Introduction
Congenital heart disease (CHD) is a multifactorial
disease; genetic and environmental factors play a key role in the
occurrence of this disease. It is considered that there are over
1,700 genes that are required for heart development. The
genetically modified mouse is a good anatomical model for common
cardiac malformations which is powerful experimental tool for
understanding human CHD (1). The
GATA4 gene is essential to normal heart development.
GATA4 deficiency is embryonically lethal in mice, as this
leads to severe ventricular developmental defects, resulting in
cardiac malformation (2).
GATA4 also regulates the expression of cardiac structural
genes, including α-myosin heavy chain (α-MHC), cardiac
troponin C (cTNC), atrial natriuretic factor (ANF),
brain natriuretic peptide (BNF), A1 adenosine receptor
(ADORA1), muscarinic acetylcholine receptor M2
(CHRM2, cholinergic receptor, muscarinic 2), and angiotensin
II 1A receptors (AngII1a) (3,4).
When GATA4 zinc finger structures and other cardiac-specific
transcription factors [NK2 homeobox 5 (NKX2.5), T-box transcription
factor (TBX5) and myocyte-specific enhancer factor 2 (MEF2C)]
interact to form complexes, these complexes play a role in
transcriptional regulation and regulate heart development (5–8).
The GATA4 gene is an important factor in the
cardiac gene network and mutations in GATA4 have been
confirmed to be associated with the occurrence of atrial septal
defect (ASD). GATA4 has been used to distinguish patients
with familial CHD from those with sporadic CHD (9–11).
GATA4 transgenic mice have been used to as a model to study
the pathogenesis of human CHD. GATA4 knockout is lethal to
mouse embryos, which suffer severe cardiac malformation (2). Double heterozygous GATA4-TBX5
mice exhibit atrioventricular septal defects (12), and double heterozygous
GATA4-GATA5 mice present with a phenotype of stenosis in the
aorta and pulmonary artery (13).
Heterozygous GATA4 mutation may result in ASD phenotypes,
ventricular septal defect, endocardial cushion defect, right
ventricular dysplasia and cardiomyopathy (14). GATA4 G295S transgenic mice
generated by Misra et al presented with a normal abdominal
cardiac morphology and cyclization pattern, but a weak ventricular
myocardium and a single ventricle in homozygous mice, which died at
the gestational age of 11.5 days; however, the heterozygous line
propagated and one subset carried the phenotypes of semilunar valve
stenosis and ASD (15). This
indicated that the complete lack of GATA4 gene activity has
a severe impact on cardiac development in mice, which cannot
survive or reproduce. The homozygous animal model limits the
functional study of GATA4 and to a certain extent cannot
represent human disease. However, heterozygous transgenic mice have
phenotypes that resemble, reasonably closely, human CHD phenotypes,
which are caused by genetic mutations.
Our group discovered a representative family with
simple ASD in 2007 (9). Since
then, thorough individual examinations were performed (e.g.,
collection of medical history, physical examination and
echocardiography) upon a total of 31 family members from 1st- to
3rd-level relatives. A total of 8 patients, including the probands,
were diagnosed with simple ASD. Subsequently, an in-depth study on
the mechanisms of GATA4 M310V mutation associated with
familial ASD by our group revealed that GATA4 M310V mutation
affects α-MHC promoter activity in in vitro
experiments (16).
In the present study, in order to identify the site
of GATA4 mutation capable of producing functional defects in
an in vivo model, a GATA4 M310V same-point mutation
was generated in transgenic mice. The mice were bred and screened,
producing a stable transgenic mouse line. We found that the
incidence of ASD in the heterozygous transgenic mice was higher
than that in their wild-type control littermates (P<0.05). In
addition, the peak pulmonary artery pressure (PPAP) and speed in
the heterozygous mice were higher compared with their wild-type
control littermates (P<0.05). The expression levels of
GATA4 downstream target genes (α-MHC) in the
homozygous mice were lower than those in their wild-type control
littermates (P<0.05). To summarize, GATA4 M310V mutations
were associated with a higher incidence of ASD-like cardiac
malformation in heterozygous transgenic mice compared with the
wild-type controls. Due to the changes in the expression of
downstream target genes in the homozygous transgenic mice, it was
inferred that the GATA4 M310V transgenic mouse model may be
used to simulate ASD reasonably well. The effects of mutant
GATA4 M310V were found to involve downstream genetic changes
that may result in the development of CHD.
Materials and methods
Construction and breeding of GATA4 M310V
transgenic mice
According to the human GATA4 encoding sequence, we
performed gene synthesis, followed by site-specific mutagenesis and
construct amplification in the pcDNA3.1-N-Myc-GATA4 (A928G) vector.
The DNA constructs were then microinjected into mouse embryos to
generate GATA4 M310V transgenic C57BL/6 mice. Each F0
transgene-positive transgenic mouse represented an individual
transgenic line and was housed in a cage with wild-type mice (each
cage contained 2 females and 1 male). All transgenic mice were
genotyped by polymerase chain reaction (PCR) to screen for positive
transgenic animals. The PCR products were verified by DNA
sequencing. The animals were bred using the aforementioned method
until the 4th generation. All animal experiments were carried out
according to the approval of the Animal Studies Ethics Committee of
the Peking University People’s Hospital, Beijing, China.
Evaluation of the genetic stability of
the exogenous genes of GATA4 M310V transgenic mice
Fluorescence-based quantitative PCR (qPCR) was used
to determine the copy number of transgenes in the mouse model and
to further screen a stably inherited transgenic line in this study.
The stably inherited transgenic mouse line was selected by qPCR
based on the following criteria: the genomic DNA of wild-type mice
was used as a control, the genomic DNA of transgenic mice was used
as a test sample, and GATA4 and GCG were used as
target fragments. Following confirmation of the sequence, the
GATA4 M310V plasmid DNA and GCG plasmid DNA were 10-fold
serially diluted with sterile distilled water to produce
109, 108, 107, 106,
105, 104 and 103 copy/μl
standards. qPCR was used to amplify 2 sets of standards (the
templates for GATA4 and GCG genes) and to generate a
standard curve for amplification efficiency. The 2−ΔΔCt
method (17) was used to measure
the exogenous gene copy number of the transgenic mice. The formula
was as follows: 2−ΔΔCt = 2−[(Ct GATA4 - Ct GCG)] in
transgenic samples - [(Ct GATA4 - Ct GCG)] in wild-type
samples.
Assessment of GATA4 protein
expression
A total of 3 (4-week-old) F3 transgene-positive
transgenic mice and their 3 wild-type littermates were selected.
