Mature miRNAs are small non-coding RNAs that consist
of 19-23 nucleotides. The first miRNAs were characterized in C.
elegans in 1993 (1).
Subsequently, an increasing number of miRNAs were identified in
plants and animals. To date, over 1,000 miRNAs have been identified
in humans, which regulate approximately 60% of mammalian gene
expression (2,3). miRNA genes are transcribed as
primary miRNAs by RNA polymerase II (4). Following transcription, the
precursor miRNA (stem-loop with approximately 80 nucleotides) is
generated by Drosha (5).
Subsequently, the precursor miRNA is exported from the nucleus to
the cytoplasm and digested into a mature miRNA by Dicer, an RNA
polymerase III enzyme (6). The
main function of miRNAs is to regulate gene expression at the
translational level. miRNAs can bind to the 3′-untranslated region
(3′-UTR) of a target mRNA and suppress its translation (1). Recent studies have indicated that
miRNAs play an essential role in a variety of diseases, including
cancer (7), type I (8) and type II diabetes (T2D) (9), autoimmunity disease (10), and cardiovascular diseases
(11). Thus, miRNAs have a
potential effect on clinical diagnosis, prognosis and therapy.
Burn injury is a complex trauma which is caused by
factors such as heat, electricity, chemicals and radiation
(12). Inflammation is one of the
host responses to injury, and following burn injury, the levels of
inflammatory mediators, such as tumor necrosis factor α (TNF-α)
(13), transformation growth
factor β (TGF-β) (14),
interleukin (IL)-2 and IL-6 (15), are markedly increased. Burn injury
also leads to cardiovascular damage and enhances vascular
permeability, which results in the loss of body fluid (16). Clinical studies have demonstrated
that burn injury may induce insulin resistance and affect glucose
and fat metabolism (17,18). Insulin resistance contributes to
the attenuation of wound healing, which enhances the risk of
infection (19). As previously
demonstrated, a topical insulin injection can accelerate wound
healing in diabetes through the activation of the Akt and Erk
signaling pathways (20).
Previous studies have indicated that miRNAs play a
critical role in regulating insulin resistance induced by
mitochondrial dysfunction or diabetes through the inhibition of
insulin receptor substrate 1 (IRS1) protein translation (21,22). The suppression of the insulin
receptor (IR)/IRS1 signaling pathway is the key mechanism
responsible for burn-induced insulin resistance; therefore, miRNAs
may also be implicated in burn-induced insulin resistance. The
identification of the specific miRNAs which are involved in
burn-induced insulin resistance may lead to the development of
novel therapeutic targets for clinical therapy.
A previous study using a rat model demonstrated that
