Introduction
Glioblastomas are the most prevalent and aggressive
form of malignant primary brain tumor (1). The pathological process of and
treatment strategies for this disease have attracted increasing
attention over the years. A large number of studies have
demonstrated that several signaling pathways are dysregulated
during the multistage carcinogenesis of human glioblastomas in
general (2–7). There is also ample evidence of an
excessive activation of the epidermal growth factor receptor (EGFR)
pathway in the majority of glioblastomas (8). Traditionally, the nuclear factor-κB
(NF-κB) family, a set of transcription factors, plays key roles in
the regulation of the immune inflammatory response, cell growth and
apoptosis, and is considered to be the major downstream component
of the EGFR pathway (7,9). However, the molecular mechanisms
underlying the EGFR/NF-κB signal pathway in glioblastomas have not
been yet been fully elucidated.
The NF-κB family consists of a group of homodimeric
and heterodimeric protein complexes (9,10),
while the p50/p65 heterodimer is the most common complex in many
cell types (11,12). In unstimulated cells, inactive
NF-κB complexes are present in the cytoplasm and bind to a class of
inhibitor proteins known as NF-κB inhibitors (NFKBI), which include
NFKBIA, NFKBIB, IκBγ, IκBɛ, Bcl-3, p100 and p105 (13). In the majority of cases, the
activation of NF-κB involves the signal-induced degradation of
NFKBIA, thus releasing the transcription factor that translocates
to the nucleus (14–16). The aberrant constitutive
activation of NF-κB has been observed in glioblastomas (17–20). In addition, current data provide
evidence for a tumor suppressor role of NFKBIA in glioblastomas
(21). Therefore, considering
that NFKBIA plays pivotal roles in the regulation of the NF-κB
signaling pathway, in the present study, we focused on the role of
NFKBIA in glioblastomas.
The NFKBI protein family is characterized by the
presence of multiple ankyrin repeats and their ability to
physically associate with the NF-κB protein. Among the NFKBI
protein family, NFKBIA has 3 regions: an N-terminal region, which
has phosphorylation sites regulating signal-dependent degradation,
an ankyrin repeat domain and a C-terminal PEST region regulating
basal degradation (22–24). The NFKBIA gene is located on
chromosome 14 and presents 6 exons. Exon 1 encodes the N-terminal
region containing the serine residues that are important
phosphorylation sites. The ankyrin repeat domain is located in
exons 2 to 5, whereas the C-terminal PEST region is found in exon
6.
Mutations, polymorphisms and haplotypes of NFKBIA
have been reported in Hodgkin’s lymphoma, colorectal cancer,
melanoma, hepatocellular carcinoma, breast cancer and multiple
myeloma (25–37). Moreover, these studies have
indicated that these alterations render the NFKBIA protein
incapable of interacting with NF-κB, thus resulting in the loss of
NFKBIA activity and the protection of tumor cells from apoptosis.
However, to the best of our knowledge, there is still no relative
study available to date on the association between the genotype and
expression of NFKBIA, while the genotype of NFKBIA in glioblastomas
remains unclear.
Given that NFKBIA modulates NF-κB activity in
physiological and pathophysiological processes and that there are
polymorphisms of NFKBIA in several types of cancer, the present
study aimed to investigate the genotype of NFKBIA in glioblastomas
and whether this is associated with its expression.
Patients and methods
Patients and samples
The study was approved by the Ethics Committee of
Soochow University, Suzhou, China and written informed consent was
obtained from all subjects or a relative family member if the
patient was unable to provide it. All experiments complied with the
current laws of our country. Twenty-four glioblastoma samples with
matching clinical data were obtained from 24 Chinese patients (13
males and 11 females) at the Department of Neurosurgery of the
First Affiliated Hospital of Soochow University from March 2009 to
March 2011. The mean age of the patients at the time of surgery was
38 years (males) and 41 years (females). All tumors were obtained
from patients with a newly diagnosed glioblastoma, who had not
received any therapy prior to sample collection. Eight adult
non-cancerous brain tissue samples were obtained from the surgical
resections of 8 trauma patients, for whom a partial resection of
normal brain tissue was required as decompression treatment to
reduce intracranial pressure. Parts of these surgically removed
samples were immediately snap-frozen in liquid nitrogen. The
remaining samples were fixed with formalin and embedded in paraffin
for histological analyses.
PCR amplification and direct sequencing
of NFKBIA mutations
Sequences for the detection of mutations of the
NFKBIA coding region were determined using an ABI 3730xl Genetic
Analyzer (Applied Biosystems, Foster City, CA, USA). The following
primers were used: exon 1 forward, 5′-GAG GAC GAA GCC AGT TCT CT-3′
and reverse, 5′-CGC GAG GTT ATT ATG AGC TG-3′; exon 2 forward,
5′-GCC AGG AAC ACT CAG CTC AT-3′ and reverse, 5′-GGT GCT GCT CCT
CCT AGA CA-3′; exon 3 forward, 5′-CAC CTG GAG CCT CTG CTA TT-3′ and
reverse, 5′-AAG CTC TTG CCT GGA CTC CT-3′; exons 4 and 5 forward,
5′-GGA GTC CAG GCA AGA GCT TA-3′ and reverse, 5′-TCT GAT AAG GAG
CAG CTC TAG G-3′; and exon 6 forward, 5′-AGT AGT GGC CTC CCC ATC
C-3′ and reverse, 5′-AGG CAG TGT GCA GTG TGG ATA-3′. PCR was
performed under standard buffer conditions using 1 μl of DNA and
0.2 μl of Platinum® Taq DNA polymerase. The conditions
included 35 cycles of denaturation at 94°C for 5 min, annealing at
94°C for 30 sec, 55°C for 30 sec, and 72°C for 30 sec, and
extension at 72°C for 5 min, in a total volume of 25 μl.
Quantitative reverse transcription PCR
(RT-qPCR)
Total RNA was extracted from the samples of tumor
and control frozen tissue using TRIzol reagent (Invitrogen,
Carslbad, CA, USA), according to the manufacturer’s instructions.
Subsequently, 2 μg of total RNA were reverse-transcribed in a 20 μl
reaction containing 10 units of M-MLV reverse transcriptase and 0.5
μg of oligo (dT) primer. A total of 2 μl of cDNA was used for qPCR.
The following primers were used: NFKBIA forward, 5′-CTC CGA GAC TTT
CGA GGA AAT AC-3′ and reverse, 5′-GCC ATT GAA GTT GGT AGC CTT
CA-3′, telomerase reverse transcriptase (TERT) forward, 5′-GAG CGT
GTG ACT TCC GAA GG-3′ and reverse, 5′-AGG AAC TGT CAC GGA GTT
TGC-3′. The NFKBIA and TERT genes were amplified using the
SYBR-Green PCR Master Mix (Takara Bio, Inc., Otsu, Japan).
Following 3 min of initial denaturation at 95°C, the cycling
conditions were 40 cycles consisting of denaturation at 95°C for 10
sec followed by annealing and extension at 60°C for 30 sec. The
results are presented as CT values, defined as the threshold PCR
cycle number at which an amplified product was first detected. The
average CT value was calculated for both NFKBIA and TERT, and the
ΔCT value was determined as the mean of the triplicate CT values
for NFKBIA minus the mean of the triplicate CT values for TERT. The
2−ΔΔCT method was used to analyze the relative changes
in gene expression.
