Introduction
Head and neck squamous cell carcinoma (HNSCC) is
comparatively frequent type of cancer worldwide and is associated
with a relatively poor prognosis despite recent improvements in
surgical and radiation therapy techniques. The majority of patients
with HNSCC present with locally and/or regionally advanced disease
which further hinders the clinical outcome, even with modern
multi-disciplinary treatment approaches.
The status of cervical lymph nodes is the primary
indicator of poor prognosis in patients with HNSCC, while distant
metastatic disease remains a major cause of mortality. Therefore,
it is critical to develop a deeper understanding of the genetic
determinants of metastasis in order to develop more effective
therapeutic approaches.
Epithelial-to-mesenchymal transition (EMT) is a
cornerstone in the molecular steps of the metastatic process,
although originally, it is a morphogenetic program of embryonic
development, through which epithelial cells are converted into
mesenchymal cells. During the process of EMT, the disruption of
cell-to-cell junctions and the loss of apical-basal polarity
results in the formation of mesenchymal cells with migratory and
invasive abilities (1).
The main molecular hallmark of the EMT process is
the loss of E-cadherin expression, while ZEB1 (also known as dEF1,
TCF8, AREB6 or Zfhx1a) and ZEB2 (also known as SIP1 or Zfhx) are Zn
finger transcription factors categorized to be master regulators of
EMT by directly repressing E-cadherin (2). Currently, various transcription
factors regulating E-cadherin transcription are also a matter of
active investigation.
MicroRNAs (miRNAs or miRs) are endogenous small
non-coding RNAs that modulate the expression of target genes
through the repression of translational or mRNA degradation. They
play significant roles in the regulation of biological and
pathological cellular process, including differentiation,
proliferation, apoptosis and metastasis. Multiple miRNAs have also
been reported to be involved in the pathogenesis of HNSCC (3).
An important regulatory link between the ZEB family
and various species of miRNA has been established. The miRNA-200
(miR-200) family has been identified as a major regulator of EMT
through the modulation of the expression of E-cadherin (4). The miR-200 family consists of five
members, which form two clusters based on the seed sequences
related with binding specificities: miR-200a/141 and
miR-200b/200c/429 (5). They
enforce the expression of E-cadherin by regulating the
transcriptional repressors, ZEB1 and ZEB2. Recent findings suggest
that the balanced expression between ZEB factors and the miR-200
family based on the reciprocal control of each other regulate EMT
and the progression of various types of cancer towards metastasis
(6). However, the effects of the
miR-200 family on EMT and the metastasis of HNSCC cells have not
been fully characterized yet.
In the present study, we evaluated the role of
miR-200c/mir-141 in the progression EMT and metastasis in HNSCC.
Our results revealed a significant reciprocal correlation between
miR-200c/mir-141 and their targets, ZEB1 and ZEB2. On the other
hand, we also noted discrepancies in some of the HNSCC cell lines
in response to the enforced expression of miR-200c/mir-141.
Therefore, we further focused on the epigenetic control of
ZEB1/ZEB2 by promoter hypermethylation, and its association with
miR-200c/mir-141 in HNSCC.
Materials and methods
Cell lines and cultures
UTSCC cell line series were kindly provided by Dr R.
Grenman (Department of Otolaryngology, Turku University, Turku,
Finland). The UTSCC cells were grown in RPMI-1640 medium
supplemented with 10% fetal bovine serum, 1%
penicillin/streptomycin and 1-μl/ml amphotericin B
(Gibco™/Invitrogen, Tokyo, Japan), and all cell lines were cultured
in a humidified incubator with 5% CO2 at 37°C. In the
definition of UTSCC cell lines, ‘A’ represents the cell lines
derived from primary tumors (e.g., UTSCC-6A cells), while ‘B’
represents the cells derived from metastatic tumors (e.g., UTSCC-6B
cells).
mRNA processing and quantitative reverse
transcription PCR (qRT-PCR)
The preparation of cDNA from mRNA was performed
directly from cultured cell lysates using the TaqMan®
Gene Expression Cells-to-CT™ kit (Ambion, Tokyo, Japan), according
to manufacturer’s instructions. Briefly, cultured cells in 96-well
plates were washed in PBS and lysed with lysis buffer and RNA was
released into this solution. Cell lysates were reverse transcribed
into cDNA using the RT Enzyme Mix and appropriate RT buffer
(Ambion). Finally the cDNA was amplified by qRT-PCR using the
included TaqMan Gene Expression Master Mix and the specific TaqMan
primer/probe assay designed for the investigated genes: E-cadherin
(Hs01023894_m1), ZEB1 (Hs00232783_m1), ZEB2 (Hs00207691_m1) and
GAPDH (Hs99999905_m1) (Applied Biosystems, Tokyo, Japan).
The gene expression levels were normalized to the
expression of the housekeeping gene, GAPDH, and were expressed as
fold changes relative to the expression of the untreated cells. The
amplification profile was initiated by 10 min of incubation at
95°C, followed by two-step amplification of 15 sec at 95°C and 60
sec at 60°C for 40 cycles. All experiments were performed with
non-template controls included to exclude reagent contamination.
Quantification was performed with the ΔΔCt calculation method
(7). For the determination of the
absolute quantity of target genes, a standard curve was generated
using ten-fold serial dilutions of a synthetic RNA duplex. Each
sample was analyzed in triplicate on the 7500 Real-time PCR system
in a 96-well plate (Applied Biosystems).
MicroRNA processing and qRT-PCR
Preparation of cDNA from miRNA was also performed
directly from cultured cell lysates using the TaqMan MicroRNA
Cells-to-CT kit (Ambion) according to the manufacturer’s
instructions. Briefly, the cultured cells in 96-well plates were
washed in PBS and lysed with lysis buffer and RNA was released into
this solution. Cell lysates were reverse transcribed into cDNA
using the RT Enzyme Mix, RT buffer and appropriate RT primer
(Applied Biosystems). Finally the cDNA of each mature miRNA was
amplified separately by qRT-PCR using the TaqMan Universal Master
Mix and the specific primer and probe mix included in pre-designed
TaqMan MicroRNA Assays: hsa-miR-200c, ID: 000385; and hsa-miR-141,
ID: 000385 (Applied Biosystems). The quantification of miRNA
expression was performed with the same method as described above
and for normalizations RNU6B RNA (RNU6B; ID: 001093, P/N: 4373381,
Applied Biosystems) was used as an internal control. Each sample
was analyzed in triplicate.