no difference in insulin secretion was detected between the sham
and burn groups; however, the sensitivity of insulin was
significantly suppressed in the burn group (23). Insulin resistance and
hyperglycemia are crucial risk factors for increased mortality in
patients with severe burn injuries (24). In another study on burned
children, it was demonstrated that insulin resistance can last up
to 3 years (25). Intensive
insulin therapy is an efficient manner to control the blood glucose
of severely burned patients, as it can decrease the risk of
infection and sepsis, improve hepatic and rental function, and
suppress acute inflammation (24).
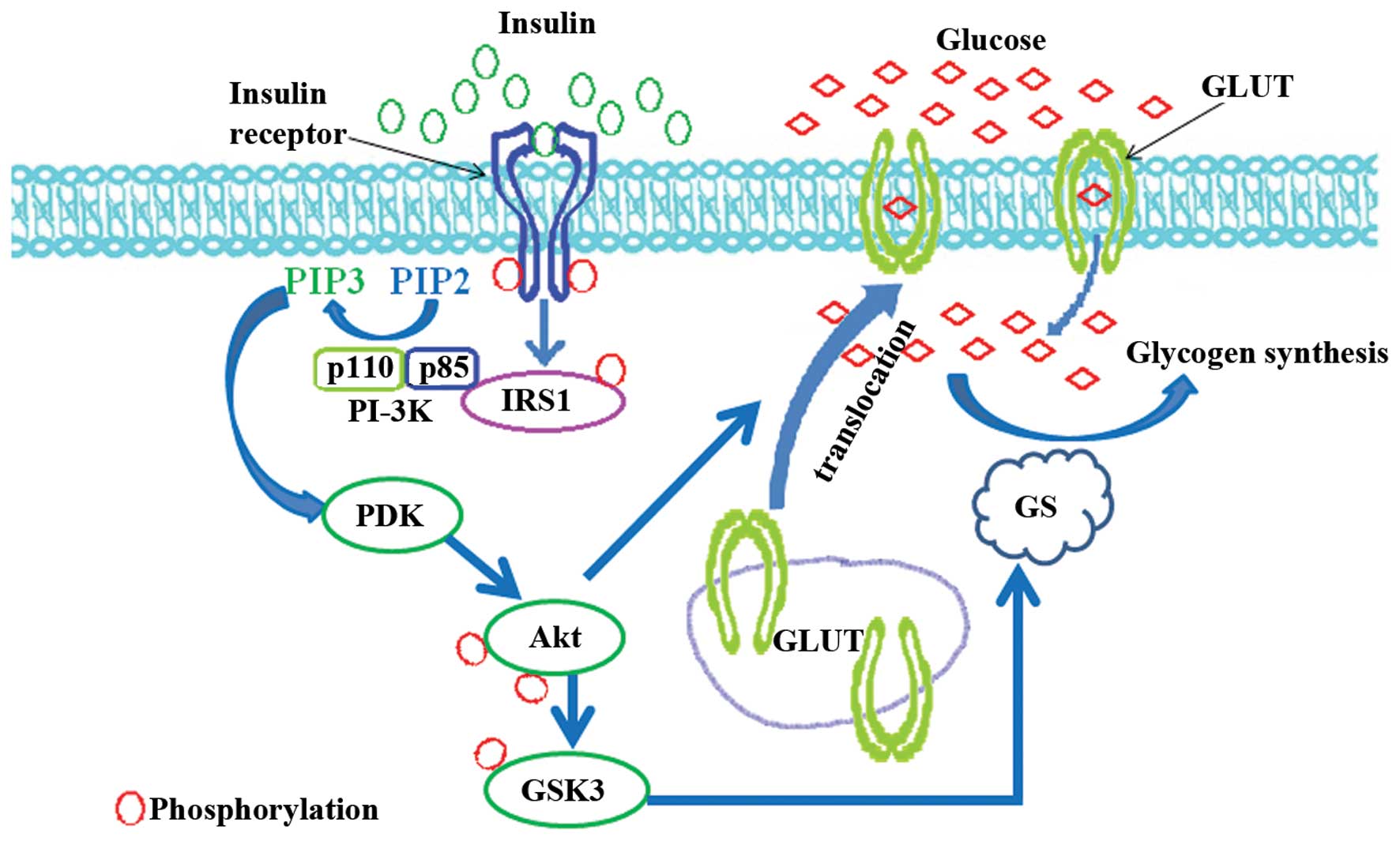
Glucose is one of the most important energy sources
for the human organism. It is usually stored in the liver and
muscle cells in the form of glycogen (26). After eating, elevated blood
glucose is converted into glycogen (27). During the conversion, blood
glucose is firstly transported into cells by the plasma membrane
protein glucose transporter (GLUT), and its four isoforms, GLUT1,
GLUT2, GLUT3 and GLUT4, have been well-characterized (28). GLUT4 is primarily expressed in
muscle and fat cells. The insulin signaling pathway plays an
important role in regulating its translocation (Fig. 1). Insulin binds to the IR and
induces the autophosphorylation of the receptor at tyrosine
residues (29). Following
autophosphorylation, the receptor further recruits the IRS and
promotes its phosphorylation at tyrosine residues (30). Phosphorylated IRS subsequently
binds to the regulatory subunit, p85, of the phosphoinositide-3
kinase (PI3K) and activates its catalytic subunit p110, which is
responsible for stimulating the phosphoinositide-dependent kinase
(PDK) (31). As the upstream
kinase of Akt, activated PDK promotes the phosphorylation of Akt at
Thr308 and Ser473 (32), and
phosphorylated Akt mediates the translocation of GLUT from the
cytoplasm to the membrane (33)
(Fig. 1). Apart from its role in
GLUT translocation, Akt has also been implicated in regulating
glycogen synthesis. Glycogen synthase (GS) is a key enzyme involved
in converting glucose into glycogen, and there are two isoforms in
mammals, the muscle isoform (34)
and the liver isoform (35). Both
isoforms are inactivated due to phosphorylation at the NH2- or
COOH-terminal residues mediated by glycogen synthesis kinase 3
(GSK3) (36). Insulin
dephosphorylates and restores the function of GS through Akt- or
protein kinase A-mediated phoshorylation and the inactivation of
GSK3 (37,38). In skeletal muscle, insulin
enhances glycogen synthesis in the absence of GSK3 phosphorylation
(39). Glucose-6 phosphate
induces glycogen synthesis through the activation of GS in a cyclic
AMP-stimulated protein kinase-dependent manner (40).