Copy number variation analysis
RT-qPCR reactions were performed to assess NFKBIA
gene expression levels in the samples using an ABI Prism 7500 Fast
Real-Time PCR System (Applied Biosystems). The target gene (NFKBIA)
TaqMan primer pair was simultaneously amplified with an endogenous
control TaqMan primer pair targeting the TERT gene located on
5p15.33, which has a diploid status in gliomas. A total of 20 μg of
genomic DNA from tumor and non-cancerous brain tissue was used for
each reaction. Thermocycling for each PCR reaction was carried out
in a final volume of 20 μl containing 20 ng of gDNA, 1 μl of 20X
TaqMan™ Gene Copy Number assay probe (6-FAM dye-labeled), 1 μl of
20X TaqMan® TERT Copy Number Reference assay probe (VIC
dye-labeled), 4 μl of nuclease-free water, and 2X
TaqMan® Genotyping Master mix (Invitrogen). After 3 min
of initial denaturation at 95°C, the cycling conditions of 40
cycles consisted of denaturation at 95°C for 10 sec followed by
annealing and extension at 60°C for 30 sec. All reactions were
performed in triplicate. The 2−ΔΔCT method was also used
to analyze the relative changes in gene copy number. The ΔCT value
was determined as the mean of the triplicate CT values for NFKBIA
minus the mean of the triplicate CT values for TERT. There was no
control group and thus, ΔCT was treated as ΔΔCT.
Western blot analysis
Total protein from the glioblastoma samples and
non-cancerous brain tissue samples was extracted directly in lysis
buffer, and the concentration of total protein was quantified using
an ultraviolet spectrophotometer. A total of 100 or 50 μg of
proteins was separated on 12% gels by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins
were then transferred onto nitrocellulose membranes and
non-specific binding was blocked by incubating the membranes in 5%
non-fat milk. Monoclonal rabbit anti-human antibody NFKBIA (Clone
ID: EP697; Epitomics, San Francisco, CA, USA) was used as the
primary antibody. To confirm equal loading, the membranes were
stripped and reprobed with rabbit anti-human β-actin antibody
(1:1000; R&D Systems, Inc., Minneapolis, MN, USA). Horseradish
peroxidase-conjugated goat anti-mouse antibody (ProSci Inc., Poway,
CA, USA) was used as the secondary antibody. Protein bands were
visualized using ECL western blot analysis detection reagents
(Pierce antibodies; Thermo Fisher Scientific, Waltham, MA,
USA).
Immunohistochemistry
The sections were deparaffinized in xylene and
rehydrated in graded alcohols. Following deparaffinization, antigen
retrieval was performed by immersing the sections in 10 mmol
citrate buffer (pH 6.0) and heating them twice in a microwave oven
(95°C) for 5 min. The sections were incubated with primary
antibodies against NFKBIA (Clone ID: EP697; Epitomics; dilution
1:100 overnight at 4°C) and NF-κB1 (Abcam, Tokyo, Japan). The
sections were subsequently incubated using the Cell and Tissue
Staining kit HRP-DAB system (R&D Systems), according to the
manufacturer’s instructions. Immunostainings were performed with
known positive and negative tumor controls, and were blindly
evaluated by a pathologist.
Statistical analysis
Survival curves were estimated using the
Kaplan-Meier product-limit method, and survival distributions were
compared across groups using the log-rank test. The genotype
frequencies were determined by direct counting in the control and
patient groups. All genotype frequencies were tested for
conformation to the Hardy-Weinberg equilibrium. Where appropriate,
we used one-way analysis of variance, Student-Newman-Keuls (SNK)
test, least significant difference procedure (LSD), Pearson’s
χ2 test and Fisher’s exact test. Statistical analyses
were performed using SPSS 17.0 software (SPSS, Inc., Chicago, IL,
USA). Values of P<0.05 were considered to indicate statistically
significant differences.
Results
NFKBIA single nucleotide polymorphism
(SNP) in glioblastoma samples and non-cancerous brain tissue
samples
We analyzed 24 glioblastomas and 8 non-cancerous
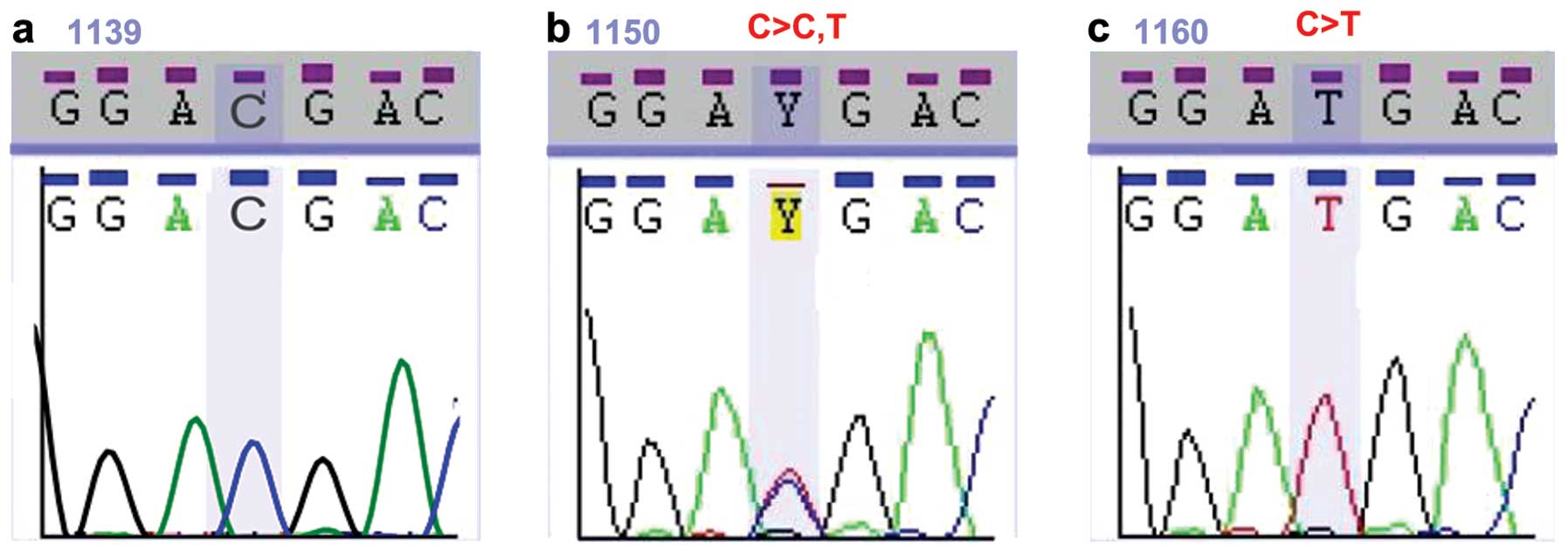
brain tissue samples. As shown in Fig. 1b, 2 different nucleotides at the same
position within the NFKBIA gene appeared as double peaks in the
chromatogram. These changes were identified as polymorphisms and
not mutations, as identical sequences were found in the
glioblastoma samples and non-cancerous brain tissue samples.
