Introduction
Bronchopulmonary dysplasia (BPD) was first reported
and defined by Northway et al in 1967 as the most common
form of chronic lung damage in premature infants, which includes
barotrauma, volutrauma and oxygen toxicity (1). It is characterized by arrested lung
growth, with decreased alveolarization and a dysmorphic vasculature
(2). It is the cause of prolonged
hospitalization with serious social and economic consequences. A
deeper understanding of the disease, such as future preventative
measures, aiming at reducing its incidence, minimizing
complications, diminishing hospital costs and improving infant
health has become the prime objective of public health (3).
Although its exact etiology and pathogenesis have
yet to be fully ascertained, several researches suggest that it
results from the complex interplay between impairments in the
premature lung, perinatal insults and resulting from the supportive
care of the infant (from mechanical ventilation and supplemental
oxygen administration) (4). In
other words, the strongest association is with pre-term birth,
although other variables, such as pre-natal and post-natal
infection, inflammation, mechanical ventilation, oxygen toxicity,
patent ductus arteriosus (PDA) also contribute to the pathogenesis
of BPD, whereas anti-angiogenesis is known to contribute
significantly to the disruption of lung development in animal
models (5). An increasing number
of studies has demonstrated impaired angiogenesis in the
development of preeclampsia (6).
Few therapies are known to effectively prevent or
treat BPD and corticosteroid therapy may decrease lung injury
through a variety of mechanisms, such as decreasing the
inflammatory response (7).
However, this type of treatment has been found to cause serious
short-term and long-term side-effects, such as gastrointestinal
bleeding, gastrointestinal perforation, hyperglycemia and
hypertension that give clinicians reason to severely limit the use
of this type of treatment (8).
The survival rate of premature neonatals has significantly
increased; however, the morbidity of BPD also shows an increasing
trend (9).
Epidermal growth factor-like domain 7 (EGFL7) is a
protein secreted from endothelial cells which plays an important
role in vascular tubulogenesis. It has been found that EGFL7 gene
expression is significantly decreased in neonatal rat lungs
following exposure to hyperoxic conditions and is important for
cell survival (10). It has been
identified as a potential therapeutic target for lung injury. Thus,
antioxidant therapy has been considered as a potential preventive
or treatment option for BPD (11–13).
Radix Astragali, a type of Chinese
traditional herb, has been used in Traditional Chinese Medicine for
over 20 centuries to strengthen the body against disease. It is
officially listed in both the Chinese and Japanese Pharmacopoeia
(14). Large numbers of
pharmacological and clinical studies have demonstrated that
Radix Astragali possesses a wide spectrum of activities,
such as immunomodulatory, antioxidant and anti-inflammatory,
cardioprotective, hepatoprotective, antihyperglycemic and antitumor
activities (15–23). It is well known that the principle
active constituents of Astragalus are polysaccharides,
saponins and isoflavonoids (24).
Several research groups have isolated and purified polysaccharides
from Astragalus termed Astragalus polysaccharides
(APS) or Astragalans (25). APS
from Radix Astragali, have attracted much attention due to
their outstanding antioxidant and anti-inflammatory effects
(26). APS have also proven to
have strong immunoregulatory properties (27).
However, the effects of APS on the mRNA expression
of EGFL7 in newborn rats with hyperoxia-induced BPD are unknown.
Thus, the aim of the present study was to determine the potent
effects of APS using a rat model of BPD and to clarify the
association between BPD and the expression of EGFL7.
Materials and methods
Animal experiments
A total of 96 Sprague Dawley newborn rats (weighing
7.82±0.63 g) and fungible mother rats were obtained from the
Experimental Animal Center of Southern Medical University
(Guangzhou, China). The present study was approved by the Ethical
and Research Committee of Southern Medical University. All
investigations were conducted according to the Guide for the Care
and Use of Laboratory Animals of the National Institutes of Health.
These rats were randomly divided into 4 groups of 24 rats in each
group (n=8 for each time point) as follows: the control group, air
room (RA) plus APS group, BPD group and the APS group (20
mg/kg/day). BPD was induced by exposing the rats to hyperoxic
conditions. Ten hours after birth, the pups in the control group
and RA plus APS group were kept in room air containing 21%
O2. The rats in the RA plus APS group received daily
injections of APS (Sainuo Pharmaceutical Co., Ltd., Tianjin, China)
[intraperitoneally (i.p.) 20 mg/kg/day] throughout the post-natal
14 days, while those in the BPD and APS groups were placed in an
oxygen chamber, into which oxygen was continuously delivered
(FiO2, 0.85±0.03) and received daily injections of
saline (i.p.) and APS (i.p. 20 mg/kg) throughout the post-natal 14
days, respectively. Temperature and humidity were maintained at
22–25°C and 60–70%, respectively. The chamber was opened for 1 h
daily to switch dams between air and the O2 environment
to protect the dams from oxygen toxicity. These pups were then
sacrificed on days 4, 10 and 14 after the experiments were
completed and the lung tissues were collected.
Assessment of lung histological
damage
Following anesthesia with pentobarbital (60 mg/kg,
i.p.), the lungs of the pups were fixed with an intratracheal
injection of 4% paraformaldehyde and post-fixed overnight at room
temperature. The tissues were paraffin-embedded, sectioned to 5 μm
thickness and stained with hematoxylin and eosin (H&E). A
quantitative analysis of the pulmonary mean linear intercept (MLI),
the mean alveolar number (MAN) was carried out according to
previously described methods (28).