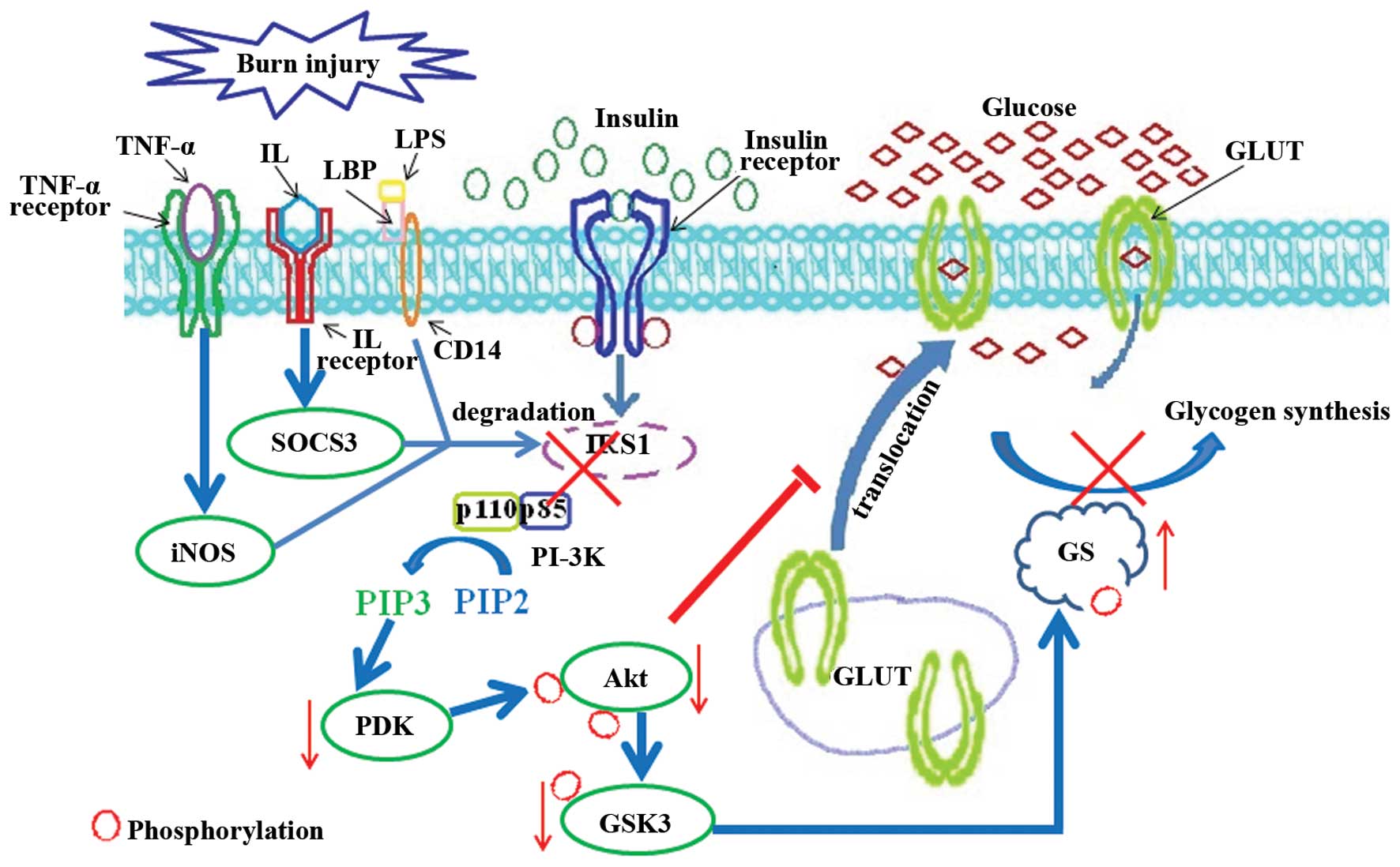
After a burn injury is sustained, the activation of
the insulin signaling pathway is significantly suppressed, and
blood glucose levels are markedly increased (41). The results from experiments
carried out in our, as well as other laboratories have indicated
that the levels of lipopolysaccharides (LPS), TNF-α and
interleukins are increased following burn injury (13,15). These factors are involved in the
regulation of the phosphorylation and degradation of IRS1 which, in
turn, results in insulin resistance (Fig. 2). LPS, the stimulator of inducible
nitric oxide synthase (iNOS), plays an essential role in inducing
hyperglycemia and insulin resistance, which can be restored by the
iNOS inhibitor (42,43). iNOS also enhances the
ubiquitination and degradation of IRS1 (44). The deficiency of iNOS attenuates
the burn-induced skeletal muscle insulin resistance (45). Glucose uptake is decreased
following exposure to TNF-α (46). TNF-α inhibits the
autophosphorylation of IR and its substrate IRS1 tyrosine
phosphorylation (46). It can
also suppress the function of IRS1 by triggering phosphorylation at
Ser307 which induces the degradation of IRS1 (47–49). IL-6 has been shown to induce
insulin resistance in HepG2 cells (50) and 3T3-L1 adipocytes (51), which can promote IRS1 degradation
by upregulating the expression of the suppressor of cytokine
signaling 3 (SOCS3) (52). IL-1β
induces adipocyte insulin resistance through the downregulation of
IRS1 (53). Due to the
burn-induced IRS1 degradation, the interaction between IRS1 and
PI3K is repressed, and insulin stimulates PKB/Akt activation, which
is impaired after burn injury (54). Subsequently, the phosphorylation
of GSK3β at Ser9 mediated by activated Akt is decreased; the
enhanced activity of GSK3β has been detected in the skeletal muscle
of rats following burn injury (55). Thus, the phosphorylation of GS
mediated by GSK3 is augmented, and the conversion of glucose into
glycogen is significantly attenuated (Fig. 2). Taken together, these data
indicate that the TNF-α-, LPS- or IL-induced IRS1 protein
degradation is involved in promoting burn-induced insulin
resistance and hyperglycemia.
