Introduction
Macrovascular diseases, such as atherosclerosis, are
the most frequent complications of diabetes (1). Diabetes mellitus impairs endothelial
function and may be an indicator of the cardiovascular disease
development (2,3). Vascular endothelium is considered to
play an essential role in diabetes-associated vascular dysfunction,
including atherosclerosis (4).
The endothelium plays a significant role in the regulation of
vascular function and the development of physiological and
pathophysiological inflammation (5–8).
Endothelial cell injury is a critical element of atherosclerosis
and hypertension (9,10). Previous studies have shown that
H2O2 induced endothelial cell apoptosis, and
causes cellular dysfunction and cell death (11,12). The vascular function pathogenesis
is complicated and there are a number of signaling pathways,
including the Notch pathway.
The Notch signaling pathway is one of the pathways
that plays a significant role in cell differentiation, primarily
determining and regulating cell survival (13,14). In mammals, four receptors
(Notch1-Notch4) and five ligands, including Jagged1, Jagged2,
Delta-like 1 (Dll), Dll3 and Dll4, have been discovered (15,16). Notch signaling also affects
cellular activities, including proliferation, migration, growth,
differentiation and death (17).
In addition, Notch activity controls the communication between
cells, signal transduction in the cytoplasm and gene transcription
in the nucleus. The genes downstream of Notch signaling include
Hairy and enhancer of split 1 (Hes1) and the Hairy-related
transcription (HRT) factor family. The binding of a ligand and
receptor induces a conformational change of the Notch receptor.
This allows an extracellular metalloprotease to cleave the
receptor, which allows the γ-secretase-mediated protease to release
the Notch intracellular domain. Subsequently, the Notch
intracellular domain travels into the nucleus where it activates
the transcription of downstream genes, such as Hes1 (18). A recent study indicates that in
H2O2-induced cell apoptosis, the Notch
signaling pathway was upregulated (19), indicating that Notch inhibition
may be a useful method in the protection of cells from
H2O2-induced apoptosis.
Thus far, detrimental effects induced by
H2O2 on human endothelial cells can be
suppressed by numerous types of plant active substances (12,20–21). Vaccariae semen, the seeds
of Vaccaria segetalis (Neck.) Garcker. ex Asch.
(Caryophyllaceae), is a famous traditional medicinal plant
(22) for activating blood to
promote menstruation, invigorating blood circulation, regulating
menstrual disturbance and dispelling edema, promoting diuresis and
milk secretion, and relieving carbuncles (23–24). It contains flavonoids, cyclic
peptides, triterpene saponins, lipids, aliphatic acids,
monosaccharides, biotin, and coumarin (25–31), with a few of these compounds
demonstrating bioactivity, such as anti-angiogenesis and
growth-inhibitory activity on luteal, HL-60 and endothelial cells
(32–33). Vaccarin is a major flavonoid
glycoside in Vaccariae semen and is considered one of the
main active constituents, which has attracted increasing attention
(34). The present study
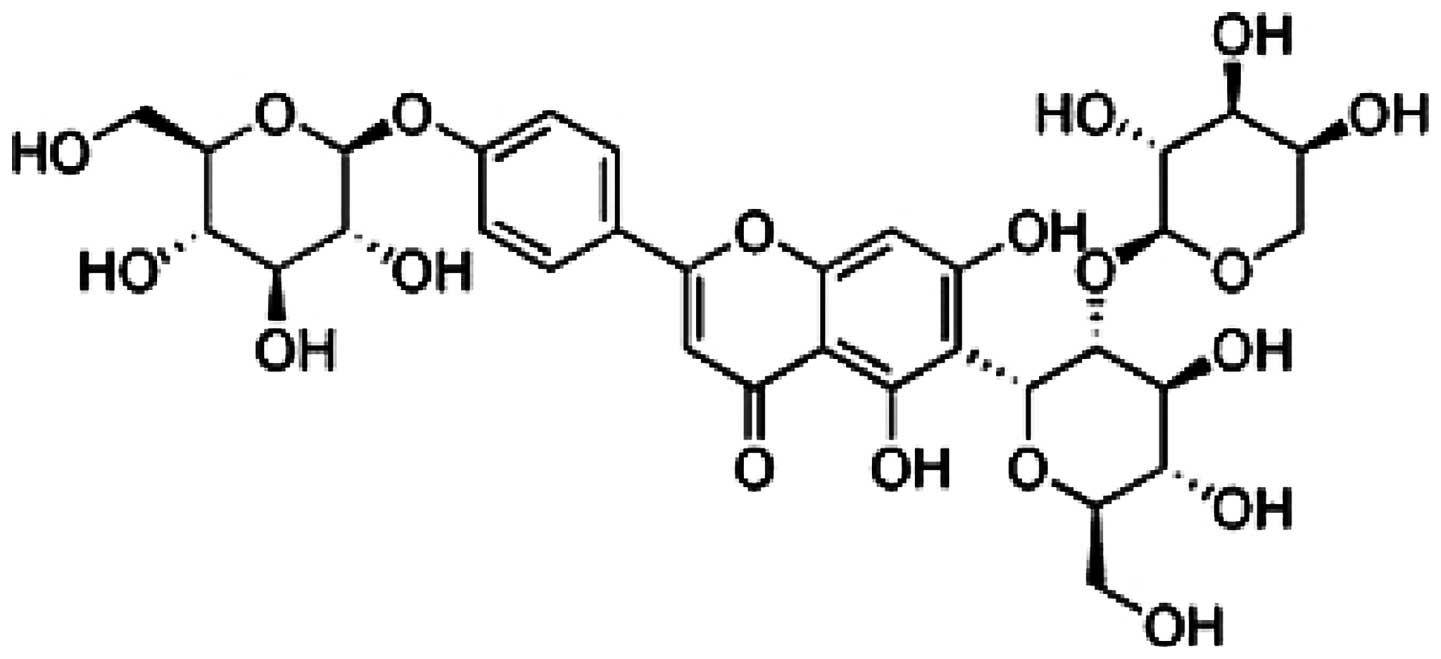
investigated the protective effect of vaccarin (Fig. 1) on human umbilical vein
endothelial cell lines (EA.hy926) injured by
H2O2 in vitro in order to further
understand the efficacy of this medicine.
