Introduction
The volatile anesthetic, sevoflurane, is widely used
in surgery as there is extensive clinical data supporting its
safety (1). Over the years has
been a growing interest in the biological effects of sevoflurane on
tissue and organ systems and the molecular mechanisms involved. The
heart and the brain are the major target organs of sevoflurane;
sevoflurane can protect the heart against ischemic damage (2). Several signal transduction pathways,
such as the activation of G protein-coupled receptors, protein
kinase C (PKC), phosphoinositide 3-kinase (PI3K), extracellular
signal-regulated kinase (Erk)1/2 and mitochondrial KATP channels,
have been implicated in the molecular mechanisms of myocardial
protection by volatile anesthetics, including sevoflurane (3). However, the cellular and molecular
processes by which sevoflurane exerts protective effects on the
heart are incompletely understood.
MicroRNAs (miRNAs or miRs), small, non-coding RNAs
of approximately 22 nucleotides (nt) in length, play a critical
role in the post-transcriptional regulation of their target genes
at the mRNA and/or protein level (4). Dysregulated miRNA expression has
been implicated in a number of pathophysiological mechanisms, such
as oncogenesis (5) and
cardiovascular disease (6).
Moreover, miRNAs are not only localized intracellularly, but are
also secreted into extracellular fluid, such as plasma, serum,
saliva and urine, via exosomes, i.e., tiny vesicles of 50–100 nm in
diameter (7). Exosomal miRNAs are
generally considered to be stable in the circulation (8). This has raised the possibility that
circulating miRNAs may serve as novel biomarkers for detecting and
monitoring various pathophysiological conditions, such as cancer
(8).
Certain studies in the field of anesthesiology have
been conducted to explore tissue- and organ-specific alterations in
miRNA expression induced by volatile anesthetics in rats (9,10).
Tanaka et al (9) reported
significant changes in miRNA expression profiles in rat lungs under
sevoflurane anesthesia. Moreover, Ishikawa et al (10) demonstrated differential miRNA
expression patterns between sevoflurane and propofol anesthesia in
the rat liver. Based on the findings regarding circulating miRNAs,
plasma miRNAs may be informative markers for monitoring and
assessing the effects of the volatile anesthetic modification of
miRNA expression in organs and body systems. However, volatile
anesthetic-associated dynamics of circulating miRNAs and the
detailed mechanisms of the effects of volatile anesthetics on
tissues and organs remain unclear.
Thus, the aim of this study was to perform a
comprehensive analysis of the time-dependent changes in circulating
miRNA levels and composition in sevoflurane-anesthetized rats using
a quantitative polymerase chain reaction (PCR)-based array. We
discovered that the persistent downregulation of muscle
specific-miRNAs (also known as myomiRNAs) occurred for 2 weeks
following sevoflurane anesthesia. Sevoflurane anesthesia may
predominantly affect cardiac and skeletal muscles and suppress
miRNA secretion from these tissues into the circulation.
Materials and methods
Sample preparation
This study was approved by the Animal Research
Committee of Nippon Medical School, Tokyo, Japan. Six-week-old male
Wistar rats (Saitama Experimental Animals Supply, Saitama, Japan),
weighing 180±20 g, were maintained under a 12/12-h light/dark cycle
in a temperature-controlled environment. Sevoflurane anesthesia was
performed using the procedure employed in our previous miRNA
studies on rats (9,10). Briefly, each rat was allowed to
breathe spontaneously, housed in an anesthesia box (width, 690 mm;
diameter, 410 mm; height, 330 mm; MAB-2; Sanplatec Corp., Osaka,
Japan), and supplied with an air-oxygen mixture (fraction of
inspired oxygen, 0.4) at a rate of 6 l/min, with body temperature
maintained at approximately 37°C using a heat lamp. Rats undergoing
sevoflurane anesthesia were supplied in the box with 2.0% (1
‘minimum alveolar concentration’) sevoflurane (Maruishi
Pharmaceutical Co., Ltd., Osaka, Japan) for 6 h (designated as 6-h
anesthesia), as previously described (11); this dose is commonly used
clinically. The control group rats received no anesthesia. There
were no significant differences in physiological data during
anesthesia between the anesthesia groups and the control group.
Hypoxia, hyper/hypocapnia, hypotension, or hypothermia did not
occur in any of the groups. The animals were sacrificed either
immediately after cessation of the 6-h anesthesia, or after the
recovery periods that lasted 1, 3, 7 and 14 days following the
termination of exposure (referred to as days 1, 3, 7 and 14
post-anesthesia, respectively).
In the initial PCR-based array, the rats were
assigned to 4 groups (n=6 per group): i) the control group (no
anesthesia), ii) the 6-h anesthesia group, iii) the day 1
post-anesthesia group and iv) the day 7 post-anesthesia group. In
all the groups, rat blood samples were obtained from the inferior
vena cava within 3 min after sacrifice by cervical dislocation.
In a subsequent validation assay, the rats were
assigned to 7 groups (n=6 per group): i) the control group, ii) the
3-h anesthesia group, iii) the 6-h anesthesia group, iv) the day 1
post-anesthesia group, v) the day 3 post-anesthesia group, vi) the
day 7 post-anesthesia group and vii) the day 14 post-anesthesia
group. Samples [blood, heart and skeletal muscle (quadriceps
femoris)] were obtained within 5 min after sacrifice by cervical
dislocation.
RNA preparation
To separate blood plasma, blood samples were
collected into EDTA vacuum blood collection tubes (Venogect II;
Terumo, Tokyo, Japan) and then centrifuged (1,700 × g, 15 min,
4°C). The collected plasma samples were transferred to
RNase/DNase-free 1.5-ml microcentrifuge tubes, and stored at −80°C
before RNA purification. Total RNA from plasma and tissue samples
was extracted using Isogen-LS and Isogen (both from Wako, Osaka,
Japan), respectively, according to the manufacturer’s
instructions.