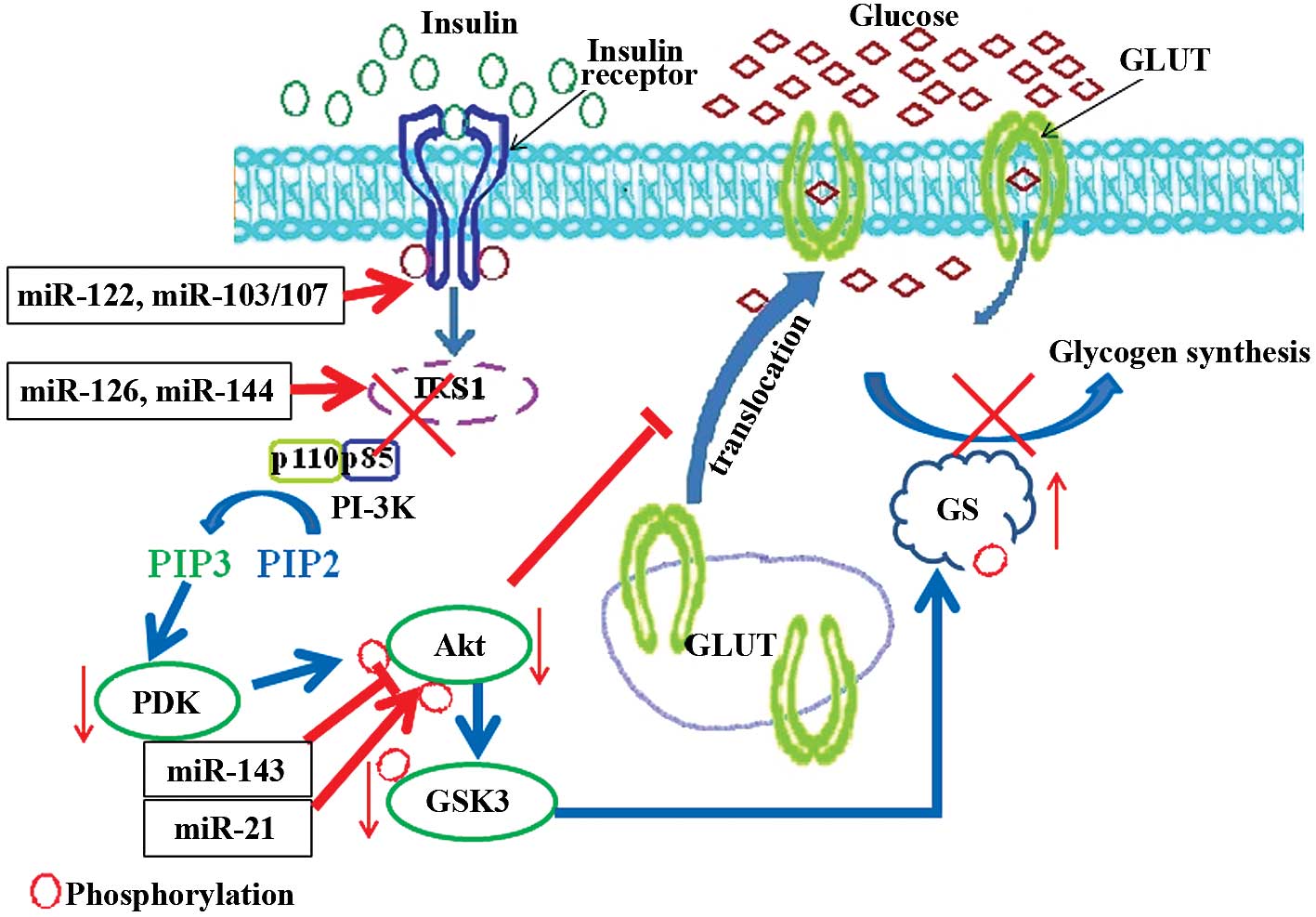
As small non-coding RNAs, miRNAs play a pivotal role
in post-transcriptional regulation. miRNAs can bind to and promote
the deadenylation and degradation of target mRNAs (56,57). Translational repression is another
important function of miRNAs. It can directly bind to the 3′-UTR of
target mRNAs and inhibit the translational initiation (58). Studies have demonstrated that
miRNAs are involved in the regulation of multiple insulin
resistance-induced diseases (Fig.