Cell transfection with miR-200c and
miR-141 precursors
The stability-enhanced miR-200c, miR-141 precursors
and the negative control, 1 ribo-oligonucleotides, were obtained
from Ambion. Transfection of miRNAs was carried out using siPORT
Amine in accordance with the manufacturer’s instructions (Ambion).
Briefly, the UTSCC cell lines were seeded (8×105 cells
in 4 ml of RPMI-1640 per dish) in 96-well culture dishes and grown
overnight (the day prior to transfection) in antibiotic-free
medium. The transfection of miR-200c and miR-141 precursors, or
each negative control (all purchased from Applied Biosystems) at
the indicated concentrations was introduced into the cells using
0.3 μl siPORT Amine Transfection Agent (Applied Biosystems) in 20
μl Opti-MEM (Gibco/Invitrogen) according to the manufacturer’s
recommendations. The negative controls were scrambled
oligonucleotides that were validated not to produce identifiable
effects on known miR functions. The level of miR-200c and miR-141
expression in the transfected cells was confirmed by qRT-PCR
(Taqman MicroRNA Assays) 24 h after transfection as described
above.
Treatment with
5-aza-2′-deoxycytidine
Treatment with 5-aza-2′-deoxycytidine (5-Aza-CdR)
(Sigma-Aldrich, Tokyo, Japan) was carried out as described in a
previous study from our group (8). Briefly, the cells were incubated for
72 h with 4 μM 5-Aza-CdR, and then harvested for RNA extraction and
qRT-PCR. For anti-miRNA transfection analysis, 48 h after being
treated with 1 μM 5-Aza-CdR, the cells were transfected for one day
and then treated again with 5-Aza-CdR for a further 24 h.
Cell proliferation assay
Cell proliferation was assessed by the
4-[3-(4-iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio]-1,3-benzene
disulfonate (WST-1) tetrazolium salt assay (Roche, Japan). The
UTSCC cells were plated at a density of 6×103 cells/well
on 96-well plates and grown overnight. The miR-200c, miR-141
precursors or the negative control were then introduced into the
cell lines as described above and 24 h later, the assay was
initiated by the addition of 20 μl of WST solution reagent to 100
μl of culture medium for each well. Following incubation for 3 h at
37°C, the plates were read on a microplate autoreader (Corona
MTP-450) at a wavelength of 450 nm. The results were expressed as
the mean optical density for selected paradigms performed in
duplicate.
Transwell invasion assay and migration
assay
The UTSCC cells were seeded at a density of
4.0×104cells/well on 24-well plates, and 24 h later, the
cells were transfected separately with 50 nM miR-200c precursor,
miR-141 precursor or the scrambled negative control. After 24 h,
the transfected cells were harvested by trypsinization, and washed
twice in PBS, and 8.0×104 cells were transferred to the
upper chamber of a BioCoat™ Matrigel™ Invasion Chamber (BD
Biosciences, Tokyo, Japan) with inserts containing an
8-μm-pore-sized membrane with a thin layer of Matrigel in the
24-well Transwell plate filled with 500 μl serum-free RPMI-1640
medium. In the lower chamber, 500 μl of the 10% FBS-containing
medium containing and 10 μg/ml fibronectin were added. Following
incubation for 24 h, the invaded cells were counted under a
microscopic using a Diff-Quick staining kit (Sysmex, Kobe,
Japan).
For the migration assay, the cells transfected with
miRNA were added to the upper chamber of a Boyden chamber (BD
Falcon cell culture insert 24-well 8 μm pole, no. 353097; BD
Biosciences). Fibronectin (10 μg/ml) and medium with 10% FBS 750 μl
are added to the lower chamber. Following incubation for 36 h at
37°C, the invaded cells were counted under a microscopic using a
Diff-Quick staining kit (Sysmex).
Wound healing assay
The UTSCC cells were transfected separately with
miR-200c precursor, miR-141 precursor or the scrambled negative
control. When cell confluence reached approximately 80% at 48-h
post-transfection, wounds were created in confluent cells using a
200-μl pipette tip. The cells were then rinsed with medium to
remove any free-floating cells and debris. Medium was then added,
and the culture plates were incubated at 37°C. Wound healing was
observed at different time points within the scrape line, and
representative scrape lines were photographed. Duplicate wells for
each condition were examined, and each experiment was repeated
three times.
Statistical analysis
Data were analyzed using GraphPad Prism 5.0
software. To evaluate significant differences between two matched
pair groups or between two independent groups of samples, paired
t-tests and the Mann-Whitney U test were used, respectively.
Pearson’s correlation coefficient (r) was used to measure
correlation, and logarithmic regression was used to calculate the
R2 and to create the equation of the slope.
Results
The miR-200 family includes five members, namely
miR-200a, miR-200b, miR-200c, miR-141 and miR-429. The miR-200
family can be grouped into two clusters according to seed sequences
related with target specifications: miR-200a/miR-141 and
miR-200b/miR-200c/miR-429. In this study, we selected miR-200c and
miR-141 as representatives of each cluster. Furthermore, both
miR-200c and miR-141 are located on the same genetic loci of
chromosome 12 (9).