Comprehensive quantitative analysis of
circulating miRNAs by quantitative PCR-based array
For the initial quantitative PCR-based array, equal
quantities from each plasma sample were pooled within each group;
total RNA were extracted from the pooled plasma samples as
described above. Total RNA extracted from the equivalent of 25-μl
plasma was reverse-transcribed using Megaplex RT Primers (Applied
Biosystems, Foster City, CA, USA). The cDNA was then pre-amplified
using Megaplex PreAmp Primers (Applied Biosystems). The
pre-amplified products were subjected to quantitative PCR using
TaqMan MicroRNA Assay Rodent Panels (A and B, v.3.0) on a 7900HT
Fast Real-Time PCR System (Applied Biosystems) according to the
manufacturer’s instructions. miRNA sequences were annotated using
the Sanger database (miRBase), release 14. Data obtained with this
assay were analyzed using RQ Manager 1.2 (Applied Biosystems). For
the quantification of each miRNA expression level, the comparative
Ct method (ΔΔCt method) was used as described below. Full array
data sets are available upon request.
Quantitative analysis of miRNA expression
by quantitative PCR
Quantitative PCR of the miRNAs was performed using
TaqMan Gene Expression assays (Applied Biosystems) in a 7300
Real-Time PCR System (Applied Biosystems), according to the
manufacturer’s instructions. To normalize the miRNA expression
levels, Caenorhabditis elegans miRNAs (cel-miRNAs) and
Rnu6 were used as exogenous internal controls for the plasma
samples and an endogenous internal control for the tissue samples,
respectively.
Normalization of PCR data for circulating
miRNAs using exogenous ‘spike-in’ cel-miRNAs
We used a fixed volume of RNA eluate (2 μl) from a
given volume of starting plasma as input for the RT reaction. For a
sample in which the starting plasma volume was 125 μl, an input of
2 μl of eluted RNA (taken from a total RNA eluate volume of ~20 μl)
into the RT reaction corresponds to the mass of RNA derived from
~12.5 μl of starting plasma.
Both PCR-based array and quantitative PCR data of
the plasma miRNAs were normalized to exogenous cel-miRNAs as a
‘spike-in’ control using a modification of the method described in
the study by Mitchell et al (8). Briefly, 2 synthetic RNA
oligonucleotides corresponding to cel-miR-39 and
cel-miR-238 (Qiagen, Valencia, CA, USA) were used. The
‘spike-in’ oligos were introduced (as a mixture of 25 fmol of each
oligonucleotide in 5-μl water) after the addition of Isogen-LS to
the plasma samples. For each RNA sample, the cel-miRNAs were
measured using TaqMan qRT-PCR assays (Applied Biosystems) as
described above. The threshold cycle (Ct) values obtained for the 2
‘spike-in’ cel-miRNAs were averaged to generate a ‘spike-in’
control Ct value. This subsequently produced a different (ΔCt)
value for each plasma miRNA based on the following formula: ΔCt =
(plasma miRNA Ct value of a given sample) − (‘spike-in’ control Ct
value of the sample). A sample in the control group (no anesthesia)
was arbitrarily designated as the calibrator sample (1× sample),
and the relative miRNA levels of samples in all other groups were
then expressed relative to the calibrator sample. Thus, the value
of ΔΔCt for each testing sample was determined by the formula: ΔΔCt
= ΔCt (testing sample) − ΔCt (calibrator sample).
Statistical analysis
We conducted all analyses using SPSS software (v.20
for Windows; IBM-SPSS). The significance of between-group
differences was assessed using ANOVA followed by Dunnett’s test.
P-values <0.05 were considered to indicate statistically
significant differences. Values are expressed as the means ±
standard deviation (SD).
Results
Comprehensive profile analysis of
circulating microRNAs using quantitative PCR-based miRNA expression
array
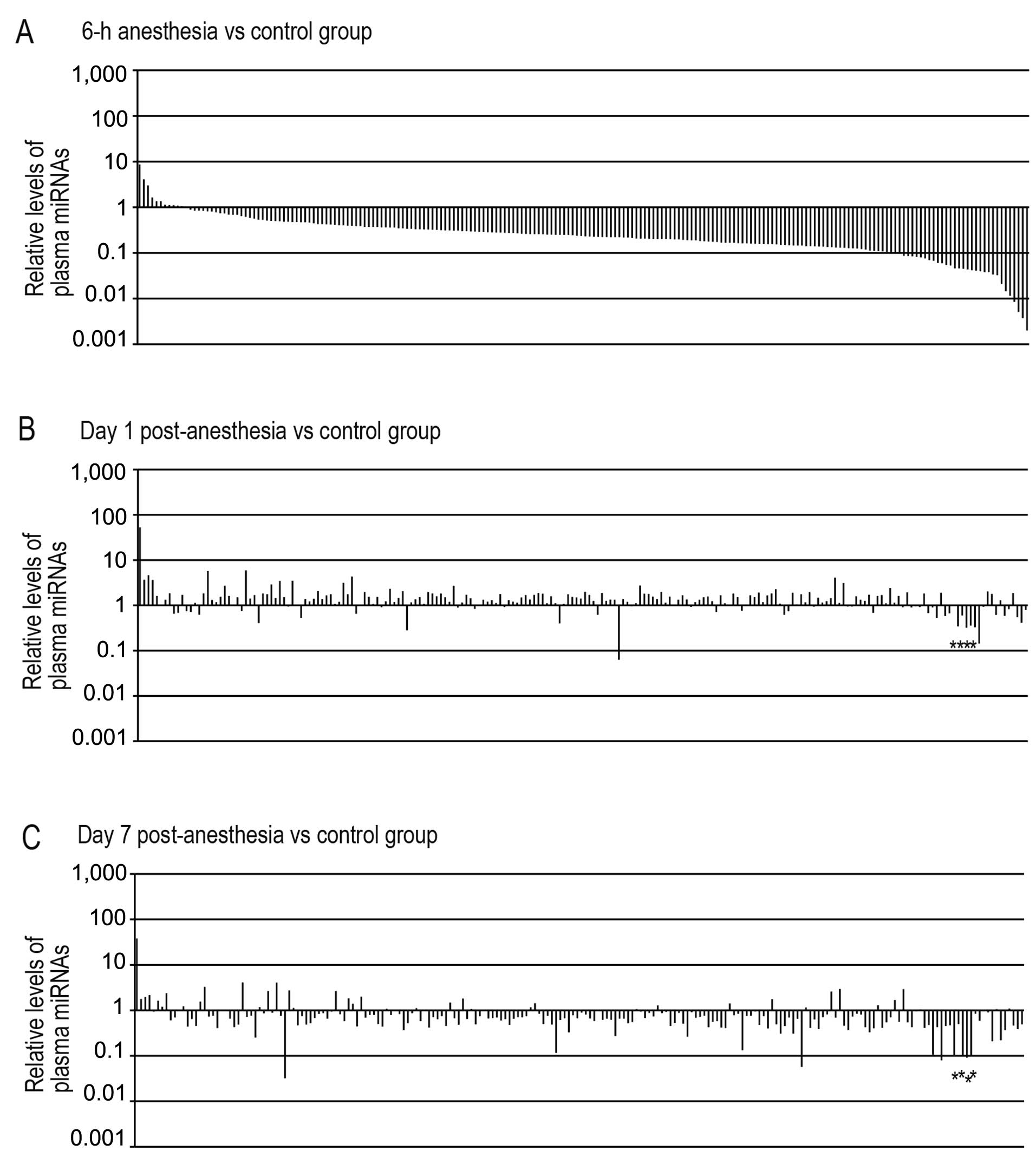
Among the 373 preloaded rat miRNAs contained in
real-time PCR-based miRNA expression profiling array cards, 210
miRNAs (56% of the preloaded miRNAs) were detected in rat plasma
from the control group (no anesthesia). Surprisingly, 161 plasma
