats
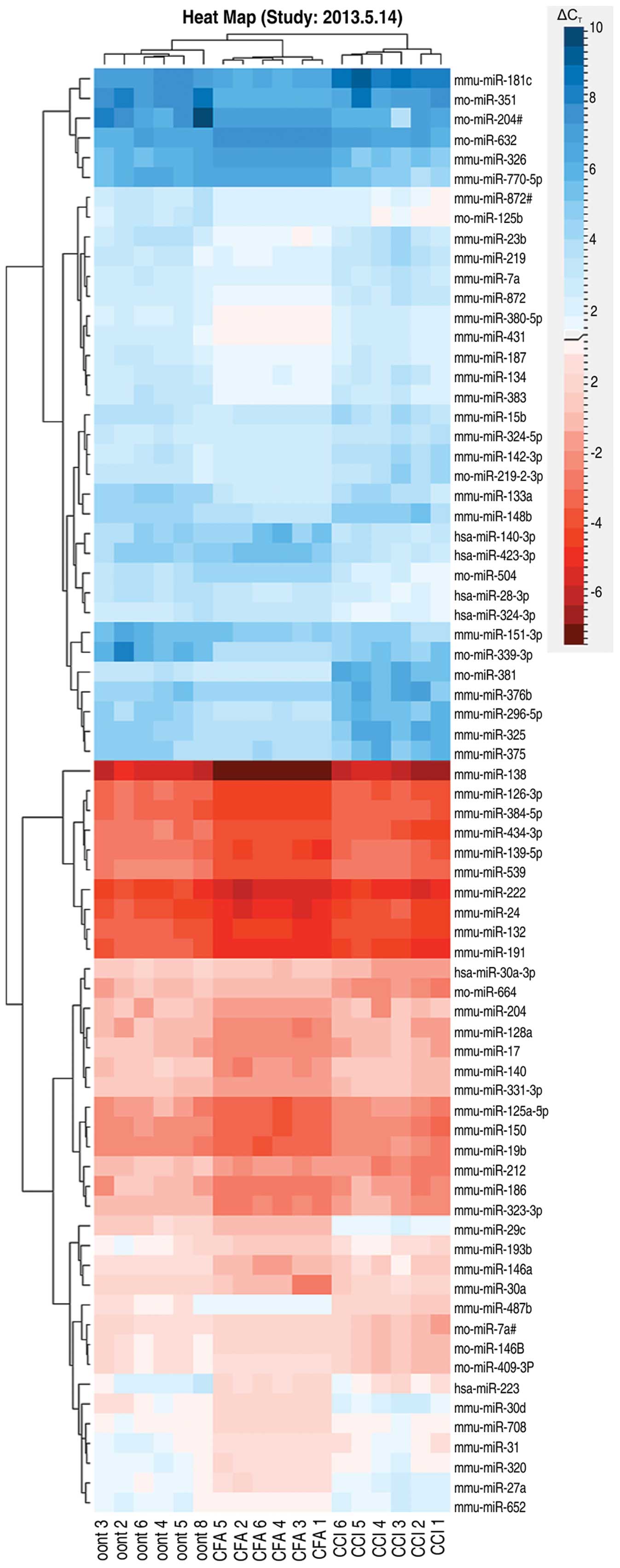
(Fig. 4). A clustergram of the
samples and the miRNAs that showed significant differences are
shown as a heat map in Fig. 3. As
can be observed in the figure, the heat map indicates 3 main
branches (control and experimental) that separate the CCI from the
CFA group.
Discussion
In the present study, we determined that hippocampal
miRNA expression differed in two pain models that have similar
symptoms (CCI and CFA models). Furthermore, we demonstrated that
miRNA expression did not differ between the left and right
hippocampus.
