Introduction
The full-length human WW domain-containing
oxidoreductase (WWOX) gene which encodes the Wwox protein located
at 16q23.3-24.1, a chromosome region that spans one of the most
active common fragile sites, termed FRA16D (1). To date, extensive research into the
WWOX gene has suggested that WWOX may function as a tumor
suppressor gene, as it behaves aberrantly and affects multiple
types of cancer, such as breast, lung, bladder, ovarian, liver,
esophageal and pancreatic cancer (2–7).
Notably, a restoration or upregulation of WWOX expression is
capable of promoting apoptosis in vitro and inhibits tumor
growth in vivo (8,9). In addition, studies using WWOX
knockout mice have demonstrated increased tumor susceptibility, as
well as defects in bone and calcium metabolism (10,11). Moreover, a reduced WWOX expression
is commonly observed in cancer cases and is associated with a poor
prognosis (12,13). Therefore, WWOX is expected to be a
potential target for the gene-targeted therapy of human
carcinomas.
The aberration or absence of WWOX expression in
primary hematopoietic malignancies has also been reported,
suggesting that WWOX also plays an important role in human
hematopoietic malignancies (14–16). Recent studies have revealed that
WWOX exerts a role as an anti-oncogene in leukemia, which inhibits
cell proliferation and promotes apoptosis through the mitochondrial
pathway (17,18). Furthermore, it has been confirmed
that WWOX is a pro-apoptotic protein that is closely related to
apoptosis-associated factors, including Bcl-2, Bcl-xL, Bax,
caspases, p73 and p53 involved in apoptotic signaling pathways
(19–21). As there is no report on the
functional role of WWOX in human multiple myeloma, and to confirm
the functional role of WWOX in suppressing multiple myeloma cells,
in this study, we transfected WWOX cDNA into U266 multiple myeloma
cells (Wwox protein-negatvie) utilizing the constructed
pCMV-WWOX-EGFP vector. We investigated the effects of Wwox protein
re-expression on the biological properties of U266 multiple myeloma
cells, as well as the underlying molecular mechanisms. Our data
revealed that Wwox re-expression resulted in a significant
suppression of cell viability and the induction of apoptosis in
U266 human multiple myeloma cells, possibly by activating the
intrinsic apoptotic pathway.
Materials and methods
Materials
The U266 multiple myeloma cell line was purchased
from the Chinese Academy of Sciences (Shanghai, China). The main
reagents used in this study are listed as follows: RPMI-1640 medium
and fetal bovine serum (FBS) (Gibco-BRL, Carlsbad, CA, USA); RT-PCR
kit (Fermentas, Glen Burnie, MD, USA); TRIzol reagent and pfu DNA
polymerase (Thermo Fisher Scientific, Waltham, MA, USA); pCMV-EGFP
vector, HindIII, Xhol enzyme, T4 DNA
ligase, RIPA lysis buffer, DAPI, Cy3-conjugated anti-rabbit IgG,
Cy3-conjugated anti-mouse IgG, mouse anti-human β-actin,
anti-cytochrome c, rabbit anti-cleaved PARP and anti-cleaved
caspase-3 (Asp175) (Beyotime Institute of Biotechnology, Shanghai,
China); rabbit anti-human Wwox, anti-Bcl-2 and anti-Bax (Abcam,
Cambridge, MA, USA); rabbit anti-caspase-9 and anti-caspase-3
(Santa Cruz Biotechnology, Santa Cruz, CA, USA); GenFectin™
Transfection kit, Plasmid Extraction Mini kit and DNA purification
kit (Beijing Zoman Biotechnology, Beijing, China); Cell Counting
Kit-8 (Dojindo Laboratories, Kumamoto, Japan).
Cell culture
U266 human myeloma cells were maintained in
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS),
and cultured at 37°C in 5% CO2.
Vector construction
Total RNA was extracted using TRIzol reagent from
normal human peripheral blood mononuclear cells and was reverse
transcribed into cDNA using a commercial RT-PCR kit. The
full-length WWOX cDNA was cloned with the primer pairs (contains
HindIII and Xhol enzyme cutting sites which are shown
by the underlined letters): forward, 5′-CCGAAGCTTGCACCATGGCAGCGCTGCGC-3′
and reverse, 5′-CCGCTCGAGTTAGCCGGACTGGC-3′. The
PCR products (1268 bp) and pCMV-EGFP vector were all digested by
both HindIII and Xhol enzymes followed by
purification using a commercial DNA purification kit. The digested
WWOX cDNA and pCMV-EGFP were integrated with the aid of the
T4 DNA ligase, followed by transformation, clone picking
and amplification. The combined plasmids were extracted using a
Plasmid Extraction Mini kit and sent for sequencing.