Immunohistochemistry
Lung tissue was obtained at each time point and
post-fixed overnight at room temperature. The tissues were then
paraffin-embedded, sectioned to 5 μm thickness and deparaffinized
in xylene and hydrated in a series of graded alcohol. After
dewaxing and rehydration, the sections were immersed in 3% hydrogen
peroxide in methanol for 20 min at room temperature to abolish
endogenous peroxidase activity and then antigen retrieval was
carried out using a microwave for 15 min before blocking with 5%
bovine serum albumin (Life Technologies Co., Carlsbad, CA, USA) at
37°C for 20 min. The sections were incubated with polyclonal CD31
antibody (diluted to 1/200; Novus Biologicals, Littleton, CO, USA)
at 37°C for 2 h. After washing with PBS, the sections were
incubated with a biotinylated peroxidase-conjugated secondary
antibody and 0.1% DAB substrate, using the standard
streptavidin-biotin-based method. A negative control was prepared
by reacting a few sections with normal mouse IgG (Abcam
Biotechnology, Cambridge, UK) at the same dilution instead of the
specific antibody. A cytoplasmic brown granule was marked as a
positive expression of CD31. We quantified vascular density by
measuring the area of CD31 immunostaining relative to the total
area of parenchymal cells using Image-Pro Plus 6.0 (Media
Cybernetics, Rockville, MD, USA) of differential interference
contrast images. The tissue sections analyzed contained mainly the
alveolar parenchyma and excluded any large airways or blood
vessels.
Reverse transcription quantitative
PCR
Total RNA from total lung tissue was isolated using
the TRIzol kit (Takara Bio, Dalian, China) following the
manufacturer’s instructions. Reverse transcription (RT) was
performed using the PrimeScript RT reagent kit (Takara Bio).
Real-time (quantitative) PCR was conducted using an Applied
Biosystems 7500 Real Time PCR System and the relative
quantification of mRNA expression was calculated using the 2-ΔΔCt
method, as previously described (29). The SYBR Premix Ex Taq (Tli RNaseH
Plus) kit was purchased from Takara Bio. This was followd by
priming with Oligo(dT) and subsequent amplification using specific
oligonucleotide primers based on the rat corresponding gene
sequence (Table I); β-actin
served as a housekeeping gene. The primers were synthesized by
Shanghai Invitrogen Biotechnology Co., Ltd. (Shanghai, China). Two
microlitres of cDNA were amplified in 20 μl of PCR solution. The
thermal cycling parameters consisted of 95°C for 30 sec, followed
by 40 cycles of 95°C for 5 sec and 60°C for 34 sec.
 | Table IList of oligonucleotides used as
primers in the quantitative PCR analysis of gene expression in lung
tissue. |
Table I
List of oligonucleotides used as
primers in the quantitative PCR analysis of gene expression in lung
tissue.
| Gene symbol | Primer | Primer sequence |
|---|
| Bax | Forward |
5′-AGAGGATGGCTGGGGAGAC-3′ |
| Reverse |
5′-CGCTCAGCTTCTTGGTGGAT-3′ |
| Bcl-2 | Forward |
5′-ACCCCTGGCATCTTCTCCT-3′ |
| Reverse |
5′-CGACGGTAGCGACGAGAG-3′ |
| EGFL7 | Forward |
5′-CCGAACCATCTACCGGACTG-3′ |
| Reverse |
5′-GCCTGTCTGTCACCCATTCA-3′ |
| β-actin | Forward |
5′-AGGGAAATCGTGCGTGACAT-3′ |
| Reverse |
5′-GAACCGCTCATTGCCGATAG-3′ |
Western blot analysis
Protein was extracted from the lung tissue of the
pups in the different groups with lysis buffer using the Total
Protein Extraction Reagent kit (Nanjing KeyGen Biotech Co., Ltd.,
Nanjing, China). The protein concentration of each sample was
measured using the KeyGen BCA Protein assay (Nanjing KeyGen
Biotech). Protein extract samples (30 μg/lane) were analyzed by 12%
SDS-PAGE and transferred onto PVDF membranes. The membranes were
blocked in 5% non-fat milk in TBS + 0.1% Tween-20, and incubated
with EGFL7 (Proteintech Group, Inc., Chicago, IL, USA), Bax
(Epitomics-an Abcam Co., Burlingame, CA, USA) and Bcl-2 (Bioworld
Technology, Inc., St. Louis Park, MN, USA) polyclonal antibody, and
diluted at 1/500; anti-β-actin (ZSGB-Bio, Beijing, China) was used
as the cytoplasmic endogenous control diluted at 1/2,000.
Anti-rabbit (H+L) HRP and anti-mouse (H+L) HRP diluted at 1:2,000
was used as the secondary antibody with SuperSignal West Pico
Chemiluminescent Substrate (Thermo Fisher Scientific Inc.,
Rockford, IL, USA) for detection. The membrane was further
incubated for 60 min at room temperature. The intensities of the
protein bands were analyzed using Quantity One software (Bio-Rad
Laboratories, Hercules, CA, USA).
Statistical analysis
The results are presented as the means ± SD.
Statistical analysis was performed using SPSS 13.0 statistical
software. One way analysis of variance and the Student-Newman-Keuls
test were used for data analysis. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
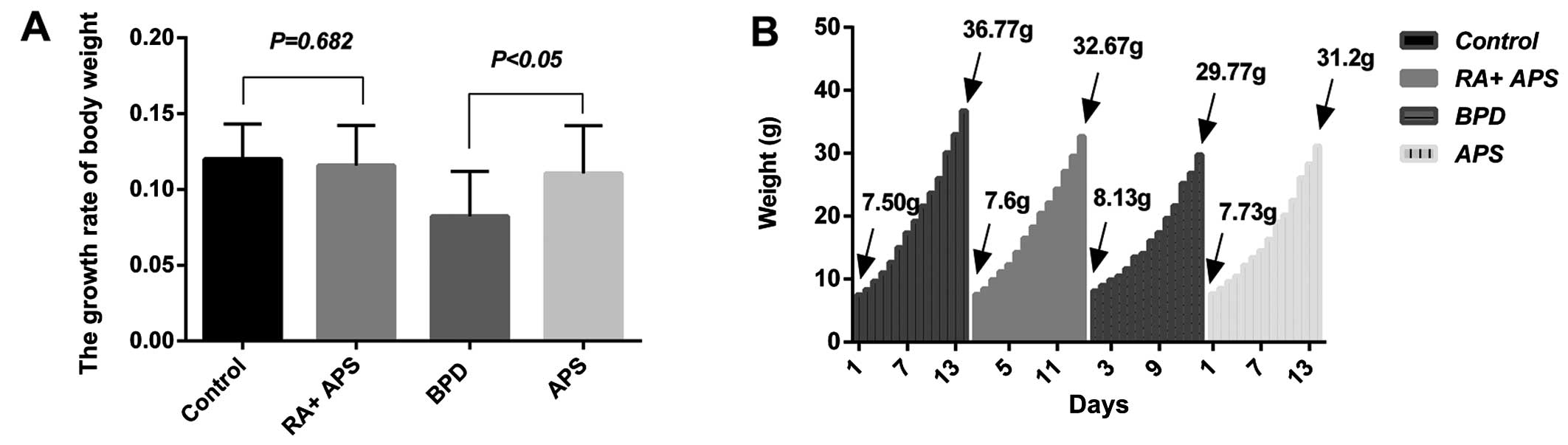
Body weight
The body weight of the hyperoxic pups was slightly
lower than that of the normoxic (control and RA plus APS groups)
pups; the body weight of the rats in the APS group was slightly
lower than that of the rats in the control and RA plus APS groups,
although higher than that of the rats in the BPD group after 14
days of exposure. The growth rate of body weight in the control
group and RA plus APS group was slightly higher than that of the
APS group and BPD group. The growth rate of the APS group was
significantly higher than that of the BPD group
(*P<0.05) and there were no statistically significant
difference among the APS group, RA plus APS group and the control
group (Fig. 1).