The involvement of the Notch signaling pathway in
the vaccarin-induced protective effects observed during
H2O2-induced cellular injury remains to be
studied. In the present study, the role of the Notch signaling
pathway in H2O2-induced cellular injury in
EA.hy926 cells was first explored. Furthermore, whether vaccarin
protected EA.hy926 cells from H2O2-induced
cellular injury via Notch signaling pathway inhibition was
investigated. To know whether vaccarin reduced Notch1 and apoptosis
in vitro, the role of vaccarin on the reduction of apoptosis
in EA.hy926 cells within the range of an effective concentration
was evaluated.
Materials and methods
Drugs and chemicals
Vaccarin was purchased from Shanghai Shifeng
Technology Co., Ltd. (Shanghai, China). Sulforhodamine B (SRB) was
purchased from Sigma (St. Louis, MO, USA). Annexin V-fluorescein
isothiocyanate (FITC)/propidium iodide (PI) Apoptosis Detection kit
was purchased from KeyGen (Nanjing, China). The
3,3′-diaminobenzidine (DAB) kit was purchased from Beyotime
Institute of Biotechnology (Jiangmen, Jiangsu, China). The Notch1
(ab52627), Hes1 (ab108937), caspase-3 (ab32351) and β-tublin
(ab6046) antibodies were purchased from Abcam (Hong Kong, China).
The goat anti-rabbit secondary antibody (AB10058) was purchased
from SangonBiotech Co., Ltd. (Shanghai, China). The kits for the
measurement of lactate dehydrogenase (LDH), methane dicarboxylic
aldehyde (MDA), superoxide dismutase (SOD) and bicinchoninic acid
(BCA) concentrations were purchased from the Institute of Jiancheng
Bioengineering (Nanjing, Jiangsu, China). M-PER Mammalian Protein
Extraction reagent was purchased from Thermo Scientific (Waltham,
MA, USA).
Cell culture and treatments
Human EA.hy926 (ATCC CRL-2922) endothelial cells
were cultured in Dulbecco’s modified Eagle’s medium (Hyclone,
Logan, UT, USA) supplemented with 10% fetal calf serum (Gibco,
Carlsbad, CA, USA) and incubated at 37°C in humidified air
containing 5% CO2.
Prior to treatment with H2O2,
cells were grown to 80–90% confluence and placed in 2%
serum-containing media for 12 h to achieve cell synchronization.
The vaccarin solution was diluted with culture medium immediately
prior to the experiment. The cells were treated with
H2O2 in the absence or presence of vaccarin.
The cell monolayers were pretreated with vaccarin (3.44, 6.88 or
13.76 μM) at 37°C for 24 h, and were subsequently induced by
H2O2. Following treatment with
H2O2, cells were maintained in 10%
serum-containing media in a 5% CO2 atmosphere at 37°C
for further experiments.
Analysis of cell viability
The SRB assay was performed to assess EA.hy926 cell
viability (35). EA.hy926 cells
were seeded in 96-well culture plates (each concentration for four
repeat holes) and cultured in medium for 24 h. Subsequently, three
different final concentrations of vaccarin (3.44, 6.88 or 13.76 μM,
dissolved in serum-free medium) were added to each well. After 24 h
incubation at 37°C, H2O2 solution (250, 500
or 1000 μM) dissolved in serum-free medium was added to each well
and cultivated (for 2 or 4 h). Subsequently, the medium was removed
and 200 μl 5% trichloroacetic acid (TCA) was added to each well to
fix the cells for 40 min at 4°C. TCA solution was removed and
replaced with 100 μl SRB and incubated at 30°C for 30 min.
Following this, SRB was removed and the cells were washed twice in
deionized water. Finally 10% tris hydroxymethyl aminomethane (Tris)
was used to dissolve the SRB and the samples were agitated for
30sec at room temperature twice. The results were determined at 540
nm using a Multiskan MK3 microplate reader (Thermo Labsystems,
Milford, MA, USA) and the cell viability was expressed as an
optical density (OD) value. In addition, the cell morphology was
observed under an inverted/phase contrast microscope, and images
were captured at 200 amplification with Olympus Nikon Eclipse Ti
(Tokyo, Japan).
Cell migration assay
The migration rate was measured by the wound healing
assay (36). Briefly, EA.hy926
cells (8×104 cells/well) were seeded and were cultured
at 37°C in a saturated humidity containing 5% CO2 for 24
h. When the cells have attached completely, the middle of the cell
plate was scraped with a line ~1 mm in width following treatment of
vaccarin (6.88 and 13.76 μM). The cells were incubated and randomly
chosen fields were photographed at 100 amplification under a
microscope video system (Olympus, Nikon Eclipse Ti). The mean
distance between the two ends of the scratch was quantified by
manual measurements. The control was defined as 100%.
Measurement of LDH release and the
intracellular contents of SOD and MDA
LDH, an indicator of cell injury, was detected with
an assay kit according to the manufacturer’s instructions. The
activity of enzyme was expressed as units per liter, and the
absorbance was read at 450 nm. As described previously (37), the activities of SOD and MDA were
determined using commercially available kits according to the
manufacturer’s instructions. The enzyme activities were expressed
as units per milligram of protein. The assay for measuring SOD
activity was based on the ability of SOD to inhibit the oxidation
of hydroxylamine by O2− produced from the
xanthine-xanthine oxidase system. One unit of SOD activity was
defined as the amount that reduced the absorbance at 450 nm by 50%.
The experiment of BCA measurement was performed prior to
determining of SOD. MDA was measured at 532 nm by its reaction with
thiobarbituric acid to form a stable chromophoric product. The MDA
level was expressed as nanomoles per milligram protein.
Cellular apoptosis assay
EA.hy926 cells were prepared for analysis according
to the instructions of the Annexin V-FITC/PI Apoptosis Detection
kit. The stained cells were quantitatively detected using the
FACScan flow cytometer (BD Biosciences, San Jose, CA, USA) in the
FL1-H and FL2-H channels. Data were analyzed using Cell Quest Pro
software (BD Biosciences). A total of 10,000 cells were
analyzed.
Hoechst staining
In the 24-well plate with cover slips, after
6×104 EA.hy926 cells were seeded onto each well and
cultured for 24 h, different doses of vaccarin (6.88 and 13.76 μM)
were applied and incubated 24 h before being subjected to 4 h of
H2O2 (1000 μM) treatment. Following the
removal of the culture medium, the cells were fixed with 0.5 ml 4%
paraformaldehyde, and washed with phosphate-buffered saline (PBS)
twice. After treatment with the Hoechst dyes (Wuhan Boster
Biological Technology, Ltd., Wuhan, China) for 10 min, the cells
were rinsed with PBS twice. The stained cells were immediately
observed under a fluorescence microscope (Olympus, Nikon Eclipse
Ti).