Expression status of E-cadherin, ZEB1 and
ZEB2 in HNSCC
To determine the EMT status of each cell line, we
evaluated the mRNA expression of E-cadherin as the key epithelial
marker and that of ZEB1 and ZEB2 as mesenchymal markers in 13 head
and neck cell carcinoma lines belonging to the UTSCC series
originating from primary or metastatic tumors. The results of
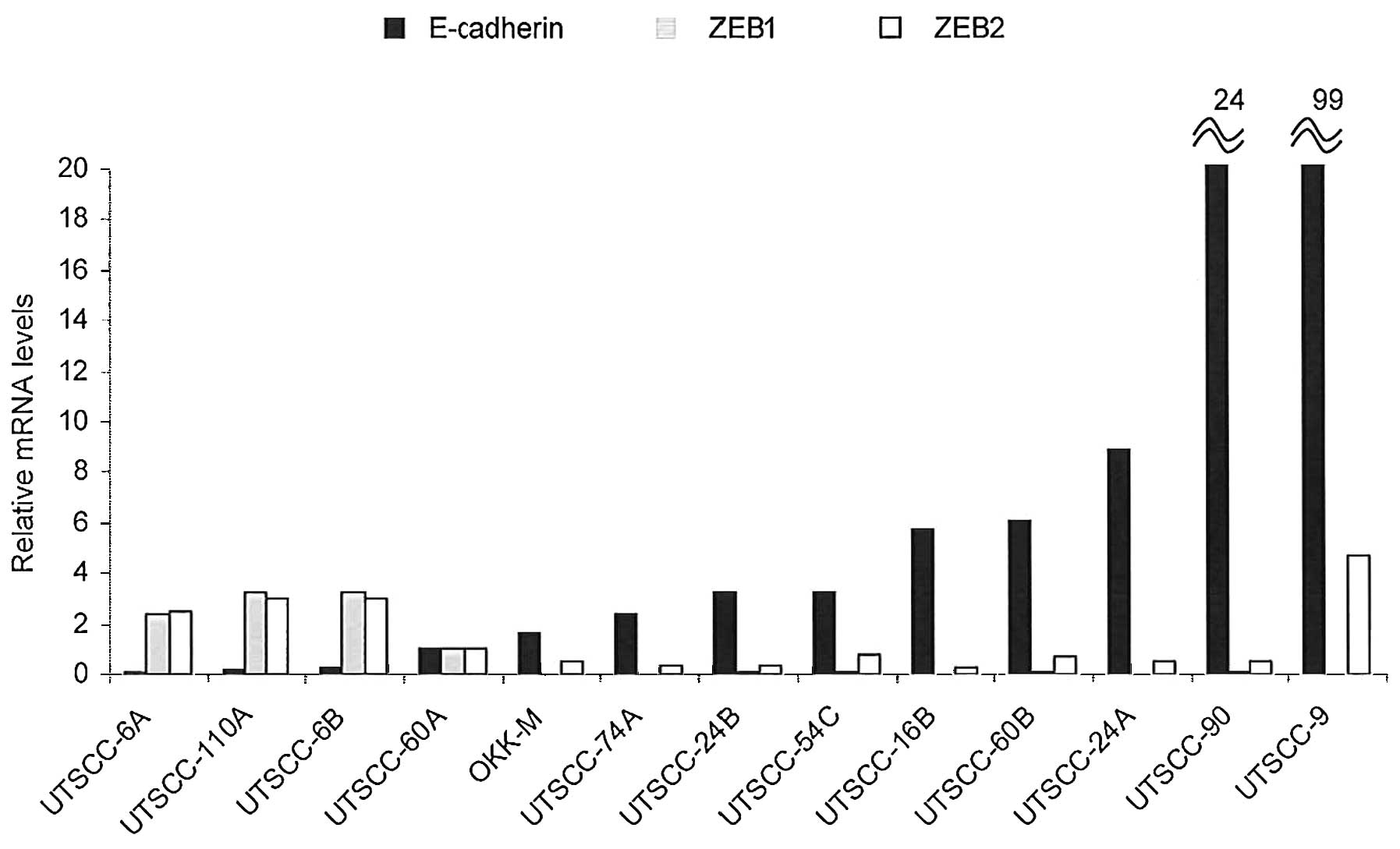
expression analysis are displayed in Fig. 1. The quantification of mRNA
expression was performed by a standard curve based on the serial
dilutions of the mRNA derived from the UTSCC-60A cell line.
Comparatively, nine of the 13 (70%) cell lines showed a
substantially increased expression of E-cadherin in contrast to the
decreased expression of both ZEB1 and ZEB2. On the other hand, the
remaining three (30%) cell lines showed an inverse trend of
expression results. Among the three pairs of cell lines originating
from primary and corresponding metastatic tumors (UTSCC-6A-B,
UTSCC-24A-B and UTSCC-60A-B), the cell lines originating from
metastatic tumors displayed a lower E-cadherin and higher ZEB1/ZEB2
expression in only one pair of cell lines (UTSCC-60A-B), in
contrast to the two other pairs of cell lines in which converse
results were obtained (Fig. 1).
The origin of the cells (namely from primary tumors or metastastic
tumors) was not found to be directly associated with the expression
levels of E-cadherin or ZEB1/ZEB2. These results suggest that the
EMT status is not constantly related with the origin of the tumor
cells (from primary or metastastic tumors).
Correlation of E-cadherin, ZEB1 and ZEB2
expression with miR200c/miR-141
In order to better understand the association
between the expression patterns of EMT markers and the miR-200
family, we analyzed the expression of miR-200c and miR-141 by
qRT-PCR in the panel of 13 UTSCC cell lines and assessed the
correlation between miR-200c/miR-141 and EMT markers, including the
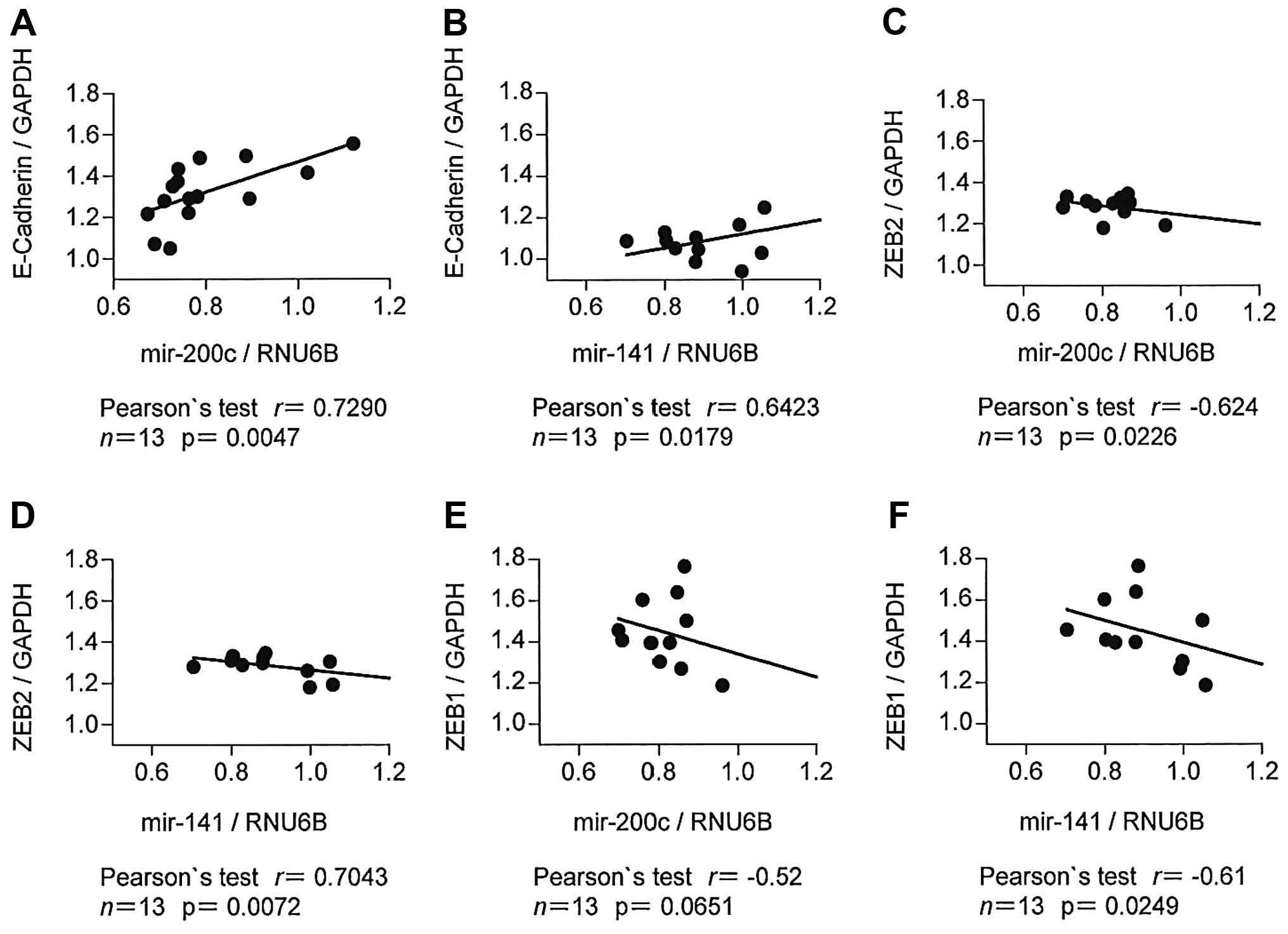
E-cadherin, ZEB1 and ZEB2 genes. The mRNA expression of miR-200c
(r=0.73, p<0.01) and miR-141 (r=0.64, p=0.02) showed a