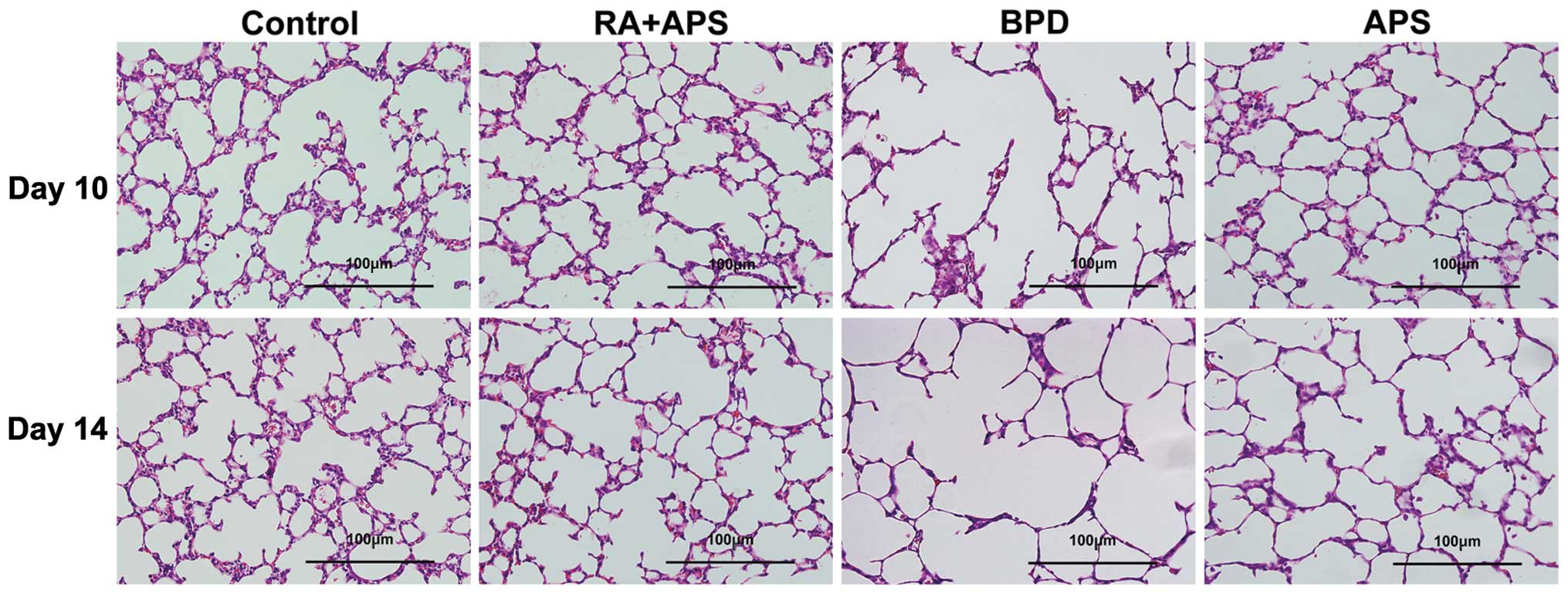
Histological evaluation
As shown in Fig.
2, no significant histological damage was observed in the
control group and RA plus APS group. In the pups with
hyperoxia-induced BPD, a number of inflammatory cella infiltrating
the interstitial lung was observed, and there were fewer and larger
simplified alveoli, variable interstitial fibroproliferation, and
fewer and dysmorphic capillaries. Treatment with APS significantly
attenuated the extent and severity of the histological signs and
evidently prevented the development of lung damage compared to the
BPD group. As compared with the BPD group, the MLI was
significantly low and the MAN per square area was significantly
high in the APS group (P<0.05 or P<0.01) (Table II).
| Table IIThe mean linear intercept value and
the number of alveolar per square area in each group. |
Table II
The mean linear intercept value and
the number of alveolar per square area in each group.