Western blot analysis
Protein levels were analyzed by western blot as
described previously (38).
Briefly, 25 μg total protein/well was loaded after denaturing in
loading buffer at 100°C for 5 min. The protein extracts were
subjected to 8–12% SDS-PAGE and transferred to a nitrocellulose
membrane (Millipore, Billerica, MA, USA). Following the transfer,
the membranes were blocked at room temperature for 2 h in 5%
skimmed dry milk/TBST and were incubated at 4°C overnight with
various primary antibodies. The primary antibodies are as follows:
Notch1 (1:500 dilution), Hes1 (1:500 dilution), caspase-3 (1:1000
dilution) and β-tublin (1:1000 dilution). The following day, the
membranes were washed three times with TBST for 10 min at room
temperature, and were subsequently incubated in secondary antibody
(anti-rabbit immunoglobulin G, 1:2000 dilution) conjugated to
horseradish peroxidase for 2 h at room temperature. Following
incubation, the membranes were washed as above, and the protein
bands were visualized using the DAB-advanced western blotting
detection kit. β-tublin was used as the protein loading
control.
Statistical analysis
The results are presented as mean ± standard
deviation. Statistical analysis was performed by one-way analysis
of variance test. P<0.05 was considered to indicate a
statistically significant difference.
Results
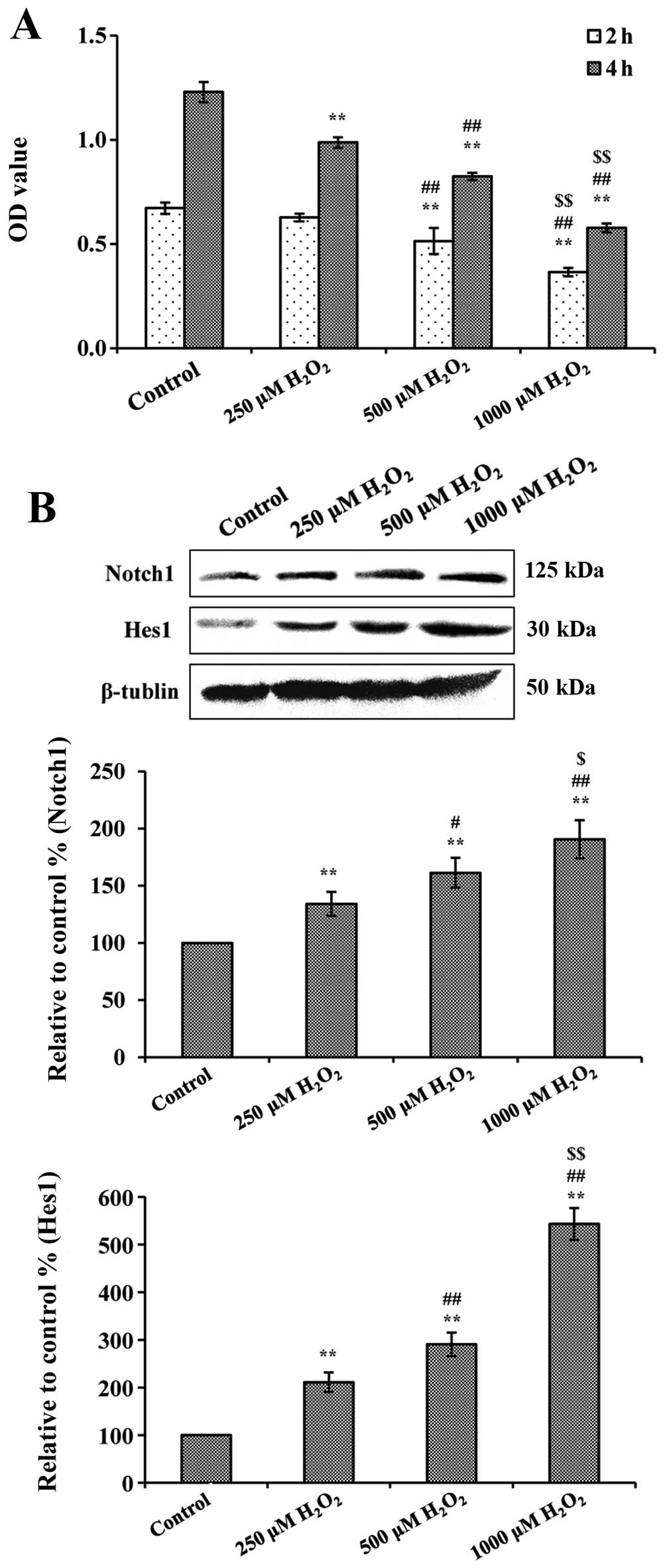
Effect of H2O2 on
the viability of EA.hy926 cells and Notch1 and Hes1 expression
In order to investigate the effect on EA.hy926 cells
induced by H2O2, the cells were induced by
H2O2 (250, 500 and 1000 μM) for 2 and 4 h and
cell viability was examined with the SRB assay. As shown in
Fig. 2A, treatment with
H2O2 alone significantly reduced cell
viability by >50% (after 4 h treatment at 1000 μM
H2O2). The OD values in the control group
were 0.673±0.027 and 1.229±0.049 after 2 and 4 h of culture. All
the groups had a significant decrease compared to the normal
groups, except 250 μM H2O2 after 2 h culture
(P<0.01, compared to respective control groups). Evenually, 4 h
treatment with 1000 μM H2O2 was selected for
the subsequent experiments. In addition, Notch1 and Hes1 expression
were detected after 4 h H2O2 culture and the
results indicated that H2O2 treatment
significantly increased the expression of Notch1 and Hes1 in a
dose-dependent manner (shown in Fig.
2B).
