Introduction
Of the malignant tumors which form in essential
liver or intrahepatic bile duct, 90% are usually hepatocellular
carcinoma (HCC) (1). HCC is the
fifth most common solid tumor worldwide. China has the the highest
incidence of liver cancer worldwide (2). The majority of patients are already
in the advanced stages of the disease at the time of diagnsis
(1,3). At present, surgical resection and
liver transplantation are the most effective treatment methods for
HCC. However, clinical research has shown that >50% of patients
with HCC relapse following liver resection, an event which is
related to the fact that HCC is a type of heterogeneous tumor with
a high invasive- and metastatic capacity. Intrahepatic spread and
extrahepatic invasion and metastasis are the leading causes of a
poor prognosis for patients with HCC (4,5).
MicroRNAs (miRNAs or miRs) are a class of
evolutionarily highly conserved endogenous, non-coding, small RNA
molecules, which can combine with the mRNA 3′-UTR region of target
genes, suppress the translation of target genes, and then inhibit
the expression of target genes (6). miRNAs play an important role in
various pathological and physiological processes, including
carcinogenesis and cancer metastasis, and promote the proliferation
and metastasis of malignant tumors, including HCC (7). The expression of miR-15a and
miR-16-1 has been shown to be downregulated or lacking in the
majority of patients with chronic lymphocytic leukemia (8). As regulatory factors, miRNAs can
either play the role of proto-oncogenes or anti-oncogenes,
affecting the biological characteristics of cells. Research has
shown that the down-regulated expression of miR-16 in HepG2,
SK-HEP-1 and Huh7 HCC cells may be associated with the
proliferation of HCC (9). miR-16
has been shown to suppress the proliferative activity of various
cancer cells and can inhibit the invasion and metastasis of human
glioma and colorectal cancer cells (10–12). Epithelial-mesenchymal transition
(EMT) is a key event in tumor invasion and metastasis. This process
is not only involved in the disappearance of cancer cell polarity,
but it is also closely related to the disappearance of the
connection between cancer cells, the remodeling of the
cytoskeleton, and changes to key signaling pathways. The induction
of EMT and the abnormal expression of miRNAs are closely related to
the occurrence, development, proliferation, migration and
recurrence of tumors, including HCC (11–13).
The occurrence and development of HCC is a
multi-step process and involves multiple different factors. It also
involves the disorder of key signaling pathways that regulate
various cellular processes, such as proliferation, apoptosis,
invasion and metastasis. Previous research has confirmed that the
abnormal activation of phosphatidylinositol 3-kinase (PI3K)/protein
kinase B (PKB or Alt) in HCC plays an important role in promoting
cancer cell proliferation, apoptosis and metastasis (14). Bax, Bcl-2, and matrix
metalloproteinases (MMPs), namely MMP-2 and MMP-9, are downstream
molecules of the PI3K/AKT signaling pathway. Following activation,
the PI3K/AKT pathway affects the expression of these downstream
molecules, and then promotes the proliferation and invasion of HCC
(15,16). Tsai et al further confirmed
that the activation of the PI3K/Akt signaling pathway was a key
mechanism in the process of EMT, and that the suppressed activation
of the pathway led to the reversal of EMT, the re-expression of
E-cadherin and β-cadherin, and the reduction of the expression of
the interstitial cell marker, vimentin (17). It has also been previously
demonstrated that miR-16 regulates the expression of EMT-related
proteins in U87 and U251 glioma cells through the Akt signaling
pathway (12).
Based on the above-mentioned data, we formulated a
hypothesis that miR-16 may inhibit the proliferation, invasion and
metastasis of HCC cells, and that this effect may be mediated
through the PI3K/Akt signaling pathway. Therefore, this study aimed
to examine the effects of miR-16 on the expression of Akt signaling
pathway-related proteins and on the proliferation, invasion and
metastasis of HCC cells.
Materials and methods
Cell lines
Human SK-Hep-1 HCC cells were purchased from the
Cell Bank of the Chinese Academy of Sciences (Beijing, China). The
human HepG2 and SMCC-7721 cells were purchased from ATCC (Manassas,
VA, USA). The Huh-7 HCC cells were purchased from Shanghai Bomaide
Biological Technology. Co. Ltd. (Shanghai, China).
Reagents
MTT solution was obtained from Sigma Co. (St. Louis,
MO, USA); rabbit anti-Bax (Cat no. 1063-1), anti-Bcl-2 (Cat no.