| MLI (μm) | MAN
(/104 μm2) |
|---|
|
|
|---|
| Group | Day 10 | Day 14 | Day 10 | Day 14 |
|---|
| Control | 32.13±2.84a | 33.26±0.74a | 34.03±2.83b | 37.65±4.12b |
| RA + APS | 31.96±3.55 | 32.04±2.32 | 33.60±1.42 | 36.39±1.53 |
| BPD | 38.43±1.01 | 38.83±1.14 | 22.45±1.82 | 25.00±2.19 |
| APS | 34.21±0.98b | 35.31±1.37a | 27.33±1.93a | 34.59±0.89b |
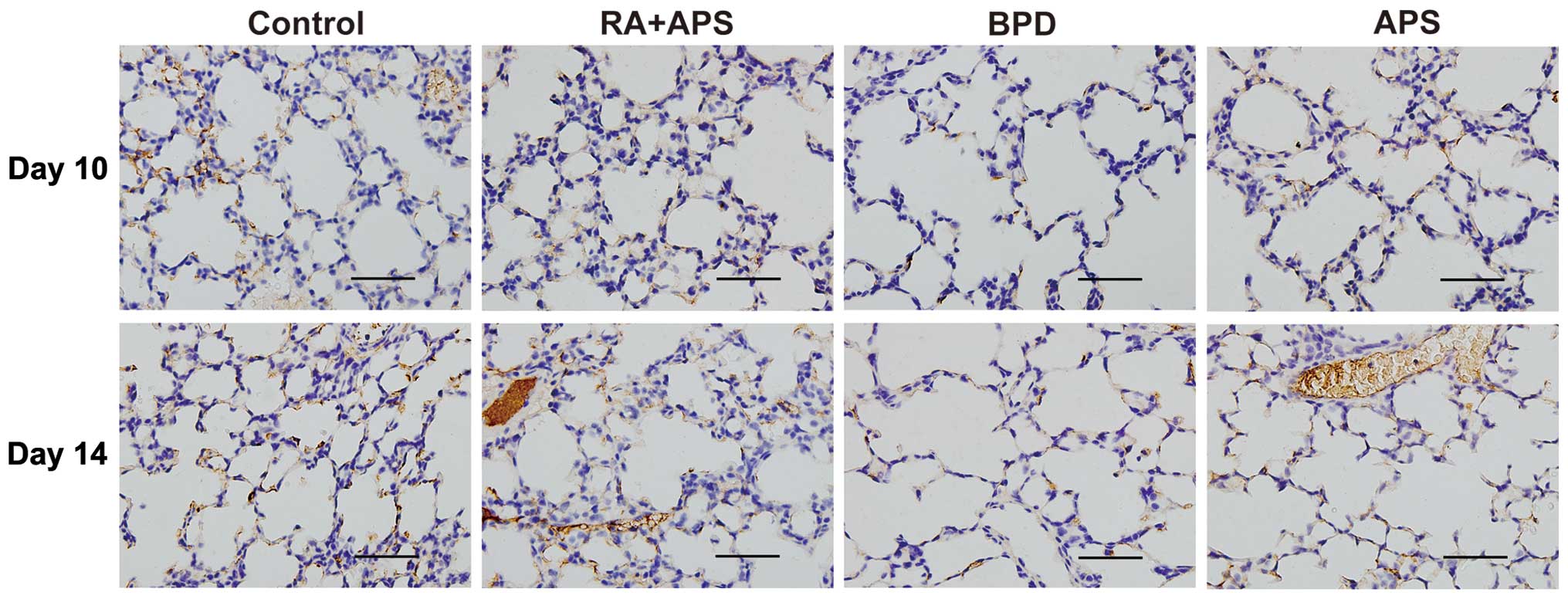
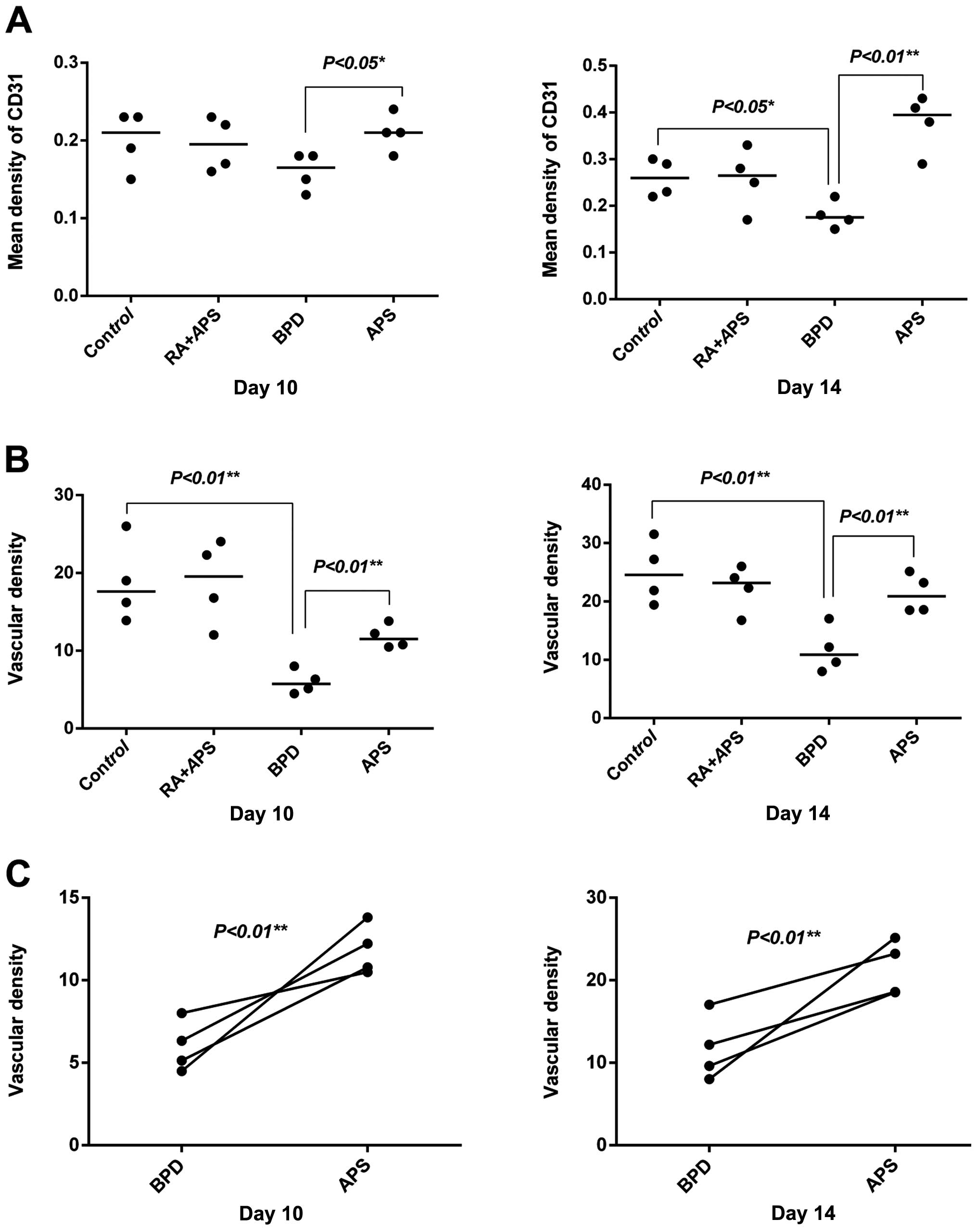
Detection of of CD31 protein expression
and vascular density by immunohistochemistry
The cells positive for CD31 show a brown-yellow
cytoplasm and the negative control shows an absence of staining.