3). In T2D, miR-144 has been shown to promote insulin
resistance by directly targeting IRS1 mRNA (22). The suppression of IRS1 mediated by
miR-126 has also been shown to result in mitochondrial dysfunction
and insulin resistance (21). The
sitmulation of Akt activation by insulin is critical for
glycometabolism, and the obesity-induced miR-143 overexpression has
been shown to lead to hyperglycemia by inactivating the Akt
signaling pathway (59). A study
using let-7 family transgenic mice demonstrated that let-7
overexpression may contribute to the development of T2D (60). Protein tyrosine phosphatase 1B
(PTP1B) impairs the insulin signaling pathway through the
dephosphorylation of IR at tyrosine residues. The 3′-UTR of
PTP1B mRNA is the target of miR-122, and decreased miR-122
expression has been shown to result in hepatic insulin resistance
(61). In mouse models of
obesity, miR-103/107 is upregulated. The blockage of miR-103/107
has been shown to promote insulin sensitivity by elevating
caveolin-1-mediated IR activation (62). Phosphatase and tensin homolog
deleted on chromosome 10 (PTEN), the direct target of miR-21
(63), is the key phosphotase of
Akt which can negatively regulate the Akt signaling pathway. In
insulin-resistant adipocytes, the suppressed expression of miR-21
and impaired Akt signaling pathway has been observed (64). The transport of glucose is also
regulated by miRNAs, and the transmembrane protein GLUTs play an
essential role in glucose transport. Elevated miR-133 levels have
been shown to reduce the insulin-stimulated glucose uptake by
downregulating GLUT4 expression (65). In cardiomyocytes, miR-223 has been
shown to promote GLUT4 expression and increase glucose uptake
(66). Insulin resistance is the
intrinsic complication of polycystic ovary syndrome (PCOS), and
overexpressed miR-93 in patients with PCOS binds to the 3′-UTR of
GLUT4 mRNA and reduces its protein translation (67). Microarray analysis has further
indicated that the expression of several miRNAs is altered
following burn injury. In comparison with normal skin tissue, a
total of 32 upregluated and 34 downregulated miRNAs were identified
in the skin tissue of patients who sustained burn injuries
(68). The expression levels of
miR-144 in the skin tissue of the burned patients, which can
directly target the 3′-UTR of IRS1 mRNA (22), were 16-fold higher than those in
normal skin tissue (68). This
suggests that miRNAs, such as miR-144, play an essential role in
promoting burn-induced insulin resistance by suppressing the
activation of the IR/IRS signaling pathway.
Although miRNAs were only discovered 20 years ago,
their molecular mechanisms of action involving the repression of
target gene expression have been elucidated. Moreover, evidence
indicates that miRNAs are associated with the development of a
number of human diseases. The process from the time of discovery of
an miRNA to the development of clinical therapeutic drug targets is
rapidly approaching. miR-34a has been shown to suppress the
development of prostate and lung cancer (69,70), and the use of a miR-34a mimic is
currently in the developmental stage for cancer therapy (www.iptonline.com, The Therapeutic Potential of
microRNAs). Another miRNA, miR-208, which is involved in promoting
chronic heart failure (71), is
also undergoing investigation in preclinical trials (www.iptonline.com, The Therapeutic Potential of
microRNAs). Its antagonist is expected to be used in the therapy of
heart disease. miR-122 has been shown to be associated with
hepatitis C virus infection (72), and its antagonist, miravirsen, has
undergone phase II clinical trials for the therapy of patients with
hepatitis C virus (73).
Miravirsen may be the first miRNA-related drug for clinical
therapy. Taken together, the mimics or antagonists of miRNAs are
expected to be widely used in clinical therapy.
In the response to burn injury, the expression of
several miRNAs, including insulin resistance-associated miRNAs, is
altered. This change may play a pivotal role in mediating
burn-induced insulin resistance, which results in hyperglycemia and
reduces wound healing. Further research focusing on the involvement
of miRNAs in the regulation of burn-induced insulin resistance may
lead to the development of novel therapeutic targets for the
treatment of burn injuries.
The present study was supported by grants from the
National Science Foundation of China (NSFC81120108014 and
NSFC81471873), the Beijing Natural Science Foundation (7144250) and
the China Postdoctoral Science Foundation (2013M532200).
|
1
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bentwich I, Avniel A, Karov Y, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee Y, Kim M, Han J, et al: MicroRNA genes
are transcribed by RNA polymerase II. EMBO J. 23:4051–4060. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zeng Y and Cullen BR: Efficient processing
of primary microRNA hairpins by Drosha requires flanking
nonstructured RNA sequences. J Biol Chem. 280:27595–27603. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Andl T, Murchison EP, Liu F, et al: The
miRNA-processing enzyme dicer is essential for the morphogenesis
and maintenance of hair follicles. Curr Biol. 16:1041–1049. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Visone R and Croce CM: MiRNAs and cancer.
Am J Pathol. 174:1131–1138. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Argyropoulos C, Wang K, McClarty S, et al:
Urinary microRNA profiling in the nephropathy of type 1 diabetes.