statistically significant positive correlation with E-cadherin
expression (Fig. 2A and B).
Conversely, both miR-200c (r=−0.62, p=0.02) and miR-141 (r=−0.70,
p<0.01) showed a significant inverse correlation with ZEB2
expression (Fig. 2C and D).
Accordingly, miR-141 showed a significant inverse correlation with
ZEB1 (r=−0.61, p=0.02), while the correlation between miR-200c and
ZEB1 was not significant (r=−0.52, p=0.07) (Fig. 2E and F). Taken together, these
data indicate that the expression of miR-200c and miR-141 is
associated with the acquisition of epithelial characteristics and
the simultaneous loss of mesenchymal features in the HNSCC cells,
the two distinguishing features that are essential for the
development of a metastatic phenotype.
Overexpression of miR200c/miR-141
inhibits the expression of ZEB1, while inducing that of
E-cadherin
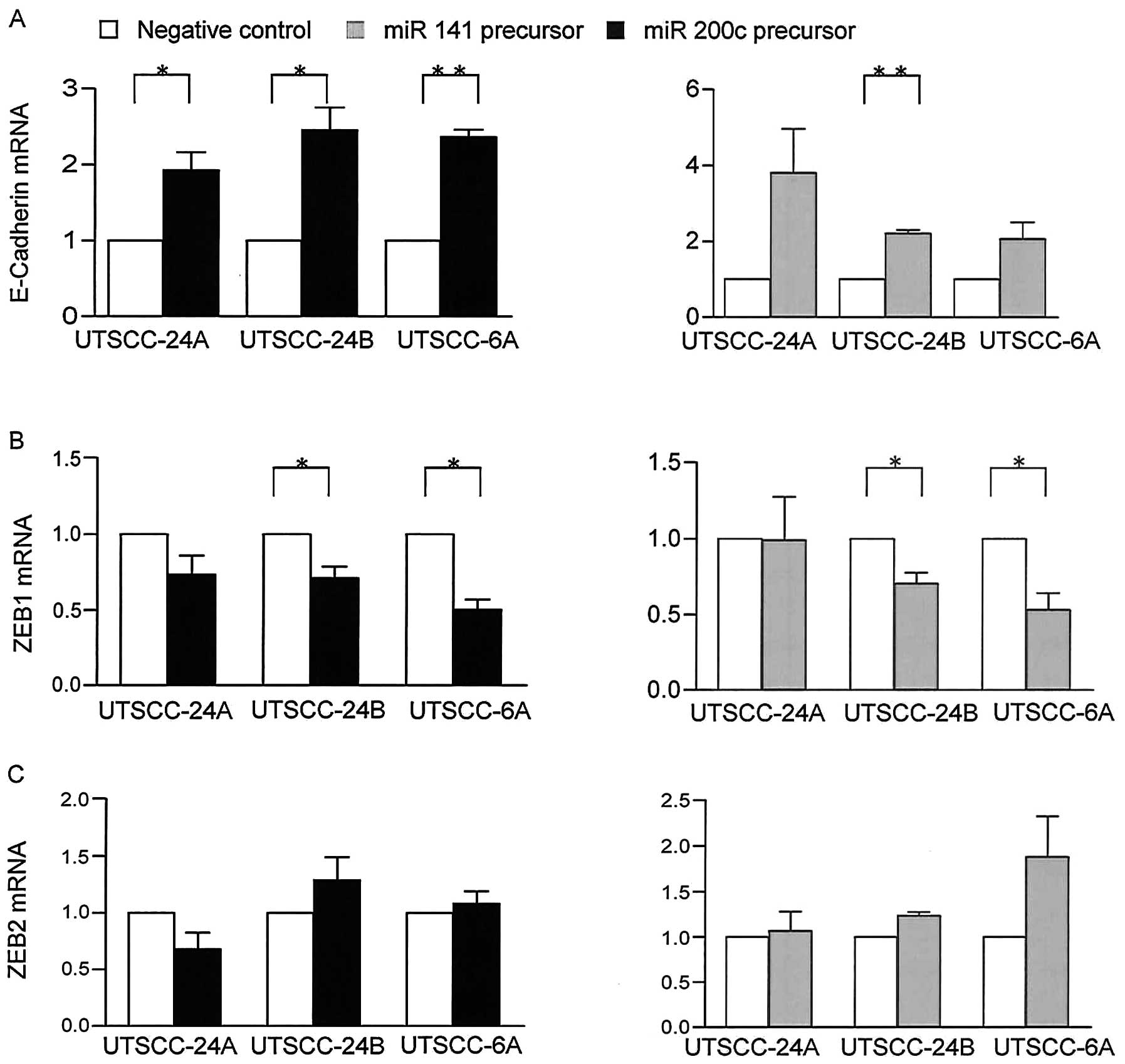
To further investigate the association between the
miR-200 family and EMT markers, we analyzed the effects of the
enforced expression of miR-200c and miR-141 on E-cadherin, ZEB1 and
ZEB2 gene epxression by transfecting the UTSCC-24A and UTSCC-24B
cells with precursor miRNAs. Following transfection with miRNA
precursors, the enforced expression of miR-200c/miR-141 was
confirmed by qRT-PCR. The UTSCC-24B cells had substantially lower
levels of endogenous miR-200c and mir-141 compared with the
UTSCC-24A cells. The enforced expression of miR-200c and mir-141
resulted in the significant upregulation of E-cadherin expression
compared with the control cells (p=0.03 and p=0.10 in UTSCC-24A
cells) and (p=0.02 and p<0.01 in UTSCC-24B cells, paired t-test)
(Fig. 3A). By contrast, ZEB1
showed a significant downregulation under the same conditions
(p=0.01 and p=0.44 in UTSCC-24A cells) and (p=0.03 and p=0.03 in
UTSCC-24B cells, paired t-test) by qRT-PCR (Fig. 3B). On the other hand, the
expression of ZEB2 remained almost unaltered (p>0.05), (Fig. 3C). These findings suggest that
ZEB1 plays a prominent role as opposed to ZEB2, in the regulation
of E-cadherin by the mir-200 family in HNSCC.