The protein expression of GATA4 in the heart tissue was measured by
western blot analysis using glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) as an internal reference. The rabbit
polyclonal antibody to GATA4 (ab84593; Abcom, Cambridge, UK) which
reacts with the human one was selected at the concentration of 1
μg/ml to assess GATA4 protein expression.
Comparative study of cardiac morphology
and functions between transgenic and wild-type mice
The dead bodies of newborn heterozygous transgenic
mice were collected [approximately one tenth of the heterozygous
transgenic mice died shortly after birth (0–5 days after birth)].
Homozygous newborn mice and their wild-type littermates were
euthanized by carbon dioxide inhalation. The animals were dissected
by cutting open the chest to expose and remove the heart by cutting
the root of the major arteries. The dissected hearts were rinsed in
0.01 mol/l phosphate buffer to remove the blood and were
immediately placed in freshly prepared 4% neural buffered formalin.
The geart tissue was then fixed for 18 h, followed by conventional
dehydration, paraffin immersion and embedding procedures. The
paraffin-embedded tissue was serially sectioned into
4-μm-thick slices and stained in hematoxylin and eosin
(H&E) solution. The 8-week-old heterozygous transgenic mice and
their wild-type littermates were selected and anesthetized with
sodium pentobarbital (50 mg/kg body weight). The cardiac index was
calculated and following imaging of the mice, their heart rates
were carefully maintained at approximately 490 beats/min. An
ultra-high resolution and real-time ultrasonic molecular imaging
system (Vevo 2100; VisualSonics, Toronto, ON, Canada) was used to
perform M-mode echocardiography, two-dimensional echocardiography
and color Doppler echocardiography on the mice. Images of apical
four-chamber view (A4C), parasternal long axis view of the left
ventricle, and parasternal short axis view of the left ventricle
(at the aortic valve level) were then collected. The basic data
measurement and calculation of the heart and parameters of cardiac
function were obtained using conventional methods. The experimental
procedures were performed under double-blind conditions with regard
to genotype. The Chi-square test and Fisher’s exact test were used
to determine the difference in the incidence of ASD. The Student’s
t-test was used to compare the independent sample results of
echocardiography. P<0.05 represents a statistically significant
difference.
Assessment of GATA4 downstream target
gene expression
Total RNA from the heart tissues of the homozygous
transgenic mice and their wild-type littermates (control mice) was
extracted using TRIzol® reagent (Life Technologies,
Grand Island, NY, USA). Two micrograms of RNA were immediately
reverse-transcribed into cDNA using the Promega Reverse
Transcription kit (Promega, Madison, WI, USA). The expression
levels of the α-MHC, cTNC, GATA6,
NKX2.5 and TBX5 genes were measured using the Applied
Biosystems StepOnePlus™ real-time PCR system using the KAPA
SYBR® FAST qPCR kit Master Mix (2X) Universal (Kapa
Biosystems, Worburn, MA, USA). The primer sequences are listed in
Table I. The relative average
gene expression in the cardiac tissues of the transgenic mice and
their wild-type littermates was calculated using GAPDH as an
internal reference and the 2−ΔΔCt formula. Each sample
was evaluated in triplicate on a qPCR plate. SPSS 13.0 statistical
software (SPSS, Inc., Chicago, IL, USA) was used to perform
non-parametric tests to screen 2 independent samples. P-values
<0.05 were considered indicative of statistically significant
differences.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Gene name | Primer
sequences |
|---|
| GATA4
(PCR) | F:
CCTTCGACAGCCCGGTCCT
R: TGCACAGATAGTGACCCGTCCC |
| GATA4
(sequencing) | F:
CTGTGCCAACTGCCAGACC
R GTGCCCGTAGTGAGATGACAG |
| GATA4 (copy
no.) | F:
GGCCTCTCCTGTGCCAACTGC
R CTTCATGTAGAGGCCGCAGGCA |
| GCG | F:
AACATTGCCAAACGTCATGATG
R: GCCTTCCTCGGCCTTTCA |
| GAPDH | F:
CATCACTGCCACCCAGAAGACTG
R: ATGCCAGTGAGCTTCCCGTTCAG |
| α-MHC | F:
GCTGGAAGATGAGTGCTCAGAG
R: CCAGCCATCTCCTCTGTTAGGT |
| cTNC | F:
GATGGTTCGGTGCATGAAGGAC
R: CTTCCGTAATGGTCTCACCTGTG |
| GATA6 | F:
ATGCGGTCTCTACAGCAAGATGA
R: CGCCATAAGGTAGTGGTTGTGG |
| TBX5 | F:
CCAAAGACAGGTCTTGCGATTCG
R: TTCTCCTCCCTGCCTTGGTGAT |
| NKX2.5 | F:
GGTCTCAATGCCTATGGCTAC
R: GCCAAAGTTCACGAAGTTGCT |
Results
Establishment and breeding of GATA4 M310V
transgenic mice
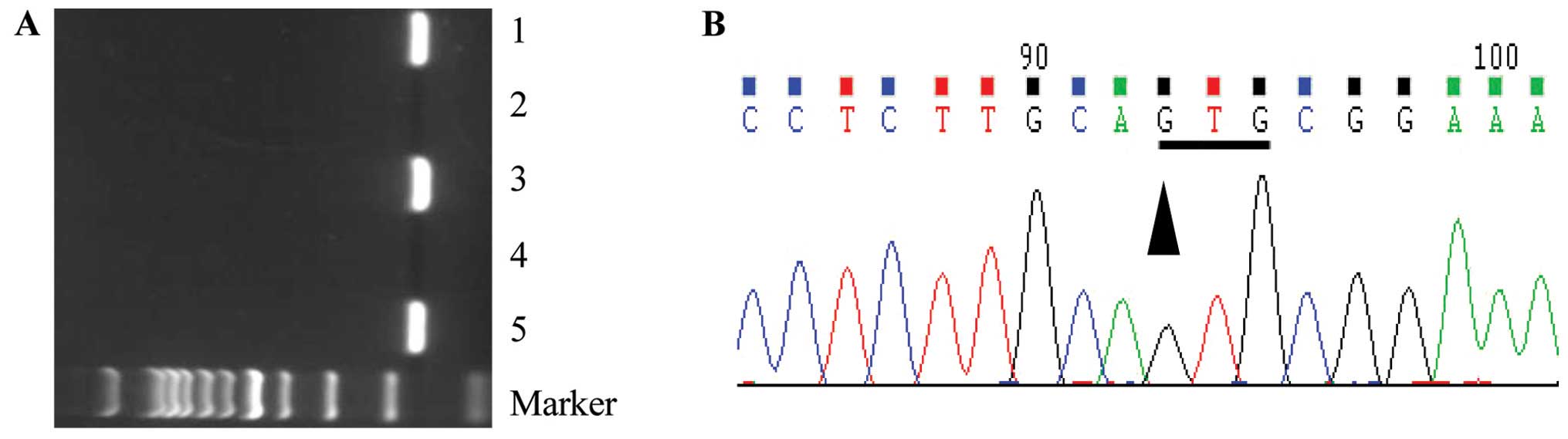
All transgenic mice were genotyped using PCR to
screen for positive transgenic animals (Fig. 1A). Genomic DNA was verified by
sequencing (Fig. 1B). The animals
were bred using the aforementioned method (as described in the
Materials and methods) to the 4th generation. A stably inherited
transgenic line was screened. The positive heterozygous transgenic
mice were inbred to generate homozygous transgenic mice.
