Introduction
Ischemic cerebrovascular disease is caused by
cerebral vascular stenosis or occlusion leading to cerebral
ischemia or thrombosis, which accounts for 70–80% of all
cerebrovascular disease. How to prevent secondary brain injury
following cerebral ischemia is currently of interest in clinical
and scientific research. Previous studies indicate that
angiogenesis is one of the key factors in the recovery of secondary
brain injury. Hypoxia-inducible factor 1α (HIF1α) plays an
important role in angiogenesis (1–4).
HIF1α promotes angiogenesis without causing blood vessel leakage,
tissue edema and inflammatory reaction (5), and is considered a promising
therapeutic target for clinical treatments.
HIF1α is an important transcription factor and a
functional subunit. Its protein stability and transcriptional
activity are mainly regulated by the intracellular oxygen
concentration (6,7). HIF1α facilitates cell survival under
hypoxic conditions by binding to the hypoxia response element in
the target gene and regulates the expression of broad spectrum of
angiogenesis-related genes, including vascular endothelial growth
factor (VEGF). As a result, VEGF promotes the angiogenesis and
revascularization, and improves the blood supply (8–10).
However, HIF1α is only expressed under hypoxia conditions and
degraded rapidly under normoxic conditions due to the hydroxylation
of the 564-proline and 803-asparagine in the coding sequences
(CDS). Hydroxylation of the 564-proline causes HIF1α degradation by
the oxygen-dependent degradation domain (ODDD) (11–14). The 803-asparagine in the
C-terminal region (COOH-terminal transactivation domain, CAD)
determines the HIF1α transcriptional activity. The prevention of
803-asparagine hydroxylation with chemical inhibitors or genetic
recombination can induce strong transcription activity of HIF1α
(15).
Thus far, there are few studies that focus on the
treatment of cerebral ischemia using mutant HIF1α-transfected
BMSCs. In the present study, the 564-proline and 803-asparagine in
the CDS were simultaneously mutated into alanine and the mutant
HIF1α recombinant adenovirus vector was constructed. The
HIF1α gene recombinant vector was transfected into BMSCs
(16), and subsequently the
infected BMSCs were transplanted to the middle cerebral artery
occlusion model (MCAO) to achieve high levels of HIF1α protein
expression and promote revascularization in the ischemic area.
Materials and methods
Animals
Male Sprague Dawley (SD) rats (280–320 g, 6–8 weeks
old) were provided by the Experimental Animal Center of Liaoning
Medical College (Jinzhou, Liaoning, China). All the experimental
procedures were carried out according to the internationally
accredited guidelines with the approval of the Institutional Animal
Care and Use Committee of Drug Safety Evaluation Center of Liaoning
Medical College.
Reagents
The recombinant adenovirus vector containing
wild-type HIF1α or mutant HIF1α was kindly given by Danping Liu
(Liaoning Medical College). Lipofectamine 2000 and Opti-MEM were
purchased from Invitrogen Life Technologies (Carlsbad, CA, USA);
pMD19-T and reverse transcription-polymerase chain reaction
(RT-PCR) kit were purchased from Takara Bio, Inc. (Otsu, Japan);
PmeI and PacI were purchased from New England
Biolabs, Inc. (Ipswich, MA, USA); Dulbecco’s modified Eagle’s
medium (DMEM), trypsin and fetal bovine serum (FBS) were purchased
from Gibco Life Technologies (Carlsbad, CA, USA); anti-HIF1α
(rabbit; no. 10790), anti-VEGF (rabbit; no. 507), anti-cluster of
differentiation 105 (CD105) (rabbit; no. 20632), anti-β-actin (no.
130656) and the corresponding secondary antibodies [horseradish
peroxidase (HRP)-conjugated; no. 2004] were purchased from Santa
Cruz Biotechnology, Inc., (Santa Cruz, CA, USA);
2,3,5-triphenyltetrazolium chloride was purchased from Sigma (St.
Louis, MO, USA); and the HEK293A cells were purchased from the Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China).
Infection of BMSCs with recombinant
adenovirus
The recombinant adenovirus vector was introduced
into the HEK293A cells for packaging and infectious viral particles
were obtained. Rats BMSCs were isolated and purified according to
the adhesion method. BMSCs were cultured in low-glucose DMEM medium
containing 10% FBS and 1% penicillin/streptomycin at 37°C, 5%
CO2. The cells were infected (multiplicity of
infection=150) and the infection efficiency was observed under
fluorescence microscope after 72 h of infection.
Animal middle cerebral artery occlusion
model and bone marrow mesenchymal stem cell transplantation
Forty-eight adult SD rats were randomly divided into
four groups. The suture occlusion method was used, as previously
described by Longa et al (17). In brief, a 0.2-mm suture was
inserted into the external carotid artery with a depth of 18.0±0.5
mm to occlude the entrance of the right middle cerebral artery to
induce ischemia. Two hours after the occlusion, the suture was
removed to create a reperfusion model. The rats were randomly
divided into four groups and 5×106 BMSCs were injected
via the tail vein 6 h after MCAO (18).
Evaluation of the cerebral infarction and
functional outcome
Functional outcome was evaluated by the modified
neurological severity score (mNSS) (19). The higher the mNSS score, the more
severe the neurological dysfunction. Triphenyltetrazolium chloride
(TTC) staining was carried out at days 1, 14 and 28 after MCAO. The
rats were sacrificed and the brains were collected. The coronal
sections (2 mm) were incubated in 2% TTC solution at 37°C for 15
min. The relative infarct volume was calculated based on methods
described by Neumann-Haefelin et al (20).