CD31 protein was mainly expressed in the cytoplasm of the pulmonary
microvascular endothelial cells. Both CD31 and vascular density in
the BPD group were decreased as compared with the control group.
The expression of CD31 and vascular density in the APS group was
significantly increased compared with the BPD group at each time
point (P<0.05 or P<0.01) (Figs.
3 and 4).
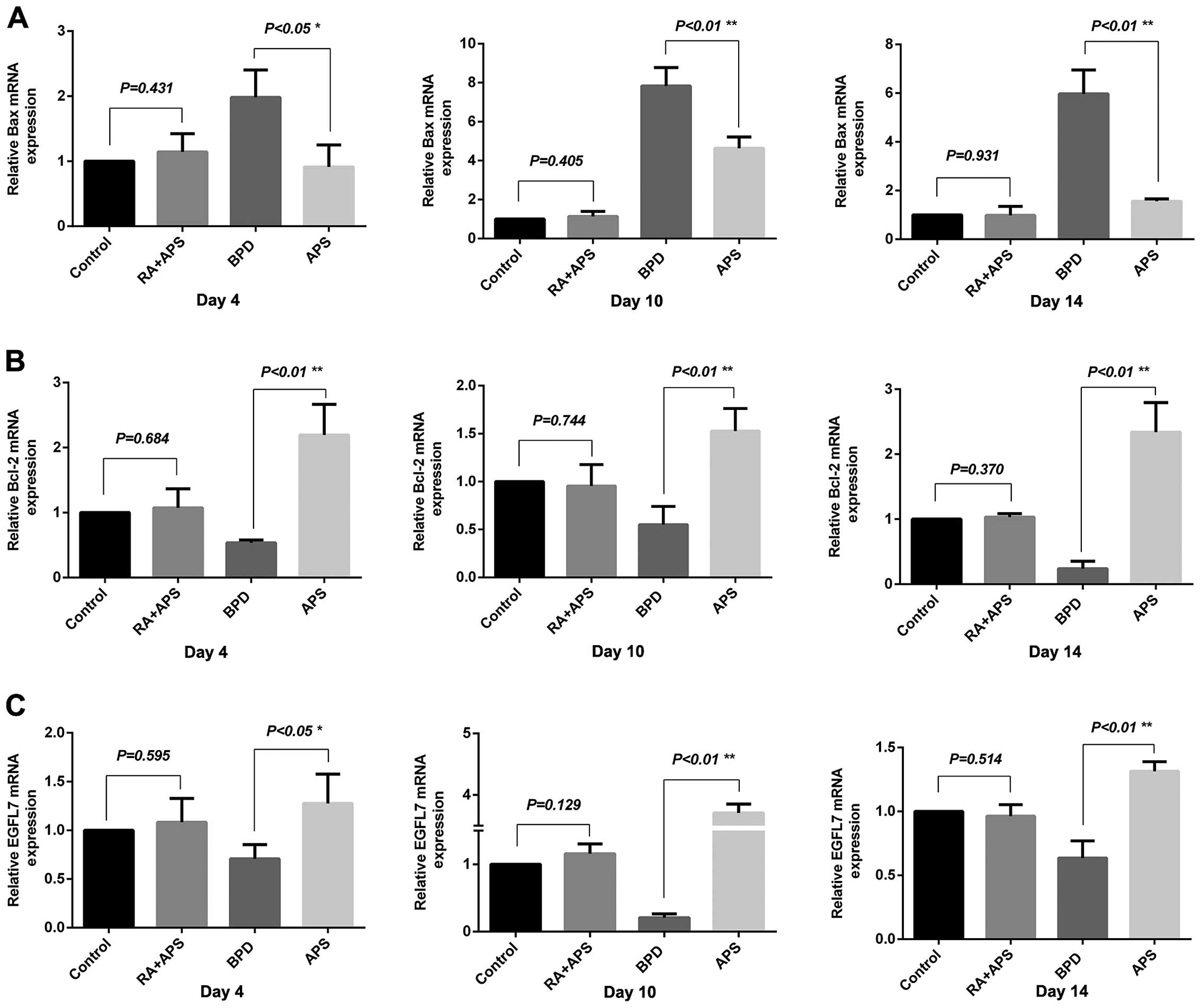
Changes in the mRNA expression of Bax,
Bcl-2 and EGFL7 detected by quantitative (real-time) PCR
The mRNA expression of EGFL7 and Bcl-2 in the rats
in the BPD group was significantly low, while the Bax mRNA level
was significantly high compared with thecontrol group at each time
point (P<0.01). In the APS group, the expression of EGFL7 and
Bcl-2 was significantly increased and that of Bax was significantly
decreased compared with the BPD group on days 4, 10 and 14
(P<0.05 or P<0.01). There was no significant difference
observed between the RA plus APS group and the control group
(Fig. 5).