Cell transfection and cell growth
assays
In this section, pCMV-WWOX-EGFP (encoding Wwox-GFP
fusion protein), pCMV-EGFP (a mock plasmid only encoding GFP) and
non-treated groups (blank control) were established. The cells were
seeded at a density of 1×105/ml, and transfected using
the GenFectin™ Transfection kit according to the specified
protocol. A commercial Cell Counting Kit-8 was used to evaluate the
growth inhibitory effects following the manufacturer’s
instructions. The optical density (OD value proportional to the
cell number) was measured using a microculture plate reader (BioTek
Instruments, Winooski, VT, USA) at both 450 and 630 nm.
Colony-forming assay
Briefly, GFP-expressing cells were sorted by a
special sterile flow cytometry using Vi-Cell counter (Beckman
Coulter, Inc., Fullerton, CA, USA), and 300 cells/ml were seeded in
24-well plates. Methylcellulose dissolved in RPMI-1640 containing
30% FBS was supplied to each well at a concentration of 0.8 g/l.
The colonies containing >50 cells were counted following
incubation at 37°C in 5% CO2 for 2 weeks.
Analysis of apoptosis
DAPI fluorescence staining and DNA fragmentation
were employed in the assessment of cell apoptosis following
transfection. For the DAPI fluorescence staining, the cells were
harvested, washed with PBS and stained with DAPI (2 mg/ml) for 3–5
min followed by washing 2 times with PBS. The morphological changes
of the stained cells were observed using a fluorescence microscope
(Olympus, Tokyo, Japan) at ×400 magnification. Non-viable apoptotic
ratio (%) = (cell numbers in phase IIb/200 cells) ×100%. DNA
fragmentation analysis was performed according to the protocols of
a commercial DNA Fragmentation kit (Beyotime Institute of
Biotechnology), followed by 1.0% agarose gel electrophoresis with a
constant voltage of 18 V for >4 h.
Immunofluorescence
Beforehand, the GFP-expressing cells were sorted by
flow cytometry using a specified sterile Vi-Cell counter (Beckman
Coulter). The cell monolayer was fixed with 4% paraformaldehyde,
and the cells were incubated at 4°C overnight with rabbit
anti-cleaved caspase-3 (Asp175) (1:300) and mouse monoclonal
anti-cytochrome c (1:300) antibodies. Cy3-conjugated
anti-rabbit and anti-mouse IgG were all diluted at 1:1,000. DAPI
was used to dye the cell nuclei. The stained cells were washed with
PBS and observed with a fluorescence microscope (Olympus) at ×400
magnification.
Real-time PCR
Real-time PCR was performed using a SYBR-Green PCR
Master Mix (Roche Diagnostics GmbH, Mannheim, Germany) under the
recommended conditions. The primer sequences for Bcl-2, Bax and
GAPDH were previously described (17). The comparative Ct method to GAPDH
was employed to calculate the mRNA expression level of Bcl-2 and
Bax.
Immunoblotting
Cell lysates were prepared using RIPA protein lysis
buffer and the protein extracts were quantified and then subjected
to electrophoresis on a 10–12% SDS-PAGE gel. The proteins were
transferred onto polyvinylidene difluoride (PVDF) membranes and
blocked in Tris-buffered saline (TBS) containing 5% non-fat milk
powder. The primary antibodies and the dilutions used were: rabbit
anti-Wwox (1:1,000), anti-Bcl-2 (1:1,000), anti-caspase-9 (1:500),
anti-caspase-3 (1:500) and anti-cleaved PARP (1:300). Mouse
monoclonal anti-cytochrome c antibody was diluted at 1:500,
while mouse monoclonal anti-β-actin antibody was diluted at 1:1,000
and used as a positive control.
Statistical analysis
Data are presented as the means ± standard deviation
(SD). One-way ANOVA, the Student’s t-test or the non-parametric
test were used for comparison differences between groups using SPSS
13.0 software (SPSS Inc., Chicago, IL, USA), and statistical
significance for the data was set at P-value <0.05.