RT-PCR
Total RNA was extracted using using TRIzol
(Invitrogen Life Technologies). The optical density (OD) 260/OD280
values were measured by UV spectrophotometer to assess the quality
of the RNA. RNA was subsequently reverse-transcribed into cDNA by
RT-PCR. The thermal cycler parameters were 95°C, 3 min; 94°C, 30
sec, 59°C, 30 sec, 72°C, 45 sec, 36 cycles; and 72°C, 7 min; prior
to storage at 4°C. Following the reaction, 5 μl of PCR product for
each cell sample was used for 1.5% agarose gel electrophoresis. The
OD of each band was analyzed by a gel imaging system (I-box; UVP,
Upland, CA, USA). The experiment was carried out in triplicate and
the relative OD values were independently calculated. The primers
used for PCR amplification were as follows: 564 forward,
CTGGCCGCAGCCGCTGGAGACAC; and reverse, GGATATAGGCAGCTAACATCTCC;
803, GATGTTAGCTGCCTATATCCCAATG; and reverse, TTCA
CCCTGCAGTAGGTTTCTGCTGCCTTGTATAGGAGCA GCAACTTCACAATC; HIF1α
forward, GAAACCACC TATGACCTGC; and reverse, GTCGTGCTGAATAA
TACCACTC; β-actin forward, GGGACCTGACTGACTA CCTC; and
reverse, TCATACTCCTGCTTGCTGAT.
Immunohistochemistry
The rats were decapitated and the right brain
hemispheres were collected. The tissues were fixed, dehydrated,
embedded in paraffin and sectioned at the coronal plane. The
sections were then incubated with rabbit anti-mouse CD105 factor
antibody (1:200) for immunohistochemical staining. Following the
3,3′-diaminobenzidine reaction, the sections were stained with
hematoxylin. Microvessel density (MVD) was calculated based on the
method described in the study by Weidener et al (21). A brown-stained single endothelial
cell (or endothelial cell cluster) was used as a vessel count; six
fields on each slice were randomly selected for the microvessel
count. The number of vessels per unit (mm2) (microvessel
density) was calculated and averaged.
Western blot analysis
The total protein was extracted. Equal amounts of
protein (50 μg) were separated by 10% SDS-PAGE and transferred to a
polyvinylidene fluoride membrane. The membranes were blocked with
5% skimmed dry milk for 1 h at room temperature and incubated with
primary antibody [anti-HIF1α (rabbit) or anti-VEGF (rabbit),
1:1,000] at 4°C overnight. The membranes were washed and incubated
with the HRP-conjugated secondary antibodies for 1 h at room
temperature. Subsequent to washing, the membranes were incubated in
developing solution in a dark room for 30 min at room temperature.
The expression levels of targeted protein were detected by enhanced
chemiluminescence and analyzed using Quantity One (Bio-Rad,
Hercules, CA, USA). The experiment was carried out in triplicate
and the relative OD values were calculated.
Statistical analysis
Data are presented as the means ± standard
deviation. One-way analysis of variance was used to analyze the
difference among groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Infection of BMSCs with the recombinant
adenovirus particles
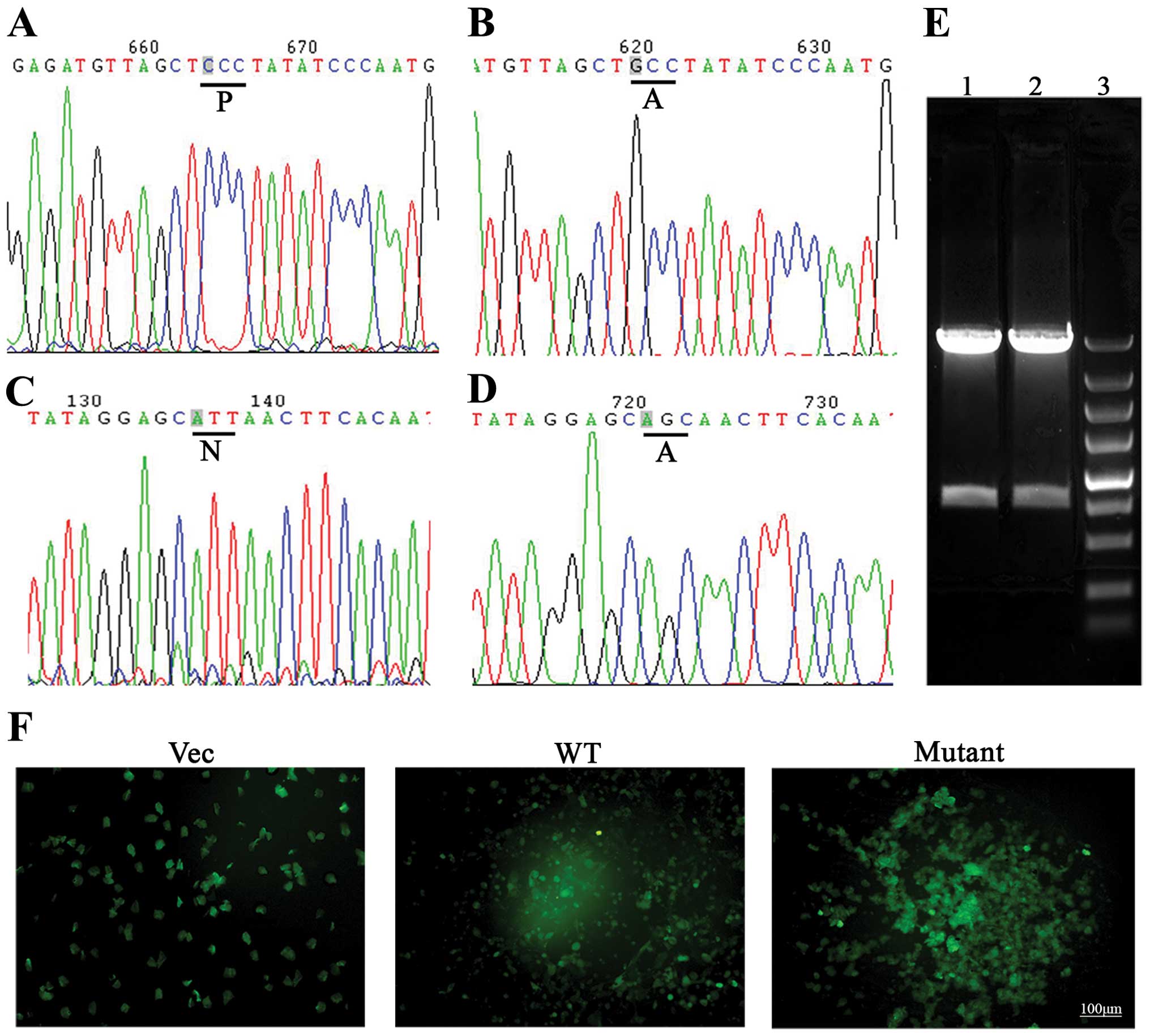
As shown in Fig.