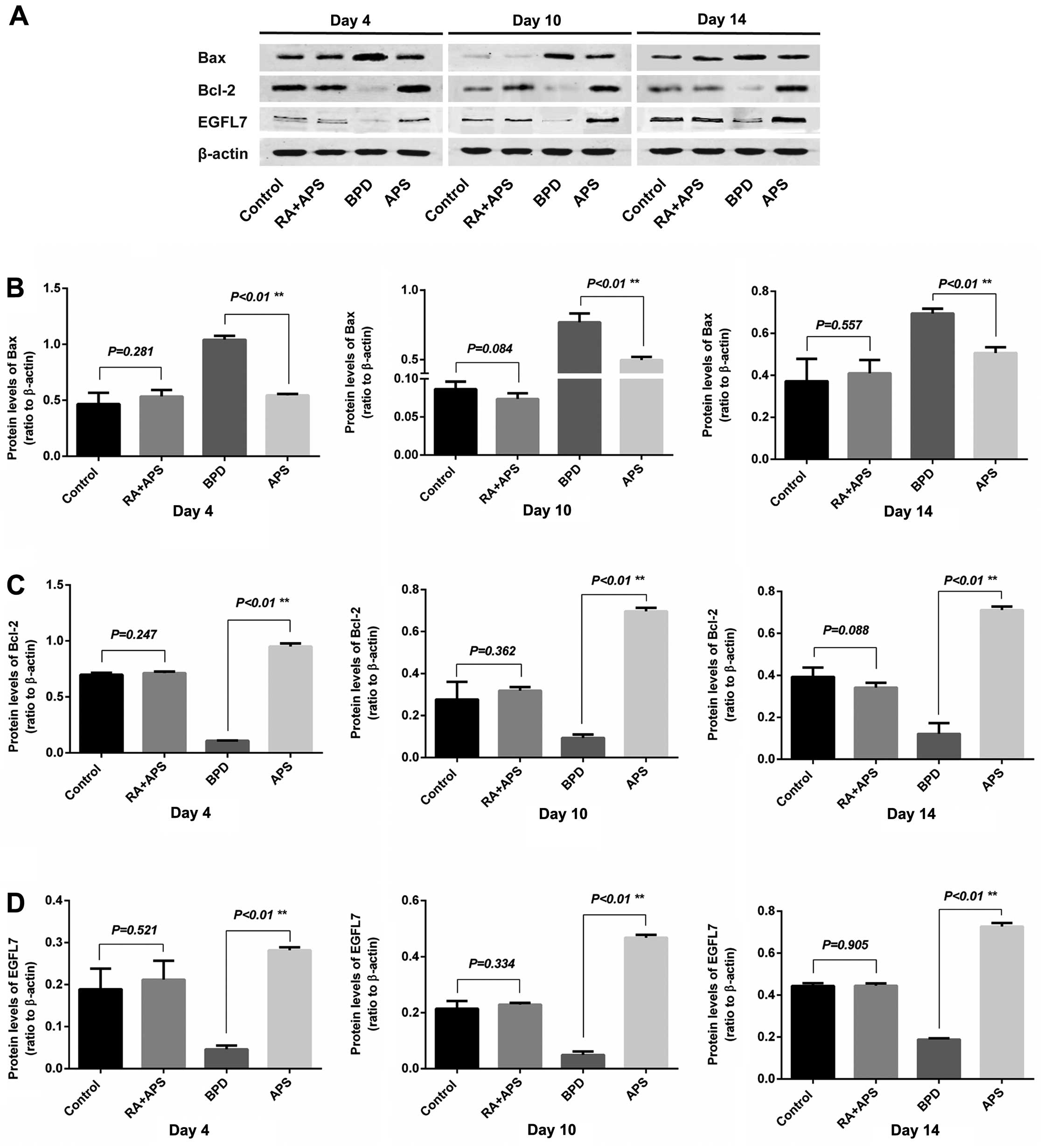
Detection of the Bax, Bcl-2 and EGFL7
protein expression in lung tissue by western blot analysis
The protein levels of Bax, Bcl-2 and EGFL7 in the
lung tissue the pups in each group were examined on days 4, 10 and
14. The results from western blot analsyis revealed that the
protein expression of Bcl-2 and EGFL7 in the BPD group was
decreased, while the protein expression of Bax was increased
compared with the control group on days 4, 10 and 14 (P<0.01).
In the APS group, the protein expression of Bcl-2 and EGFL7 was
significantly increased, while that of Bax was significantly
decreased compared with the BPD group. The protein expression of
Bax, Bcl-2 and EGFL7 in the RA plus APS groups did not differ
significantly from that of control group at any time point
(Fig. 6).
Discussion
Bronchopulmonary dysplasia (BPD) is the product of a
heterogeneous group of lung disorders that begin in the neonatal
period. The overall incidence of BPD has not changed over the past
decades. What is more, it remains a challenge for the future
(3).
There is increasing evidence indicating that
oxidative stress plays a key role in the development of BPD. In
this study, we found serious damage to the lung tissue in the BPD
model group. The balance of the production of reactive oxygen
species (ROS) and the antioxidant defenses disturbed by exposrue to
high oxygen concentrations, leads to biochemical and histological
effects being observed in the lung tissue. Furthermore, hyperoxia
has the ability to modify cellular macromolecules, thereby
promoting cell death (30).
Previous studies have demonstrated that the
disruption of normal lung vascular growth plays a vital role in the
pathogenesis of BPD (31). CD31,
a marker for endothelial cells, is decreased in lung tissues in BPD
(32). However, in the present
study, the damage to the lung tissue of newborn rats in the APS
group was mild, with vascular development, compared with the BPD
model group. CD31 expression in lung tissue increased by almost
2-fold in the APS group compared to the BPD group, and vascular
density also significantly improved in the APS group compared with
the BPD group. This indicated that APS markedly improved pulmonary
injury in the newborn rats with BPD by reducing oxidative damage.
The antioxidant effects of Astragalus in brain and kidney
tissue following ischemia-reperfusion injury have also been
demonstrated (33).