miRNAs (77% of the miRNAs detected in the control group) were
decreased by >2.5-fold in the 6-h anesthesia group compared with
the control group (Fig. 1A). By
contrast, 46 plasma mRNA levels (22%) remained unlatered (i.e.,
between 2.5-fold underexpressed and overexpressed), and only 3
miRNAs (1%) were increased by >2.5-fold in the 6-h anesthesia
group compared with the control group. Fold differences in miRNAs
ranged from 0.002- to 8.6-fold change in the 6-h anesthesia group
relative to the control group (median, 0.234). It should be noted
that the majority of the miRNAs detected in the control group were
downregulated in the rat plasma during sevoflurane anesthesia.
Following sevoflurane anesthesia, the levels of the
plasma miRNAs which had decreased within 6 h of anesthesia,
substantially increased in the day 1 post-anesthesia group: of the
161 plasma miRNAs decreased by >2.5-fold in the 6-h anesthesia
group compared with the control group, the levels of 148 of these
miRNAs (92%) returned to control group levels (between 2.5-fold
underexpressed and overexpressed) (Fig. 1B). Fold differences in miRNAs
ranged from 0.063- to 52.6-fold changes in the day 1
post-anesthesia group relative to the control group (median,
1.331). In the day 7 post-anesthesia group, of the 210 miRNAs
detected in rat plasma from the control group, the levels of 176 of
these miRNAs (84%) were between 2.5-fold underexpressed and
overexpressed as compared with those of the control group (Fig. 1C). Fold differences in miRNAs
ranged from 0.032- to 38.0-fold change in the day 7 post-anesthesia
group relative to the control group (median, 0.695). Only 6 miRNAs
were downregulated by >2.5-fold in both the day 1 and day 7
post-anesthesia groups compared with the control group (Table I). Of note, 4 miRNAs
(miR-1, miR-133a, miR-133b and
miR-206), which have been reported to be muscle-specific
miRNAs (12), were included in
the 6 plasma miRNAs that were persistently downregulated following
sevoflurane anesthesia. In terms of persistently upregulated
miRNAs, only 1 miRNA, miR-148b-5p, was upregulated by
>2.5-fold in both the day 1 and day 7 post-anesthesia groups
compared with the normal (control) group (Table I).
 | Table IRepresentative miRNAs significantly
downregulated or upregulated in rat plasma in a quantitative
PCR-based array. |
Table I
Representative miRNAs significantly
downregulated or upregulated in rat plasma in a quantitative
PCR-based array.
| miRNA | Ct value control
group | Fold change 6-h
anesthesia | Fold change day 1
post-anesthesia | Fold change day 7
post-anesthesia |
|---|
| Downregulated
miRNAsa |
|
miR-133a | 19.86 | 0.032 | 0.311 | 0.075 |
|
miR-206 | 22.61 | 0.046 | 0.329 | 0.225 |
|
miR-133b | 22.89 | 0.033 | 0.333 | 0.072 |
| miR-1 | 23.80 | 0.030 | 0.348 | 0.066 |
|
miR-433 | 25.60 | 0.243 | 0.273 | 0.265 |
|
miR-350 | 29.20 | 0.221 | 0.063 | 0.272 |
| Upregulated
miRNAsb |
|
miR-148b-5p | 35.88 | 8.601 | 52.552 | 37.969 |
Although many miRNAs are expressed ubiquitously in
mammals, some miRNAs exhibit specific expression patterns in a
tissue- or cell type-dependent manner (13). Several miRNAs, such as
miR-1, miR-133a, miR-133b and miR-206,
are specifically expressed in striated muscle tissue. It seems
reasonable that plasma muscle-specific miRNAs mirror an altered
miRNA status in cardiac and skeletal muscle tissue. Thus, it may be
that the decreased plasma levels of muscle-specific miRNAs were
associated with the sevoflurane-induced inhibition of the release
of miRNAs from cardiac and skeletal muscle tissues. Additionally,
considering the experimental and technical aspects of plasma miRNA
application as potential biomarkers to evaluate the effects of
volatile anesthetics, the 4 muscle-specific miRNAs detected at low
Ct values in rat plasma may be advantageous in terms of sensitivity
and reliability as a low Ct value corresponds to a high miRNA level
in the plasma (Table I). Thus, we
then focused on the muscle-specific miRNAs that were persistently
downregulated following sevoflurane anesthesia.