1A–D, DNA sequencing assay showed that the proline at 564 and
the asparagine at 803 of HIF1α were mutated into alanine. A 30- and
a 3-kb band were observed following PacI cleavage (Fig. 1E). This mutant was transfected
into HEK293A cells using Lipofectamine 2000 (Fig. 1F). The infectious recombinant
adenovirus particles were harvested and the titer was
2.1×109 pfu/ml. Subsequently, the BMSCs were infected
using these virus particles containing P564A and N803A, and the
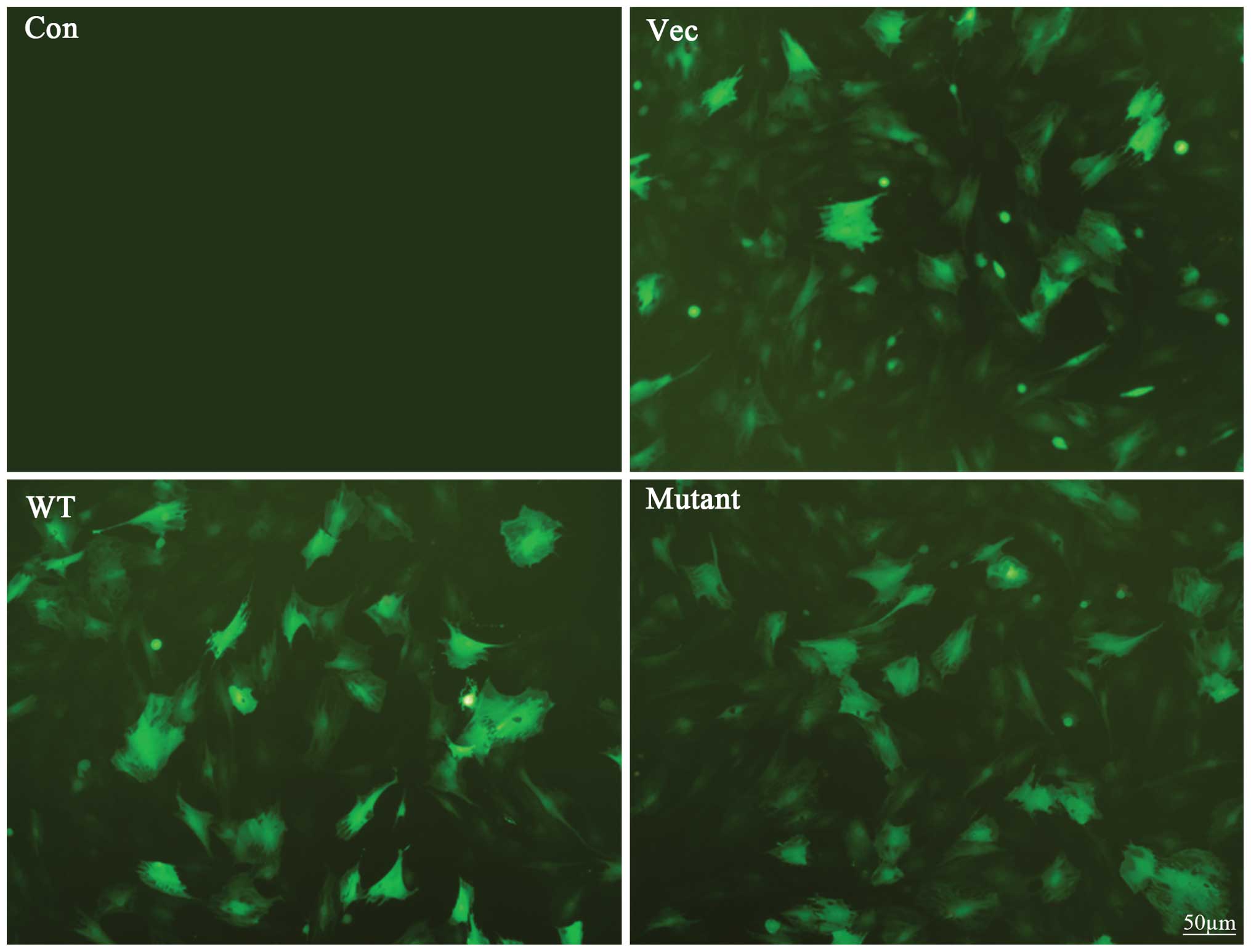
infectious status was observed by an immunofluorescence microscope.