1017-1), anti-MMP-2 (Cat no. 1948-1) and anti-MMP-9 (Cat no.
1939-1) antibodies were purchased from Epitomics Co. (Burlingame,
CA, USA); rabbit anti-PI3K (Cat no. 11889S), anti-Akt (Cat no.
2920) and anti-p-Akt (Cat no. 11962) antibodies were from Cell
Signaling Techonology (Danvers, MA, USA); rabbit anti-E-cadherin
(Cat no. 2707-1) and anti-vimentin (Cat no. 5409-1) antibodies were
purchased from Abcam (Cambridge, MA, USA); β-actin antibody (Cat
no. AA128), the Annexin V/PI double dye detection kit (Cat no.
C1063), goat anti-rabbit and anti-mice antibody labeled with
horseradish peroxidase (Cat no. A0208 and A0258) were all from the
Beyotime Institute of Biotechnology (Jiangsu, China); Transwell
Chambers were purchased from Corning Inc. (Cat no. 3413, Corning,
NY, USA); fetal bovine serum, DMEM culture medium and trypsin were
all purchased from Gibco Life Technologies (Cat no. 10099-141 and
11965-092, Carlsbad, CA, USA); methyl thiazolyltetrazolium and
crystal violet staining solution were from Sigma Co. (Cat no.
M-2128 and 219215).
Instruments
The CO2 incubator was obtained from
Thermo Scientific, Waltham, MA, USA; the super clean workbench was
from Thermo Scientific; the inverted microscope was from Nikon,
Tokyo, Japan; the flow cytometer was obtained from BD Biosciences
(San Diego, CA, USA; the Mini dual vertical electrophoresis
apparatus, the Mini transfer electrophoresis unit, and the
ChemiDoc™ XRS Gel Imaging System were all from Bio-Rad, Hercules,
CA, USA.
miRNA synthesis, preparation and
storage
Synthetic miR-16 mimic, miR-16 inhibitor and mimic
negative (NC) control, and inhibitor negative control were
purchased from Guangzhou RiboBo Co., Ltd., (Guangzhou, China).
Synthetic samples were first centrifuged (10,000 rpm, 25°C) then
diluted per 5 nmol miRNA, and subsequently added to 250 µl
diethylpyrocarbonate (DEPC)-treated water, which was added to 20
µM mother liquor; the samples were repackaged and stored at
−20°C. miR-16 mimic and NC mimic chemical synthesis were undertaken
by Applied Biosystems, Foster City, CA, USA). The miR sequences
were as follows: miR-16 mimic,
5′-UAGCAGCACGUAAAUAUUGGCGCCAAUAUUUACGUGCUGCUAUU-3′; miR-16 mimic
negative control, 5′-UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT-3′;
miR-16 inhibitor, 5′-CGCCAAUAUUUACGUGCUGCUA-3′; and miR-16
inhibitor negative control, 5′-CAGUACUUUUGUGUAGUACAA-3′.
Transfection of cells with miRNAs
On the day prior to transfection, the cells were
inoculated in medium without penicillin-streptomycin solution
(6-well plates, 2,000 µl per well; increase or decrease in
proportion). Cells were transfected when they reached a confluence
of 30–50%. The miRNAs were diluted with a moderate amount of
serum-free Opti-MEM® I culture medium (Cat. no.
31985-070; Invitrogen, Carlsbad, CA, USA) (250 µl) by
blending gently. Lipofectamine 2000 was diluted in Opti-MEM I by
blending gently, followed by incubation for 5 min at room
temperature. The diluted Lipofectamine 2000 (Cat. no. 13778150;
Invitrogen) was gently mixed with the diluted miRNAs, and the
mixture was incubated for 20–30 min at room temperature. Following
the addition of the miRNA/Lipofectamine 2000 compound to the 6-well
plates, the plates were gently shaken. After 6 h, the DMEM
containing 10% serum was added, and the cells were cultivated at
37°C in a CO2 incubator for 24–72 h.
Reverse-transcription PCR (RT-PCR)
validation of miR-16 expression in 4 different HCC cell lines
Total RNA was extracted using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA), according to the manufacturer's
instructions. The primer sequences were as follows: miR-16,
5′-TAGCAGCACGTAAATATTGGCG-3′; and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH; internal control),
5′-TGACTTCAACAGCGACACCCA-3′. RNA was reverse transcribed into cDNA
and PCR amplification was carried out using a one-step RT-PCR kit
(Cat. no. C81401180; Invitrogen). Approximately 5 µl
amplification product was used in the next step of 2% agarose gel
detection. Following electrophoresis, bands were detected and
images were acquired using an ultraviolet spectrophotometer
(Quawell, San Jose, CA USA).