Evaluation of genetic stability of
exogenous genes in GATA4 M310V transgenic mice
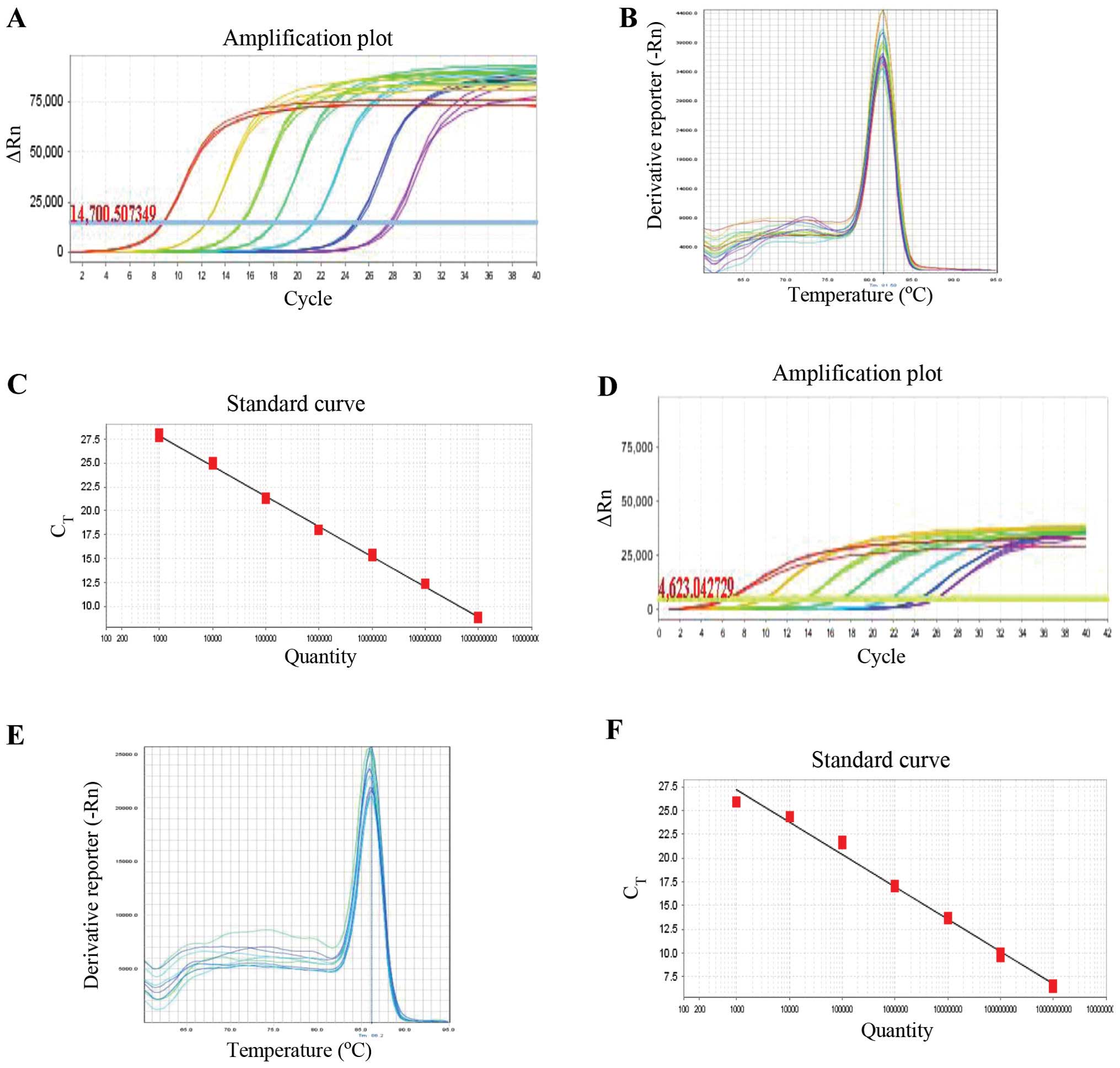
The GATA4 M310V application and melting
curves are illustrated in Fig. 2C
and D, and the GATA4 M310V standard curve is illustrated in
Fig. 2F. No sign peaks were found
in the melting curve and no primer dimers or other non-specific
amplification were observed. The data obtained by PCR amplification
were therefore considered reliable. The correlation coefficient of
the GCG standard curve coefficient was R2=0.999
with 101% amplification efficiency, and the correlation coefficient
of the GATA4 standard curve was R2=0.988, with
97% amplification efficiency. The 2−ΔΔCt method of qPCR
revealed that the amplification efficiency between the reference
and target genes was very close, ranging from 90–110%.