HIF1α- and mutant-HIF1α transfected BMSCs showed strong expression
of green fluorescent protein (Fig.
2) indicating that mutant HIF1α can be expressed at high level
under nomorxic conditions.
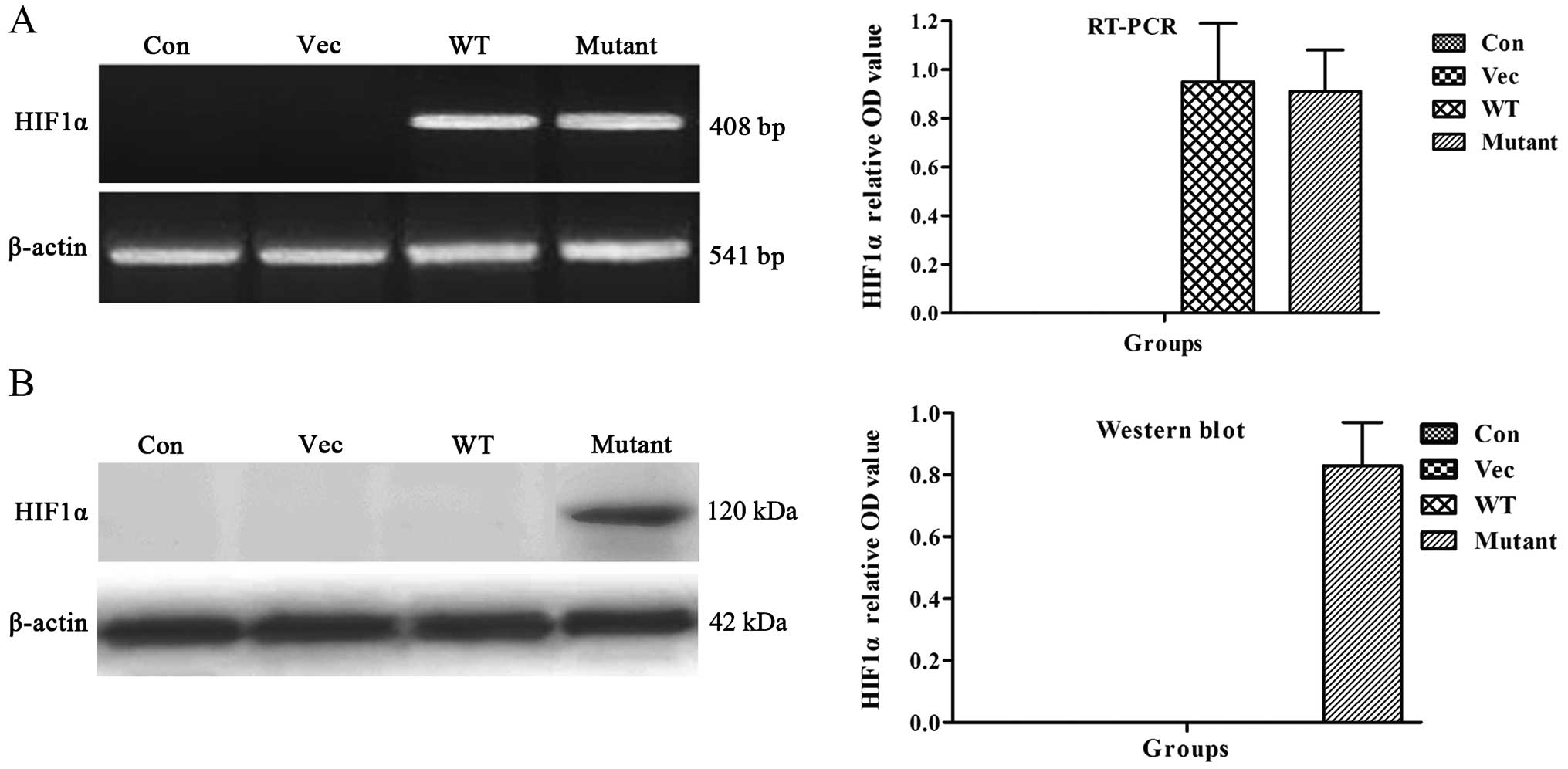
The expression of mutant HIF1α were quantitatively
examined in BMSCs at mRNA and protein levels under normoxic
conditions. RT-PCR showed that the expression of mutant
HIF1α mRNA could be detected under normoxic conditions
(Fig. 3A). Western blot analysis
revealed that the mutant HIF1α protein was only detected under
normoxic conditions. These data further indicated that the HIF1α
mutant could be expressed under normoxic conditions as compared to
HIF1α, which is only expressed under hypoxia conditions (Fig. 3B).
Mutant HIF1α improves the functional
outcome and decreases the infarct size
The mutant-HIF1α therapy outcomes in rats were
evaluated using the mNSS on days 1, 7, 14, 21 and 28. A lower mNSS
score indicates that less neurological defects resulted from MCAO
and represents a more improved outcome. As shown in Fig. 4A, there was a significant
improvement in neurological function in the mutant-HIF1α group
compared to the other three groups from days 14–28 (P<0.05).