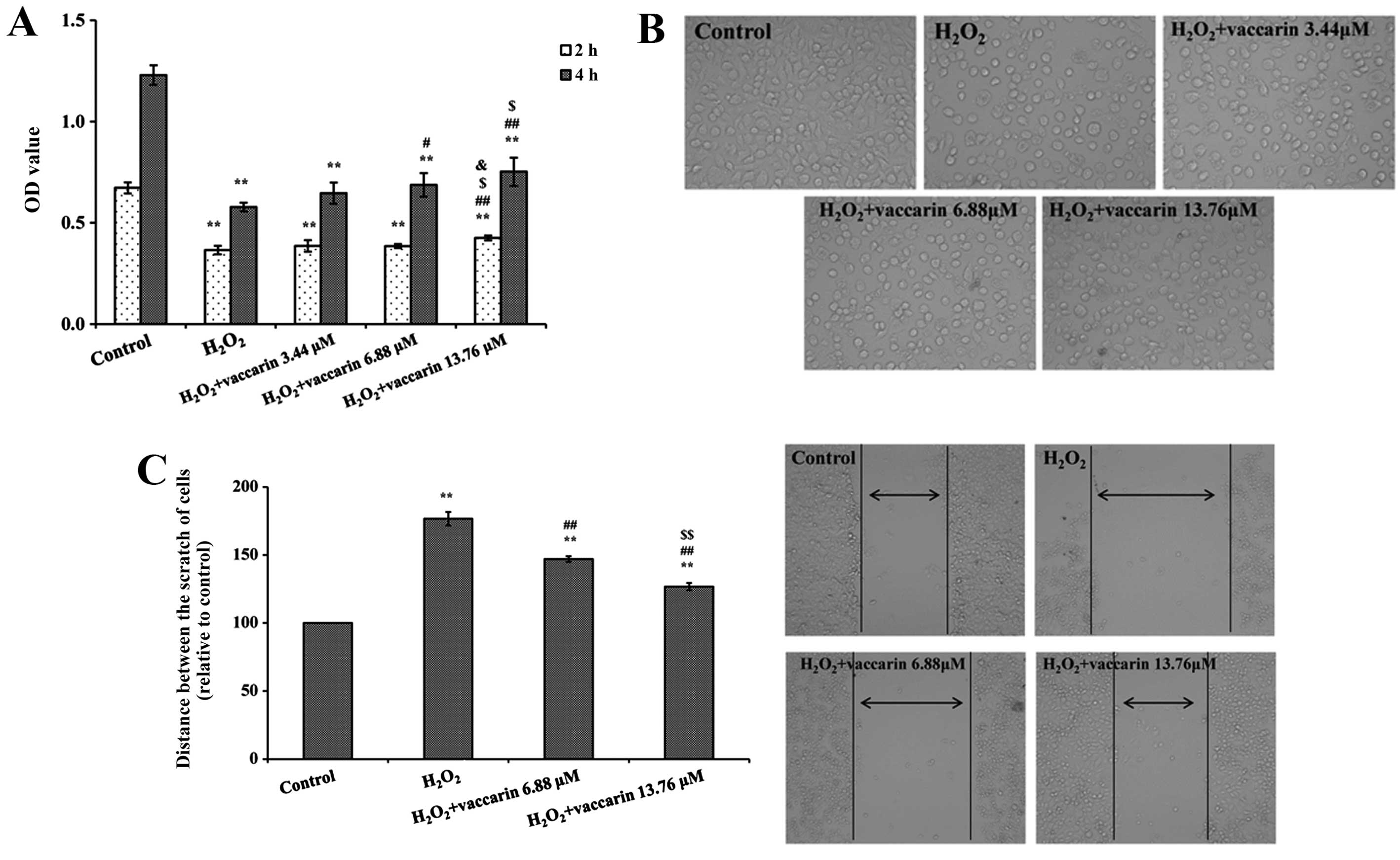
Effects of vaccarin on the viability and
migratory ability of H2O2-injured EA.hy926
cells
The effects of vaccarin on EA.hy926 cell
proliferation were examined after 2 and 4 h treatment with 1000 μM
H2O2. As shown in Fig. 3A, the cell viability in the
presence of vaccarin groups significantly increased compared to the
absence groups of vaccarin (P<0.05 or P<0.01), respectively.
Vaccarin provided dose-dependent protection against the reduction
in cell viability induced by H2O2 for the
concentration range, 3.44–13.76 μM. As observed under the
microscope, H2O2 treatment resulted in
significant cell shrinkage compared to the control group. However,
pretreatment with three different vaccarin concentrations (3.44,
6.88 and 13.76 μM) attenuated H2O2-injured
cell shrinkage (shown in Fig.
3B). Based on these results, pretreatment with 6.88 and 13.76
μM vaccarin and 1000 μM H2O2 for 4 h was
chosen for further studies.
As shown in Fig.
3C, following treatment with H2O2 the
migratory ability of cells was decreased and the distance between
the scratch (μm) of cells was 176.68±4.89% (P<0.01, compared to
the normal cells). However, vaccarin (6.88 and 13.76 μM) treatment
groups significantly decreased the distances (147.01±2.14% and
126.76±2.68%;, P<0.01, compared to the
H2O2 group, respectively).
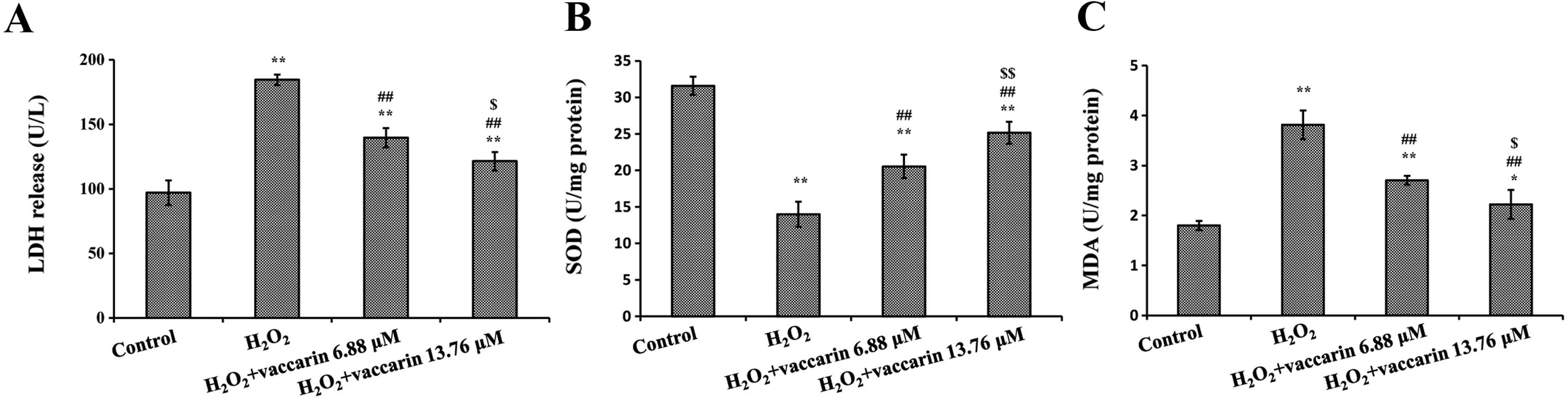
Effects of vaccarin on SOD, LDH release
and MDA levels in H2O2-injured EA.hy926
cells
Treating the cells with H2O2
for 4 h decreased the SOD levels, but increased the LDH release and
MDA levels (P<0.01, compared to the control group,
respectively). As demonstrated in Fig. 4, following incubation of EA.hy926
cells in the presence of vaccarin (6.88 and 13.76 μM) with
H2O2 significantly increased SOD activity
(Fig. 4A) and decreased the level
of MDA and LDH release, respectively (Fig. 4B and C). According to these
results, vaccarin significantly changed the SOD activity, LDH
leakage and MDA level in H2O2-induced
endothelial cells in a concentration-dependent manner.