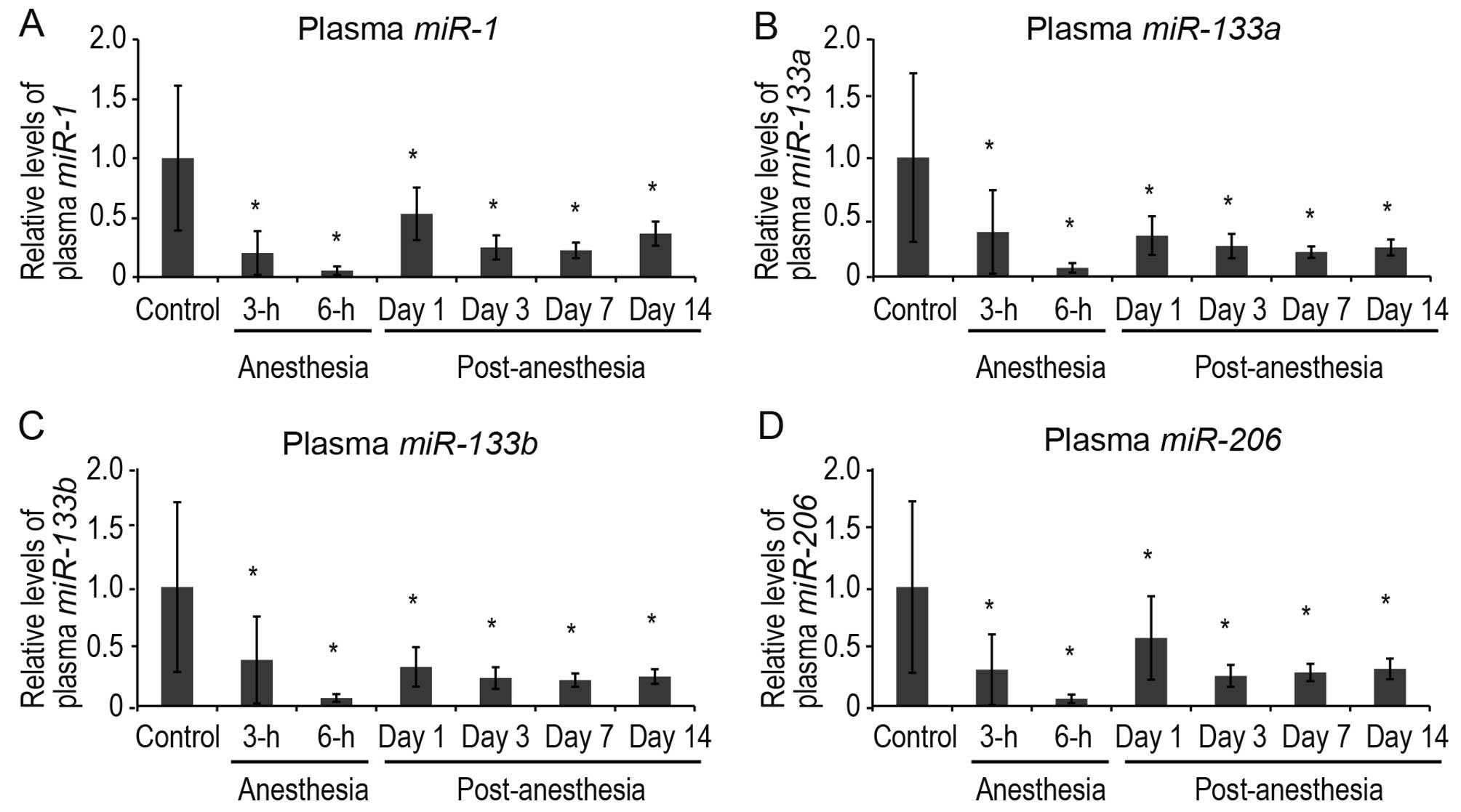
Validation of dynamics of plasma
muscle-specific miRNAs by quantitative PCR
To confirm whether plasma muscle-specific miRNAs
were indeed downregulated by sevoflurane, the plasma levels of 4
muscle-specific miRNAs (miR-1, miR-133a,
miR-133b and miR-206) were analyzed during and after
sevoflurane anesthesia by quantitative PCR (Fig. 2). Quantitative PCR was performed
at extended time points: at 3 and 6 h of anesthesia, and at 1, 3, 7
and 14 days post-anesthesia. The plasma levels of all 4
muscle-specific miRNAs decreased significantly at 3 h after the
induction of anesthesia and were downregulated until 14 days
post-sevoflurane anesthesia (Fig.
2).
Expression levels of muscle-specific
miRNAs in cardiac and skeletal muscle
It is conceivable that the suppression of the
release of miRNAs from cardiac and/or skeletal muscle tissue is the
major cause of the decrease in plasma muscle-specific miRNA levels.