However, no significant difference were observed in all the groups
at day 1 and day 7 after MCAO (P>0.05). The mNSS score was
significantly lower in the mutant HIF1α group than the other three
groups at day 14 (P<0.05), whereas the scores were similar among
the other three groups (P<0.05). Compared to the other groups,
the scores of the mutant-HIF1α group decreased more significantly
at day 21 and day 28, indicating that the mutant HIF1α could
effectively mitigate motor function (P<0.05) (Fig. 4A). Compared to the other groups,
TTC staining showed that the infarcted area decreased significantly
in the mutant-HIF1α groups. This indicated that mutant HIF1α could
attenuate cerebral ischemic injury in rats. The infarct area (white
area) was similar among all the groups at day 1 (P<0.05). On day
14, the infarct size in the mutant-HIF1α group was significantly
reduced, compared to the other three groups (P<0.05); at day 28,
the reduction in the infarct size became clearer in comparison to
the other groups (P<0.05), indicating that mutant HIF1α
effectively reduced ischemic injury in rats (Fig. 4B and C).
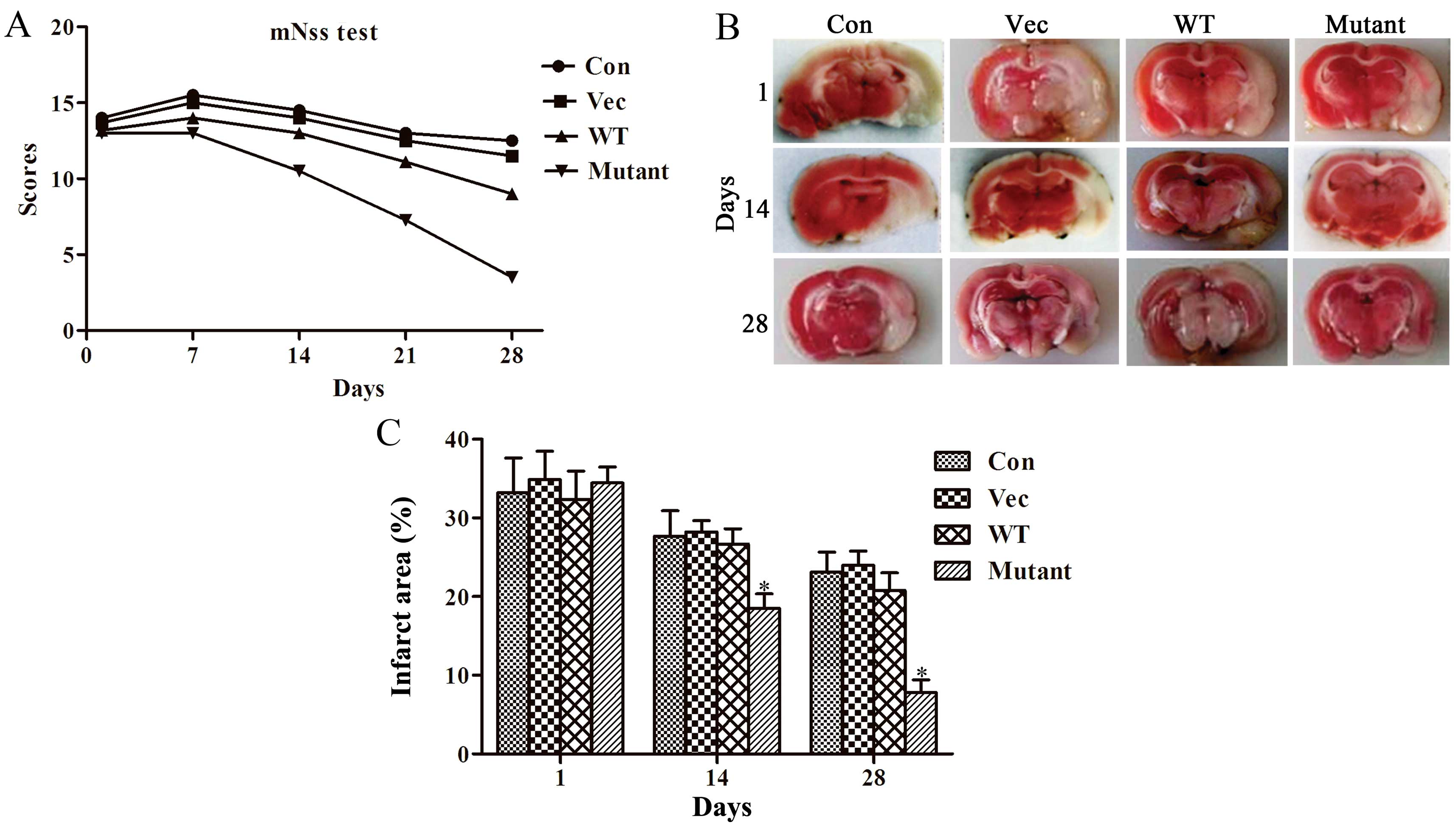 | Figure 4Mutant HIF1α improved the functional
outcome and decreased infarct size. (A) Neurological function
evaluation at days 1, 7, 14, 21, 28 after MCAO. These experiments
were repeated three times. (B) TTC staining of the infarcted area