Fig. 1a–c represents the CC, CT
and TT genotypes of the SNP rs1957106, respectively. Table I summarizes the polymorphisms
found in the NFKBIA gene of the glioblastoma and non-cancerous
brain samples. Through direct sequencing of PCR-amplified products,
we identified a single polymorphism of NFKBIA. The prevalence of
this polymorphism (rs1957106) in exon 1 was found to be greater in
the glioblastoma samples, although the difference was not
statistically significant (Table
I). The SNP was located on amino acid 27 of NFKBIA and induced
no change in the amino acid.
 | Table IGenotype frequencies in glioblastoma
and non-cancerous patients. |
Table I
Genotype frequencies in glioblastoma
and non-cancerous patients.
| Accession code | Genotype | Change | Glioblastoma
samples (n=24) | Non-cancerous
samples (n=8) | OR | 95% CI | P-value |
|---|
|
|
|
|---|
| n | (%) | n | (%) | Lower | Upper |
|---|
| rs1957106 | CC | - | 14 | 58.3 | 6 | 75 | 0.467 | 0.080 | 2.810 | 0.340 |
| CT | C>CT | 8 | 33.3 | 2 | 25 | 1.5 | 0.245 | 9.179 | 0.512 |
| TT | C>T | 2 | 12.5 | 0 | 0 | 0.917 | 0.813 | 1.034 | 0.556 |
| | | HWE (Pa=0.5862) | HWE (Pa=0.6862) | | | | |
Association between different genotypes
of SNP rs1957106 in NFKBIA and the protein expression of NFKBIA
detected by western blot analysis
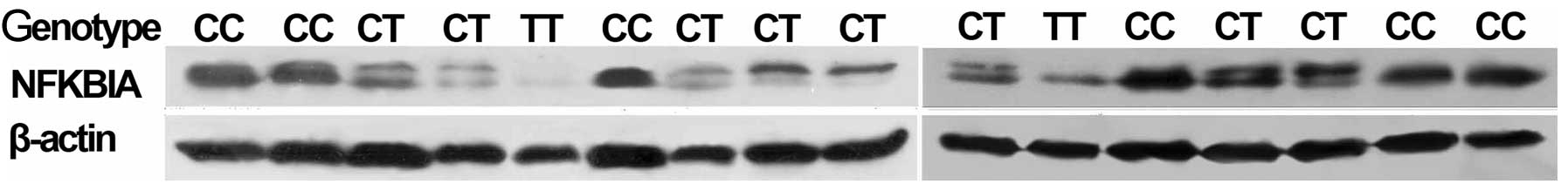
As shown in Fig.
2, the NFKBIA protein levels detected by western blot analysis
(using 100 ng of sample) were significantly lower in the
glioblastomas harboring the SNP rs1957106 CT and TT genotypes than
in the samples harboring the SNP rs1957106 CC genotype. On the blot
images, the CT and TT genotypes show 2 parallel bands, representing
the mature protein and its degradation precursor, as prevoiusly
described (38). In the CC
genotype, the 2 bands were merged. Similarly, there was only 1 band
in the non-cancerous brain tissue samples.
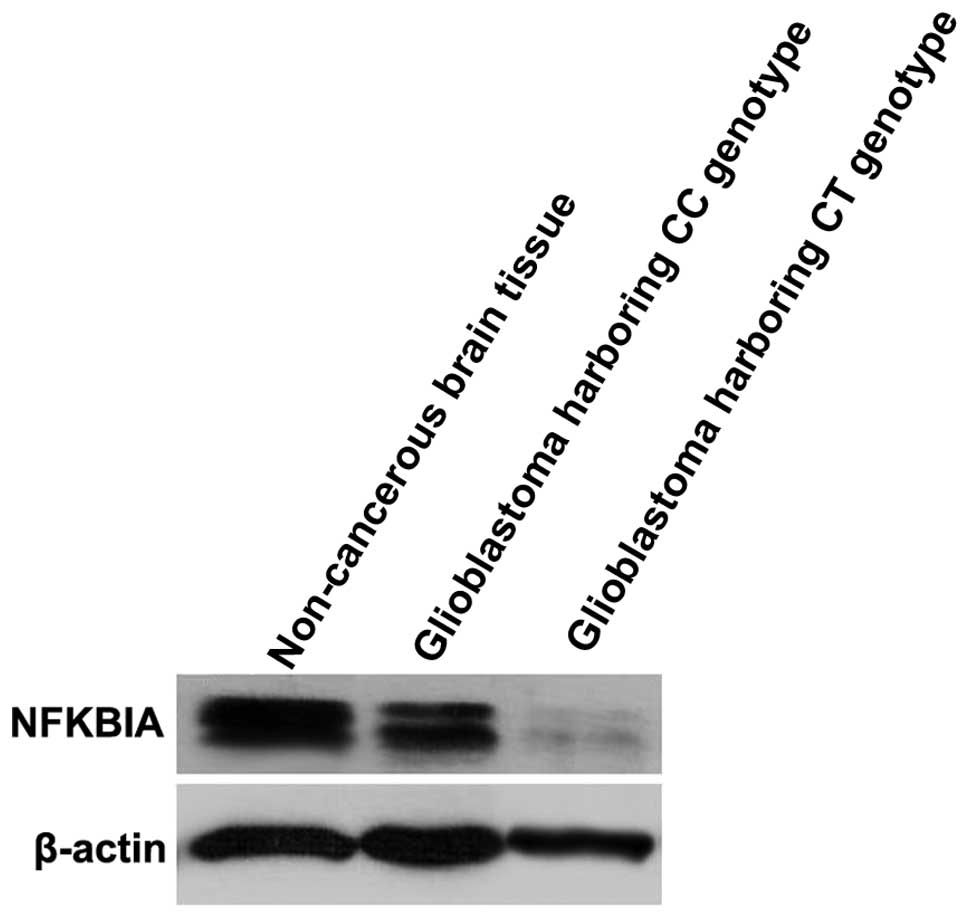
In order to examine whether there were significant
differences in the expression of NFKBIA protein in glioblastomas
compared with non-cancerous brain tissues, we decreased the amount
of protein sample. As shown in Fig.
3, the NFKBIA protein levels detected by western blot analysis
(using 50 ng of protein sample) were significantly lower in the
glioblastomas harboring the CC and CT genotypes than in the
non-cancerous brain tissue samples.