In the present study, both the mRNA and protein
levels of EGFL7 in the BPD model group were significantly decreased
compared with the control, as shown by RT-qPCR and western bolt
analysis. However, the mRNA and protein expression of EGFL7 was
significantly increased in the APS intervention group compared with
the BPD model group. It is known that EGFL7 is a secreted protein
specifically expressed by endothelial cells and is highly expressed
in the lungs, heart, kidneys, spleen and uterus (34,35), and is an important tubulogenic
factor in the process of vasculogenesis. Xu et al (10) proposed that the
endothelial-specific growth factor, EGFL7, may play a role in
hyperoxia-induced vascular injury. The knockdown of the gene in
zebrafish has been shown to result in a severe impairment of
arterial and venous endothelial cell cord segregation, leading to
the formation of midline angioblast aggregates (36,37). The overexpression of EGFL7 reduces
the expression of the pro-apoptotic protein, Bax, and increases the
expression of the anti-apoptotic protein, Bcl-2, which prevents
hyperoxia-induced endothelial cell death and promotes lung vascular
development (10).
Hockenbery et al (39) found that the overexpression of
Bcl-2 completely suppressed lipid peroxidation. In other words,
Bcl-2 functions in an antioxidant pathway to prevent apoptosis. It
is known that Bax, a pro-apoptotic member of the Bcl-2 family of
proteins, promotes apoptosis by binding Bcl-2 and inhibiting its
anti-apoptotic function during mitochondria-regulated programmed
cell death (40). Previous
studies have demonstrated that the exposure of newborn mice to
hyperoxia increased the mRNA expression pro-apoptotic Bax and tge
number of apoptotic lung cells (41). In this study, we found that the
expression of Bcl-2 was significantly decreased in the BPD group
and was significantly increased in the APS group both at the mRNA
and protein level, while the expression of Bax showed an opposite
trend. This suggests that APS exert protective effects against BPD.
The protective effect of APS against hyperoxia-induced lung injury
may occur through the upregulation EGFL7 expression. Emerging
evidence links the pathophysiology of BPD to an imbalance between
anti-apoptotic and pro-apoptotic signaling pathways and an arrest
in alveolar and vascular development (42).
The present study demonstrates that APS upregulate
the expression of EGFL7 and Bcl-2 and downregulates Bax expression
and significantly reduces alveolar damage, exerting protective
effects on lung tissue in BPD, which was closely related to the
inhibition of the endothelial cell apoptotic pathway. Our results
demonstrated that APS exerted beneficial effects (antioxidant and
anti-inflammatory effects) in the rats with hyperoxia-induced BPD.
The exposure of newborn rats to a high oxygen environment caused
lung injury which was very similar to human BPD. We speculate that
the upregulation of the expression of EGFL7 may be the target of
potential drugs for BPD. Thus, APS may have potential for use as a
therapeutic strategy for BPD in neonates.
Acknowledgements
The present study was supported by the Guangdong
Province Science and Technology Plan Project (2012B031800303 to
W.-M.H.).
References
|
1
|
Northway WH Jr, Rosan RC and Porter DY:
Pulmonary disease following respirator therapy of hyaline-membrane
disease. Bronchopulmonary dysplasia. N Engl J Med. 276:357–368.
1967. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jobe AH: The new bronchopulmonary
dysplasia. Curr Opin Pediatr. 23:167–172. 2011. View Article : Google Scholar
|
|
3
|
Gien J and Kinsella JP: Pathogenesis and
treatment of bronchopulmonary dysplasia. Curr Opin Pediatr.
23:305–313. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hayes D Jr, Feola DJ, Murphy BS, Shook LA
and Ballard HO: Pathogenesis of bronchopulmonary dysplasia.
Respiration. 79:425–436. 2010. View Article : Google Scholar
|
|
5
|
Tang JR, Markham NE, Lin YJ, et al:
Inhaled nitric oxide attenuates pulmonary hypertension and improves
lung growth in infant rats after neonatal treatment with a VEGF
receptor inhibitor. Am J Physiol Lung Cell Mol Physiol.
287:L344–L351. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Foidart JM, Schaaps JP, Chantraine F, et
al: Dysregulation of anti-angiogenic agents (sFlt-1, PLGF, and
sEndoglin) in preeclampsia: a step forward but not the definitive
answer. J Reprod Immunol. 82:106–111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Speer CP: Chorioamnionitis, postnatal
factors and proinflammatory response in the pathogenetic sequence
of bronchopulmonary dysplasia. Neonatology. 95:353–361. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Soll RF: Corticosteroids for the treatment
and prevention of bronchopulmonary dysplasia. Neonatology.
98:109–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jensen EA and Schmidt B: Epidemiology of
bronchopulmonary dysplasia. Birth Defects Res A Clin Mol Teratol.
100:145–157. 2014. View Article : Google Scholar
|
|
10
|
Xu D, Perez RE, Ekekezie II, Navarro A and
Truog WE: Epidermal growth factor-like domain 7 protects
endothelial cells from hyperoxia-induced cell death. Am J Physiol
Lung Cell Mol Physiol. 294:L17–L23. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Asikainen TM and White CW: Pulmonary
antioxidant defenses in the preterm newborn with respiratory
distress and bronchopulmonary dysplasia in evolution: implications
for antioxidant therapy. Antioxid Redox Signal. 6:155–167. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Welty SE: Antioxidants and oxidations in
bronchopulmonary dysplasia: there are no easy answers. J Pediatr.
143:697–698. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Welty SE and Smith CV: Rationale for
antioxidant therapy in premature infants to prevent
bronchopulmonary dysplasia. Nutr Rev. 59:10–17. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chu Chu, Qi Lian-Wen, Liu E-Hu, Li Bin,
Gao Wen and Li Ping: Radix Astragali (Astragalus):
latest advancements and trends in chemistry, analysis, pharmacology
and pharmacokinetics. Curr Org Chem. 14:1792–1807. 2010. View Article : Google Scholar
|
|
15
|
Yang ZG, Sun HX and Fang WH: Haemolytic
activities and adjuvant effect of Astragalus membranaceus
saponins (AMS) on the immune responses to ovalbumin in mice.