To investigate this matter, we examined the expression levels of
these miRNAs in cardiac and skeletal muscle (n=6 per group)
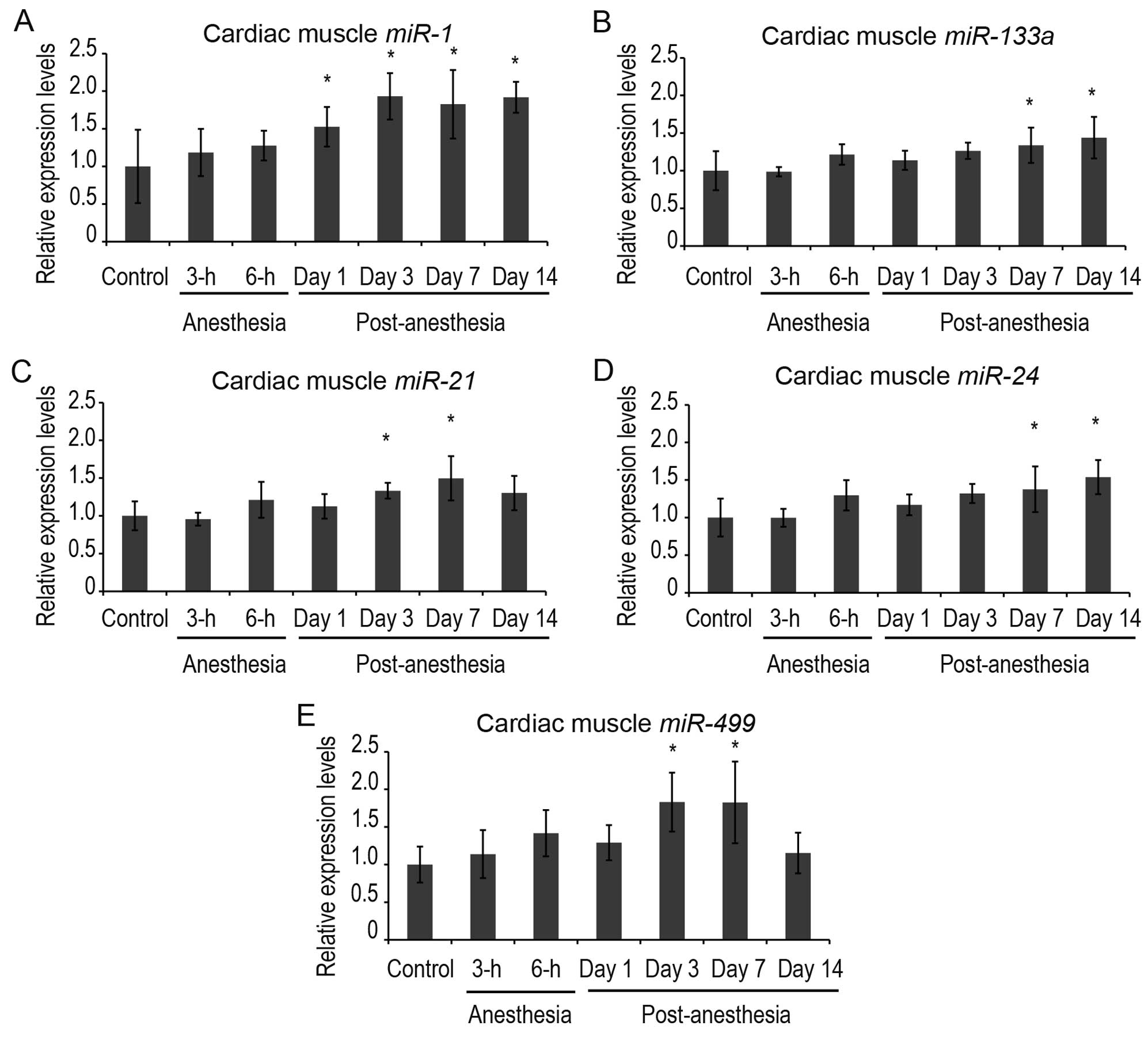
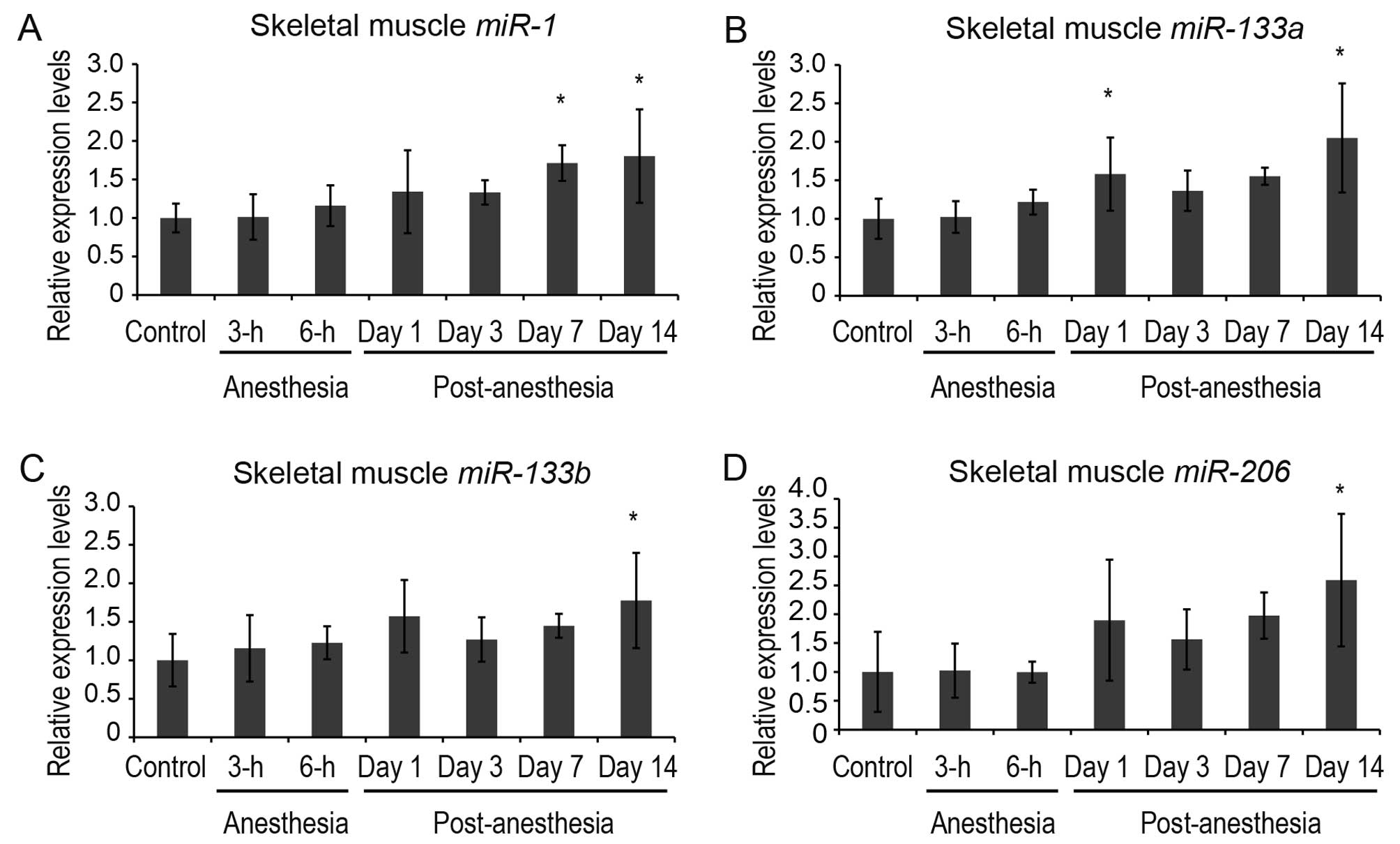
(Figs. 3 and 4). miR-1 and miR-133a were
expressed in both skeletal and cardiac muscle, while
miR-133b and miR-206 were expressed solely in
skeletal muscle, in accordance with a previous report (12).
In cardiac muscle, the expression levels of
miR-1 were increased significantly at 1 day post-anesthesia
and were upregulated until 14 days post-anesthesia (Fig. 3). The expression levels of
miR-133a were also significantly increased at 7 and 14 days
post-anesthesia. In the skeletal muscle, the expression levels of
muscle-specific miRNAs tended to increase following sevoflurane
anesthesia (Fig. 4). The
expression levels of miR-1 were increased significantly at 7
and 14 days post-anesthesia. A significant increase in
miR-133a levels was detected at 1 and 14 days
post-anesthesia. The expression levels of miR-133b
and miR-206, skeletal muscle-specific miRNAs, were increased
significantly at 14 days post-anesthesia. These findings, in
conjunction with the results presented above for the plasma
muscle-specific miRNAs, indicate that the expression levels of
muscle-specific miRNAs in cardiac and skeletal muscles and their
plasma levels are substantially inversely correlated following
sevoflurane anesthesia.
We also examined miR-21, miR-24 and
miR-499 in cardiac muscle (Fig. 3) as these miRNAs have been
reported to protect cardiomyocytes against
ischemia/reperfusion-induced apoptosis (14). A significant increase in
miR-21 and miR-499 levels was detected at 3 and 7
days post-anesthesia (Fig. 3).
The expression levels of miR-24 increased significantly at 7
and 14 days post-anesthesia.
Discussion
In the present study, we examined the dynamics of
circulating miRNAs as potentially informative markers for
monitoring and assessing changes in miRNA expression due to
sevoflurane anesthesia in organs and body systems. The main finding
was that the majority of the circulating miRNAs in rat plasma were