To confirm these results, we also examined another
pair of cells (UTSCC-60A and UTSCC-60B), using same experimental
conditions. In contrast to the UTSCC-24A and UTSCC-24B cells, the
UTSCC-60A and UTSCC-60B cells did not show a significant change in
the expression of E-cadherin or ZEB1/ZEB2 in response to the
enforced expression of miR-200c and miR-141. These results suggest
that the effects of the miR-200 family on EMT-related target gene
expression may be hindered by another epigenetic mechanism rather
than the one involving miRNAs.
Hypermethylation of ZEB1 and ZEB2 blocks
the modulation by miR-200c/miR-141 in HNSCC
The expression of ZEB2 in both UTSCC pairs of cell
lines and ZEB1 in one pair of cell lines failed to be suppressed by
the enforced expression of miR-200c and miR-141. To address this
issue, we aimed to identify the effects of other epigenetic
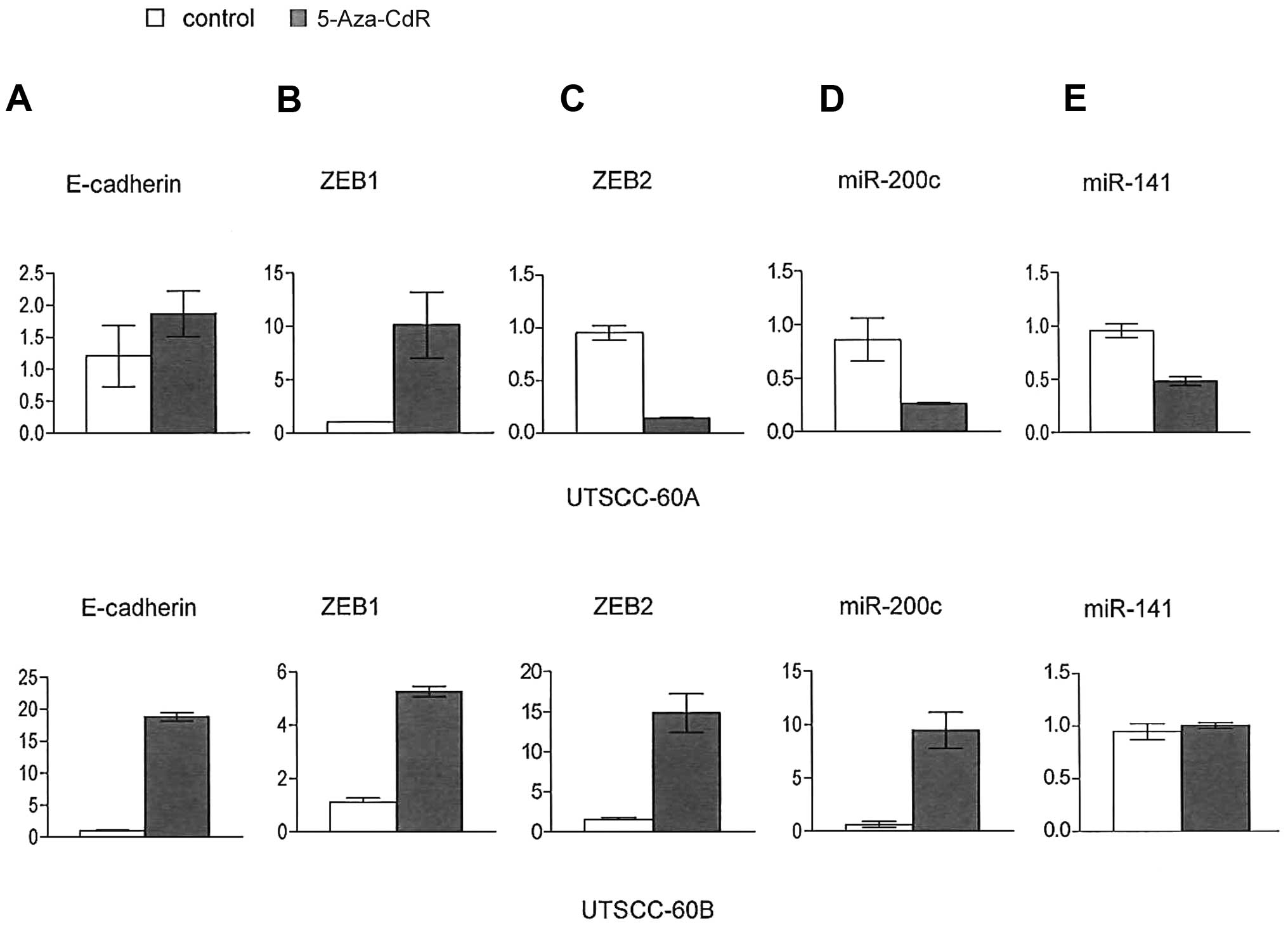
mechanisms in the regulation of ZEB1/ZEB2. Both ZEB1 and ZEB2 genes
have wide CpG islands in their promoter regions. To clarify whether
the CpG methylation of ZEB1 and ZEB2 gene promoters is involved in
their transcriptional repression, we examined the expression mRNA
before and after treatment with 5-Aza-CdR, a DNA demethylating
agent, by qRT-PCR in the UTSCC-60A and UTSCC-60B cells. We observed
a marked upregulation in both ZEB1 and ZEB2 mRNA expression
following treatment with 5-Aza-CdR in both cell lines, while the
expression of E-cadherin was also increased, although to a lesser
degree (Fig. 4A–C). On the other
hand, mir-200c and mir-141 expression was decreased in those cells
(Fig. 4D and E). These results
suggest that the effects of the miR-200 family on EMT-related gene
expression are blocked by the methylation of target gene
promoters.