at days 1, 14 and 28 after MCAO. (C) Quantitative analysis of TTC
staining (P<0.05). The data are represented by the mean ±
standard deviation and analyzed by one-way ANOVA. The difference is
considered statistically significant when P<0.05. HIF1α,
hypoxia-inducible factor 1α; Con, control; Vec, vector; WT,
wild-type HIF1α; MCAO, middle cerebral artery occlusion model; TTC,
triphenyltetrazolium chloride; ANOVA, analysis of variance. |
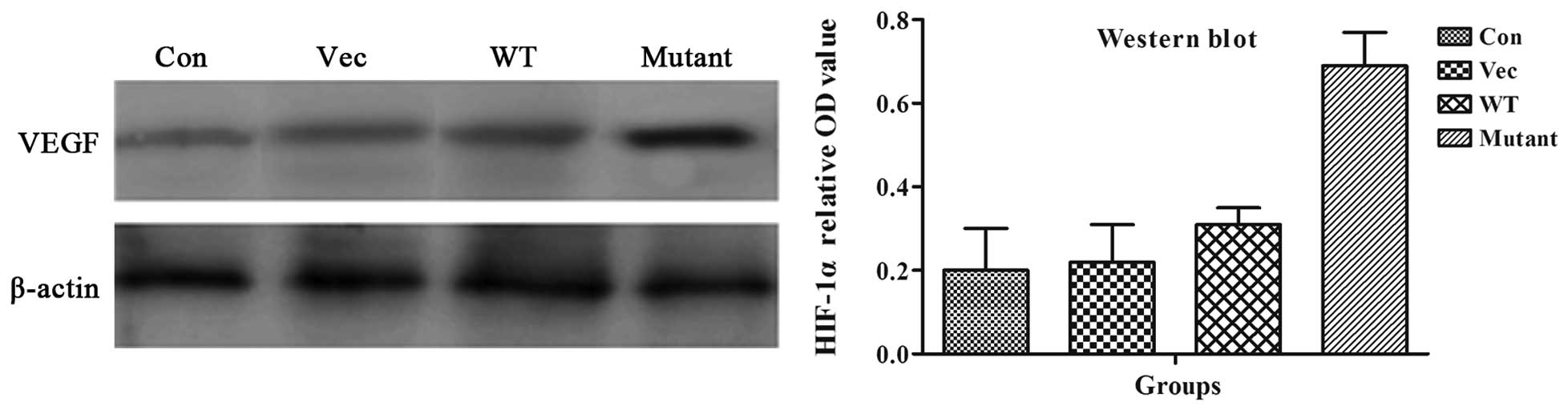
VEGF protein expression and revasculation
in the mutant group
Western blot analysis showed that VEGF protein
expression was significantly higher in the mutant-HIF1α group than
the other three groups (P<0.05), indicating that the mutant
HIF1α significantly increased the expression of the downstream
gene, VEGF, which was consistent with the mutant-HIF1α function
under normoxic conditions (Fig.
5).
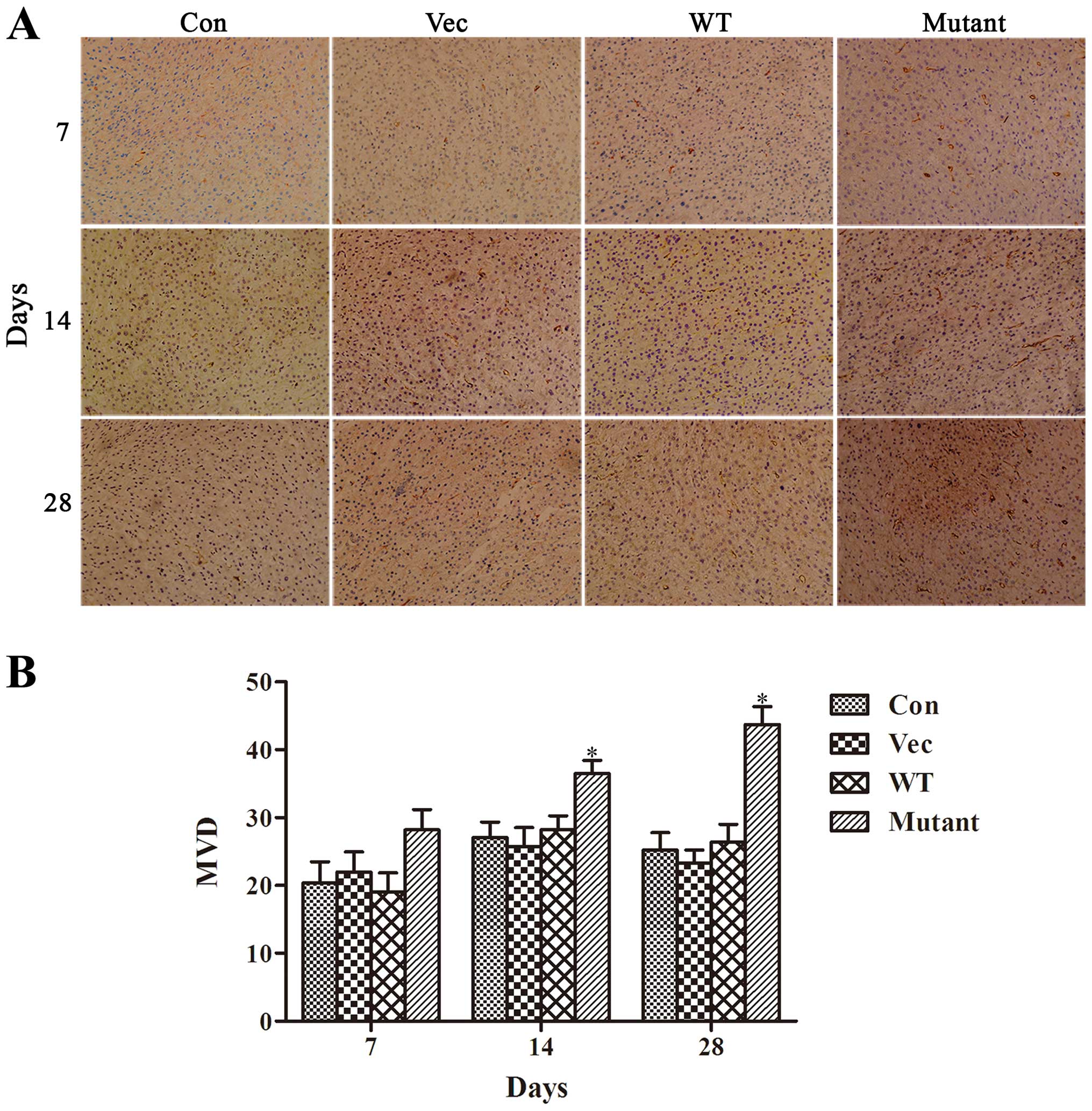
The number of new blood vessels at the ischemia
areas were also evaluated at the same time. For the wild-type HIF1α
group, a small amount of scattered microvascular could be observed
at 7 days after cerebral ischemia. The revasculation peaked at day
14 after ischemia and subsequently decreased gradually. Whereas,
the mutant-HIF1α group showed that the density of neomicrovascular
vessels was denser at day 7, peaked at day 14 and maintained at a
relatively higher level at day 28 after MCAO (Fig. 6).