transiently downregulated as a result of sevoflurane anesthesia.
With the exception of muscle-specific miRNAs (miR-1,
miR-133a, miR-133b and miR-206), the levels of
the plasma miRNAs which were downregulated increased substantially
immediately after the anesthesia was terminated (in the recovery
period). Furthermore, we revealed distinct plasma profiles of the
muscle-specific miRNAs following sevoflurane anesthesia; the plasma
levels of the muscle-specific miRNAs were persistently
downregulated until 14 days post-anesthesia. Finally, we
demonstrated that the expression levels of muscle-specific miRNAs
in cardiac and skeletal muscle and their plasma levels were
substantially inversely correlated following sevoflurane
anesthesia, suggesting that sevoflurane predominantly affects
cardiac and skeletal muscle and suppresses the release of miRNAs
from these tissues into the circulation. To the best of our
knowledge, the present study is the first to demonstrate the
circulating miRNA expression signature induced by sevoflurane
anesthesia in rats.
Several miRNA genes are specifically expressed or
highly enriched in cardiac and/or skeletal muscle, namely the
muscle-specific miRNAs (12,15). Muscle-specific miRNAs regulate the
differentiation and proliferation of muscle cells (16). Recently, a number of studies have
reported that miRNAs in plasma or serum are promising biomarkers
for muscle diseases and myocardial injury (17–19). Mizuho et al (17) demonstrated that serum levels of
miR-1, miR-133a and miR-206 were increased in
both the dystrophin-deficient muscular dystrophy mouse model, mdx,
and the canine X-linked muscular dystrophy in Japan (CXMDJ) dog
model. Cheng et al (18)
reported that in a rat model of acute myocardial infarction (AMI)
induced by coronary ligation, serum levels of miR-1 were
increased significantly following AMI and that serum miR-1
levels with AMI showed a positive correlation with myocardial
infarct size. The increase in circulating muscle-specific miRNAs in
these muscle dystrophies and AMI models appears to be caused by an
increase in the leakage of miRNAs from muscle tissues.