PLoS One. 8:e546622013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van de Bunt M, Gaulton KJ, Parts L, et al:
The miRNA profile of human pancreatic islets and beta-cells and
relationship to type 2 diabetes pathogenesis. PLoS One.
8:e552722013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McMichael AJ, Borrow P, Tomaras GD,
Goonetilleke N and Haynes BF: The immune response during acute
HIV-1 infection: clues for vaccine development. Nat Rev Immunol.
10:11–23. 2010. View
Article : Google Scholar
|
|
11
|
Ono K, Kuwabara Y and Han J: MicroRNAs and
cardiovascular diseases. FEBS J. 278:1619–1633. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Evers LH, Bhavsar D and Mailander P: The
biology of burn injury. Exp Dermatol. 19:777–783. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duan H, Chai J, Sheng Z, et al: Effect of
burn injury on apoptosis and expression of apoptosis-related
genes/proteins in skeletal muscles of rats. Apoptosis. 14:52–65.
2009. View Article : Google Scholar
|
|
14
|
Nishimura T, Nishiura T, deSerres S,
Nakagawa T, Brenner DA and Meyer AA: Impact of burn injury on
hepatic TGF-beta1 expression and plasma TGF-beta1 levels. J Trauma.
48:39–44. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kowal-Vern A, Walenga JM, Hoppensteadt D,
Sharp-Pucci M and Gamelli RL: Interleukin-2 and interleukin-6 in
relation to burn wound size in the acute phase of thermal injury. J
Am Coll Surg. 178:357–362. 1994.PubMed/NCBI
|
|
16
|
Soejima K, Traber LD, Schmalstieg FC, et
al: Role of nitric oxide in vascular permeability after combined
burns and smoke inhalation injury. Am J Respir Crit Care Med.
163:745–752. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cree MG, Zwetsloot JJ, Herndon DN, et al:
Insulin sensitivity and mitochondrial function are improved in
children with burn injury during a randomized controlled trial of
fenofibrate. Ann Surg. 245:214–221. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cree MG, Aarsland A, Herndon DN and Wolfe
RR: Role of fat metabolism in burn trauma-induced skeletal muscle
insulin resistance. Crit Care Med. 35:S476–S483. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sharp A and Clark J: Diabetes and its
effects on wound healing. Nurs Stand. 25:41–47. 2010. View Article : Google Scholar
|
|
20
|
Lima MH, Caricilli AM, de Abreu LL, et al:
Topical insulin accelerates wound healing in diabetes by enhancing
the AKT and ERK pathways: a double-blind placebo-controlled
clinical trial. PLoS One. 7:e369742012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ryu HS, Park SY, Ma D, Zhang J and Lee W:
The induction of microRNA targeting IRS-1 is involved in the
development of insulin resistance under conditions of mitochondrial
dysfunction in hepatocytes. PLoS One. 6:e173432011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Karolina DS, Armugam A, Tavintharan S, et
al: MicroRNA 144 impairs insulin signaling by inhibiting the
expression of insulin receptor substrate 1 in type 2 diabetes
mellitus. PLoS One. 6:e228392011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Frayn KN: Effects of burn injury on
insulin secretion and on sensitivity to insulin in the rat in vivo.
Eur J Clin Invest. 5:331–337. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jeschke MG, Kulp GA, Kraft R, et al:
Intensive insulin therapy in severely burned pediatric patients: a
prospective randomized trial. Am J Respir Crit Care Med.
182:351–359. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gauglitz GG, Herndon DN, Kulp GA, Meyer WJ
III and Jeschke MG: Abnormal insulin sensitivity persists up to
three years in pediatric patients post-burn. J Clin Endocrinol
Metab. 94:1656–1664. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kokubun E, Hirabara SM, Fiamoncini J, Curi
R and Haebisch H: Changes of glycogen content in liver, skeletal
muscle, and heart from fasted rats. Cell Biochem Funct. 27:488–495.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chari-Bitron A, Lepkovsky S, Lemmon RM and
Dimick MK: Conversion of glucose to glycogen after ingestion of a
high-carbohydrate diet. Am J Physiol. 198:787–792. 1960.PubMed/NCBI
|
|
28
|
Bell GI, Kayano T, Buse JB, et al:
Molecular biology of mammalian glucose transporters. Diabetes Care.
13:198–208. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kasuga M, Zick Y, Blithe DL, Crettaz M and
Kahn CR: Insulin stimulates tyrosine phosphorylation of the insulin
receptor in a cell-free system. Nature. 298:667–669. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Boura-Halfon S and Zick Y: Phosphorylation
of IRS proteins, insulin action, and insulin resistance. Am J
Physiol Endocrinol Metab. 296:E581–E591. 2009. View Article : Google Scholar
|
|
31
|
Metz HE and Houghton AM: Insulin receptor
substrate regulation of phosphoinositide 3-kinase. Clin Cancer Res.