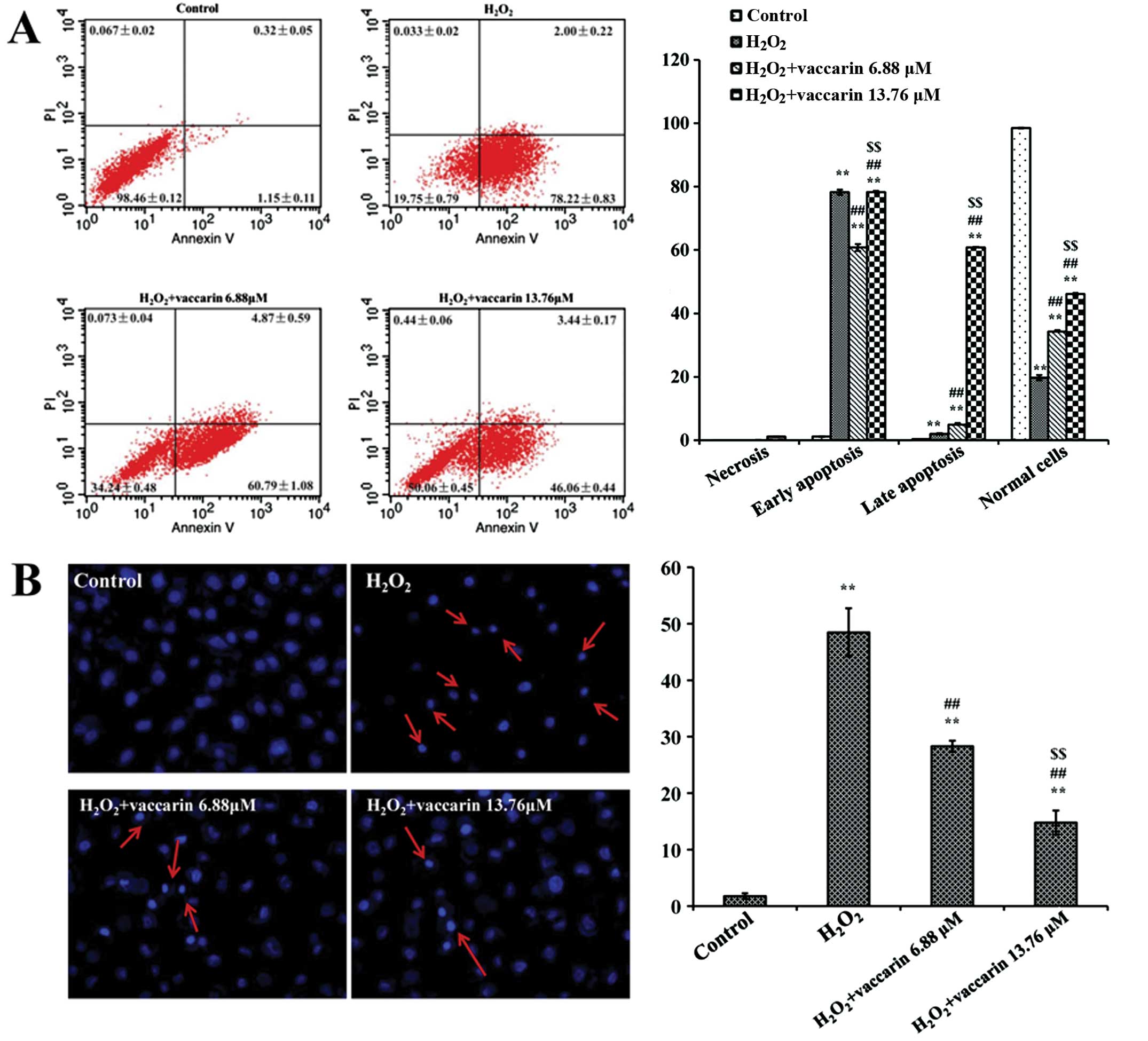
Effects of vaccarin on the apoptotic
index of H2O2-injured EA.hy926 cells
Induction of apoptosis was measured by Annexin V and
PI-double staining. The flow cytometric analysis of the
H2O2 group showed an increase in apoptosis.
The ratio of prophase and late apoptosis reached 78.22±0.83% and
2.00±0.22%, respectively (P<0.01, compared to the control
group). However, the apoptosis ratio for the treatment with
vaccarin (6.88 and 13.76 μM) groups significantly reduced
(P<0.01, compared to the H2O2 group, shown
in Fig. 5A).
In the Hoechst stain experiment, it was also found
that apoptosis was significantly higher in the
H2O2 group (shown in Fig. 5B). Following treatment with
vaccarin (6.88 and 13.76 μM), the cell index was markedly decreased
(P<0.01, compared to the H2O2 group).