Pain hypersensitivity and
dysesthesia
In both the CCI and CFA models, we observed
hyperalgesia and mechanical allodynia on the injured side beginning
on the first post-operative day (day 1). Pain hypersensitivity
continued until day 14, which indicated that a similar chronic pain
was induced in the two models. In the CFA model, we observed
hyperalgesia and allodynia on the uninjured side beginning on day
3. We suspect that this was due to CFA-mediated inflammation that
was caused by systemic inflammation, which led to pain
hypersensitivity on the uninjured side.
miRNA expression
On day 7, when chronic pain was thought to be
complete, we investigated hippocampal miRNA expression using
cluster analysis.
Cluster analysis is a method used for grouping
samples same with very similar expression levels; it aids in the
identification of group-specific expression patterns. We observed
differential miRNA expression in the hippocampus. Furthermore, the
CCI and CFA group each produced differential miRNA expression
patterns.
The limbic system, which includes the hippocampus,
has been reported to be associated with memory and emotional
responses (19). Both the cause
and persistence of chronic pain is complex and cannot be easily
explained. Thus, differences at the miRNA level are likely
involved, even when the symptoms of chronic pain are identical.
Therefore, treatments for the different forms of chronic pain may
not be the same. We believe that our findings may lead to the
future assessment of treatment methods, and the development of
therapeutic agents with greater efficacies.
Difference between the left and right
hippocampus
Previous studies have indicated a correlation
between chronic pain and the hippocampus (20). Although previous studies have
examined anatomical differences between the hippocampus
bilaterally, as well as the effect of peripheral pain on the
hippocampus, the responsible pathways remain unelucidated.
Therefore, we in this study, investigated how unilateral chronic
pain affects the hippocampus bilaterally. In addition, we examined
the hypothesis that a model of inflammatory pain would induce a
systemic reaction, but not bilateral differences in the
hippocampus. In both the CCI and CFA groups, we searched for
bilateral differences in miRNA expression that showed significant
changes. We found no evidence that indicated a difference between
the hippocampus bilaterally. We concluded that the bilateral
hippocampal miRNA expression is equal in the two chronic pain
models.
Although the hippocampus functions bilaterally, it
is widely known that its asymmetrical nature is an essential
feature for high-order brain functioning (21,22). Bilateral differences in
hippocampal projections related to chronic pain have not been
investigated thus far. Although bilateral differences in miRNA
expression have been previously demonstrated in the spinal cord, in
this study, to our knowledge, we show for the first time that there
was no difference in miRNA expression between the left and right
hippocampus.
Symptomatic bilateral differences disappeared in the
model of inflammatory pain (CFA group), but remained in the CCI
model. In both models, however, we observed no differences between
the left and right hippocampus. In the pain pathway, the left and
right sides of the hippocampus are thought to be morphologically
connected or in a compartment, and thus, symptomatic bilateral
differences were not affected.
In conclusion, two chronic pain models that show
similar pain actions also showed differential changes in
hippocampal miRNA expression patterns. Such a finding indicates
that each model may possess a unique mechanism for regulating mRNA
and protein expression. We observed no differences in miRNA
expression between the left and right hippocampus. Thus, we believe
that clarifying the mechanisms underlying pain development may lead
to improved treatment techniques, as well as to the development of
therapeutic agents with greater efficacies.
References
|
1
|
Lee HL, Lee KM, Son SJ, Hwang SH and Cho
HJ: Temporal expression of cytokines and their receptors mRNAs in a
neuropathic pain model. Neuroreport. 15:2807–2811. 2004.PubMed/NCBI
|
|
2
|
Urban MO and Gebhart GF: Supraspinal
contributions to hyperalgesia. Proc Natl Acad Sci USA.
96:7687–7692. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kuss AW and Chen W: MicroRNAs in brain
function and disease. Curr Neurol Neurosci Rep. 8:190–197. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Farh KK, Grimson A, Jan C, Lewis BP,
Johnston WK, et al: The widespread impact of mammalian MicroRNAs on
mRNA repression and evolution. Science. 310:1817–1821. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, et al: Microarray analysis shows that some
microRNAs downregulate large numbers of target mRNAs. Nature.