Results
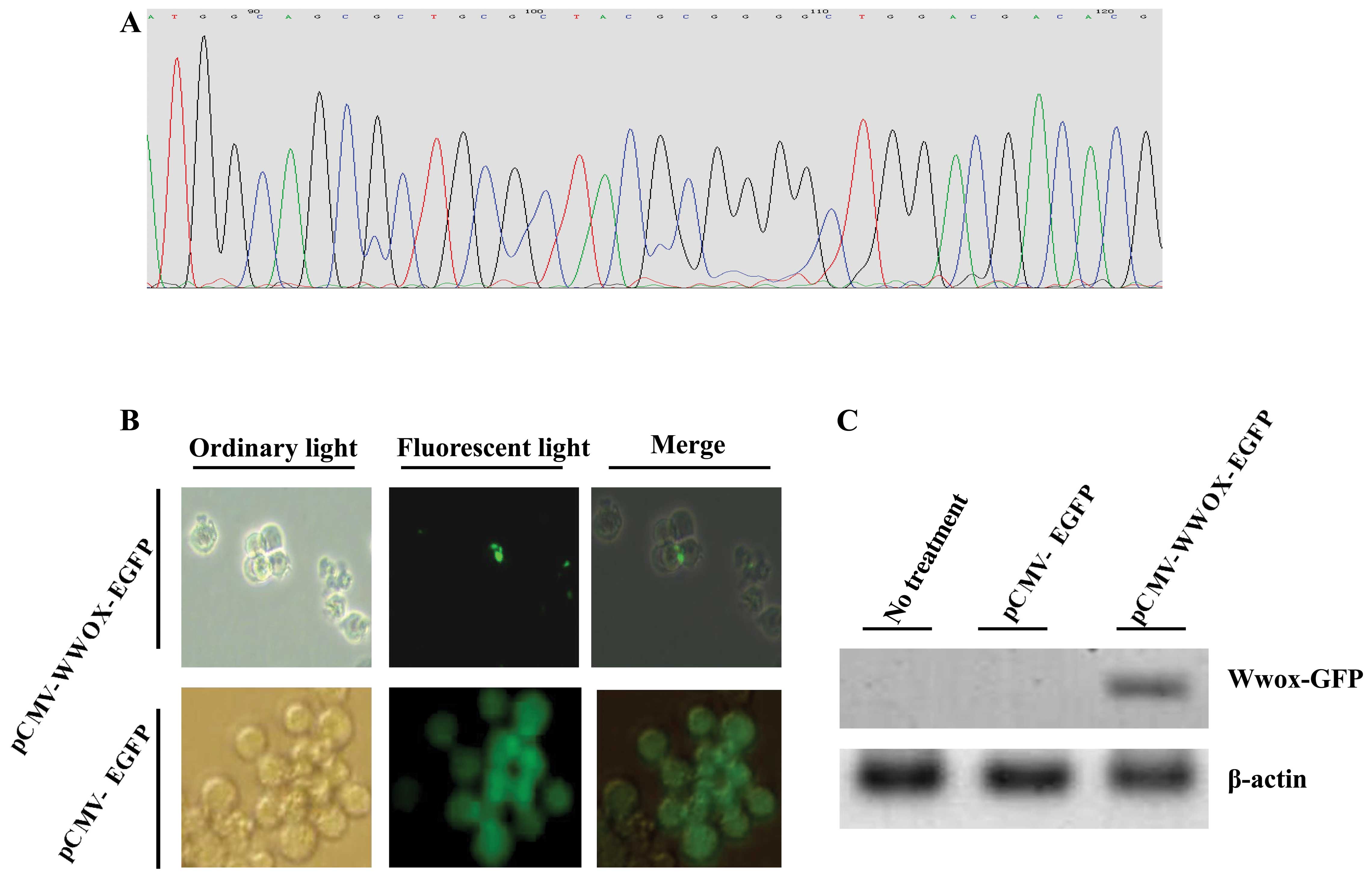
Exogenous WWOX cDNA was successfully
cloned and effectively transfected into the U266 cells
We first examined whether the pCMV-WWOX-EGFP vector
was correctly recombinated: the DNA sequencing results exhibited
that the full length WWOX cDNA was correctly connected to the
plasmid (Fig. 1A). We further
evaluated whether Wwox protein was successfully restored or
re-expressed in the U266 cells using a fluorescence microscope and
immunoblotting. The results indicated that the pCMV-WWOX-EGFP- and
pCMV-EGFP-transfected cells all expressed GFP at 36 h following
transfection (Fig. 1B). In
addition, the re-expression of Wwox protein was further confirmed
by immunoblotting using rabbit anti-human Wwox antibody: an
expected 74-kDa Wwox-GFP fusion protein at 72 h following
transfection was detected (Fig.
1C).