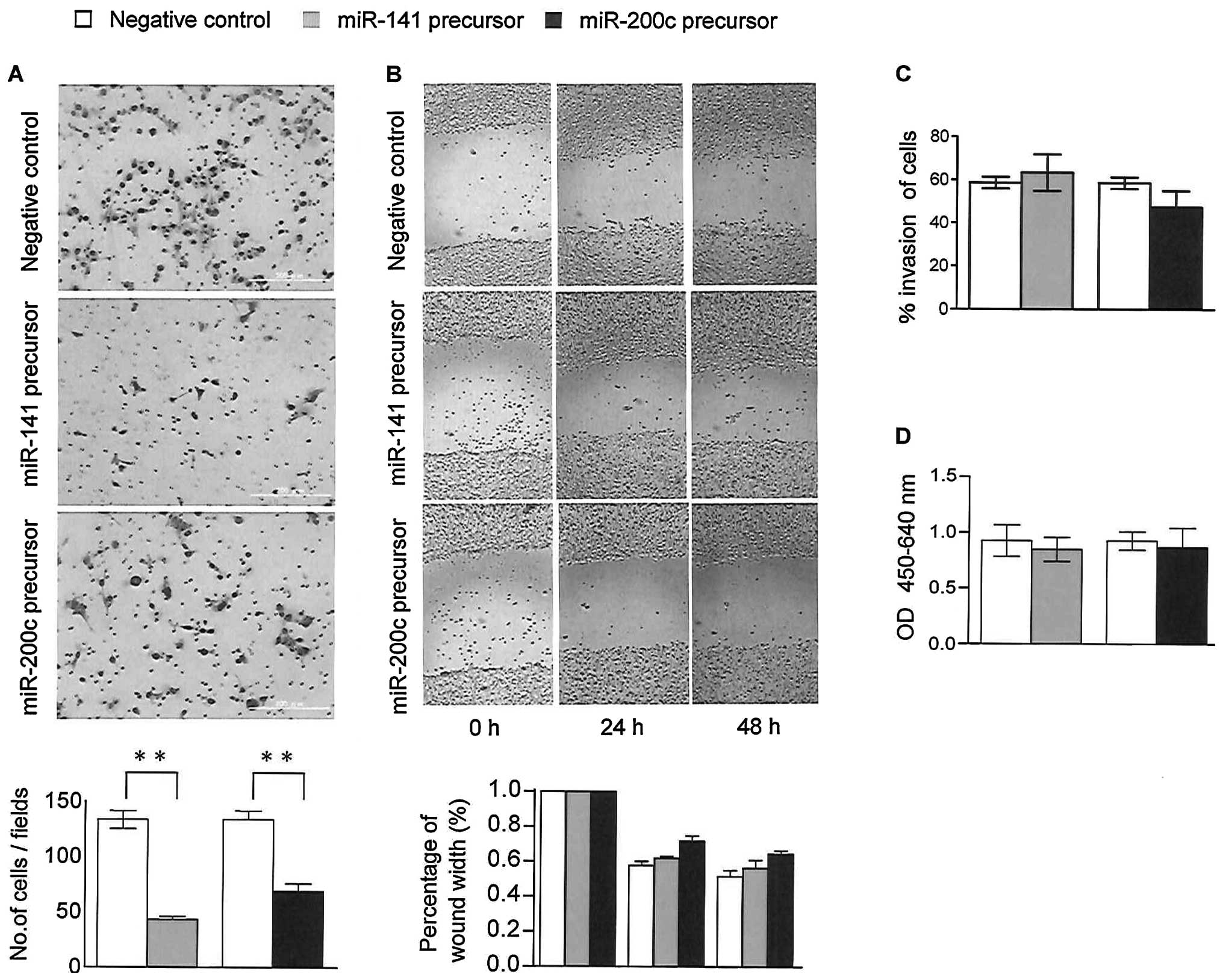
Overexpression of miR200c/miR-141 impairs
HNSCC cell migration but not cell invasion and proliferation
To determine the role of the microRNA-200 family in
the metastatic process, we assessed the effects of the induced
expression of miR-200c and miR-141 on cell motility and invasion by
transfecting the UTSCC-24A and UTSCC-24B cells with precursor
microRNAs. Cell migration assays demonstrated that the
overexpression of miR-200c and miR-141 significantly reduced the
migration capacity of both cell lines compared to the controls
(p<0.01; Fig. 5A). Similarly,
a wound healing assay confirmed these results in 48-h time course
of transfection compared to the controls (Fig. 5B). On the other hand, Matrigel
invasion assay revealed no significant change in the invasion
abilities of the cells in response to the overexpression of
miR-200c and miR-141 (Fig.
5C).
To identify the role of the microRNA-200 family in
the progression of HNSCC, we determined whether miR-200c and
miR-141 may also affect the proliferation of HNSCC cells. In the
WST assay, the proliferation rates of the UTSCC-24A and UTSCC-24B
cells transfected with precursor miRNAs were not altered
significantly compared to the control cells (p>0.2) (Fig. 5D). These results indicate that the
miR-200 family inhibits the migration of HNSCC cells, independent
of cell invasion and proliferation in vitro.