Association between different genotypes
of SNP rs1957106 in NFKBIA and the relative mRNA expression levels
of NFKBIA
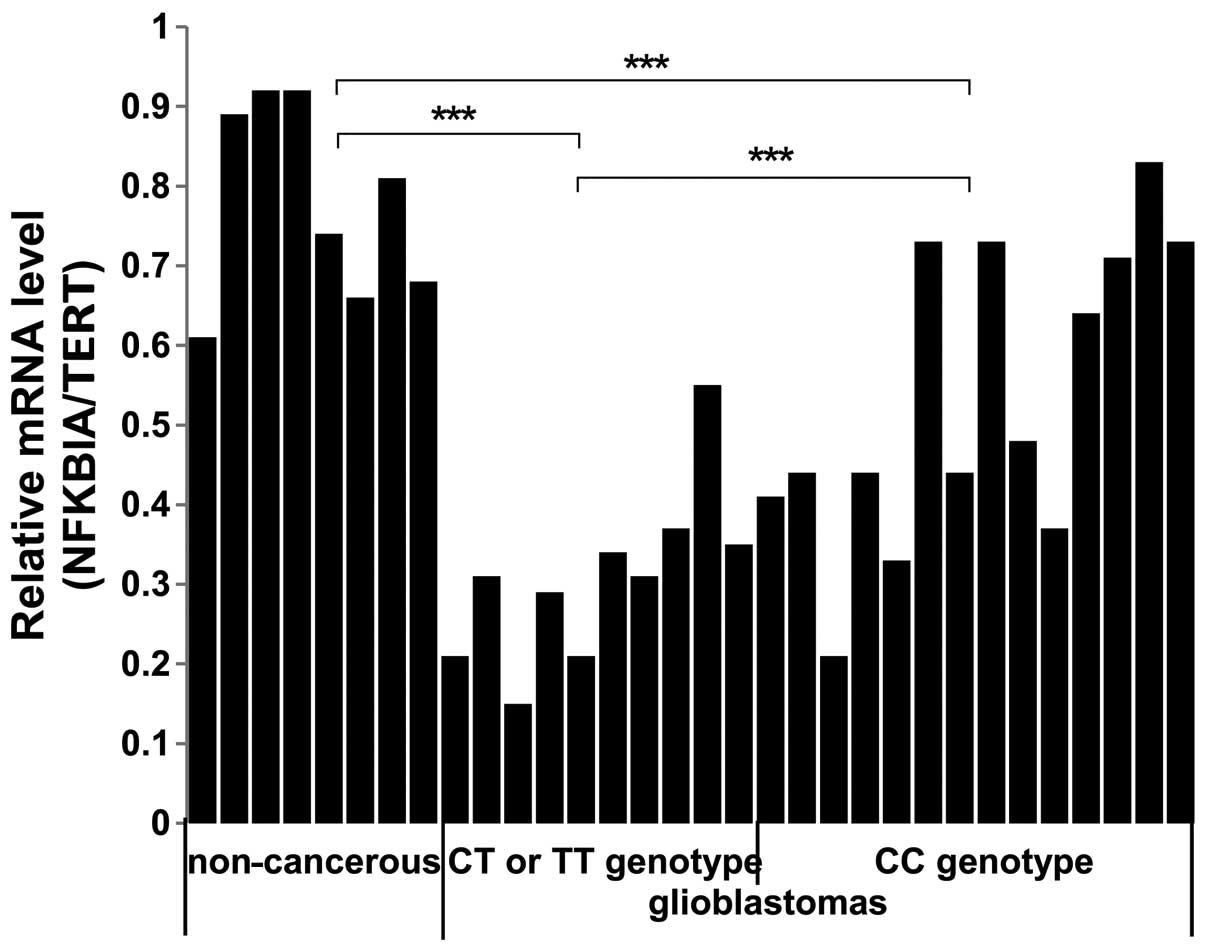
We performed qPCR to examine whether there were
significant differences in the mRNA expression of NFKBIA in
glioblastomas compared with non-cancerous brain tissue samples and
in glioblastomas harboring different genotypes of SNP rs1957106. As
shown in Fig. 4, the NFKBIA mRNA
levels were lower in the glioblastoma compared with the
non-cancerous brain tissue samples. Furthermore, the NFKBIA mRNA
levels were significantly lower in the glioblastomas harboring the
SNP rs1957106 CT and TT genotypes than in the samples harboring the
SNP rs1957106 CC genotype (CT/TT 0.31±0.11 vs. CC 0.54±0.18 vs.
non-cancerous 0.78±0.12, P<0.001). These data suggest that the
lower protein expression levels of NFKBIA are in accordance with
the lower mRNA expression levels of NFKBIA in glioblastomas.
Association between different genotypes
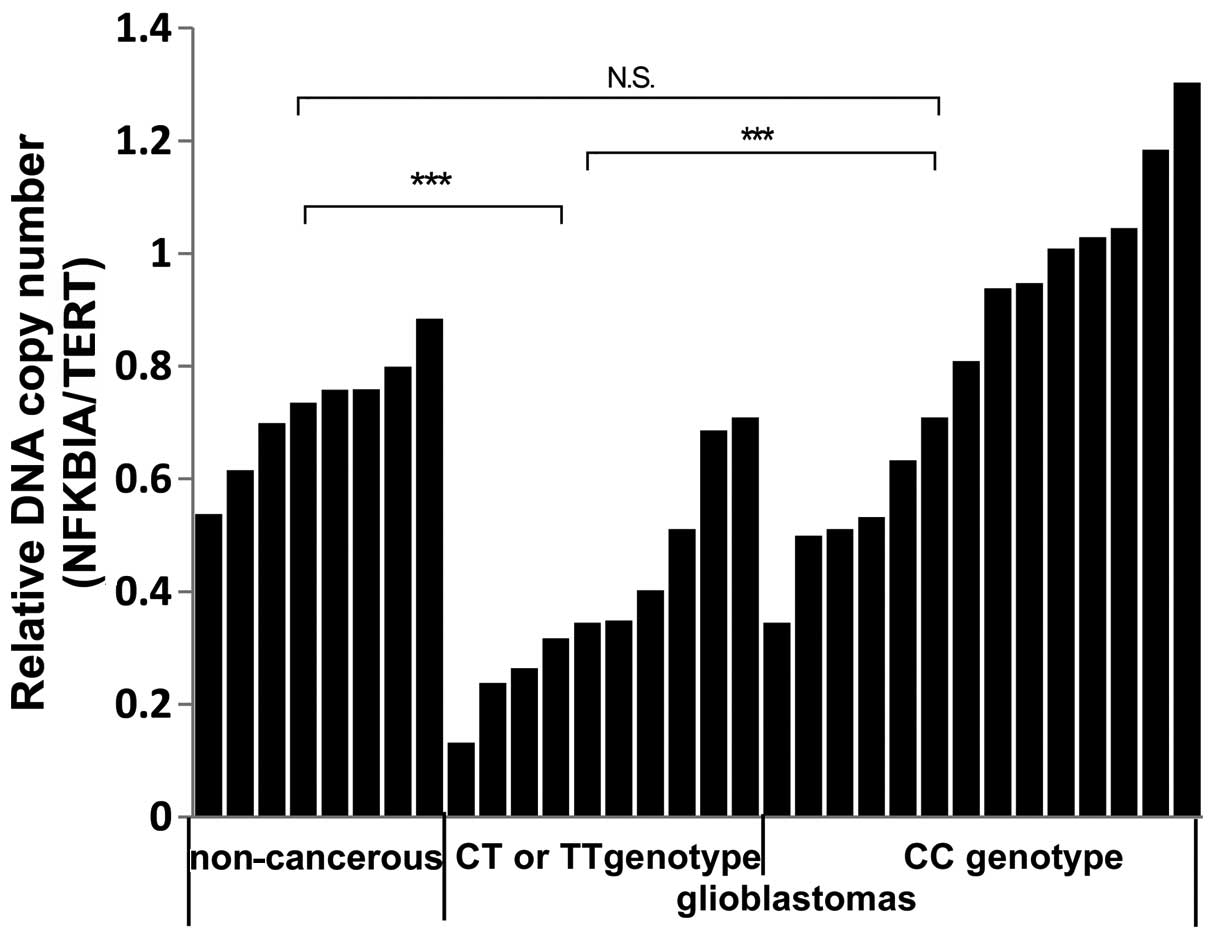
of SNP rs1957106 in NFKBIA and the relative copy number of NFKBIA
in glioblastoma tissue samples
In order to examine whether there was a variation in
the copy number of NFKBIA, we used a specific TaqMan®
primer pair producing the allele, rs1957106. We detected the copy
number of NFKBIA in 24 cases of glioblastoma samples and in 8 cases
of non-cancerous brain tissue samples. We observed that the
relative copy number of NFKBIA was significantly lower in 7 of the
10 glioblastomas, all of which had the SNP rs1957106 CT and TT
genotypes and low NFKBIA protein levels. Thus, it is possible that
some CT and TT genotypes are accompanied by a decrease in the
NFKBIA copy number (Fig. 5).