In the present study, we found that sevoflurane
anesthesia significantly altered the plasma levels of
muscle-specific miRNAs compared with the levels of other miRNAs.
However, the mechanisms involved in this sevoflurane-induced
downregulation of circulating muscle-specific miRNAs need to be
elucidated. There are several possible explanations for these
effects of sevoflurane. First, it is possible that the
exosome-mediated secretion of miRNAs is attenuated in cardiac and
skeletal muscle, resulting in the relatively lower plasma levels of
muscle-specific miRNAs compared with those of other miRNAs. Another
explanation may be that de novo miRNA synthesis is blocked
in cardiac and skeletal muscle. To confirm whether the expression
levels of muscle-specific miRNAs were altered by sevoflurane
anesthesia in cardiac and skeletal muscle, we performed
quantitative PCR analysis and found that the expression levels of
muscle-specific miRNAs showed a negative correlation with the
plasma levels (Figs. 3 and
4). Taking these findings into
consideration, it seems likely that sevoflurane anesthesia affects
miRNA secretion rather than de novo miRNA synthesis in
cardiac and skeletal muscle. Circulating miRNAs are generally
considered to be the result of exosomes that can protect
circulating miRNAs from plasma RNase (8). However, a recent study raised an
objection to this. Specifically, Roberts et al (20) demonstrated that the majority of
serum muscle-specific miRNAs were protected from serum nucleases by
the association with protein/lipoprotein complexes, rather than
extracellular vesicles, in mdx mice. Thus, further studies are
required in order to elucidate the molecular mechanisms of the
sevoflurane-dependent muscle cell type-specific inhibition of miRNA
secretion.
Sevoflurane exerts cardioprotective effects in
anesthetic preconditioning (APC) when administered before a period
of myocardial ischemia and reperfusion (21,22). However, there have been few
studies on the role of cardioprotective miRNAs in APC. Several
myocardial ischemia-associated miRNAs have been reported to serve
as cardioprotective miRNAs (14).
For example, miR-21 has been shown to be upregulated in the
mouse heart following ischemic preconditioning (IPC) and sublethal
heat shock, and exerts a protective effect on
hypoxia/reoxygenation-induced cell apoptosis, targeting programmed
cell death 4 (23). IPC-induced
miRNAs (miR-1, miR-21 and miR-24) exert
cardioprotective effects similar to the delayed phase of IPC,
possibly through upregulating endothelial nitric oxide synthase
(eNOS), heat shock protein (HSP)70 and the HSP70 transcription
factor, HSF-1 (24).
miR-24 has been shown to suppress cardiomyocyte apoptosis,
by repressing the gene encoding the BH3-only domain-containing
protein ‘Bim’, which positively regulates apoptosis (25). miR-499 inhibits
cardiomyocyte apoptosis through its suppression of the
calcineurin-mediated dephosphorylation of dynamin-related protein-1
(26). In the present study, we
investigated the effects of sevoflurane anesthesia on the
expression levels of some cardioprotective miRNAs (miR-21,
miR-24 and miR-499) in cardiac muscle and found that
a significant increase in their expression levels was also detected
within 2 weeks post-anesthesia (Fig.
3). Sevoflurane may have cardioprotective effects by
persistently upregulating endogenous miRNAs in cardiac muscle.
Unlike miR-133b and miR-206,
miR-1 and miR-133a, examined in this study, could be
derived from cardiac and/or skeletal muscle (12,15). miR-208 and miR-499
are primarily expressed in cardiac muscle (26); however, we did not determine
significant changes in the levels of these miRNAs in the plasma. At
present, the estimation of the proportion of circulating
muscle-specific miRNAs derived from individual muscles remains an
issue.
In conclusion, we demonstrted a circulating miRNA
expression signature induced by sevoflurane anesthesia in rats. We
revealed the persistent downregulation of muscle-specific miRNAs
(miR-1, miR-133a, miR-133b and miR-206)
in rat plasma for 2 weeks following sevoflurane anesthesia. The
expression levels of muscle-specific miRNAs in cardiac and skeletal
muscle showed a negative correlation with their plasma levels. Our
data suggest that sevoflurane anesthesia suppresses the release of
miRNAs from cardiac and skeletal muscle tissue into the
circulation, and this may contribute to the cardioprotective
effects of sevoflurane. Although further functional and
pathological studies on muscle-specific modification induced by
sevoflurane are required, this new information provides novel
insight towards a better understanding of the molecular mechanisms
of action of the anesthetic, sevoflurane.
Acknowledgements
The authors are indebted to Mr. Takuji Kosuge
(Department of Molecular Medicine and Anatomy, Nippon Medical
School, Tokyo, Japan) for providing technical assistance. The
present study was supported by Grants-in-Aid for Scientific
Research from the Ministry of Education, Culture, Sports, Science
and Technology (MEXT), Japan, and by a grant from the
MEXT-Supported Program for the Strategic Research Foundation at
Private Universities (2013–2017).
References
|
1
|
Brown BR Jr and Frink EJ: The safety of
sevoflurane in humans. Anesthesiology. 79:201–202. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Obal D, Preckel B, Scharbatke H,
Müllenheim J, Höterkes F, Thämer V and Schlack W: One MAC of
sevoflurane provides protection against reperfusion injury in the
rat heart in vivo. Br J Anaesth. 87:905–911. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu ZY and Liu J: Mechanism of cardiac
preconditioning with volatile anaesthetics. Anaesth Intensive Care.