433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gozzi A, Crestan V, Turrini G, Clemens M
and Bifone A: Antagonism at serotonin 5-HT(2A) receptors modulates
functional activity of frontohippocampal circuit.
Psychopharmacology (Berl). 209:37–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gemma C, Imeri L and Opp MR: Serotonergic
activation stimulates the pituitary-adrenal axis and alters
interleukin-1 mRNA expression in rat brain.
Psychoneuroendocrinology. 28:875–884. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Norman GJ, Karelina K, Zhang N, Walton JC,
Morris JS and Devries AC: Stress and IL-1beta contribute to the
development of depressive-like behavior following peripheral nerve
injury. Mol Psychiatry. 15:404–414. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Klein AB, Santini MA, Aznar S, Knudsen GM
and Rios M: Changes in 5-HT2A-mediated behavior and 5-HT2A- and
5-HT1A receptor binding and expression in conditional brain-derived
neurotrophic factor knock-out mice. Neuroscience. 169:1007–1016.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bordukalo-Niksic T, Mokrovic G, Stefulj J,
Zivin M, Jernej B and Cicin-Sain L: 5HT-1A receptors and
anxiety-like behaviours: studies in rats with constitutionally
upregulated/downregulated serotonin transporter. Behav Brain Res.
213:238–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bennett GJ and Xie YK: A peripheral
mononeuropathy in rat that produces disorders of pain sensation
like those seen in man. Pain. 33:87–107. 1988. View Article : Google Scholar
|
|
12
|
Zhang W, Liu LY and Xu TL: Reduced
potassium-chloride co-transporter expression in spinal cord dorsal
horn neurons contributes to inflammatory pain hypersensitivity in
rats. Neuroscience. 152:502–510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Okabe T, Sato C and Sakamoto A: Changes in
neuropeptide Y gene expression in the spinal cord of chronic
constrictive injury model rats after electroconvulsive stimulation.
Biomed Res. 31:287–292. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sato C, Sakai A, Ikeda Y, Suzuki H and
Sakamoto A: The prolonged analgesic effect of epidural ropivacaine
in a rat model of neuropathic pain. Anesth Analg. 106:313–320.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiu K, Lau WM, Lau HT, So KF and Chang
RC: Micro-dissection of rat brain for RNA or protein extraction
from specific brain region. J Vis Exp. 7:2692007.PubMed/NCBI
|
|
16
|
Wang B, Howel P, Bruheim S, Ju J, Owen LB,
Fodstad O and Xi Y: Systematic evaluation of three microRNA
profiling platforms: microarray, beads array, and quantitative
real-time PCR array. PLoS One. 6:e171672011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hui AB, Shi W, Boutros PC, et al: Robust
global micro-RNA profiling with formalin-fixed paraffin-embedded
breast cancer tissues. Lab Invest. 89:597–606. 2009. View Article : Google Scholar
|
|
18
|
Ishikawa M, Tanaka S, Arai M, Genda Y and
Sakamoto A: Differences in microRNA changes of healthy rat liver
between sevoflurane and propofol anesthesia. Anesthesiology.
117:1245–1252. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Devinsky O, Morrell MJ and Vogt BA:
Contributions of anterior cingulate cortex to behavior. Brain.
118:279–306. 1995.PubMed/NCBI
|
|
20
|
Arai M, Genda Y, Ishikawa M, Shunsuke T,
Okabe T and Sakamoto A: The miRNA and mRNA changes in rat
hippocampi after chronic constriction injury. Pain Med. 14:720–729.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Corballis MC: From mouth to hand: gesture,
speech, and the evolution of right-handedness. Behav Brain Sci.
26:199–208. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cooke J: Developmental mechanism and
evolutionary origin of vertebrate left/right asymmetries. Biol Rev
Camb Philos Soc. 79:377–407. 2004. View Article : Google Scholar : PubMed/NCBI
|