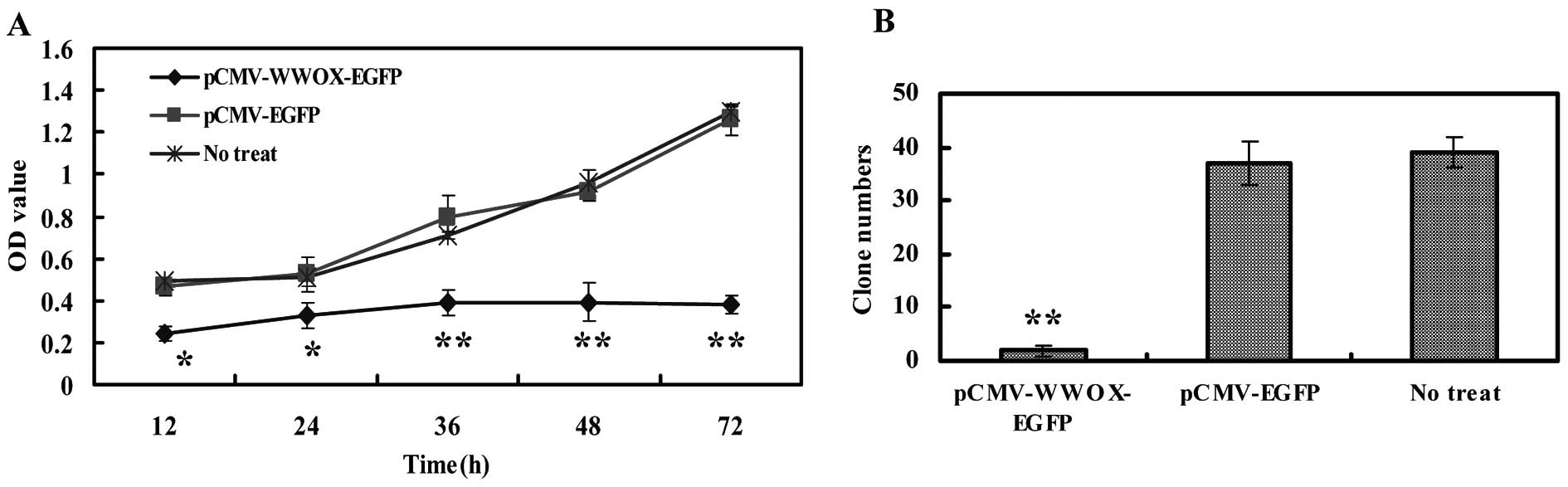
Re-expression Wwox reduces cell viability
and colony formation
The effects of Wwox protein re-expression or
restoration on the viability of U266 cells were assessed by CCK-8
assay. The results revealed that pCMV-WWOX-EGFP transfection led to
a detection of 0.242±0.037, 0.326±0.059, 0.386±0.061, 0.393±0.089
and 0.379±0.042 of the OD values (proportional to the cell numbers)
at 12, 24, 36, 48 and 72 h following transfection, respectively;
significantly lower than those of non-treated cells with P<0.05
or 0.01. By contrast, pCMV-EGFP mock vector transfection revealed
no difference with P>0.05 when compared with the non-treated
U266 cells (Fig. 2A). For the
colony formation assay, significantly fewer and smaller colonies
were formed for the pCMV-WWOX-EGFP-transfected cells compared with
the untreated cells, while the pCMV-EGFP-transfected cells showed
no statistical significance vs. the untreated cells with P>0.05
(Fig. 2B).
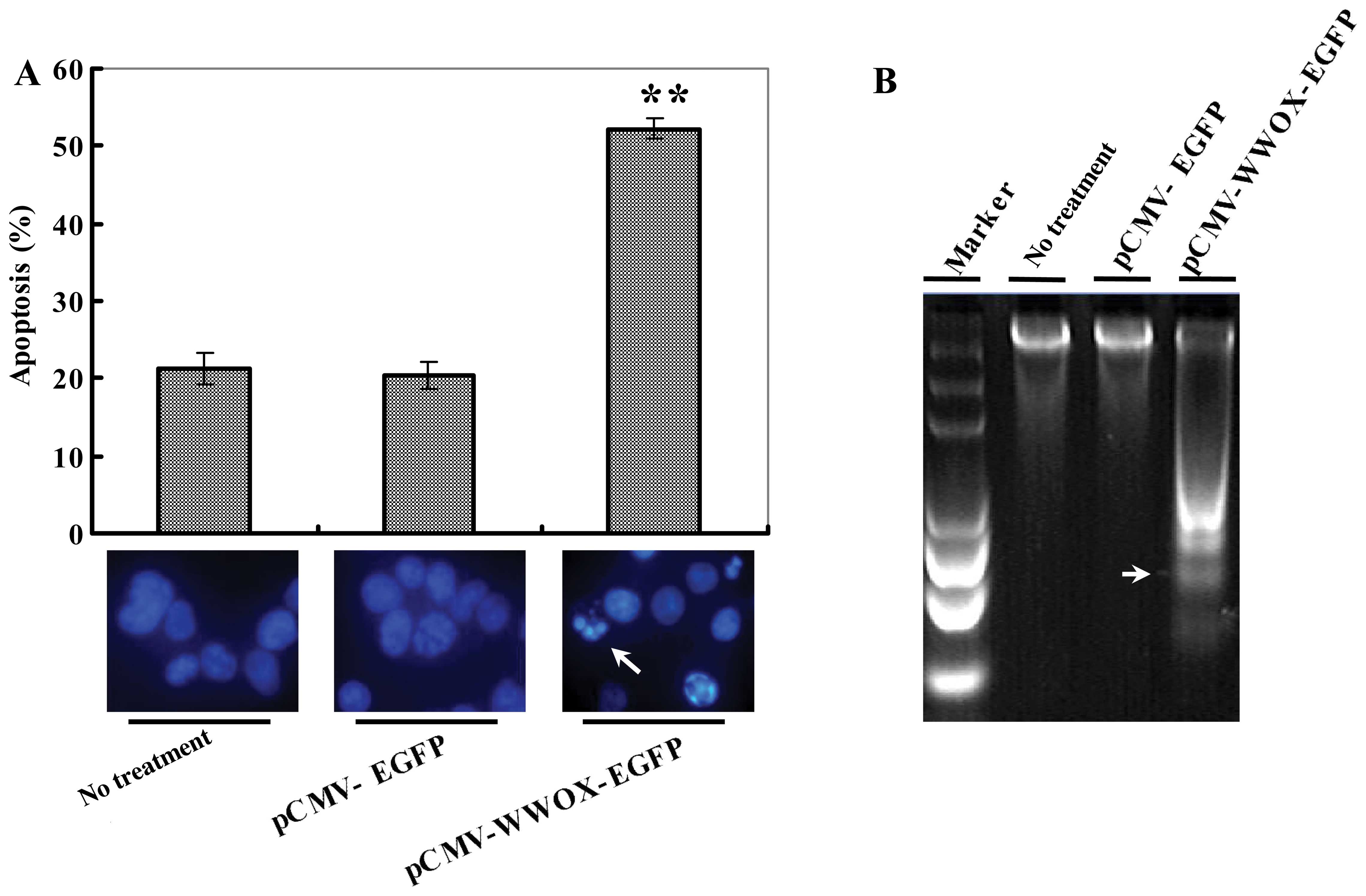
pCMV-WWOX-EGFP transfection promotes cell
apoptosis
The apoptosis-inducing effects were measured with
the aid of DAPI staining and DNA ladder electrophoresis. As
displayed in Fig. 3A, DAPI
staining exhibited apparent microscopic changes in the nucleis in
the pCMV-WWOX-EGFP-transfected U266 cells characterized by a
morphology of chromatin condensation and shrinkage in phase IIb of