Association between different genotypes
of SNP rs1957106 in NFKBIA and the protein expression of NFKBIA and
NF-κB1 detected by immunohistochemistry
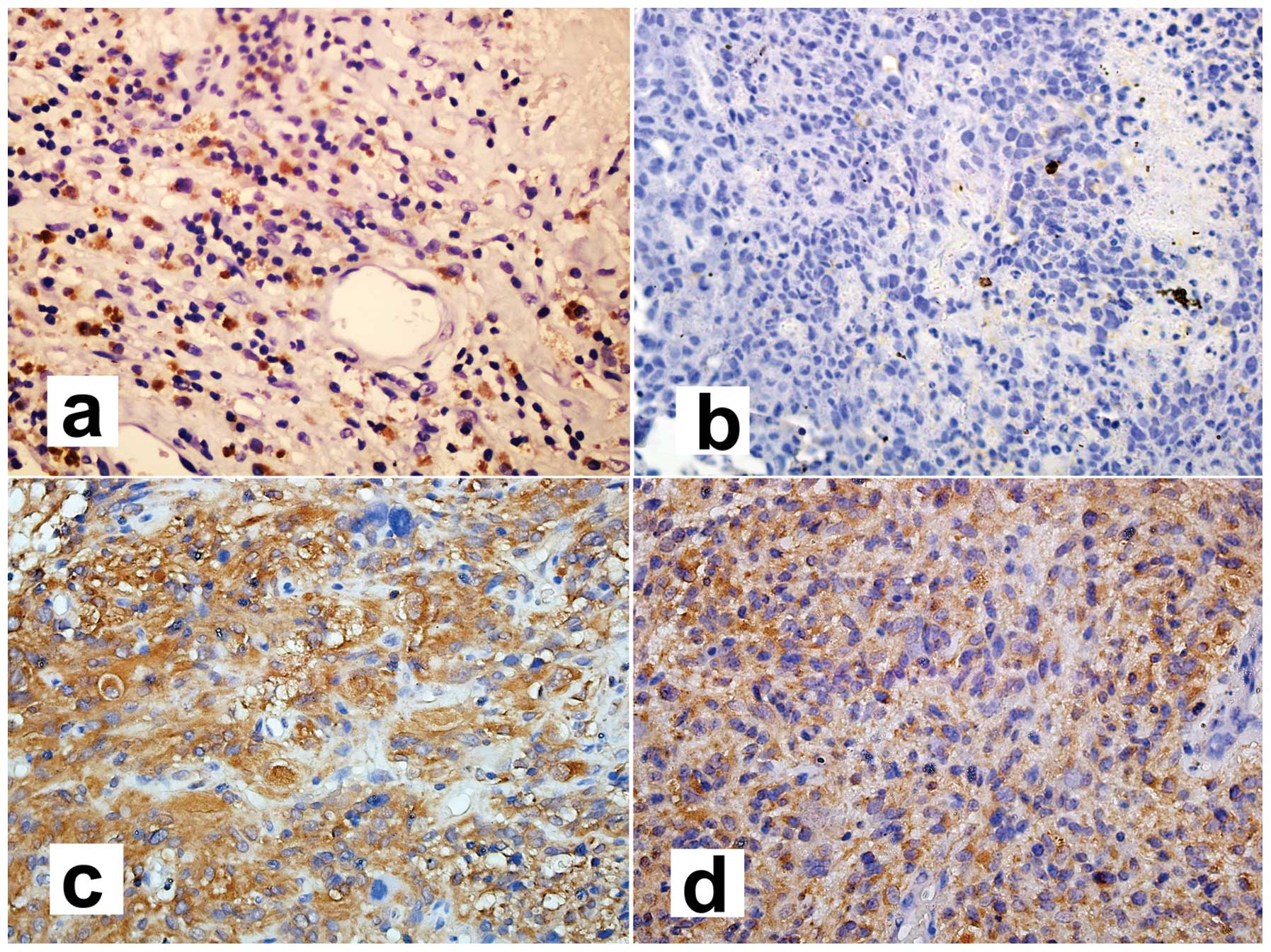
Immunohistochemical staining for NFKBIA and NF-κB1
protein is illustrated in Fig. 6.
Positivity is indicated by brown cytosolic staining. The intensity
of staining indicated the level of protein expression according to
the percentage of stained tumor cells. NF-κB1 expression was
classified as ‘low’ (<33% positive carcinoma cells),
‘intermediate’ (≥33 and <66% positive carcinoma cells) or ‘high’
(≥66% positive carcinoma cells). There was no protein expression of
NFKBIA in the non-cancerous brain tissue and glioblastoma samples,
and only 1 case of pilocytic astrocytoma showed staining for NFKBIA
in a total of 94 gliomas (from tissue microarray analysis in our
laboratory). There was a high NF-κB1 expression in 9 of 10
glioblastomas harboring the SNP rs1957106 CT and TT genotypes, and
in 6 of 14 glioblastomas harboring the SNP rs1957106 CC genotype;
the difference was statistically significant (P=0.033) (data not
shown). This finding suggests that this SNP affects the expression
of NF-κB1 through the downregulation of NFKBIA.
Association between different genotypes
of SNP rs1957106 in NFKBIA and outcomes in patients with
glioblastoma
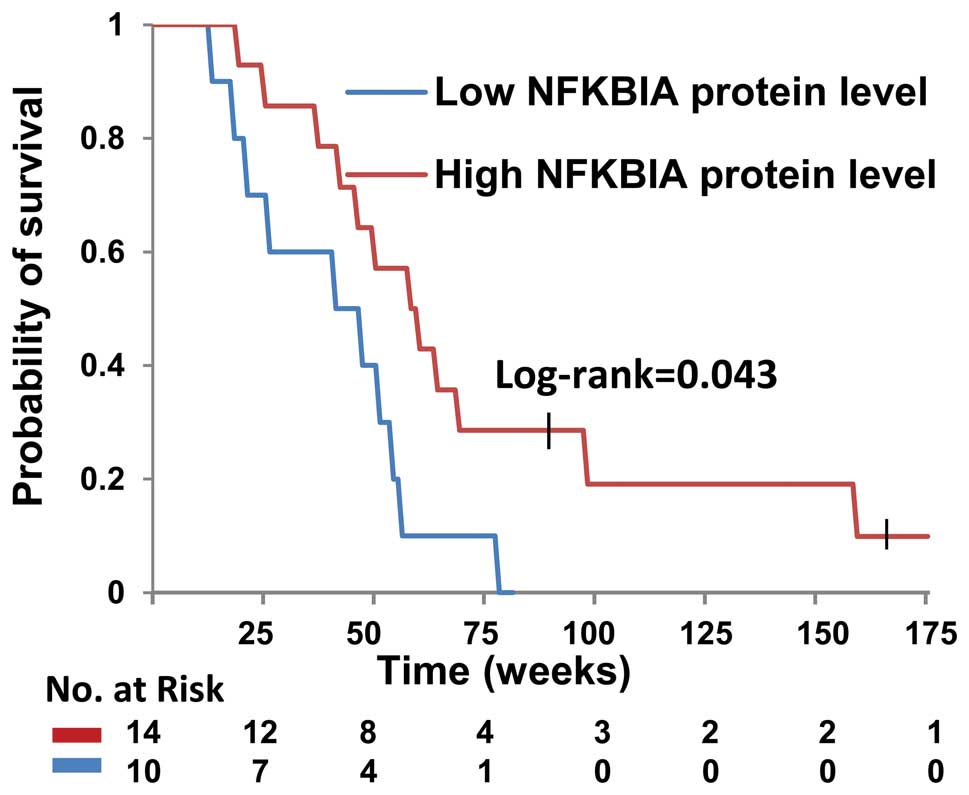
Patients with glioblastoma harboring the SNP
rs1957106 CT or TT genotypes had a significantly shorter overall
survival than those harboring the SNP rs1957106 CC genotype
(Fig. 7). The estimated median
survival times were 43 weeks for the patients harboring the SNP
rs1957106 CT and TT genotypes, and 55 weeks for those harboring the
SNP rs1957106 CC genotype. Furthermore, no statistically
significant differences were observed as regards the outcome of
patients harboring the different genotypes of SNP rs1957106 in
NFKBIA in relation to age, gender, tumor size, tumor surgery, tumor
radiotherapy or chemotherapy (Table
II).