Vaccine. 23:5196–5203. 2005.PubMed/NCBI
|
|
16
|
Xu XL, Ji H, Gu SY, Shao Q, Huang QJ and
Cheng YP: Cardioprotective effects of Astragali Radix
against isoproterenol-induced myocardial injury in rats and its
possible mechanism. Phytother Res. 22:389–394. 2008.
|
|
17
|
Ohkawara S, Okuma Y, Uehara T, Yamagishi T
and Nomura Y: Astrapterocarpan isolated from Astragalus
membranaceus inhibits proliferation of vascular smooth muscle
cells. Eur J Pharmacol. 525:41–47. 2005.
|
|
18
|
Yan F, Zhang QY, Jiao L, Han T, Zhang H,
Qin LP and Khalid R: Synergistic hepatoprotective effect of
Schisandrae lignans with Astragalus polysaccharides
on chronic liver injury in rats. Phytomedicine. 16:805–813.
2009.
|
|
19
|
Wang S, Li J, Huang H, Gao W, Zhuang C, Li
B, Zhou P and Kong D: Anti-hepatitis B virus activities of
astragaloside IV isolated from radix Astragali. Biol Pharm
Bull. 32:132–135. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mao XQ, Yu F, Wang N, Wu Y, Zou F, Wu K,
Liu M and Ouyang JP: Hypoglycemic effect of polysaccharide enriched
extract of Astragalus membranaceus in diet induced insulin
resistant C57BL/6J mice and its potential mechanism. Phytomedicine.
16:416–425. 2009.PubMed/NCBI
|
|
21
|
Auyeung KK, Cho CH and Ko JK: A novel
anticancer effect of Astragalus saponins: transcriptional
activation of NSAID-activated gene. Int J Cancer. 125:1082–1091.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu DH, Bao YM, Wei CL and An LJ: Studies
of chemical constituents and their antioxidant activities from
Astragalus mongholicus Bunge. Biomed Environ Sci.
18:297–301. 2005.PubMed/NCBI
|
|
23
|
Choi SI, Heo TR, Min BH, Cui JH, Choi BH
and Park SR: Alleviation of osteoarthritis by
calycosin-7-O-beta-D-glucopyranoside (CG) isolated from
Astragali radix (AR) in rabbit osteoarthritis (OA) model.
Osteoarthritis Cartilage. 15:1086–1092. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song JZ, Yiu HH, Qiao CF, Han QB and Xu
HX: Chemical comparison and classification of Radix Astragali by
determination of isoflavonoids and astragalosides. J Pharm Biomed
Anal. 47:399–406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu DJ, Xia Q, Wang JJ and Wang PP:
Molecular weight and monosaccharide composition of Astragalus
polysaccharides. Molecules. 13:2408–2415. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang WM, Liang YQ and Wang XH:
Antioxidant and anti-inflammatory effects of Astragalus
polysaccharide on EA.hy926 cells. Exp Ther Med. 36:199–203. 201
|
|
27
|
Yang B, Xiao B and Sun T: Antitumor and
immunomodulatory activity of Astragalus membranaceus
polysaccharides in H22 tumor-bearing mice. Int J Biol Macromol.
62:287–290. 2013.
|
|
28
|
Robbesom AA, Versteeg EM, Veerkamp JH, et
al: Morphological quantification of emphysema in small human lung
specimens: comparison of methods and relation with clinical data.
Mod Pathol. 16:1–7. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morcillo EJ, Estrela J and Cortijo J:
Oxidative stress and pulmonary inflammation: pharmacological
intervention with antioxidants. Pharmacol Res. 40:393–404. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
D’Angio CT and Maniscalco WM: The role of
vascular growth factors in hyperoxia-induced injury to the
developing lung. Front Biosci. 7:D1609–D1623. 2002.PubMed/NCBI
|
|
32
|
Coalson JJ, Winter VT, Siler-Khodr T and
Yoder BA: Neonatal chronic lung disease in extremely immature
baboons. Am J Respir Crit Care Med. 160:1333–1346. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Krasteva I, Nikolova I, Danchev N and
Nikolov S: Phytochemical analysis of ethyl acetate extract from
Astragalus corniculatus Bieb. and brain antihypoxic
activity. Acta Pharm. 54:151–156. 2004.
|
|
34
|
Bhatt AJ, Pryhuber GS, Huyck H, Watkins
RH, Metlay LA and Maniscalco WM: Disrupted pulmonary vasculature
and decreased vascular endothelial growth factor, Flt-1, and TIE-2
in human infants dying with bronchopulmonary dysplasia. Am J Respir
Crit Care Med. 164:1971–1980. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Soncin F, Mattot V, Lionneton F, Spruyt N,
Lepretre F, Begue A and Stehelin D: VE-statin, an endothelial
repressor of smooth muscle cell migration. EMBO J. 22:5700–5711.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Campagnolo L, Leahy A, Chitnis S,
Koschnick S, Fitch MJ, Fallon JT, Loskutoff D, Taubman MB and
Stuhlmann H: EGFL7 is a chemoattractant for endothelial cells and
is up-regulated in angiogenesis and arterial injury. Am J Pathol.
167:275–284. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
De Maziere A, Parker L, Van Dijk S, Ye W
and Klumperman J: Egfl7 knockdown causes defects in the extension
and junctional arrangements of endothelial cells during zebrafish
vasculogenesis. Dev Dyn. 237:580–591. 2008.PubMed/NCBI
|
|
38
|
Parker LH, Schmidt M, Jin SW, et al: The
endothelial-cell-derived secreted factor Egfl7 regulates
vascular tube formation. Nature. 428:754–758. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hockenbery DM, Oltvai ZN, Yin XM, Milliman
CL and Korsmeyer SJ: Bcl-2 functions in an antioxidant pathway to
prevent apoptosis. Cell. 75:241–251. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Oltval ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kroon AA and Post M: Apoptotic cell death
in bronchopulmonary dysplasia. Curr Pediatr Rev. 7:285–292. 2011.
View Article : Google Scholar
|
|
42
|
Schmidt M, Paes K, De Maziere A, et al:
EGFL7 regulates the collective migration of endothelial cells by
restricting their spatial distribution. Development. 134:2913–2923.
2007. View Article : Google Scholar : PubMed/NCBI
|