17:206–211. 2011. View Article : Google Scholar
|
|
32
|
Bhaskar PT and Hay N: The two TORCs and
Akt. Dev Cell. 12:487–502. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thorell A, Hirshman MF, Nygren J, et al:
Exercise and insulin cause GLUT-4 translocation in human skeletal
muscle. Am J Physiol. 277:E733–E741. 1999.PubMed/NCBI
|
|
34
|
Browner MF, Nakano K, Bang AG and
Fletterick RJ: Human muscle glycogen synthase cDNA sequence: a
negatively charged protein with an asymmetric charge distribution.
Proc Natl Acad Sci USA. 86:1443–1447. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bai G, Zhang ZJ, Werner R, Nuttall FQ, Tan
AW and Lee EY: The primary structure of rat liver glycogen synthase
deduced by cDNA cloning. Absence of phosphorylation sites 1a and
1b. J Biol Chem. 265:7843–7848. 1990.PubMed/NCBI
|
|
36
|
Hojlund K, Birk JB, Klein DK, et al:
Dysregulation of glycogen synthase COOH- and
NH2-terminal phosphorylation by insulin in obesity and
type 2 diabetes mellitus. J Clin Endocrinol Metab. 94:4547–4556.
2009. View Article : Google Scholar
|
|
37
|
Cross DA, Alessi DR, Cohen P, Andjelkovich
M and Hemmings BA: Inhibition of glycogen synthase kinase-3 by
insulin mediated by protein kinase B. Nature. 378:785–789. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fang X, Yu SX, Lu Y, Bast RC Jr, Woodgett
JR and Mills GB: Phosphorylation and inactivation of glycogen
synthase kinase 3 by protein kinase A. Proc Natl Acad Sci USA.
97:11960–11965. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bouskila M, Hirshman MF, Jensen J,
Goodyear LJ and Sakamoto K: Insulin promotes glycogen synthesis in
the absence of GSK3 phosphorylation in skeletal muscle. Am J
Physiol Endocrinol Metab. 294:E28–E35. 2008. View Article : Google Scholar
|
|
40
|
Villar-Palasi C and Guinovart JJ: The role
of glucose 6-phosphate in the control of glycogen synthase. FASEB
J. 11:544–558. 1997.PubMed/NCBI
|
|
41
|
Ikezu T, Okamoto T, Yonezawa K, Tompkins
RG and Martyn JA: Analysis of thermal injury-induced insulin
resistance in rodents. Implication of postreceptor mechanisms. J
Biol Chem. 272:25289–25295. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pilon G, Charbonneau A, White PJ, et al:
Endotoxin mediated-iNOS induction causes insulin resistance via
ONOO− induced tyrosine nitration of IRS-1 in skeletal
muscle. PLoS One. 5:e159122010. View Article : Google Scholar
|
|
43
|
Sugita H, Kaneki M, Tokunaga E, et al:
Inducible nitric oxide synthase plays a role in LPS-induced
hyperglycemia and insulin resistance. Am J Physiol Endocrinol
Metab. 282:E386–E394. 2002.PubMed/NCBI
|
|
44
|
Sugita H, Fujimoto M, Yasukawa T, et al:
Inducible nitric-oxide synthase and NO donor induce insulin
receptor substrate-1 degradation in skeletal muscle cells. J Biol
Chem. 280:14203–14211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sugita M, Sugita H, Kim M, et al:
Inducible nitric oxide synthase deficiency ameliorates skeletal
muscle insulin resistance but does not alter unexpected lower blood
glucose levels after burn injury in C57BL/6 mice. Metabolism.
61:127–136. 2012. View Article : Google Scholar :
|
|
46
|
Hotamisligil GS, Murray DL, Choy LN and
Spiegelman BM: Tumor necrosis factor alpha inhibits signaling from
the insulin receptor. Proc Natl Acad Sci USA. 91:4854–4858. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pederson TM, Kramer DL and Rondinone CM:
Serine/threonine phosphorylation of IRS-1 triggers its degradation:
possible regulation by tyrosine phosphorylation. Diabetes.
50:24–31. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang Q, Carter EA, Ma BY, White M,
Fischman AJ and Tompkins RG: Molecular mechanism(s) of burn-induced
insulin resistance in murine skeletal muscle: role of IRS
phosphorylation. Life Sci. 77:3068–3077. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rui L, Aguirre V, Kim JK, et al:
Insulin/IGF-1 and TNF-α stimulate phosphorylation of IRS-1 at
inhibitory Ser307 via distinct pathways. J Clin Invest.