| Table IIAssociation between outcome and
clinicopathologic characteristics in patients with glioblastoma
harboring different genotypes of SNP rs1957106 in NFKBIA. |
Table II
Association between outcome and
clinicopathologic characteristics in patients with glioblastoma
harboring different genotypes of SNP rs1957106 in NFKBIA.
| | Glioblastoma
patients | |
|---|
| |
| |
|---|
| Characteristic | n | CT/TT genotype | CC genotype | P-value |
|---|
| Gender |
| Male | 13 | 6 | 7 | 0.647 |
| Female | 11 | 5 | 6 | |
| Age (years) |
| <60 | 19 | 8 | 11 | 0.668 |
| ≥60 | 5 | 2 | 3 | |
| Surgery |
| Total | 20 | 8 | 12 | 0.563 |
| Subtotal | 4 | 2 | 2 | |
| Radiotherapy | 24 | All | All | - |
| Chemotherapy |
| Temozolomide | 14 | 6 | 8 | 0.500 |
| Me-CCNU+VM26 | 10 | 4 | 6 | |
| Tumor size
(cm) |
| <5 | 17 | 7 | 10 | 0.643 |
| ≥5 | 7 | 3 | 4 | |
Discussion
Genetic polymorphisms in NFKBIA have attracted much
interest due to their potential role in the etiology and outcomes
of Hodgkin’s lymphoma, colorectal cancer, melanoma, hepatocellular
carcinoma, breast cancer and multiple myeloma (25–37). In the present study, we determined
whether alterations of this gene also occur in glioblastomas. We
analyzed the DNA sequence of the NFKBIA gene from brain tissue
samples obtained from 24 glioblastomas patients and 8 non-cancerous
subjects. We identified the SNP rs1957106 as a novel polymorphism.
We also detected that CT and TT genetypes were associated with a
decreased gene copy number, as well as a decreased mRNA and protein
level of NFKBIA. Taken together, these results provide the basis
for the future evaluation of the role of the NF-κB/NF-KBIA pathway
in glioblastoma patients.
Our results demonstrated that CT and TT genetypes
were associated with decreased mRNA and protein levels of NFKBIA,
which suggests that SNP rs1957106 affects the expression of NFKBIA.
Individual and/or combinations of SNPs within the NFKBIA gene may
affect the expression and function of the protein. Allelic
differences in the NFKBIA promoter and the 3′-UTR may alter NFKBIA
expression, while allelic differences located around important
sites, such as in exon 1, may determine the rate of NFKIBA protein
degradation, and these 2 factors may influence the formation of the
NF-κB/NFKBIA complex, subsequently affecting cell growth and
apoptosis. In addition, the prevalence of this polymorphism was not
significantly higher in the glioblastoma patients compared with the
non-cancerous subjects. This finding suggests that this SNP has a
functional significance, but only in some special contexts.
Alternatively, the SNP itself may be in linkage disequilibrium with
a polymorphism directly influencing NFKBIA expression located
either upstream or downstream of the gene.
The observation that the relative copy number of
NFKBIA was significantly lower in glioblastomas that had the SNP
rs1957106 CT and TT genotypes and low NFKBIA protein levels,
suggests that the lower protein and mRNA levels of NFKBIA were at
least partially due to the decrease in the NFKBIA copy number. The
polymorphism may also affect the transcription of the NFKBIA
gene.
Our results also indicate that synonymous SNPs, such
as NFKBIA SNP rs1957106, may have biological significance. The
NFKBIA SNP rs1957106 has previously been reported in gastric
cancer, hepatocellular carcinoma, multiple myeloma and epithelial
ovarian cancer (32,34,37,39). White et al (39) found that the minor allele for the
synonymous rs1957106 SNP in NFKBIA was associated with a decreased
risk of epithelial ovarian cancer by querying 19 tag SNPs and
putative-functional SNPs in NFKBIA and NFKBIB among 930 epithelial
ovarian cancer cases and 1,037 controls. However, considering the
number of single-SNP tests performed and the null gene-level
results, they concluded that NFKBIA and NFKBIB were not likely to
harbor any alleles associated with the risk of ovarian cancer.
As described above, EGF has been demonstrated to
activate NF-κB (40–44). NFKBIA degradation preceded by the
phosphorylation of NFKBIA is critical for NF-κB activation. EGF
induces NFKBIA phosphorylation at tyrosine 42 within exon 1
(45) and tumor necrosis factor
(TNF) induces NFKBIA phosphorylation at serines 32 and 36 within
exon 1 (46–49). In brief, phosphorylation plays an
important role in the regulation of NFKBIA degradation. However, as
this SNP is located on amino acid 27 of NFKBIA, the association
between genotype and phosphorylation requires further
investigation.
Our observations revealed that the SNP rs1957106 CT
and TT genotypes in glioblastomas were associated with a
comparatively shorter survival rate. Patients with the SNP
rs1957106 CT or TT genotype had a lower NFKBIA expression. Thus,
our data support a role for NFKBIA in the suppression of
glioblastoma tumors. Our results are in agreement with those
presented in the study by Bredel et al (21), who demonstrated that a higher
NFKBIA expression was associated with a longer survival by studying
a wider sample of patients. They also indicated that glioblastoma
cells that did not respond to temozolomide chemotherapy had lower
mRNA expression levels of NFKBIA, and that the increased expression
of NFKBIA inhibited the malignant behavior of the tumor. Our
observations suggest that it would be useful to include the SNP or
NFKBIA expression in models predicting survival. Our findings,
taken together with those by Bredel et al (21), suggest that NFKBIA-stabilizing
therapies may be effective against glioblastomas. The limited
efficacy of molecular therapies targeting EGFR in glioblastomas
suggests that the therapeutic efficacy of EGFR inhibition may be
circumvented through cross-coupled signaling from other growth
factor receptors that are mutated, amplified or overexpressed in
these tumors, such as platelet-derived growth factor receptor,
alpha polypeptide (PDGFRA), ERBB2 and MET (7). Due to the fact that NFKBIA is a
major node downstream of such cross-coupled signaling, therapies
stabilizing NFKBIA may inhibit oncogenic signaling more
effectively.
In addition, Bredel et al (21) reported that there were no
alterations (or mutations) of NFKBIA in either coding or promoter
sequences in glioblastoma patients (from Western countries). Our
results differ from those by presented in the study by Bredel et
al (21). It is possible that
these differences are due to racial differences.