Protein expression of GATA4
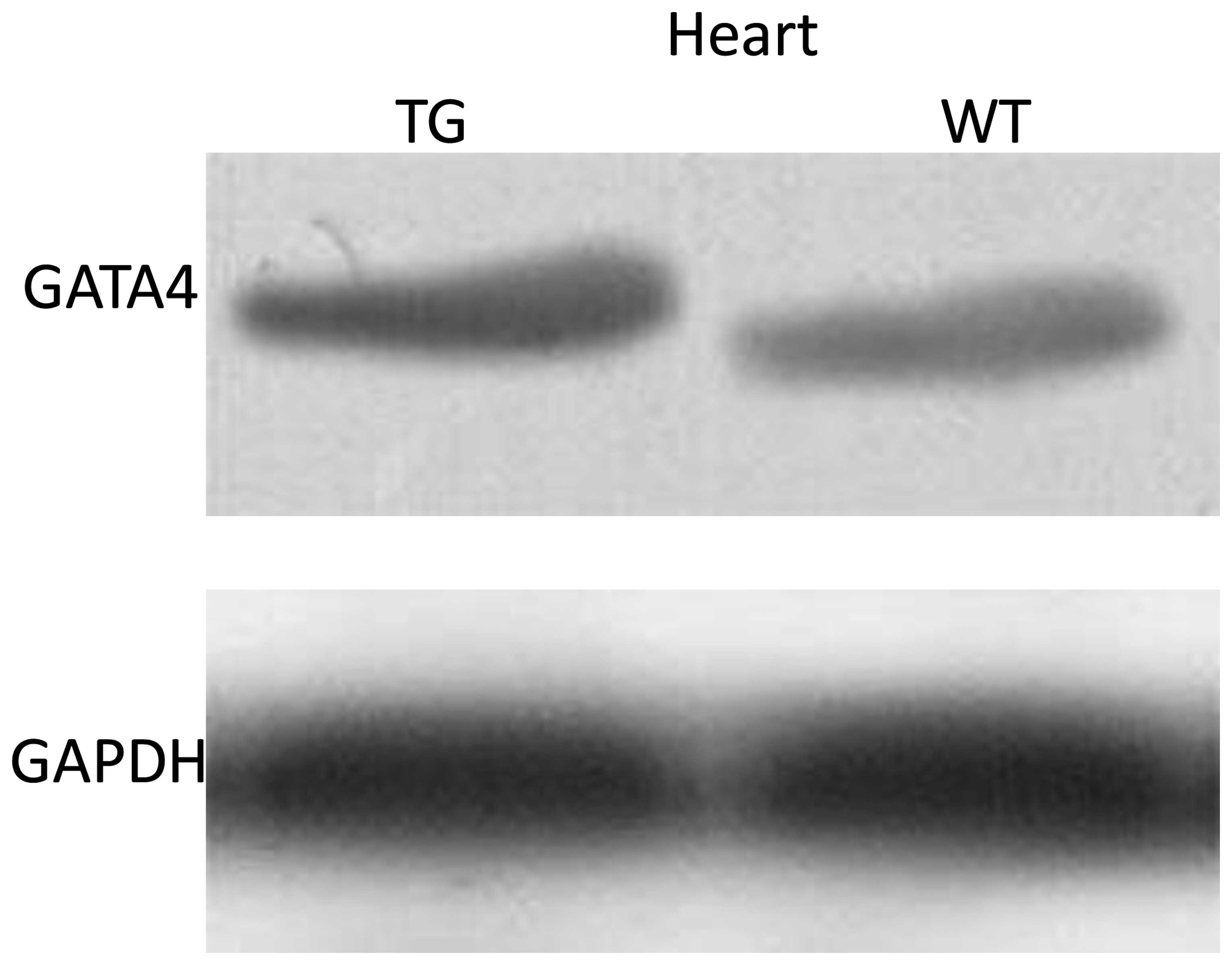
To assess the changes in GATA4 protein expression in
the hearts of F3 transgene-positive transgenic mice, western blot
analysis was used to analyze the protein expression levels of
GATA4. The results revealed that the protein expression level of
GATA4 was higher in the hearts of the transgenic mice than in those
of the wild-type mice (P<0.05; Fig. 3).
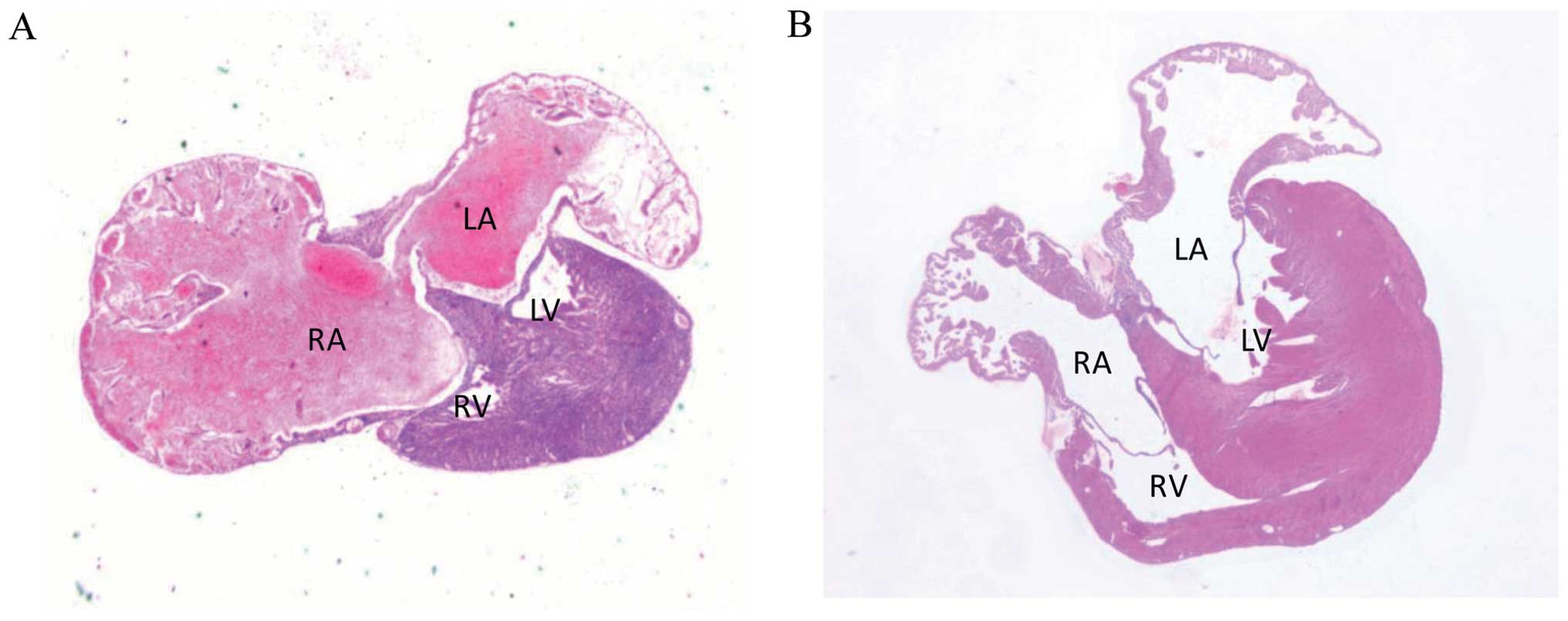
Cardiac phenotype of heterozygous
transgenic mice Observation of cardiac structure in newborn
mice
Approximately one tenth of the heterozygous
transgenic mice died shortly after birth (0–5 days after birth). To
observe the cardiac structural defects of the GATA4 M310V
heterozygous transgenic mice, the dead newborn pups were collected
and their hearts were dissected and subjected to conventional
histological analysis. H&E staining of the serial heart
sections revealed that, among the 15 newborn heterozygous mice, 9
mice (60%) had the ASD phenotype in their hearts (Fig. 4A) and 6 mice (40%) had no obvious
structural defect. A total of 20 newborn wild-type littermates were
selected as the controls and only 1 mouse (5%) had the ASD
phenotype in its heart. The remaining mice had no obvious
structural defects (Fig. 4B). The
details of the transgenic and wild-type mice regarding cardiac
structure are presented in Table
II, suggesting that GATA4 M310V produces the phenotype
of ASD in the heart.