the apoptotic phase at 72 h following transfection, while the 2
contrast groups presented no marked apoptotic morphology.
Similarly, the non-viable apoptotic ratio (%) for pCMV-WWOX-EGFP at
72 h after transfection was (52.25±1.38%), significantly higher
than that of the 2 control groups with P<0.01 (Fig. 3A). DNA degradative fragments were
detected by DNA ladder electrophoresis, which also presented
typical apoptosis ‘DNA ladders’ in the pCMV-WWOX-EGFP-transfected
cells when compared with the controls as shown in Fig. 3B.
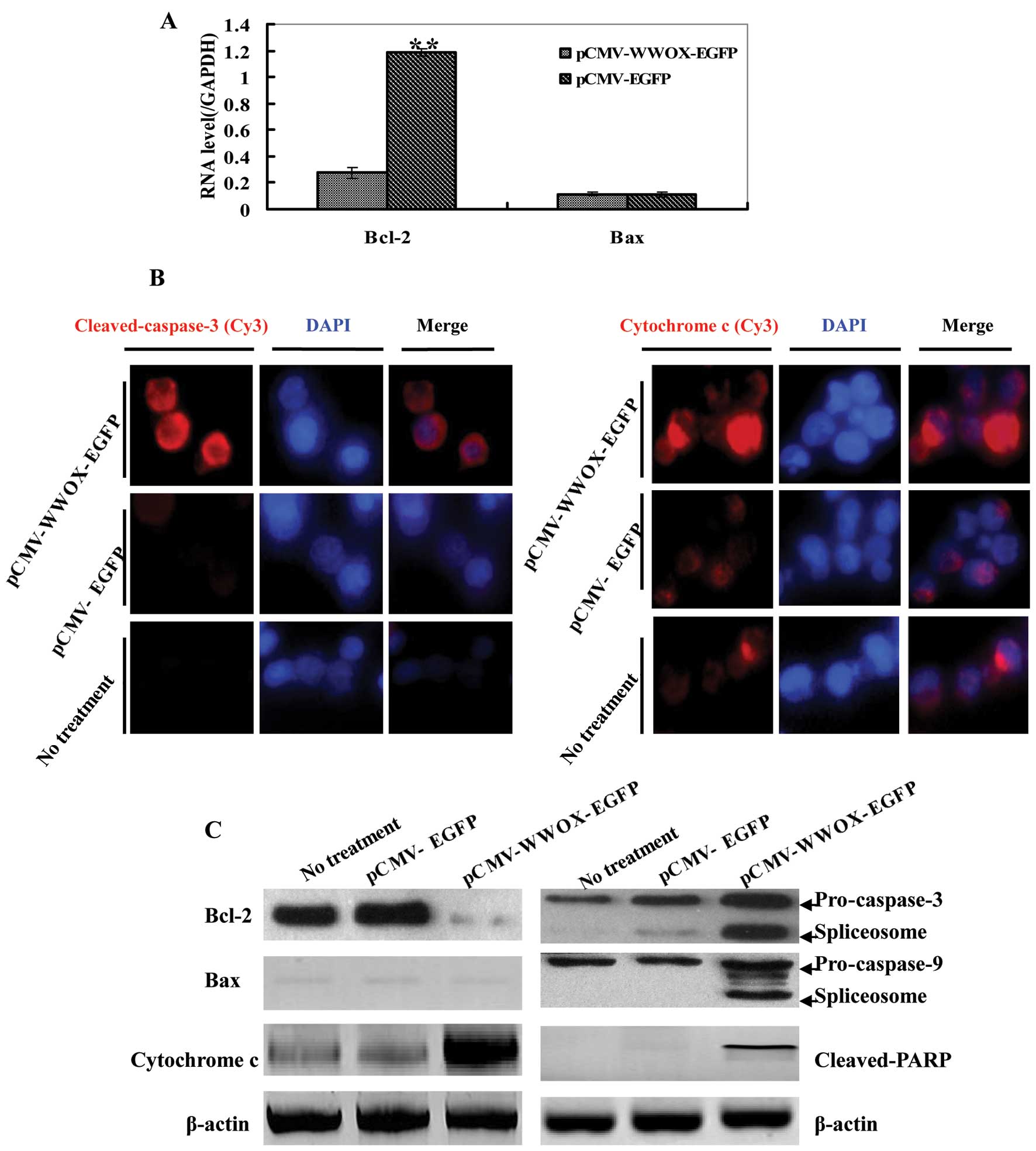
The intrinsic apoptotic pathway is
involved in the WWOX-mediated induction of apoptosis
Real-time PCR, immunoblotting and immunofluorescence
were performed for the detection of apoptosis-related proteins,
such as Bcl-2, Bax, PARP, cytochrome c, caspase-3 and
caspase-9. Real-time PCR showed that the mRNA expressio of Bcl-2 in
the pCMV-WWOX-EGFP-transfected cells decreased with P<0.01 vs.
the pCMV-EGFP-transfected cells, while the Bax mRNA expression
changed unconspicuously (Fig.
4A). As shown in Fig. 4B,
immunofluorescence revealed that caspase-3 was activated as well,
accompanied by an increase in the levels of cytochrome c.
Enhanced red fluorescence was observed in the
pCMV-WWOX-EGFP-transfected cells, utilizing anti-cleaved caspase-3
(Asp175) and anti-cytochrome c antibodies, recognized by
Cy3-conjugated anti-mouse or rabbit IgG. Immunoblotting also