In conclusion, and to the best of our knowledge, the
findings of the present study provide the first evidence that the
rs1957106 SNP in NFKBIA is frequently present in glioblastoma
patients. Specific pharmacological targeting of the SNP rs1957106
CC genotype may aid in the development of novel therapeutic
strategies for glioblastoma.
Acknowledgements
This study was partially supported by the Foundation
of Health Department in Jiangsu Province (Grant no. K201106).
References
|
1
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parsons DW, Jones S, Zhang X, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, Olivi
A, McLendon R, Rasheed BA, Keir S, Nikolskaya T, Nikolsky Y, Busam
DA, Tekleab H, Diaz LA Jr, Hartigan J, Smith DR, Strausberg RL,
Marie SK, Shinjo SM, Yan H, Riggins GJ, Bigner DD, Karchin R,
Papadopoulos N, Parmigiani G, Vogelstein B, Velculescu VE and
Kinzler KW: An integrated genomic analysis of human glioblastoma
multiforme. Science. 321:1807–1812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yan H, Parsons DW, Jin G, McLendon R,
Rasheed BA, Yuan W, Kos I, Batinic-Haberle I, Jones S, Riggins GJ,
Friedman H, Friedman A, Reardon D, Herndon J, Kinzler KW,
Velculescu VE, Vogelstein B and Bigner DD: IDH1 and IDH2 mutations
in gliomas. N Engl J Med. 360:765–773. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Phillips HS, Kharbanda S, Chen R, Forrest
WF, Soriano RH, Wu TD, Misra A, Nigro JM, Colman H, Soroceanu L,
Williams PM, Modrusan Z, Feuerstein BG and Aldape K: Molecular
subclasses of high-grade glioma predict prognosis, delineate a
pattern of disease progression, and resemble stages in
neurogenesis. Cancer Cell. 9:157–173. 2006. View Article : Google Scholar
|
|
5
|
Bredel M, Scholtens DM, Harsh GR, Bredel
C, Chandler JP, Renfrow JJ, Yadav AK, Vogel H, Scheck AC,
Tibshirani R and Sikic BI: A network model of a cooperative genetic
landscape in brain tumors. JAMA. 302:261–275. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yadav AK, Renfrow JJ, Scholtens DM, Xie H,
Duran GE, Bredel C, Vogel H, Chandler JP, Chakravarti A, Robe PA,
Das S, Scheck AC, Kessler JA, Soares MB, Sikic BI, Harsh GR and
Bredel M: Monosomy of chromosome 10 associated with dysregulation
of epidermal growth factor signaling in glioblastomas. JAMA.
302:276–289. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cancer Genome Atlas Research Network.
Comprehensive genomic characterization defines human glioblastoma
genes and core pathways. Nature. 455:1061–1068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Watanabe K, Tachibana O, Sata K, Yonekawa
Y, Kleihues P and Ohgaki H: Overexpression of the EGF receptor and
p53 mutations are mutually exclusive in the evolution of primary
and secondary glioblastomas. Brain Pathol. 6:217–224. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bargou RC, Leng C, Krappmann D, Emmerich
F, Mapara MY, Bommert K, Royer HD, Scheidereit C and Dörken B:
High-level nuclear NF-kappa B and Oct-2 is a common feature of
cultured Hodgkin/Reed-Sternberg cells. Blood. 87:4340–4347.
1996.PubMed/NCBI
|
|
10
|
Grüssel T and Busch W: Experimental
studies of the effect of peracetic acid on the endometrium of
cattle. Tierarztl Prax. 25:28–34. 1997.(In German).
|
|
11
|
Barkett M and Gilmore TD: Control of
apoptosis by Rel/NF-kappaB transcription factors. Oncogene.
18:6910–6924. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
May MJ and Ghosh S: Signal transduction
through NF-kappa B. Immunol Today. 19:80–88. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chiao PJ, Miyamoto S and Verma IM:
Autoregulation of I kappa B alpha activity. Proc Natl Acad Sci USA.
91:28–32. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baeuerle PA and Baltimore D: NF-kappa B:
ten years after. Cell. 87:13–20. 1996.PubMed/NCBI
|
|
15
|
Matthews JR, Nicholson J, Jaffray E, Kelly
SM, Price NC and Hay RT: Conformational changes induced by DNA
binding of NF-kappa B. Nucleic Acids Res. 23:3393–3402. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Roulston A, Lin R, Beauparlant P, Wainberg
MA and Hiscott J: Regulation of human immunodeficiency virus type 1
and cytokine gene expression in myeloid cells by NF-kappa B/Rel
transcription factors. Microbiol Rev. 59:481–505. 1995.PubMed/NCBI
|
|
17
|
Raychaudhuri B, Han Y, Lu T and Vogelbaum
MA: Aberrant constitutive activation of nuclear factor kappaB in
glioblastoma multiforme drives invasive phenotype. J Neurooncol.
85:39–47. 2007. View Article : Google Scholar
|
|
18
|
Nagai S, Washiyama K, Kurimoto M, Takaku
A, Endo S and Kumanishi T: Aberrant nuclear factor-kappaB activity
and its participation in the growth of human malignant astrocytoma.
J Neurosurg. 96:909–917. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bredel M, Bredel C, Juric D, Duran GE, Yu
RX, Harsh GR, Vogel H, Recht LD, Scheck AC and Sikic BI: Tumor
necrosis factor-alpha-induced protein 3 as a putative regulator of
nuclear factor-kappaB-mediated resistance to O6-alkylating agents
in human glioblastomas. J Clin Oncol. 24:274–287. 2006. View Article : Google Scholar
|
|
20
|
Tsunoda K, Kitange G, Anda T, Shabani HK,
Kaminogo M, Shibata S and Nagata I: Expression of the
constitutively activated RelA/NF-kappaB in human astrocytic tumors
and the in vitro implication in the regulation of urokinase-type
plasminogen activator, migration, and invasion. Brain Tumor Pathol.
22:79–87. 2005. View Article : Google Scholar
|
|
21
|
Bredel M, Scholtens DM, Yadav AK, Alvarez
AA, Renfrow JJ, Chandler JP, Yu IL, Carro MS, Dai F, Tagge MJ,
Ferrarese R, Bredel C, Phillips HS, Lukac PJ, Robe PA, Weyerbrock
A, Vogel H, Dubner S, Mobley B, He X, Scheck AC, Sikic BI, Aldape
KD, Chakravarti A and Harsh GR IV: NFKBIA deletion in
glioblastomas. N Engl J Med. 364:627–637. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Davis N, Ghosh S, Simmons DL, Tempst P,
Liou HC, Baltimore D and Bose HR Jr: Rel-associated pp40: an
inhibitor of the rel family of transcription factors. Science.
253:1268–1271. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Haskill S, Beg AA, Tompkins SM, Morris JS,
Yurochko AD, Sampsonjohannes A, Mondal K, Ralph P and Baldwin AS
Jr: Characterization of an immediate-early gene induced in adherent
monocytes that encodes I-kappa-B-like activity. Cell. 65:1281–1289.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jacobs MD and Harrison SC: Structure of an
IkappaBalpha/NF-kappaB complex. Cell. 95:749–758. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Spink CF, Gray LC, Davies FE, Morgan GJ
and Bidwell JL: Haplotypic structure across the I kappa B alpha
gene (NFKBIA) and association with multiple myeloma. Cancer Lett.