| Table IIResults obtained from the heart
sections on the incidence of ASD between the transgenic mice and
the wild-type mice. |
Table II
Results obtained from the heart
sections on the incidence of ASD between the transgenic mice and
the wild-type mice.
| Heart section
results | Mouse group
|
|---|
| GATA4 M310V
heterozygous transgenic mice (n=15) | Wild-type mice
(n=20) |
|---|
| ASD | 60% (9)a | 5% (1) |
| Normal | 40% (6) | 95% (19) |
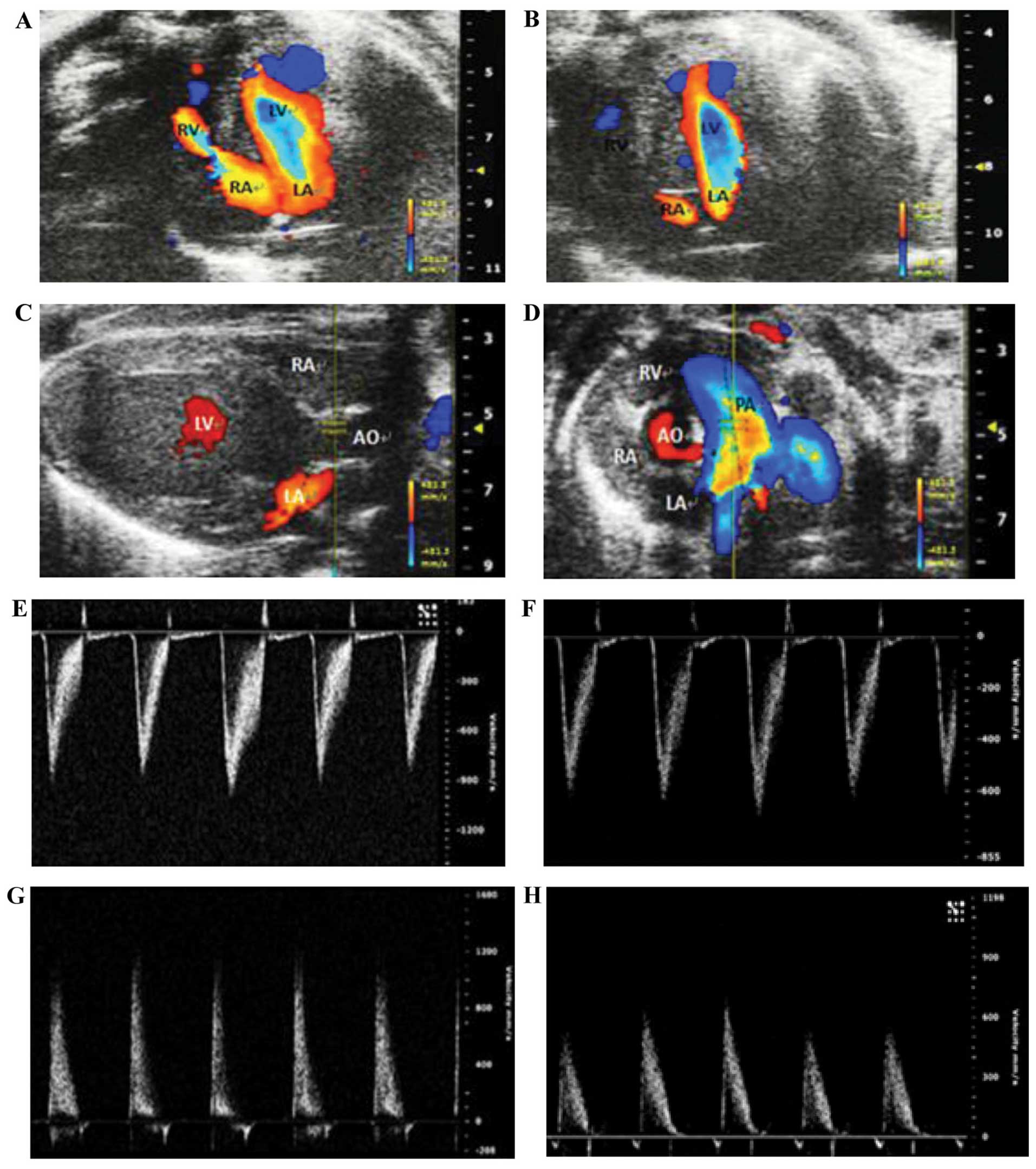
Echocardiography for the diagnosis and
assessment of cardiac structure and function in the mice
To access the differences in cardiac structure and
function between the transgenic mice and their wild-type
littermates, 10 transgenic and 6 wild-type mice (8-weeks old) were
selected for M-mode echocardiography, two-dimensional
echocardiography and color Doppler echocardiography using the Vevo
2100 ultra-high resolution and real-time ultrasonic molecular
imaging system. A4C was selected when using color Doppler
echocardiography to assess atrial septal integrity. Horizontal
sections of the parasternal long axis view of the left ventricle
and the parasternal short axis view of the left ventricle (at the
aortic valve level) were selected under color Doppler
echocardiography and M-mode echocardiography to indicate the aortic
and pulmonary artery flow velocities. Among the heterozygous
transgenic mice, 8 out of the 10 heterozygous transgenic mice
presented with intermittent blood flow occurring between the left
and right atria; however, only 1 out of the 6 wild-type littermates
presented with the similar symptom (significant difference between
groups, P<0.05) (Table III
and Fig. 5. The results of color
Doppler echocardiography on assessing aortic and pulmonary artery
peak velocities and pressure revealed that 6 out of the 10
heterozygous transgenic mice exhibited mild pulmonary stenosis,
whereas no abnormality was observed in their wild-type littermates.