displayed that the expression of Bcl-2 protein in the
pCMV-WWOX-EGFP-transfected U266 cells decreased, while cytochrome
c expression increased, all with P<0.05 when comparing
the pCMV-EGFP-transfected cells with the untreated U266 cells. Both
caspase-3 and -9 were activated, as indicated by presenting their
spliceosomes: a 17 kDa spliceosome for pro-caspase-3 and a 37 kDa
spliceosome for pro-caspase-9, both observed in the
pCMV-WWOX-EGFP-transfected U266 cells. Moreover, cleaved PARP was
also detected in the pCMV-WWOX-EGFP-transfected U266 cells
(Fig. 4B).
Discussion
In the present study, we tranfected exogenous WWOX
cDNA into U266 human multiple myeloma cells (Wwox protein-negative)
using the pCMV-WWOX-EGFP combination vector and investigated the
effects of Wwox re-expression on the biological properties of U266
multiple myeloma cells. Our data exhibited that Wwox re-expression
or restoration resulted in a significant suppression of cell
viability and the induction of apoptosis in the U266 multiple
myeloma cells. We further investigated whether Wwox re-expression
activates apoptosis-related factors involved in the intrinsic
apoptotic signaling pathway. As expected, we found that
re-expressed Wwox protein downregulated Bcl-2, upregulated
cytochrome c and activated PARP, caspase-3 and caspase-9,
indicating that Wwox plays a role as a tumor suppressor in U266
cells by activating the intrinsic apoptotic pathway.