Determination of the cell proliferation
rate by MTT assay
The HepG2 cells were inoculated in 96-well plates,
with 100 µl in each well and 4 plates for each group. When
cell confluence reached 50%, miR-16 mimic, miR-16 inhibitor and the
corresponding negative control were transfected into the cells, and
the transfection concentrations were 50 and 200 nmol, respectively.
One day prior to transfection, the cells were digested and counted,
and inoculated in 96-well plates. Following transfection for 48 h,
20 µl 5 mg/ml MTT were added and followed by cultivation for
4 h. After the nutrient solution was removed, 150 µl DMSO
were added to each well followed by shaking until the crystals had
dissolved. The optical density (OD) was measured at 560 nm using an
enzyme standard instrument, and the relative proliferation rate was
calculated with 630 nm as the reference wavelength.
Analysis of cell migration
The HepG2 cells were inoculated in 96-well plates at
100 µl in each well and 4 plates for each group. When the
confluence of the cells reached 50%, miR-16 mimic, miR-16 inhibitor
and the corresponding negative controls were transfected into the
cells, and the transfection concentrations were 50 and 200 nmol,
respectively. On the day prior to transfection, the cells were
digested and counted, and inoculated in 96-well plates. Following
transfection for 48 h, the cells were digested and added to a
Transwell upper chamber, and the cells in the lower chamber
continued to be cultured for a further 24 h in DMEM with 5% fetal
bovine serum. The Transwell chamber was subsequently removed and
washed, and the cells were fixed with paraformaldehyde and stained
with crystal violet. The number of migrated cells was counted in a
total of 5 fields of view under an inverted optical microscope. The
average number of cells per field of view was calculated, and this
represented the migratory ability of the cells.
Analysis of cell invasion
The Matrigel was evenly spread on the microfilm of
the Transwell chamber. The remaining steps were the same as those
in described in 'RT-PCR validation of miR-16 expression in 4
different HCC cells lines'. The average number of HepG2 cells per
field of view was calculated, which represented the invasive
ability of the cells.
Detection of cell apoptosis
The HepG2 HCC cells were inoculated in 6-well
plates. When the confluence of the cells reached 50%, miR-16 mimic,
miR-16 inhibitor and the corresponding negative controls were
transfected into the cells, and the transfection concentrations
were 50 and 200 nmol, respectively, with 3 wells/group. After 24 h,
the cells were digested and harvested, and then detected using a
FACSCanto flow cytometry instrument (BD Biosciences) after being
stained with Annexin V-FITC/PI for 30 min in the dark.
Western blot analysis
The HepG2 cells were inoculated in 96-well plates,
100 µl/well and 4 plates/group. When the cell confluence
reached 50%, miR-16 mimic, miR-16 inhibitor and the corresponding
negative controls were transfected into the cells, and the
transfection concentrations were 50 and 200 nmol, respectively.
Following transfection for 48 h, the cells were scraped and
centrifuged (10,000 rpm for 10 min at 25°C). Following the addition
of the appropriate amount of RIPA lysis buffer, the cells were
placed in a vortex meter and shaken for 30 sec. After 40 min, they
were centrifuged (4°C, 10,000 rpm) for 10 min, and the supernatant
was then removed, and thus we obtained the total protein. The
protein concentration was measured using a BCA kit. The proteins
separated by SDS gel electrophoresis, and then transferred to the
membrane using the wet transfer method. The membrane was immersed
and incubated in primary antibody solution (Bax, Bcl-2, MMP 2, MMP
9, E-cadherin, vimentin, PI3K, β-actin, Akt and p-Akt) overnight at
4°C. After being rinsed, the proteins were immersed and incubated
in the secondary antibody solution (goat anti-rabbit and anti-mouse
antibody labeled with horseradish peroxidase) at room temperature
for 1–2 h. The membrane was removed and rinsed, followed by the
addition of ECL solution, and were then examined using a gel
imaging system. Each antibody gray value stripes were detected
using Quantity One software.
Statistical analysis
Means and standard deviations (SD) were used to
summarize continuous variables. To determine the differences
between groups, the independent t-test and one-way ANOVA were used
where appropriate. A two-sided P-value <0.05 was considered to
indicate a statistically significant difference. All analyses were
performed using SPSS software, version 17.0.