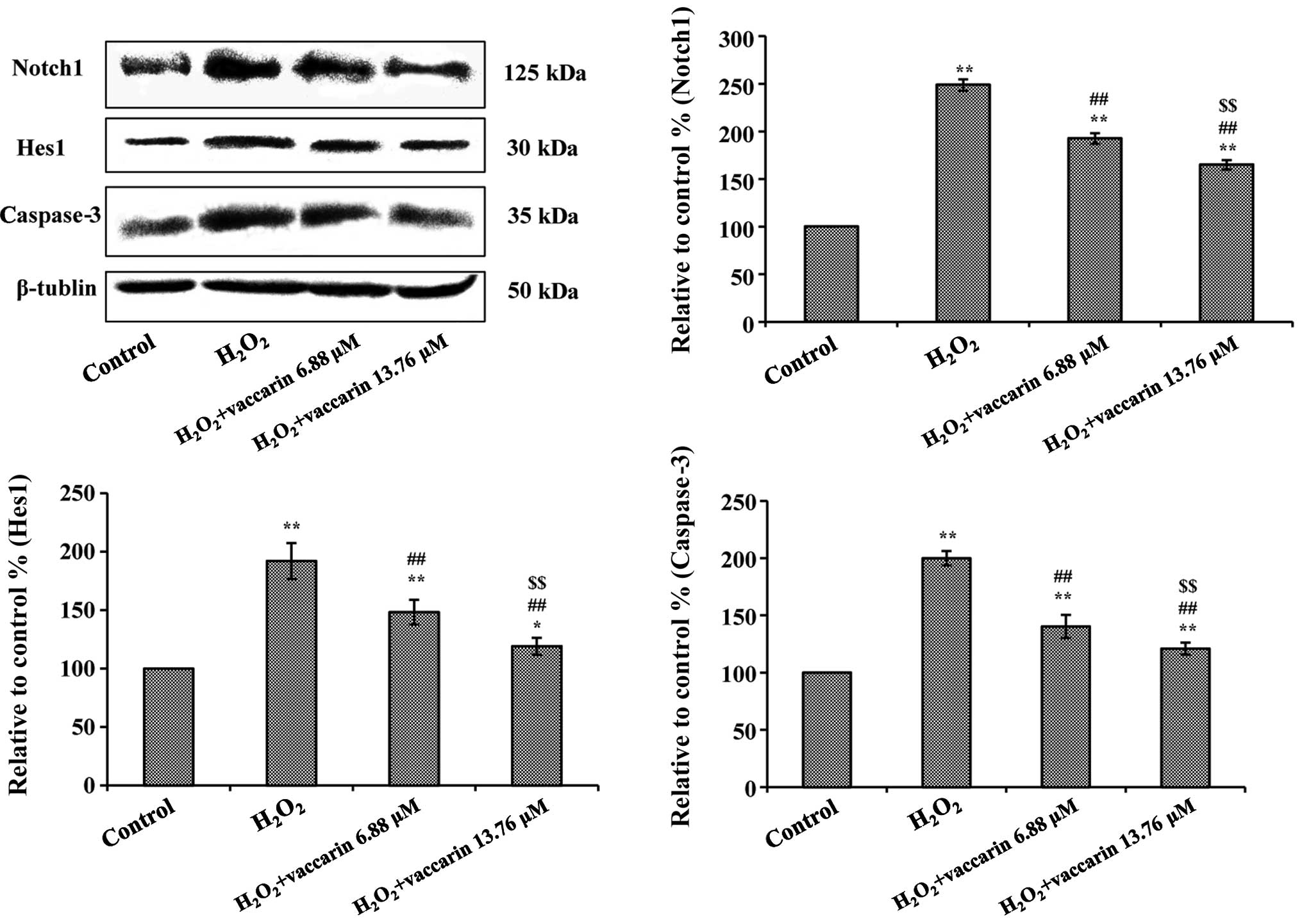
Effects of vaccarin on the expression of
Notch1, Hes1 and caspase-3 in H2O2-injured
EA.hy926 cells
To further investigate the effect and mechanism of
vaccarin in H2O2-injured EA.hy926 cells, the
expression of Notch1, Hes1 and caspase-3 were detected though
western blotting analysis. As demonstrated in Fig. 6, treatment with
H2O2 significantly increased the expression
of Notch1, Hes1 and caspase-3 (% relative to control group,
P<0.01). When the cells were pretreated with vaccarin (6.88 and
13.76 μM), the expression of Notch1, Hes1 and caspase-3 decreased
significantly (P<0.01, compared to the
H2O2 group).
Discussion
Endothelial barrier dysfunction plays a critical
role in the pathogenesis of diabetic vascular complications
(39). Exposure of the vascular
endothelial tissue to H2O2 causes endothelial
dysfunction and further complications of atherosclerosis and
hypertension (40). A previous
study showed that H2O2 induced reactive
oxygen species (ROS), which can cause cellular dysfunction, cell
apoptosis and cell death (41).
Notch signaling has been widely connected in endothelial to
mesenchymal transformation (42),
endothelia cell proliferation (43) and apoptosis control (44). In the cultured renal proximal
tubular cell model, puromycin aminonucleoside induced the
upregulation of the Notch1 signaling components, such as Notch
intracellular domain and the downstream molecule Hes1, and also the
downregulation of Numb, an intrinsic Notch antagonist (45).
The present study has indicated that in human
EA.hy926 endothelial cells, H2O2 caused
vascular endothelial cell apoptosis via activating Notch1 and Hes1.
Vaccarin contributed a protective effect against
H2O2 induced endothelial injury, as shown by
improved cell viability, migratory ability and a decreased
apoptotic index. The protective effects of vaccarin against cell
injury are, in part, dependent on Notch1 inhibition. Numerous
anti-H2O2-induced cell injury drugs protect
against apoptosis by regulation of the cell apoptosis pathway
(46–47). B-cell lymphoma 2, Bax, Bak and
caspase-3 have significant roles in cell apoptosis and are
important members of the cell survival pathway (48–49). Caspase-3 is the main component of
the execution steps of the apoptotic process, and the activation of
caspase-3 is the central link of apoptosis (50). A previous study showed that
caspase-3 may be a main target involved in the ROS-mediated
H2O2-induced apoptosis in human endothelial
cells (51). In the present
study, vaccarin was shown to effectively suppress caspase-3
overexpression induced by H2O2.
H2O2-induced free radicals can
have irreversible effects on a number of biomolecules, including
lipids, leading to lipid peroxidation. LDH leakage, which is
associated with membrane damage, and MDA, a by-product of lipid
peroxidation induced by excessive ROS exposure, are commonly used
biomarkers of oxidative stress injury (37). Antioxidants, such as SOD, are
important in providing protection against
H2O2 injury. Thus, the combined action of SOD
and other endogenous antioxidants can reduce intracellular ROS
(37). In the present study,
significant decreases in SOD were observed in EA.hy926 cells
following H2O2 exposure, indicating the
impairment in antioxidant defenses. In addition, a clear increase
in MDA production was correlated with an increase of LDH release.