107:181–189. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Senn JJ, Klover PJ, Nowak IA and Mooney
RA: Interleukin-6 induces cellular insulin resistance in
hepatocytes. Diabetes. 51:3391–3399. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Rotter V, Nagaev I and Smith U:
Interleukin-6 (IL-6) induces insulin resistance in 3T3-L1
adipocytes and is, like IL-8 and tumor necrosis factor-α,
overexpressed in human fat cells from insulin-resistant subjects. J
Biol Chem. 278:45777–45784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang L, Du J, Hu Z, et al: IL-6 and serum
amyloid A synergy mediates angiotensin II-induced muscle wasting. J
Am Soc Nephrol. 20:604–612. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jager J, Gremeaux T, Cormont M, Le
Marchand-Brustel Y and Tanti JF: Interleukin-1beta-induced insulin
resistance in adipocytes through down-regulation of insulin
receptor substrate-1 expression. Endocrinology. 148:241–251. 2007.
View Article : Google Scholar :
|
|
54
|
Sugita H, Kaneki M, Sugita M, Yasukawa T,
Yasuhara S and Martyn JA: Burn injury impairs insulin-stimulated
Akt/PKB activation in skeletal muscle. Am J Physiol Endocrinol
Metab. 288:E585–E591. 2005. View Article : Google Scholar
|
|
55
|
Fang CH, Li B, James JH, et al: GSK-3β
activity is increased in skeletal muscle after burn injury in rats.
Am J Physiol Regul Integr Comp Physiol. 293:R1545–R1551. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Beilharz TH, Humphreys DT, Clancy JL, et
al: microRNA-mediated messenger RNA deadenylation contributes to
translational repression in mammalian cells. PLoS One. 4:e67832009.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Behm-Ansmant I, Rehwinkel J, Doerks T,
Stark A, Bork P and Izaurralde E: mRNA degradation by miRNAs and
GW182 requires both CCR4:NOT deadenylase and DCP1:DCP2 decapping
complexes. Genes Dev. 20:1885–1898. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Mathonnet G, Fabian MR, Svitkin YV, et al:
MicroRNA inhibition of translation initiation in vitro by targeting
the cap-binding complex eIF4F. Science. 317:1764–1767. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Jordan SD, Kruger M, Willmes DM, et al:
Obesity-induced overexpression of miRNA-143 inhibits
insulin-stimulated AKT activation and impairs glucose metabolism.
Nat Cell Biol. 13:434–446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Frost RJ and Olson EN: Control of glucose
homeostasis and insulin sensitivity by the Let-7 family of
microRNAs. Proc Natl Acad Sci USA. 108:21075–21080. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yang YM, Seo SY, Kim TH and Kim SG:
Decrease of microRNA-122 causes hepatic insulin resistance by
inducing protein tyrosine phosphatase 1B, which is reversed by
licorice flavonoid. Hepatology. 56:2209–2220. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Trajkovski M, Hausser J, Soutschek J, et
al: MicroRNAs 103 and 107 regulate insulin sensitivity. Nature.
474:649–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang JG, Wang JJ, Zhao F, Liu Q, Jiang K
and Yang GH: MicroRNA-21 (miR-21) represses tumor suppressor PTEN
and promotes growth and invasion in non-small cell lung cancer
(NSCLC). Clin Chim Acta. 411:846–852. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ling HY, Hu B, Hu XB, et al: MiRNA-21
reverses high glucose and high insulin induced insulin resistance
in 3T3-L1 adipocytes through targeting phosphatase and tensin
homologue. Exp Clin Endocrinol Diabetes. 120:553–559. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Horie T, Ono K, Nishi H, et al:
MicroRNA-133 regulates the expression of GLUT4 by targeting KLF15
and is involved in metabolic control in cardiac myocytes. Biochem
Biophys Res Commun. 389:315–320. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Lu H, Buchan RJ and Cook SA: MicroRNA-223
regulates Glut4 expression and cardiomyocyte glucose metabolism.
Cardiovasc Res. 86:410–420. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen YH, Heneidi S, Lee JM, et al:
miRNA-93 inhibits GLUT4 and is overexpressed in adipose tissue of
polycystic ovary syndrome patients and women with insulin
resistance. Diabetes. 62:2278–2286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Liang P, Lv C, Jiang B, et al: MicroRNA
profiling in denatured dermis of deep burn patients. Burns.
38:534–540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu C, Kelnar K, Liu B, et al: The
microRNA miR-34a inhibits prostate cancer stem cells and metastasis
by directly repressing CD44. Nat Med. 17:211–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wiggins JF, Ruffino L, Kelnar K, et al:
Development of a lung cancer therapeutic based on the tumor
suppressor microRNA-34. Cancer Res. 70:5923–5930. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Callis TE, Pandya K, Seok HY, et al:
MicroRNA-208a is a regulator of cardiac hypertrophy and conduction
in mice. J Clin Invest. 119:2772–2786. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Henke JI, Goergen D, Zheng J, et al:
microRNA-122 stimulates translation of hepatitis C virus RNA. EMBO
J. 27:3300–3310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Janssen HL, Reesink HW, Lawitz EJ, et al:
Treatment of HCV infection by targeting microRNA. N Engl J Med.
368:1685–1694. 2013. View Article : Google Scholar : PubMed/NCBI
|