Discussion
In this study, we demonstrated that the miR-200
family plays an essential role both in the suppression of the EMT
process and in the metastatic ability of HNSCC cells. Our results
highlight the importance of the upregulation of the
miR-200c/miR-141 cluster in the prevention of the metastatic
ability of the HNSCC cells. On the other hand, we also observed
that other mechanisms of epigenetic silencing, such as the promoter
hypermethylation of target genes may disrupt the role of the
miR-200 family in the regulation of EMT-related gene
expression.
We observed that ZEB2 mRNA expression was not
suppressed by the enforced expression of mir-200c/miR-141 in all
four HNSCC cell lines examined. Similarly, ZEB1 mRNA expression was
not inhibited in one group of HNSCC cells by mir-200c/miR-141.
Since wide CpG islands exist in the promoter regions of both genes,
the suppression of expression by hypermethylation seems to be a
possible mechanism to explain this condition. Our expression
analysis after inhibiting DNA methylation by 5-Aza-CdR revealed a
marked upregulation in the mRNA expression of ZEB1 and ZEB2. To our
knowledge, this is the first study to demonstrate that both ZEB1
and ZEB2 genes have already been suppressed by hypermethylation,
and are able to block the regulatory role of the miR-200 family.
Although a number of studies have defined the status of
hypermethylation in the promoter region of the miR-200 family
itself (10–12), the role of methylation in the
promoter regions of their target genes has not yet been fully
elucidated. To date, only one study investigated the association
between miR-200a/miR-200b and the hypermethylation of ZEB2 in
pancreatic carcinomas. Li et al found that the majority of
pancreatic cancers highly express miR-200a and miR-200b; however,
this expression does not affect ZEB2 expression, since the promoter
of ZEB2 has already been silenced by hypermethylation (13). Similarly, the promoter methylation
of ZEB2 was also identified in two other types of epithelial
cancers, including breast carcinoma (14) and hepatocellular carcinoma
(15). However, the methylation
status of the ZEB1 promoter was not investigated in these studies.
Previously, Hidaka et al observed the activation of ZEB1
expression in adult T-cell leukemia/lymphoma (ATLL) cell lines
follwoing treatment with the DNA methylation agent, 5-Aza-CdR
(16); however, the association
between the miR-200 family and ZEB1 was not investigated.
DNA methylation (transcriptional level) and miRNA
modulation (post-transcriptional level) are two major epigenetic
regulatory mechanisms, which affect opposite sites of genes at the
5′ promoter and 3′ untranslated regions, respectively. A number of
studies have investigated each epigenetic mechanism separately
(10–12,14); however, the combined effects of
both mechanisms have not yet been elucidated. A recent
bioinformatics analysis demonstrated that there is a complementary
regulation between these two mechanisms at the genome level, while
cancer-related genes have a low methylation status and more miRNA
target sites (17). Thus, the
role of promoter methylation of target genes in the modulation by
the miR-200 family in the EMT process in HNSCC and other types of
cancer requires further investigation.
In EMT, E-cadherin is a representative marker of
epithelial transition, while ZEB1 and ZEB2 inhibit E-cadherin
expression and induce mesenchymal transition in cancer progression.
Our preliminary survey revealed that there was a broad range in the
expression patterns of E-cadherin as opposed to those of ZEB1/ZEB2;
there was an inverse correlation between the expression of
E-cadherin and ZEB1/ZEB2 in the HNSCC cell lines. Our results
suggested that the EMT status differs between cancer cells,
occurring at different time points. Of note, the origin of the cell
lines (from primary or metastatic tumors) did not demonstrate a
clear association with the expression range of E-cadherin or
ZEB1/ZEB2. Consistent with these findings, Wiklund et al did
not identify any specific changes in miRNA expression profiles,
including the miR-200 family between non-metastatic and metastatic
tumors of oral squamous cell carcinoma (18). By contrast, Lo et al
demonstrated that miR-200c expression in the regional metastatic
lymph node of HNSCC tissues was significantly decreased compared
with primary tumors (19). In
literature, there are also controversial results concerning the
expression status of the miR-200 family and other EMT-related
factors between primary and metastatic tumors. To explain these
discrepancies, a recent study on breast carcinomas suggested a
dynamic dual role for the miR-200 family in the progression of
metastasis, hindering the entry of tumor cells into the
circulation, as opposed to promoting colonization in distant organs
of those cells that enter into the intravascular circulation
(20).
In this study, we demonstrated that miR-200c/miR-141
expression correlated with the increased E-cadherin and reduced
ZEB1/ZEB2 expression, suggesting that during cancer progression,
HNSCC cells initially lose epithelial features, while
simultaneously gaining the mesenchymal characteristics required for
EMT. We also showed that miR-200c/miR-141 suppressed the migration
ability of HNSCC cells. These results are consistent with those of
previous studies on other epithelial cancers (21). However, studies which
systematically analyze the correlation between the miR-200 family
and EMT in HNSCC are limited. Lo et al examined the role of
the miR-200 family in HNSCC cancer stem cells specifically and
revealed that the overexpression of miR-200c in those cells
downregulated ZEB1/ZEB2 expression and blocked the ability of the
cells to invade and metastasize (19). Another study compared the miR-200
family and desmosomal cadherins between spindle cell carcinoma and
squamous cell carcinoma of the head and neck region. They found
that the expression of the miR-200 family was significantly
downregulated in spindle cell carcinoma, while it was unaltered in
squamous cell carcinoma tumor tissues compared to normal mucosa
(22). Desmosomal cadherins,
including E-cadherin and N-cadherin also did not show a significant
change in expression in HNSCC. These findings suggest that only a
small part of the HNSCC cell population with a low miR-200 status
has the potential to undergo the EMT process.
One limitation of our study was that the experiments
were performed in vitro by using cell lines only. Cell lines
derived from human solid tumors may not adequately represent the
tumors of origin. Therefore, the results may need to be validated
by subsequent in vivo studies using primary tumor cells from
tissue samples of human tumors.
In conclusion, our molecular pathological findings
indicate that the expression of the miR-200 family positively
correlates with the epithelial marker, E-cadherin, while inversely
correlates with the mesenchymal markers, ZEB1 and ZEB2.
Furthermore, we found that the miR-200 family negatively regulates
the expression of ZEB1. These results confirm the reciprocal
balance between the miR-200 family and EMT in HNSCC. Notably, we
revealed that the promoter hypermethylation of ZEB1 and ZEB2 plays
an essential role and may overshadow the effects of the miR-200
family in the regulation of EMT during carcinogenesis. Thus, the
role of epigenetic mechanisms and their regulation concerning the
miR-200 family-ZEB1/ZEB2 feedback loop in EMT requires further
investigation.