Discussion
In the present study, a recombinant adenovirus HIF1α
was established in which the proline at 564 and asparagine at 803
were mutated simultaneously and explored the significance in the
clinical treatment of cerebral ischemia. Numerous data have
demonstrated that the evidence indicates that the 564-proline and
the 803-asparagine are important loci that regulate HIF1α
expression. The 564-proline in the ODDD of HIF1α is degraded by
proline hydroxylase and hydroxylation of the 803-asparagine can
inhibit HIF1α transcriptional activity under normoxic conditions
(11–15). For these reasons, HIF1α
accumulates only under hypoxia conditions. In the present study,
the P to A mutation at 564 prevented oxygen-dependent degradation
of HIF1α and the N to A mutation of the 803-asparagine increased
transcriptional activity of HIF1α, which all lead to a stable HIF1α
expression under normoxic conditions. The present data have
demonstrated that the mutant HIF1α could be expressed at a
relatively higher level under normoxic conditions. In addition, the
mutant HIF1α is more stable compared to wild-type HIF1α under
normoxic conditions with a higher expression level without
degradation.
HIF1α mRNA expression, but not protein
expression, was found under normoxic conditions, which indicates
that wild-type HIF1α protein can be completely degraded under
normoxic conditions. However, the mutant HIF1α was expressed under
normoxic conditions. These data indicated that the degradation of
wild-type HIF1α occurs at the post-transcriptional translation
stage.
BMSCs were infected with adenovirus particles
containing the mutant-HIF1α gene in vitro and found
that transfection with the mutant HIF1α effectively induced the
expression of HIF1α under normoxic conditions, which provided a
foundation for our in vivo animal experiments. In addition,
VEGF protein expression in the animals treated with BMSCs
overexpressing the mutant HIF1α were significantly higher than that
of the animals treated with BMSCs overexpressing the wild-type
HIF1α. VEGF is a HIF1α-downstream gene, indicating that
mutant HIF1α is not only expressed under normoxic conditions, but
also effectively promotes downstream gene expression, including
VEGF, which is consistent with the wild-type HIF1α function
under hypoxic conditions.
Immunohistochemistry results showed that the MVD was
significantly higher in the animals treated with BMSCs
overexpressing the mutant HIF1α at days 7, 14 and 28 compared to
the other three groups, indicating that mutant HIF1α promoted
angiogenesis and revascularization in the cerebral ischemia area,
thereby accelerate the process of motor function recovery.
Correspondingly, the infarct volume was significantly reduced in
the animals treated with BMSCs overexpressing the mutant HIF1α than
that of the animals treated with BMSCs overexpressing the wild-type
HIF1α, which is consistent with the behavioral test results
(22–24).
A previous study has shown that mutant HIF1α protein
can be efficiently expressed in vitro and in vivo
under normoxic conditions, which accelerates the angiogenesis and
the formation of microvessel system, improve local blood supply and
thus, facilitate blood perfusion to ischemic brain tissue (25). Wild-type HIF1α is unstable in a
complex in vivo environment and less effective in promoting
in vivo angiogenesis. In the present study, the unknown
consequence of gene mutation was not encountered. The results
showed that mutant HIF1α has generated new potential for the
treatment of ischemic cerebrovascular disease.
Numerous data have demonstrated that rat bone marrow
mesenchymal stem cells can pass the blood brain barrier (26–29). In the present study, mutant-HIF1α
infected-BMSCs was injected into into rats and the BMSCs passed
through the blood-brain barrier and achieved therapeutic effects.
However, due to the limited experimental conditions, the amount of
HIF1α that successfully transferred into brain tissue was not
measured. This area requires exploring further in future
studies.
References
|
1
|
Giaccia A, Siim BG and Johnson RS: HIF-1
as a target for drug development. Nat Rev Drug Discov. 2:803–811.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khan TA, Sellke FW and Laham RJ: Gene
therapy progress and prospects: therapeutic angiogenesis for limb
and myocardial ischemia. Gene Ther. 10:285–291. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Semenza GL: Evaluation of HIF-1 inhibitors
as anticancer agents. Drug Discov Today. 12:853–859. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wan C, Gilbert SR, Wang Y, et al:
Activation of the hypoxia-inducible factor-1α pathway accelerates
bone regeneration. Proc Natl Acad Sci USA. 105:686–691. 2008.
|
|
5
|
Pugh CW and Ratcliffe PJ: Regulation of
angiogenesis by hypoxia: role of the HIF system. Nat Med.