No statistically significant difference was observed in left
ventricular function, left ventricular size and left ventricular
wall thickness between the heterozygous transgenic mice and their
wild-type littermates (Table
IV).
| Table IIIThe echocardiography results about
the incidence of ASD in transgenic and the wild-type mice. |
Table III
The echocardiography results about
the incidence of ASD in transgenic and the wild-type mice.
|
Echocardiography | Mouse group
|
|---|
| GATA4 M310V
heterozygous transgenic mice (n=10) | Wild-type mice
(n=6) |
|---|
| ASD | 80% (8)a | 17% (1) |
| Normal | 20% (2) | 83% (5) |
| Table IVEchocardiographic measurements. |
Table IV
Echocardiographic measurements.
| Measurement | Wild-type mice
(n=6) | Transgenic mice
(n=9) |
|---|
| Aortic peak
velocity (AV Peak Vel; mm/sec) | 788.47±72.71 | 919.40±195.79 |
| Aortic (AV) peak
pressure (mmHg) | 2.43±0.53 | 3.51±1.51 |
| Pulmonary artery
peak velocity (PV Peak Vel; mm/sec) |
−640.20±32.89a | −879.44±213.58 |
| Pulmonary (PV)
artery peak pressure (mmHg) | 1.66±0.16a | 3.13±1.63 |
| End-diastolic left
ventricular anterior wall thickness (LVAW; day, mm) | 0.80±0.07 | 0.89±0.11 |
| End-systolic left
ventricular anterior wall thickness (LVAW; sec, mm) | 1.29±0.23 | 1.45±0.16 |
| End-diastolic left
ventricular diameter (LVID; day, mm) | 3.30±0.48 | 3.39±0.28 |
| End-systolic left
ventricular diameter (LVID; sec, mm) | 2.17±0.66 | 2.04±0.24 |
| End-diastolic left
ventricular posterior wall thickness (LVPW; day, mm) | 0.71±0.09 | 0.80±0.13 |
| End-systolic left
ventricular posterior wall thickness (LVPW; sec, mm) | 1.16±0.18 | 1.28±0.17 |
| Ejection fraction
(EF, %) | 64.17±17.23 | 71.05±7.28 |
| Fractional
shortening (FS, %) | 35.29±12.13 | 39.72±5.79 |
| Left ventricular
end-diastolic volume (LV Vol; day, μl) | 45.45±14.94 | 47.49±9.34 |
| Left ventricular
end-systolic volume (LV Vol; sec, μl) | 17.75±13.17 | 13.68±4.09 |
| Heart rate (HR,
bpm) | 491.00±7.75 | 487.67±6.02 |
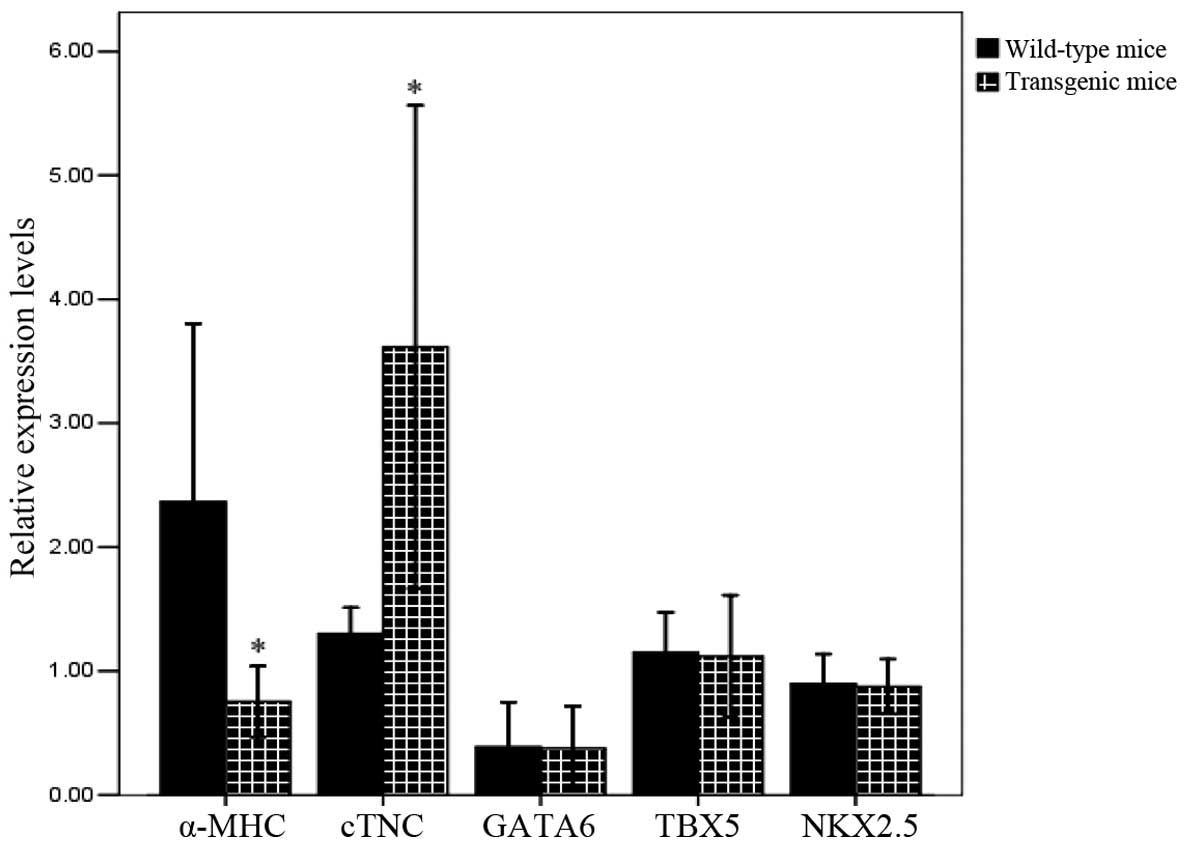
Expression of GATA4 target genes
To assess the changes in GATA4 transcript
target gene expression in the hearts of teh homozygous transgenic
mice, RNA was extracted from the hearts of newborn homozygous mice
and qPCR was used to analyze the expression levels of the
α-MHC, cTNC, GATA6, NKX2.5 and
TBX5 genes. The results revealed that expression levels of
α-MHC, as a GATA4 downstream target gene, was lower
in the hearts of the homozygous transgenic mice than in those of
the wild-type mice (P<0.05; Fig.
6), suggesting that the introduction of GATA4 M310V
inhibits the normal expression of this downstream gene. However,
the expression level of cTNC, as a GATA4 downstream target
gene, was higher in the hearts of the homozygous transgenic mice
than in those of the wild-type mice (P<0.05), in accordance with
the increased protein expression of GATA4. There was no
change in the expression levels of other GATA4 downstream
genes (NKX2.5 and GATA6) and GATA4
coactivating genes (TBX5). It was thus inferred that the
introduction of the GATA4 M310V exogenous gene suppressed
the expression of the GATA4 downstream gene, α-MHC,
which may subsequently cause the abnormal development of atrial
septum in the hearts of mice.