Preincubation with vaccarin protected EA.hy926 cells from
H2O2-induced cellular oxidative injury as
shown by inhibition of the levels of LDH and MDA, but increased SOD
activity. Notably, in addition to downregulation of
H2O2-induced Notch signaling, vaccarin
treatment also downregulated H2O2-induced the
apoptotic pathway-related protein, caspase-3. These results
indicate that increased endogenous antioxidant preservation and
attenuation of the cell apoptotic pathway may represent a major
mechanism of cellular protection by vaccarin.
In conclusion, the present study demonstrates that
vaccarin, the major bioactive compound of Vaccariae semen,
can prevent H2O2-induced apoptosis of human
EA.hy926 endothelial cells. Vaccarin was effective on attenuating
cells injury via the inhibition of the Notch signaling pathway.
These findings indicated that vaccarin has anti-apoptotic activity
in the diabetic vascular lesion, which leads to the development of
atherosclerosis or hypertension. The study shows that vaccarin is a
possible therapeutic in the prevention of diabetic vascular lesion
or atherosclerosis.
Acknowledgements
The present study was supported by Fundamental
Research Funds for the Central Universities (grant no.
JUSRP51412B).
References
|
1
|
Beckman JA, Creager MA and Libby P:
Diabetes and atherosclerosis: epidemiology, pathophysiology, and
management. JAMA. 287:2570–2581. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Malakul W, Thirawarapan S, Suvitayavat W
and Woodman OL: Type 1 diabetes and hypercholesterolaemia reveal
the contribution of endothelium-derived hyperpolarizing factor to
endothelium-dependent relaxation of the rat aorta. Clin Exp
Pharmacol Physiol. 35:192–200. 2008.
|
|
3
|
McNulty PH, Tulli MA, Robertson BJ, et al:
Effect of simulated postprandial hyperglycemia on coronary blood
flow in cardiac transplant recipients. Am J Physiol Heart Circ
Physiol. 293:H103–H108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vita JA: Endothelial function and clinical
outcome. Heart. 91:1278–1279. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schulz E, Dopheide J, Schuhmacher S, et
al: Suppression of the JNK pathway by induction of a metabolic
stress response prevents vascular injury and dysfunction.
Circulation. 118:1347–1357. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yada T, Shimokawa H, Hiramatsu O, et al:
Hydrogen peroxide, an endogenous endothelium-derived
hyperpolarizing factor, plays an important role in coronary
autoregulation in vivo. Circulation. 107:1040–1045. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
West XZ, Malinin NL, Merkulova AA, et al:
Oxidative stress induces angiogenesis by activating TLR2 with novel
endogenous ligands. Nature. 467:972–976. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen CA, Wang TY, Varadharaj S, et al:
S-glutathionylation uncouples eNOS and regulates its cellular and
vascular function. Nature. 468:1115–1118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu XG and Li L: Rosiglitazone suppresses
lipopolysaccharide-induced matrix metalloproteinase-2 activity in
rat aortic endothelial cells via Ras-MEK1/2 signaling. Int J
Cardiol. 158:54–58. 2012. View Article : Google Scholar
|
|
10
|
Zanchetti A, Hennig M, Hollweck R, et al:
Baseline values but not treatment-induced changes in carotid
intima-media thickness predict incident cardiovascular events in
treated hypertensive patients: findings in the European Lacidipine
Study on Atherosclerosis (ELSA). Circulation. 120:1084–1090. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang Y, Duan WX, Liang ZX, et al: Curcumin
attenuates endothelial cell oxidative stress injury through Notch
signaling inhibition. Cell Signal. 25:615–629. 2013. View Article : Google Scholar
|
|
12
|
Rosenbaum MA, Miyazaki K and Graham LM:
Hypercholesterolemia and oxidative stress inhibit endothelial cell
healing after arterial injury. J Vasc Surg. 55:489–496. 2012.
View Article : Google Scholar :
|
|
13
|
Cook KM and Figg WD: Angiogenesis
inhibitors: Current strategies and future prospects. CA Cancer J
Clin. 60:222–243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ji X, Wang Z, Geamanu A, Sarkar FH and
Gupta SV: Inhibition of cell growth and induction of apoptosis in
non-small cell lung cancer cells by delta-tocotrienol is associated
with notch-1 down-regulation. J Cell Biochem. 112:2773–2783. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Samon JB, Champhekar A, Minter LM, et al:
Notch1 and TGFb1 cooperatively regulate Foxp3 expression and the
maintenance of peripheral regulatory T cells. Blood. 112:1813–1821.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maciej J and Krzysztof S: Notch: A new
player in MS mechanisms. J Neuroimmunol. 218:3–11. 2010. View Article : Google Scholar
|
|
17
|
Androutsellis-Theotokis A, Leker RR,
Soldner F, et al: Notch signalling regulates stem cell numbers in
vitro and in vivo. Nature. 442:823–826. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McCright B: Notch signaling in kidney
development. Curr Opin Nephrol Hypertens. 12:5–10. 2003. View Article : Google Scholar
|
|
19
|
Archana VB, Karl DP, Pao LC, Yoon YS and
Michael ED: Oxidative stress-induced Notch1 signaling promotes
cardiogenic gene expression in mesenchymal stem cells. Stem Cell
Res Ther. 4:2–15. 2013.
|
|
20
|
Campos J, Schmeda-Hirschmann G, Leiva E,
et al: Lemon grass (Cymbopogon citratus (D.C) Stapf) polyphenols
protect human umbilical vein endothelial cell (HUVECs) from
oxidative damage induced by high glucose, hydrogen peroxide and
oxidised low-density lipoprotein. Food Chem. 151:175–181. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jin RG, Wang QJ and Yan TH: Protective
effects of salidroside on human umbilical vein endothelial cells.
Pharmacology and Clinics of Chinese Materia Medical. 28:41–44.
2012.
|
|
22
|
China Pharmacopoeia Committee. Chinese
Pharmacopoeia: The 2010 edition. China Medical Science Press;
Beijing, China: pp. 49–50. 2010
|
|
23
|
Sang SM, Lao A and Chen ZL: Chemistry and
bioactivity of the seeds of Vaccaria segetalis. Orient Foods Herbs.