246:92–99. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Krappmann D, Emmerich F, Kordes U,
Scharschmidt E, Dörken B and Scheidereit C: Molecular mechanisms of
constitutive NF-kappaB/Rel activation in Hodgkin/Reed-Sternberg
cells. Oncogene. 18:943–953. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cabannes E, Khan G, Aillet F, Jarrett RF
and Hay RT: Mutations in the IkBa gene in Hodgkin’s disease suggest
a tumour suppressor role for IkappaBalpha. Oncogene. 18:3063–3070.
1999.PubMed/NCBI
|
|
28
|
Emmerich F, Meiser M, Hummel M, Demel G,
Foss HD, Jundt F, Mathas S, Krappmann D, Scheidereit C, Stein H and
Dörken B: Overexpression of I kappa B alpha without inhibition of
NF-kappaB activity and mutations in the I kappa B alpha gene in
Reed-Sternberg cells. Blood. 94:3129–3134. 1999.PubMed/NCBI
|
|
29
|
Jungnickel B, Staratschek-Jox A,
Brauninger A, Spieker T, Wolf J, Diehl V, Hansmann ML, Rajewsky K
and Kuppers R: Clonal deleterious mutations in the IkappaBalpha
gene in the malignant cells in Hodgkin’s lymphoma. J Exp Med.
191:395–402. 2000.PubMed/NCBI
|
|
30
|
Lake A, Shield LA, Cordano P, Chui DT,
Osborne J, Crae S, Wilson KS, Tosi S, Knight SJ, Gesk S, Siebert R,
Hay RT and Jarrett RF: Mutations of NFKBIA, encoding IkappaBalpha,
are a recurrent finding in classical Hodgkin lymphoma but are not a
unifying feature of non-EBV-associated cases. Int J Cancer.
125:1334–1342. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sjöblom T, Jones S, Wood LD, Parsons DW,
Lin J, Barber TD, Mandelker D, Leary RJ, Ptak J, Silliman N, Szabo
S, Buckhaults P, Farrell C, Meeh P, Markowitz SD, Willis J, Dawson
D, Willson JK, Gazdar AF, Hartigan J, Wu L, Liu C, Parmigiani G,
Park BH, Bachman KE, Papadopoulos N, Vogelstein B, Kinzler KW and
Velculescu VE: The consensus coding sequences of human breast and
colorectal cancers. Science. 314:268–274. 2006.
|
|
32
|
Gao J, Pfeifer D, He LJ, Qiao F, Zhang Z,
Arbman G, Wang ZL, Jia CR, Carstensen J and Sun XF: Association of
NFKBIA polymorphism with colorectal cancer risk and prognosis in
Swedish and Chinese populations. Scand J Gastroenterol. 42:345–350.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Osborne J, Lake A, Alexander FE, Taylor GM
and Jarrett RF: Germline mutations and polymorphisms in the NFKBIA
gene in Hodgkin lymphoma. Int J Cancer. 116:646–651. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He Y, Zhang H, Yin J, Xie J, Tan X, Liu S,
Zhang Q, Li C, Zhao J, Wang H and Cao G: IkappaBalpha gene promoter
polymorphisms are associated with hepatocarcinogenesis in patients
infected with hepatitis B virus genotype C. Carcinogenesis.
30:1916–1922. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bu H, Rosdahl I, Sun XF and Zhang H:
Importance of polymorphisms in NF-kappaB1 and NF-kappaBIalpha genes
for melanoma risk, clinicopathological features and tumor
progression in Swedish melanoma patients. J Cancer Res Clin Oncol.
133:859–866. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu X, Yu H, Yang W, Zhou X, Lu H and Shi
D: Mutations of NFKBIA in biopsy specimens from Hodgkin lymphoma.
Cancer Genet Cytogenet. 197:152–157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Parker KM, Ma MH, Manyak S, Altamirano CV,
Tang YM, Frantzen M, Mikail A, Roussos E, Sjak-Shie N, Vescio RA
and Berenson JR: Identification of polymorphisms of the
IkappaBalpha gene associated with an increased risk of multiple
myeloma. Cancer Genet Cytogenet. 137:43–48. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tanaka K, Kawakami T, Tateishi K,
Yashiroda H and Chiba T: Control of IkappaBalpha proteolysis by the
ubiquitin-proteasome pathway. Biochimie. 83:351–356. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
White KL, Vierkant RA, Phelan CM, Fridley
BL, Anderson S, Knutson KL, Schildkraut JM, Cunningham JM, Kelemen
LE, Pankratz VS, Rider DN, Liebow M, Hartmann LC, Sellers TA and
Goode EL: Polymorphisms in NF-kappaB inhibitors and risk of
epithelial ovarian cancer. BMC Cancer. 9:1702009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Obata H, Biro S, Arima N, Kaieda H, Kihara
T, Eto H, Miyata M and Tanaka H: NF-kappa B is induced in the
nuclei of cultured rat aortic smooth muscle cells by stimulation of
various growth factors. Biochem Bioph Res Commun. 224:27–32. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Habib AA, Högnason T, Ren J, Stefánsson K
and Ratan RR: The epidermal growth factor receptor associates with
and recruits phosphatidylinositol 3-kinase to the platelet-derived
growth factor beta receptor. J Biol Chem. 273:6885–6891. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sun L and Carpenter G: Epidermal growth
factor activation of NF-kappaB is mediated through IkappaBalpha
degradation and intracellular free calcium. Oncogene. 16:2095–2102.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Biswas DK, Cruz AP, Gansberger E and
Pardee AB: Epidermal growth factor-induced nuclear factor kappa B
activation: A major pathway of cell-cycle progression in
estrogen-receptor negative breast cancer cells. Proc Natl Acad Sci
USA. 97:8542–8547. 2000. View Article : Google Scholar
|
|
44
|
Haussler U, von Wichert G, Schmid RM,
Keller F and Schneider G: Epidermal growth factor activates nuclear
factor-kappaB in human proximal tubule cells. Am J Physiol Renal
Physiol. 289:F808–F815. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sethi G, Ahn KS, Chaturvedi MM and
Aggarwal BB: Epidermal growth factor (EGF) activates nuclear
factor-kappaB through IkappaBalpha kinase-independent but EGF
receptor-kinase dependent tyrosine 42 phosphorylation of
IkappaBalpha. Oncogene. 26:7324–7332. 2007. View Article : Google Scholar
|
|
46
|
Beg AA and Baldwin AS Jr: The I kappa B
proteins: multifunctional regulators of Rel/NF-kappa B
transcription factors. Genes Dev. 7:2064–2070. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Brown K, Park S, Kanno T, Franzoso G and
Siebenlist U: Mutual regulation of the transcriptional activator
NF-kappa B and its inhibitor, I kappa B-alpha. Proc Natl Acad Sci
USA. 90:2532–2536. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Henkel T, Machleidt T, Alkalay I, Kronke
M, Ben-Neriah Y and Baeuerle PA: Rapid proteolysis of I kappa
B-alpha is necessary for activation of transcription factor
NF-kappa B. Nature. 365:182–185. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mellits KH, Hay RT and Goodbourn S:
Proteolytic degradation of MAD3 (I kappa B alpha) and enhanced
processing of the NF-kappa B precursor p105 are obligatory steps in
the activation of NF-kappa B. Nucleic Acids Res. 21:5059–5066.
1993. View Article : Google Scholar : PubMed/NCBI
|