Discussion
The findings of the present study are as follows: i)
the GATA4 gene plays an important role in the development of
ASD; ii) the GATA4 downstream gene, α-MHC, is
associate with GATA4 function. The altered expression of
this gene affects the normal cardiac atrial septal formation during
development. This is associated with the incidence of ASD. The
conclusions of this study were based on the following: i) in a
previous study of ours on GATA4 mutation screening among
patients with CHD revealed that 8 patients with simple ASD from a
Chinese family with a history of CHD demonstrated the point
mutation of GATA4 M310V (9). ii) In another previous study of
ours, in vitro experiments revealed that GATA4
mutation affected its dose-positive correlation with α-MHC
activity (16). iii) GATA4
M310V transgenic mice developed for the purpose of this study
revealed ASD phenotypes. iv) GATA4 downstream gene
(α-MHC) expression in the hearts of GATA4 M310V
transgenic mice was significantly lower compared with the wild-type
littermates (P<0.05). In short, it was here inferred that
GATA4 M310V mutation may result in some functional defects,
which suppress the normal expression of GATA4 downstream
genes and affect the normal development of the cardiac septum,
resulting in the development of ASD.
ASD, in which blood flows between the left and right
atria of the heart, is a very common form of CHD. However, the
incidence of ASD and its etiology remain unclear. GATA4 gene
mutations have been demonstrated in many human families and are
associated with ASD and pulmonary stenosis (9,11,18). In a previous study by our group,
we presented a representative family with familial ASD (9).
The GATA4 gene is required for normal heart
development (2). In addition, the
GATA4 gene is a regulatory factor for heart development and
dose-related adjustment (19).
Different degrees of GATA4 gene defect have been found to
produce ASD in a mouse model, producing a variety of heart
malformations (14). In this
study, a GATA4 M310V transgenic mouse model presenting with
ASD phenotypes was developed. Echocardiography and histological
examination were used on the cardiac sections to determine the
differences in cardiac structure between the heterozygous
transgenic mice and their wild-type littermates. The heterozygous
transgenic mice carried the same ASD phenotypes as the 8 patients
with simple ASD from a previously tested family with CHD. It was
here inferred that these specific phenotypes of simple ASD are
triggered by GATA4 gene mutation during heart formation,
resulting in partially functional defects in the heart.
GATA4 is involved in the regulation of the
expression of cardiac structural genes, including α-MHC,
cTNC and ANF (3,4).
It has been found that the conserved GATA sites are in the
GATA4 regulatory region of many cardiac promoters and
GATA4 and potently activate their promoters (20). α-MHC is the major
contractile protein in the heart. Two putative GATA-binding
sites are in the proximal enhancer of the α-MHC gene which
suggests that GATA4 regulates its expression (21). Both MEF2C and GATA4
activate the gene expression of ANF and α-MHC
(22). The cTNC gene has
been used to examine the molecular mechanisms that regulate cardiac
muscle-specific transcription. It has been proven that GATA4 binds
specifically to the CEF-1 site of the cTNC enhancer (23). GATA6, another member of the
GATA family, is also critical to cardiac development and disorders.
It has been reported that GATA4 and GATA6 act cooperatively in the
heart (24). NKX2.5 is a
restrictive transcription factor of the heart and is involved in
normal cardiac development and the function of the cardiac
conduction system (25–27). NKX2.5 regulates
transcription and often interacts with other transcriptional
factors, including GATA4, MEF2C (25,28,29) and Hand2 (30). The TBX5 gene plays a key
role in cardiac morphology and the conduction system. Model mice
with a missing copy of the T-box transcription factor, TBX,
have ASD and sporadic ventricular septal defect (VSD) (31).
In a previous in vitro study published by our
group, we demonstrated that following the introduction of the
GATA4 expression vector using α-MHC promoter, the
increased concentration of GATA4 expression vector also
enhanced its promoter activity (16) (P<0.05). However, as we
demonstrated in the present study, the increased concentration of
the mutant GATA4 expression vector reduced the activity of
the α-MHC promoter. qPCR revealed that the expression of the
GATA4 downstream target gene, α-MHC, in the heart
tissues of the homozygous mice was lower than that in the heart
tiussues of the wild-type control mice (P<0.05), suggesting that
the introduction of GATA4 M310V inhibits the normal
expression of this downstream gene. However, the expression level
of cTNC, as a GATA4 downstream target gene, were
higher in the hearts of the homozygous transgenic mice than in
those of the wild-type mice (P<0.05), in accordance with the
increased protein expression of GATA4. However, the
expression of GATA6, NKX2.5 and TBX5 was not
altered. It was thus inferred that the GATA4 M310V point
mutation may result in the amino acid substitution of methionine to
valine in a corresponding position, leading to functional defects
in GATA4. This change also caused the suppression of
downstream gene α-MHC expression. The α-MHC gene may
be associated with the ASD phenotype of GATA4 M310V
transgenic mice.
This study focused on an important familial mutation
of the GATA4 gene found in a clinical study to further
establish the findings of basic research. In this study, a
GATA4 M310V transgenic mouse model of ASD phenotypes was
developed. Compared with the heart autopsies of embryonically
lethal mice and hearts from newborn GATA4 knockout mice,
this transgenic mouse line had a longer survival time and ensured
the stable inheritance of the exogenous gene through the transgenic
line. This mutant transgenic mouse model may be suitable for
research into human CHD. This mouse model may facilitate the study
of the genetic and environmental modifications that cause cardiac
malformations. It may also help the in-depth understanding of the
mechanisms underlying septal defects. This mouse model demonstrated
a specific gene mutation which may lead to the functional defects
in the animals. Studies on the potential function of GATA4
genes are warranted to determine the effect and changes associated
with GATA4 mutation.
References
|
1
|
Bentham J and Bhattacharya S: Genetic
mechanisms controlling cardiovascular development. Ann NY Acad Sci.
1123:10–19. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Molkentin JD, Lin Q, Duncan SA and Olson
EN: Requirement of the transcription factor GATA4 for heart tube
formation and ventral morphogenesis. Genes Dev. 11:1061–1072. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brunskill EW, Witte DP, Yutzey KE and
Potter SS: Novel cell lines promote the discovery of genes involved
in early heart development. Dev Biol. 235:507–520. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Temsah R and Nemer M: GATA factors and
transcriptional regulation of cardiac natriuretic peptide genes.
Regul Pept. 128:177–185. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sarkozy A, Conti E, Neri C, D’Agostino R,
Digilio MC, Esposito G, Toscano A, Marino B, Pizzuti A and
Dallapiccola B: Spectrum of atrial septal defects associated with
mutations of NKX2.5 and GATA4 transcription factors. J Med Genet.