21:279–291. 2000.
|
|
24
|
Li F and Liang JY: Research progress of
Vaccaria segetalis. Straits Pharm J. 3:1–5. 2007.
|
|
25
|
Koike K, Jia ZH and Nikaido T:
Triterpenoid saponins from Vaccaria segetalis. Phytochemistry.
7:13431998. View Article : Google Scholar
|
|
26
|
Sang SM, Lao AN and Wang HC: A
phenylpropanoid glycoside from Vaccaria segetalis. Phytochemistry.
48:569–571. 1998. View Article : Google Scholar
|
|
27
|
Sang S, Lao A and Wang H: Triterpenoid
saponin from Vaccaria segetalis. J Asian Nat Prod Res. 1:199–205.
1999. View Article : Google Scholar
|
|
28
|
Yun YS, Morita H, Takeya K, et al: Cyclic
peptides from higher plants. 34. segetalins G and H, structures and
estrogen-like activity of cyclic pentapeptides from Vaccaria
segetalis. J Nat Prod. 60:216–218. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Morita H, Young SY, Takeya K, et al: A
cyclic heptapeptide from Vaccaria segetalis. Phytochemistry.
42:439–441. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dong Z and Chen N: Study on the TLC
identification method of Wangbuliuxing tablets. China For Med
Treat. 30:1282011.
|
|
31
|
Li N, Ma CH, Liu D, et al: Chemical
constituents analysis of fried Vaccaria segetafis. Chin J Exptl
Tradit Med Form. 19:73–75. 2013.
|
|
32
|
Sun YX, Liang HT, Zhang XT, et al:
Structural elucidation and immunological activity of a
polysaccharide from the fruiting body of Armillaria mellea. Biores
Technol. 100:1860–1863. 2009. View Article : Google Scholar
|
|
33
|
Sun Y, Wang S, Li T, et al: Purification
structure and immunobiological activity of a new water-soluble
polysaccharide from the mycelium of Polyporus albicans (Imaz) Teng.
Biores Technol. 99:900–904. 2008. View Article : Google Scholar
|
|
34
|
Meng H, Chen Y, Qin W, et al:
Determination of vaccarin in Vaccariae Semen by HPLC. Zhongguo
Zhong Yao Za Zhi. 35:2072–2074. 2010.PubMed/NCBI
|
|
35
|
Xia MX, Feng L and Zhang LF: Isolation,
Purification and Identification of Chemical Compound from Semen
vaccariae to Inhibit Endothelial Cells. Biotechnology Bulletin.
2:93–97. 2009.
|
|
36
|
Cheong SM, Choi H, Hong BS, Gho YS and Han
JK: Dab2 is pivotal for endothelial cell migration by mediating
VEGF expression in cancer cells. Exp Cell Res. 318:550–557. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu HT, Li WM, Xu G, et al: Chitosan
oligosaccharides attenuate hydrogen peroxide-induced stress injury
in human umbilical vein endothelial cells. Pharmacol Res.
59:167–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li B, Qiu T, Zhang P, Wang X, Yin Y and Li
S: IKVAV regulates ERK1/2 and Akt signalling pathways in BMMSC
population growth and proliferation. Cell Prolif. 47:133–145. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liang KW, Lee WJ, Lee WL, Chen YT, Ting CT
and Sheu WH: Diabetes exacerbates angiographic coronary lesion
progression in subjects with metabolic syndrome independent of CRP
levels. Clin Chim Acta. 388:41–45. 2008. View Article : Google Scholar
|
|
40
|
Dikalov SI, Dikalova AE, Bikineyeva AT,
Schmidt HH, Harrison DG and Griendling KK: Distinct roles of Nox1
and Nox4 in basal and angiotensin II-stimulated superoxide and
hydrogen peroxide production. Free Radical Biol Med. 45:1340–1351.
2008. View Article : Google Scholar
|
|
41
|
Li JK, Ge R, Tang L and Li QS: Protective
effects of farrerol against hydrogen-peroxide-induced apoptosis in
human endothelium-derived EA.hy926 cells. Can J Physiol Pharmacol.
91:733–740. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Noseda M, McLean G, Niessen K, et al:
Notch activation results in phenotypic and functional changes
consistent with endothelial-to-mesenchymal transformation. Circ
Res. 94:910–917. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Williams CK, Li JL, Murga M, Harris AL and
Tosato G: Up-regulation of the Notch ligand Delta-like 4 inhibits
VEGF-induced endothelial cell function. Blood. 107:931–939. 2006.
View Article : Google Scholar
|
|
44
|
MacKenzie F, Duriez P, Wong F, Noseda M
and Karsan A: Notch4 inhibits endothelial apoptosis via
RBP-Jkappa-dependent and -independent pathways. J Biol Chem.
279:11657–11663. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ding X, Zhu F, Li T, Zhou Q, Hou FF and
Nie J: Numb Protects Renal Proximal Tubular Cells from Puromycin
Aminonucleoside-Induced Apoptosis through Inhibiting Notch
Signaling Pathway. Int J Biol Sci. 7:269–278. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Stampfer MJ, Hennekens CH, Manson JE,
Colditz GA, Rosner B and Willett WC: Vitamin E consumption and the
risk of coronary disease in women. N Engl J Med. 328:1444–1449.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rimm EB, Stampfer MJ, Ascherio A,
Giovannucci E, Colditz GA and Willett WC: Vitamin E consumption and
the risk of coronary heart disease in men. N Engl J Med.
328:1450–1456. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen SD, Yin JH, Hwang CS, Tang CM and
Yang DI: Anti-apoptotic and anti-oxidative mechanisms of
minocycline against sphingomyelinase/ceramide neurotoxicity:
implication in Alzheimer’s disease and cerebral ischemia. Free
Radic Res. 46:940–950. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Luo P, Chen T, Zhao Y, et al: Protective
effect of Homer 1a against hydrogen peroxide-induced oxidative
stress in PC12 cells. Free Radic Res. 46:766–776. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fan G, Ma X, Wong PY, Rodrigues CM and
Steer CJ: p53 dephosphorylation and p21 (Cip1/Waf1) translocation
correlate with caspase-3 activation in TGF-beta1-induced apoptosis
of HuH-7 cells. Apoptosis. 9:211–221. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Duan W, Yang Y and Yi W: New Role of
JAK2/STAT3 signaling in endothelial cell oxidative stress injury
and protective effect of melatonin. PLoS One. 8:1–13. 2013.
View Article : Google Scholar
|