37:532–538. 2009.PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Le Quesne J and Caldas C: Micro-RNAs and
breast cancer. Mol Oncol. 4:230–241. 2010.PubMed/NCBI
|
|
6
|
Small EM, Frost RJ and Olson EN: MicroRNAs
add a new dimension to cardiovascular disease. Circulation.
121:1022–1032. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weber JA, Baxter DH, Zhang S, Huang DY,
Huang KH, Lee MJ, Galas DJ and Wang K: The microRNA spectrum in 12
body fluids. Clin Chem. 56:1733–1741. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mitchell PS, Parkin RK, Kroh EM, Fritz BR,
Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O’Briant
KC, Allen A, Lin DW, Urban N, Drescher CW, Knudsen BS, Stirewalt
DL, Gentleman R, Vessella RL, Nelson PS, Martin DB and Tewari M:
Circulating microRNAs as stable blood-based markers for cancer
detection. Proc Natl Acad Sci USA. 105:10513–10518. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tanaka S, Ishikawa M, Arai M, Genda Y and
Sakamoto A: Changes in microRNA expression in rat lungs caused by
sevoflurane anesthesia: a TaqMan® low-density array
study. Biomed Res. 33:255–263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ishikawa M, Tanaka S, Arai M, Genda Y and
Sakamoto A: Differences in microRNA changes of healthy rat liver
between sevoflurane and propofol anesthesia. Anesthesiology.
117:1245–1252. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Orliaquet G, Vivien B, Langeron O,
Bouhemad B, Coriat P and Riou B: Minimum alveolar concentration of
volatile anesthetics in rats during postnatal maturation.
Anesthesiology. 95:734–739. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu N and Olson EN: MicroRNA regulatory
networks in cardiovascular development. Dev Cell. 18:510–525. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Landgraf P, Rusu M, Sheridan R, Sewer A,
Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M,
Lin C, Socci ND, Hermida L, Fulci V, Chiaretti S, Foà R, Schliwka
J, Fuchs U, Novosel A, Müller RU, Schermer B, Bissels U, Inman J,
Phan Q, Chien M, Weir DB, Choksi R, De Vita G, Frezzetti D,
Trompeter HI, Hornung V, Teng G, Hartmann G, Palkovits M, Di Lauro
R, Wernet P, Macino G, Rogler CE, Nagle JW, Ju J, Papavasiliou FN,
Benzing T, Lichter P, Tam W, Brownstein MJ, Bosio A, Borkhardt A,
Russo JJ, Sander C, Zavolan M and Tuschl T: A mammalian microRNA
expression atlas based on small RNA library sequencing. Cell.
129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu H and Fan GC: Role of microRNAs in the
reperfused myocardium towards post-infarct remodelling. Cardiovasc
Res. 94:284–292. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Koutsoulidou A, Mastroyiannopoulos NP,
Furling D, Uney JB and Phylactou LA: Expression of miR-1, miR-133a,
miR-133b and miR-206 increases during development of human skeletal
muscle. BMC Dev Biol. 11:342011. View Article : Google Scholar
|
|
16
|
Townley-Tilson WH, Callis TE and Wang D:
MicroRNAs 1, 133, and 206: critical factors of skeletal and cardiac
muscle development, function, and disease. Int J Biochem Cell Biol.
42:1252–1255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mizuho H, Nakamura A, Aoki Y, Ito N, Kishi
S, Yamamoto K, Sekiguchi M, Takeda S and Hashido K: Identification
of muscle-specific microRNAs in serum of muscular dystrophy animal
models: promising novel blood-based markers for muscular dystrophy.
PloS One. 6:e183882011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng Y, Tan N, Yang J, Liu X, Cao X, He
P, Dong X, Qin S and Zhang C: A translational study of circulating
cell-free microRNA-1 acute myocardial infarction. Clin Sci (Lond).
119:87–95. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tijsen AJ, Pinto YM and Creemers EE:
Circulating microRNAs as diagnostic biomarkers for cardiovascular
diseases. Am J Physiol Heart Circ Physiol. 303:H1085–H1095. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roberts TC, Godfrey C, McClorey G, Vader
P, Briggs D, Gardiner C, Aoki Y, Sargent I, Morgan JE and Wood MJ:
Extracellular microRNAs are dynamic non-vesicular biomarkers of
muscle turnover. Nucleic Acids Res. 41:9500–9513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Toller WG, Kersten JR, Pagel PS, Hettrick
DA and Warltier DC: Sevoflurane reduces myocardial infarct size and
decreases the time threshold for ischemic preconditioning in dogs.
Anesthesiology. 91:1437–1446. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jakobsen CJ, Berg H, Hindsholm KB, Faddy N
and Sloth E: The influence of propofol versus sevoflurane
anesthesia on outcome in 10,535 cardiac surgical procedures. J
Cardiothorac Vasc Anesth. 21:664–671. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng Y, Zhu P, Yang J, Liu X, Dong S,
Wang X, Chun B, Zhuang J and Zhang C: Ischaemic
preconditioning-regulated miR-21 protects heart against
ischaemia/reperfusion injury via anti-apoptosis through its target
PTCD4. Cardiovasc Res. 87:431–439. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yin C, Salloum FN and Kukreja RC: A novel
role of microRNA in late preconditioning: upregulation of
endothelial nitric oxide synthase and heat shock protein 70. Circ
Res. 104:572–575. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qian L, Van Laake LW, Huang Y, Liu S,
Wendland MF and Srivastava D: miR-24 inhibits apoptosis and
represses Bim in mouse cardiomyocytes. J Exp Med. 208:549–560.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang JX, Jiao JQ, Li Q, Long B, Wang K,
Liu JP, Li YR and Li PF: miR-499 regulates mitochondrial dynamics
by targeting calcineurin and dynamin-related protein-1. Nat Med.
17:71–78. 2011. View
Article : Google Scholar : PubMed/NCBI
|