Acknowledgements
We thank Dr Hirohashi for his comments and
suggestions regarding the manuscript. This study was partially
presented at the 8th International Conference on Head and Neck
Cancer. This study was partially supported by Grants-in-Aid for
scientific research from the Ministry of Education, Culture,
Sports, Science and Technology of Japan (to S.T. and L.B.B.).
References
|
1
|
Tsuji T, Ibaragi S and Hu GF:
Epithelial-mesenchymal transition and cell cooperativity in
metastasis. Cancer Res. 69:7135–7139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hill L, Browne G and Tulchinsky E:
ZEB/miR-200 feedback loop: At the crossroads of signal transduction
in cancer. Int J Cancer. 132:745–754. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tran N, McLean T, Zhang X, Zhao CJ,
Thomson JM, O’Brien C and Rose B: MicroRNA expression profiles in
head and neck cancer cell lines. Biochem Biophys Res Commun.
358:12–17. 2007. View Article : Google Scholar
|
|
4
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park SM, Gaur AB, Lengyel E and Peter ME:
The miR-200 family determines the epithelial phenotype of cancer
cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes
Dev. 22:894–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brabletz S and Brabletz T: The ZEB/miR-200
feedback loop - a motor of cellular plasticity in development and
cancer? EMBO Rep. 11:670–677. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Beder LB, Gunduz M, Hotomi M, Fujihara K,
Shimada J, Tamura S, Gunduz E, Fukushima K, Yaykasli K, Grenman R,
Shimizu K and Yamanaka N: T-lymphocyte maturation-associated
protein gene as a candidate metastasis suppressor for head and neck
squamous cell carcinomas. Cancer Sci. 100:873–880. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Griffiths-Jones S, Grocock RJ, van Dongen
S, Bateman A and Enright AJ: miRBase: microRNA sequences, targets
and gene nomenclature. Nucleic Acids Res. 1:D140–D144. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vrba L, Jensen TJ, Garbe JC, Heimark RL,
Cress AE, Dickinson S, Stampfer MR and Futscher BW: Role for DNA
methylation in the regulation of miR-200c and miR-141 expression in
normal and cancer cells. PLoS One. 5:e86972010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wiklund ED, Bramsen JB, Hulf T, Dyrskjøt
L, Ramanathan R, Hansen TB, Villadsen SB, Gao S, Ostenfeld MS,
Borre M, Peter ME, Ørntoft TF, Kjems J and Clark SJ: Coordinated
epigenetic repression of the miR-200 family and miR-205 in invasive
bladder cancer. Int J Cancer. 128:1327–1334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lim YY, Wright JA, Attema JL, Gregory PA,
Bert AG, Smith E, Thomas D, Lopez AF, Drew PA, Khew-Goodall Y and
Goodall GJ: Epigenetic modulation of the miR-200 family is
associated with transition to a breast cancer stem-cell-like state.
J Cell Sci. 126:2256–2266. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li A, Omura N, Hong SM, Vincent A, Walter
K, Griffith M, Borges M and Goggins M: Pancreatic cancers
epigenetically silence SIP1 and hypomethylate and overexpress
miR-200a/200b in association with elevated circulating miR-200a and
miR-200b levels. Cancer Res. 70:5226–5237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rodenhiser DI, Andrews J, Kennette W,
Sadikovic B, Mendlowitz A, Tuck AB and Chambers AF: Epigenetic
mapping and functional analysis in a breast cancer metastasis model
using whole-genome promoter tiling microarrays. Breast Cancer Res.
10:R622008. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Acun T, Oztas E, Yagci T and Yakicier MC:
SIP1 is downregulated in hepatocellular carcinoma by promoter
hypermethylation. BMC Cancer. 11:2232011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hidaka T, Nakahata S, Hatakeyama K,
Hamasaki M, Yamashita K, Kohno T, Arai Y, Taki T, Nishida K,
Okayama A, Asada Y, Yamaguchi R, Tsubouchi H, Yokota J, Taniwaki M,
Higashi Y and Morishita K: Down-regulation of TCF8 is involved in
the leukemogenesis of adult T-cell leukemia/lymphoma. Blood.
112:383–393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su Z, Xia J and Zhao Z: Functional
complementation between transcriptional methylation regulation and
post-transcriptional microRNA regulation in the human genome. BMC
Genomics. 12(Suppl 5): S152011. View Article : Google Scholar
|
|
18
|
Wiklund ED, Gao S, Hulf T, Sibbritt T,
Nair S, Costea DE, Villadsen SB, Bakholdt V, Bramsen JB, S⊘rensen
JA, Krogdahl A, Clark SJ and Kjems J: MicroRNA alterations and
associated aberrant DNA methylation patterns across multiple sample
types in oral squamous cell carcinoma. PLoS One. 6:e278402011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lo WL, Yu CC, Chiou GY, Chen YW, Huang PI,
Chien CS, Tseng LM, Chu PY, Lu KH, Chang KW, Kao SY and Chiou SH:
MicroRNA-200c attenuates tumour growth and metastasis of
presumptive head and neck squamous cell carcinoma stem cells. J
Pathol. 223:482–495. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Korpal M, Ell BJ, Buffa FM, Ibrahim T,
Blanco MA, Celià-Terrassa T, Mercatali L, Khan Z, Goodarzi H, Hua
Y, Wei Y, Hu G, Garcia BA, Ragoussis J, Amadori D, Harris AL and
Kang Y: Direct targeting of Sec23a by miR-200s influences cancer
cell secretome and promotes metastatic colonization. Nat Med.
17:1101–1108. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Korpal M, Lee ES, Hu G and Kang Y: The
miR-200 family inhibits epithelial-mesenchymal transition and
cancer cell migration by direct targeting of E-cadherin
transcriptional repressors ZEB1 and ZEB2. J Biol Chem.
283:14910–14914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zidar N, Bostjancic E, Gale N, Kojc N,
Poljak M, Glavac D and Cardesa A: Down-regulation of microRNAs of
the miR-200 family and miR-205, and an altered expression of
classic and desmosomal cadherins in spindle cell carcinoma of the
head and neck - hallmark of epithelial-mesenchymal transition. Hum
Pathol. 42:482–488. 2011. View Article : Google Scholar : PubMed/NCBI
|