42:e162005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Garg V, Kathiriya IS, Barnes R, et al:
GATA4 mutations cause human congenital heart defects and reveal an
interaction with TBX5. Nature. 424:443–447. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Christoforou N, Chellappan M, Adler AF,
Kirkton RD, Wu T, Addis RC, Bursac N and Leong KW: Transcription
factors MYOCD, SRF, Mesp1 and SMARCD3 enhance the cardio-inducing
effect of GATA4, TBX5, and MEF2C during direct cellular
reprogramming. PLoS One. 8:e635772013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dodou E, Verzi MP, Anderson JP, Xu SM and
Black BL: Mef2c is a direct transcriptional target of ISL1 and GATA
factors in the anterior heart field during mouse embryonic
development. Development. 131:3931–3942. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Y, Han ZQ, Yan WD, Tang CZ, Xie JY,
Chen H and Hu DY: A novel mutation in GATA4 gene associated with
dominant inherited familial atrial septal defect. J Thorac
Cardiovasc Surg. 140:684–687. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiang R, Fan LL, Huang H, Cao BB, Li XP,
Peng DQ and Xia K: A novel mutation of GATA4 (K319E) is responsible
for familial atrial septal defect and pulmonary valve stenosis.
Gene. 534:320–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garg V, Kathiriya IS, Barnes R,
Schluterman MK, King IN, Butler CA, Rothrock CR, Eapen RS,
Hirayama-Yamada K, Joo K, Matsuoka R, Cohen JC and Srivastava D:
GATA4 mutations cause human congenital heart defects and reveal an
interaction with TBX5. Nature. 424:443–447. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maitra M, Schluterman MK, Nichols HA,
Richardson JA, Lo CW, Srivastava D and Garg V: Interaction of Gata4
and Gata6 with Tbx5 is critical for normal cardiac development. Dev
Biol. 326:368–377. 2009. View Article : Google Scholar :
|
|
13
|
Laforest B and Nemer M: GATA5 interacts
with GATA4 and GATA6 in outflow tract development. Dev Biol.
358:368–378. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rajagopal SK, Ma Q, Obler D, et al:
Spectrum of heart disease associated with murine and human GATA4
mutation. J Mol Cell Cardiol. 43:677–685. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Misra C, Sachan N, McNally CR, Koenig SN,
Nichols HA, Guggilam A, Lucchesi PA, Pu WT, Srivastava D and Garg
V: Congenital heart disease-causing Gata4 mutation displays
functional deficits in vivo. PLoS Genet. 8:e10026902012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang Y, Han Z, Han H and Chen Y: Mechanism
involved in the familial atrial septal defects caused by GATA-4
M310V single gene mutation. China Circ J. 3:211–214. 2013.In
Chinese.
|
|
17
|
Ballester M, Castelló A, Ibáñez E, Sánchez
A and Folch JM: Real-time quantitative PCR-based system for
determining transgene copy number in transgenic animals.
Biotechniques. 37:610–613. 2004.PubMed/NCBI
|
|
18
|
Nemer G, Fadlalah F, Usta J, Nemer M,
Dbaibo G, Obeid M and Bitar F: A novel mutation in the GATA4 gene
in patients with Tetralogy of Fallot. Hum Mutat. 27:293–294. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zeisberg EM, Ma Q, Juraszek AL, Moses K,
Schwartz RJ, Izumo S and Pu WT: Morphogenesis of the right
ventricle requires myocardial expression of Gata4. J Clin Invest.
115:1522–1531. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pu WT, Ishiwata T, Juraszek AL, Ma Q and
Izumo S: GATA4 is a dosage-sensitive regulator of cardiac
morphogenesis. Dev Biol. 275:235–244. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McBride K and Nemer M: Regulation of the
ANF and BNP promoters by GATA factors: Lessons learned for cardiac
transcription. Can J Physiol Pharmacol. 79:673–681. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Molkentin JD, Kalvakolanu DV and Markham
BE: Transcription factor GATA-4 regulates cardiac muscle-specific
expression of the alpha-myosin heavy-chain gene. Mol Cell Biol.
14:4947–4957. 1994.PubMed/NCBI
|
|
23
|
Morin S, Charron F, Robitaille L and Nemer
M: GATA-dependent recruitment of MEF2 proteins to target promoters.
EMBO J. 19:2046–2055. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ip HS, Wilson DB, Heikinheimo M, Tang Z,
Ting CN, Simon MC, Leiden JM and Parmacek MS: The GATA-4
transcription factor transactivates the cardiac muscle-specific
troponin C promoter-enhancer in nonmuscle cells. Mol Cell Biol.
14:7517–7526. 1994.PubMed/NCBI
|
|
25
|
Zhao R, Watt AJ, Battle MA, Li J, Bondow
BJ and Duncan SA: Loss of both GATA4 and GATA6 blocks cardiac
myocyte differentiation and results in acardia in mice. Dev Biol.
317:614–619. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jay PY, Harris BS, Maguire CT, Buerger A,
Wakimoto H, Tanaka M, Kupershmidt S, Roden DM, Schultheiss TM,
O’Brien TX, Gourdie RG, Berul CI and Izumo S: Nkx2-5 mutation
causes anatomic hypoplasia of the cardiac conduction system. J Clin
Invest. 113:1130–1137. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Terada R, Warren S, Lu JT, Chien KR,
Wessels A and Kasahara H: Ablation of Nkx2-5 at mid-embryonic stage
results in premature lethality and cardiac malformation. Cardiovasc
Res. 91:289–299. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Takeda M, Briggs LE, Wakimoto H, Marks MH,
Warren SA, Lu JT, Weinberg EO, Robertson KD, Chien KR and Kasahara
H: Slow progressive conduction and contraction defects in loss of
Nkx2-5 mice after cardiomyocyte terminal differentiation. Lab
Invest. 89:983–993. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schlesinger J, Schueler M, Grunert M,
Fischer JJ, Zhang Q, Krueger T, Lange M, Tönjes M, Dunkel I and
Sperling SR: The cardiac transcription network modulated by Gata4,
Mef2a, Nkx2.5, Srf, histone modifications, and microRNAs. PLoS
Genet. 7. pp. e10013132011, View Article : Google Scholar
|
|
30
|
Yamagishi H, Yamagishi C, Nakagawa O,
Harvey RP, Olson EN and Srivastava D: The combinatorial activities
of Nkx2.5 and dHAND are essential for cardiac ventricle formation.
Dev Biol. 239:190–203. 2001. View Article : Google Scholar
|
|
31
|
Bruneau BG, Nemer G, Schmitt JP, Charron
F, Robitaille L, Caron S, Conner DA, Gessler M, Nemer M, Seidman CE
and Seidman JG: A murine model of Holt-Oram syndrome defines roles
of the T-box transcription factor Tbx5 in cardiogenesis and
disease. Cell. 106:709–721. 2001. View Article : Google Scholar : PubMed/NCBI
|