9:677–684. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Majmundar AJ, Wong WJ and Simon MC:
Hypoxia-inducible factors and the response to hypoxic stress. Mol
Cell. 40:294–309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl
Acad Sci USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fiorenzo P, Mongiardi MP, Dimitri D, et
al: HIF1-positive and HIF1-negative glioblastoma cells compete
in vitro but cooperate in tumor growth in vivo. Int J
Oncol. 36:785–791. 2010.PubMed/NCBI
|
|
9
|
Giatromanolaki A, Fiska A, Pitsiava D,
Kartalis G, Kouourakis MI and Sivridis E: Erythropoietin receptors
in endometrial carcinoma as related to HIF1α and VEGF expression.
In vivo. 23:699–703. 2009.PubMed/NCBI
|
|
10
|
Rey S and Semenza GL: Hypoxia-inducible
factor-1-dependent mechanisms of vascularization and vascular
remodelling. Cardiovasc Res. 86:236–242. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bruick RK and McKnight SL: A conserved
family of prolyl-4-hydroxylases that modify HIF. Science.
294:1337–1340. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dang EV, Barbi J, Yang HY, et al: Control
of T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell.
146:772–784. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Evans CE, Humphries J, Mattock K, et al:
Hypoxia and upregulation of hypoxia-inducible factor 1α stimulate
venous thrombus recanalization. Arterioscler Thromb Vasc Biol.
30:2443–2451. 2010.
|
|
14
|
Kazi AA, Molitoris KH and Koos RD:
Estrogen rapidly activates the PI3K/AKT pathway and
hypoxia-inducible factor 1 and induces vascular endothelial growth
factor A expression in luminal epithelial cells of the rat uterus.
Biol Reprod. 81:378–387. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lando D, Peet DJ, Whelan DA, Gorman JJ and
Whitelaw ML: Asparagine hydroxylation of the HIF transactivation
domain a hypoxic switch. Science. 295:858–861. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ralph GS, Parham S, Lee SR, et al:
Identification of potential stroke targets by lentiviral vector
mediated overexpression of HIF-1α and HIF-2α in a primary neuronal
model of hypoxia. J Cereb Blood Flow Metab. 24:245–258.
2004.PubMed/NCBI
|
|
17
|
Longa EZ, Weinstein PR, Carlson S, Cummins
R, et al: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu N, Zhang Y, Fan L, et al: Effects of
transplantation with bone marrow-derived mesenchymal stem cells
modified by Survivin on experimental stroke in rats. J Transl Med.
9:1052011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boltze J, Kowalski I, Geiger K, et al:
Experimental treatment of stroke in spontaneously hypertensive rats
by CD34+ and CD34− cord blood cells. Ger Med
Sci. 3:Doc092005.PubMed/NCBI
|
|
20
|
Neumann-Haefelin T, Kastrup A, de
Crespigny A, Yenari MA, Ringer T, Sun GH and Moseley ME: Serial MRI
after transient focal cerebral ischemia in rats: dynamics of tissue
injury, blood-brain barrier damage, and edema formation. Stroke.
31:1965–1972. 2000. View Article : Google Scholar
|
|
21
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis-correlation in invasive breast
carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mustafa AK, Ahmad AS, Zeynalov E, et al:
Serine racemase deletion protects against cerebral ischemia and
excitotoxicity. J Neurosci. 30:1413–1416. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ruiz-Crespo S, Trejo-Gabriel-Galán JM and
Coma-del-Corral MJ: Localizing coordinates of cerebral ischemic
tissue without the need of staining in a rat model of focal
cerebral infarct. Metab Brain Dis. 28:21–24. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu J, Zhu H, Ko D and Kindy MS:
Motoneuronotrophic factor analog GM6 reduces infarct volume and
behavioral deficits following transient ischemia in the mouse.
Brain Res. 1238:143–153. 2008.PubMed/NCBI
|
|
25
|
Elson DA, Thurston G, Huang LE, Ginzinger
DG, McDonald DM, Johnson RS and Arbeit JM: Induction of
hypervascularity without leakage or inflammation in transgenic mice
overexpressing hypoxia-inducible factor-1α. Genes Dev.
15:2520–2532. 2001.
|
|
26
|
Menge T, Zhao Y, Zhao J, et al:
Mesenchymal stem cells regulate blood-brain barrier integrity
through TIMP3 release after traumatic brain injury. Sci Transl Med.
4:161ra1502012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pati S, Khakoo AY, Zhao J, et al: Human
mesenchymal stem cells inhibit vascular permeability by modulating
vascular endothelial cadherin/β-catenin signaling. Stem Cells Dev.
20:89–101. 2011.PubMed/NCBI
|
|
28
|
Warach S and Latour LL: Evidence of
reperfusion injury, exacerbated by thrombolytic therapy, in human
focal brain ischemia using a novel imaging marker of early
blood-brain barrier disruption. Stroke. 35:2659–2661. 2004.
View Article : Google Scholar
|
|
29
|
Zacharek A, Chen J, Cui X, et al:
Angiopoietin1/Tie2 and VEGF/Flk1 induced by MSC treatment amplifies
angiogenesis and vascular stabilization after stroke. J Cereb Blood
Flow Metab. 27:1684–1691. 2007. View Article : Google Scholar : PubMed/NCBI
|