Multiple myeloma is considered the second most
common hematological malignancy and is characterized by the clonal
proliferation of neoplastic plasma cells in the bone marrow. The
improved understanding of the molecular mechanisms of multiple
myeloma development may provide a basis for the development of
effective treatment strategies. The WWOX gene is located at the
chromosomal area 16q23.3-24.1, which spans as the common
chromosomal fragile site, FRA16D (1). The abnormal expression of WWOX,
including high frequency of loss of heterozygosity (LOH), aberrant
WWOX mRNA transcripts and reduced expression have been verified in
a variety of solid carcinomas (2–5,22),
while the restoration or overexpression of WWOX in such cancer
cells can sensitize them to apoptosis, indicating that the WWOX
gene plays a role in carcinogenesis and possesses tumor suppressor
characteristics (6,8–10,17).
The aberration or absence of WWOX expression in
primary hematopoietical malignancies has also been reported
(14–18). Our findings are consistent with
those findings: we observed that the re-expression of Wwox protein
in the Wwox-negative U266 myeloma cells led to a marked inhibition
of cell growth and colony formation, and the apoptotic effects were
exhibited by the microscopic changes in the nucleus in phase IIb of
apoptosis at 72 h after transfection, accompanied by a higher
non-viable apoptotic ratio. Furthermore, typical apoptotic DNA
ladders were observed in the pCMV-WWOX-EGFP-transfected U266 cells,
suggesting that WWOX promoted the apoptosis of U266 multiple
myeloma cells in vitro. Nevertheless, there are opposing
viewpoints as to the function role of WWOX as a tumor suppressor
gene (23); in contrary with
these opposing views, we have confirmed the functional concept that
the re-expression of Wwox protein leads to apoptosis in U266
multiple myeloma cells.
WWOX is closely related to apoptosis-associated
factors, including Bcl-2, Bcl-xL, Bax, p73 and caspases in its
antitumor activities (17–21),
indicating that WWOX may play a role in triggering
apoptosis-associated pathways. Previous studies have demonstrated
that WWOX overexpression results in a downregulation of Bcl-2, as
well as a cleavage of both pro-caspase-3 and -9 (19,20). It has also been shown that the
upregulation of WWOX in cancer cell lines enhances apoptosis by
activating the intrinsic apoptotic caspase cascade (8,9).
Cui et al (17) recently
presented similar results. The present study showed that the
ectopic expression of WWOX cDNA resulted in changes in the
expression of Bcl-2, cytochrome c, PARP, pro-caspase-3 and
pro-caspase-9, characterized by a lower expression of Bcl-2 and the
increased expression of cytochrome c, although Bax
expression was not altered significantly. The activation of
caspase-3 and -9 was further verified by immunoblotting and
immunofluorescence. Cleaved-PARP was also detected, which further
provides evidence of the activation of caspase-3 and -9. Taken
together, it these data suggest that WWOX induces cell death by
triggering the intrinsic apoptotic pathway in multiple myeloma.
Yet, our findings are supported in part and fully by those of
previous studies (8,9,17–21).
In conclusion, the present study reveals a
functional role of WWOX in U266 human multiple myeloma cells, as
well as the primary molecular mechanisms responsible for its
pro-apoptotic effects. However, in the present study we did not
evaluate the functional role of WWOX in multiple myeloma in
vivo and the clinical characterization of WWOX in multiple
myeloma patients. As regards the analysis of apoptosis, we failed
to evaluate the apoptotic ratio by flow cytometry marked with
PE/7-AAD fluorescent dye, due to the non-detectable cell fragments
that may affect by Wwox re-expression. The release of cytochrome
c from the mitochondria should also be verified. The
mechanisms underlying the molecular action of WWOX in multiple
myeloma warrant further investigation and more detailed analyses
need to be performed in future studies.
Acknowledgements
The authors thank Professor Donghong Lin for
providing editorial assistance.
References
|
1
|
Bednarek AK, Laflin KJ, Daniel RL, Liao Q,
Hawkins KA and Aldaz CM: WWOX, a novel WW domain-containing protein
mapping to human chromosome 16q23.3-24.1, a region frequently
affected in breast cancer. Cancer Res. 60:2140–2145.
2000.PubMed/NCBI
|
|
2
|
Iliopoulos D, Guler G, Han SY, et al:
Fragile genes as biomarkers: epigenetic control of WWOX and FHIT in
lung, breast and bladder cancer. Oncogene. 24:1625–1633. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lan C, Chenggang W, Yulan B, Xiaohui D,
Junhui Z and Xiao W: Aberrant expression of WWOX protein in
epithelial ovarian cancer: a clinicopathologic and
immunohistochemical study. Int J Gynecol Pathol. 31:125–132. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ekizoglu S, Muslumanoglu M, Dalay N and
Buyru N: Genetic alterations of the WWOX gene in breast cancer. Med
Oncol. 29:1529–1535. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo W, Wang G, Dong Y, Guo Y, Kuang G and
Dong Z: Decreased expression of WWOX in the development of
esophageal squamous cell carcinoma. Mol Carcinog. 52:265–274. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hu BS, Tan JW, Zhu GH, Wang DF, Zhou X and
Sun ZQ: WWOX induces apoptosis and inhibits proliferation of human
hepatoma cell line SMMC-7721. World J Gastroenterol. 18:3020–3026.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakayama S, Semba S, Maeda N, Matsushita
M, Kuroda Y and Yokozaki H: Hypermethylation-mediated reduction of
WWOX expression in intraductal papillary mucinous neoplasms of the
pancreas. Br J Cancer. 100:1438–1443. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fabbri M, Iliopoulos D, Trapasso F, et al:
WWOX gene restoration prevents lung cancer growth in vitro and in
vivo. Proc Natl Acad Sci USA. 102:15611–15616. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Iliopoulos D, Fabbri M, Druck T, Qin HR,
Han SY and Huebner K: Inhibition of breast cancer cell growth in
vitro and in vivo: effect of restoration of Wwox expression. Clin
Cancer Res. 13:268–274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aqeilan RI, Trapasso F, Hussain S, et al:
Targeted deletion of Wwox reveals a tumor suppressor function. Proc
Natl Acad Sci USA. 104:3949–3954. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aqeilan RI, Hassan MQ, de Bruin A, et al:
The WWOX tumor suppressor is essential for postnatal survival and
normal bone metabolism. J Biol Chem. 283:21629–21639. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Göthlin Eremo A, Wegman P, Stål O,
Nordenskjöld B, Fornander T and Wingren S: Wwox expression may
predict benefit from adjuvant tamoxifen in randomized breast cancer
patients. Oncol Rep. 29:1467–1474. 2013.PubMed/NCBI
|
|
13
|
Cancemi L, Romei C, Bertocchi S, et al:
Evidences that the polymorphism Pro-282-Ala within the tumor
suppressor gene WWOX is a new risk factor for differentiated
thyroid carcinoma. Int J Cancer. 129:2816–2824. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ishii H, Vecchione A, Furukawa Y, et al:
Expression of FRA16D/WWOX and FRA3B/FHIT genes in hematopoietic
malignancies. Mol Cancer Res. 1:940–947. 2003.PubMed/NCBI
|
|
15
|
Ishii H and Furukawa Y: Alterations of
common chromosome fragile sites in hematopoietic malignancies. Int
J Hematol. 79:238–242. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Zhang H, Li P, Yang Z, Qin L and
Mo W: Gene expression of WWOX, FHIT and p73 in acute lymphoblastic
leukemia. Oncol Lett. 6:963–969. 2013.PubMed/NCBI
|
|
17
|
Cui Z, Lin D, Cheng F, et al: The role of
the WWOX gene in leukemia and its mechanisms of action. Oncol Rep.
29:2154–2162. 2013.PubMed/NCBI
|
|
18
|
Lin D, Cui Z, Kong L, Cheng F, Xu J and
Lan F: p73 participates in WWOX-mediated apoptosis in leukemia
cells. Int J Mol Med. 31:849–854. 2013.PubMed/NCBI
|
|
19
|
Chang NS: A potential role of p53 and WOX1
in mitochondrial apoptosis (Review). Int J Mol Med. 9:19–24.
2002.PubMed/NCBI
|
|
20
|
Yang JL and Zhang W: WWOX tumor suppressor
gene. Histol Histopathol. 23:877–882. 2008.
|
|
21
|
Zhang P, Jia R, Ying L, Liu B, Qian G, Fan
X and Ge S: WWOX-mediated apoptosis in A549 cells mainly involves
the mitochondrial pathway. Mol Med Rep. 6:121–124. 2012.PubMed/NCBI
|
|
22
|
Płuciennik E, Kośla K, Wójcik-Krowiranda
K, Bieńkiewicz A and Bednarek AK: The WWOX tumor suppressor gene in
endometrial adenocarcinoma. Int J Mol Med. 32:1458–1464.
2013.PubMed/NCBI
|
|
23
|
Watanabe A, Hippo Y, Taniguchi H, et al:
An opposing view on WWOX protein function as a tumor suppressor.
Cancer Res. 63:8629–8633. 2003.PubMed/NCBI
|