Results
Expression of the miR-16 gene in 4 HCC
cell lines
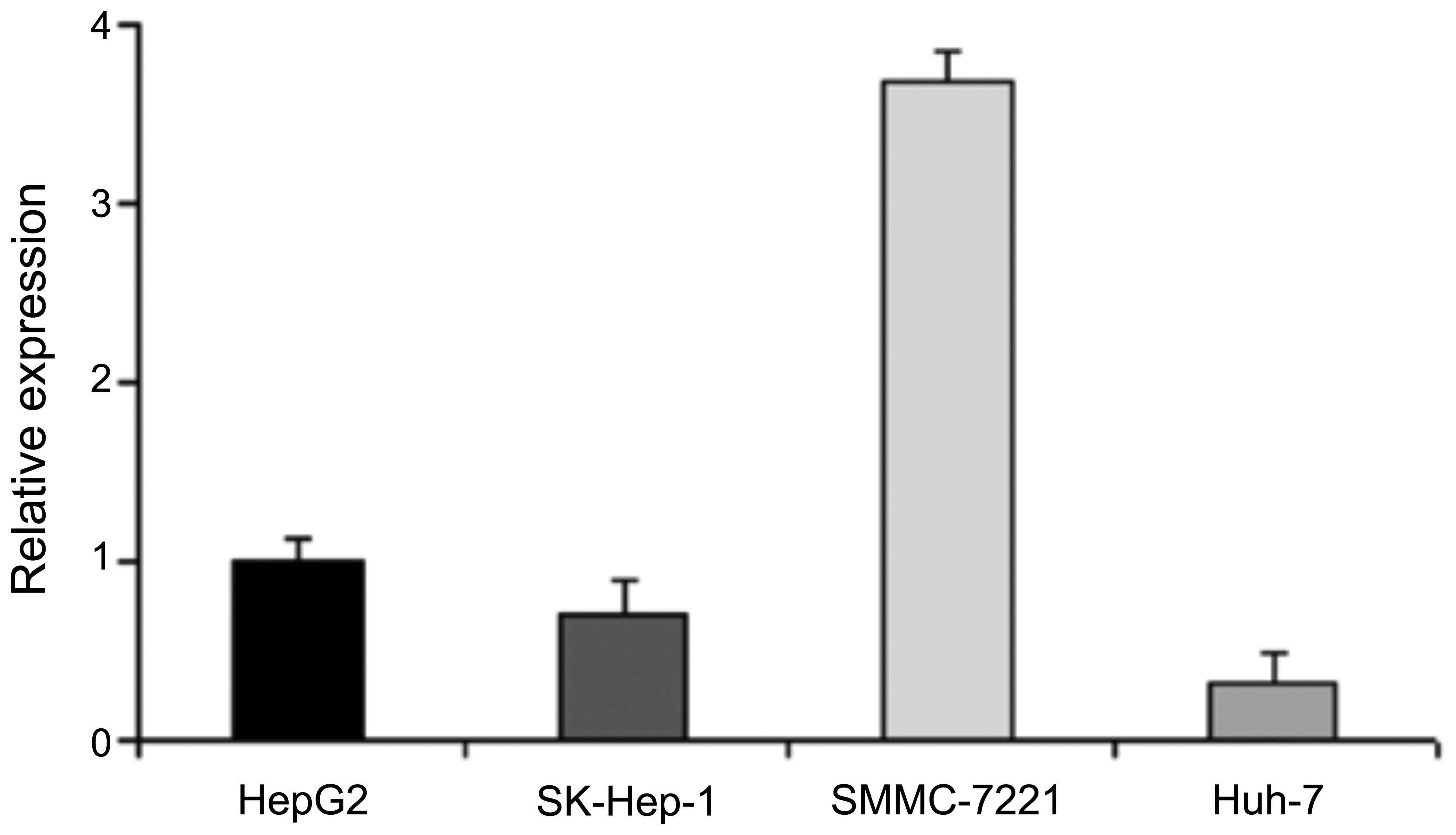
The results from RΤ-PCR revealed that of the 4 HCC
cell lines, the mRNA expression level of miR-16 was highest in the
SMMC-7721 cells and lowest in the SK-Hep-1 and Huh-7 cells;
moderate expression levels were observed in the HepG2 cells
(Fig. 1). Therefore, we selected
HepG2 as the cell line for use in followup experiments.
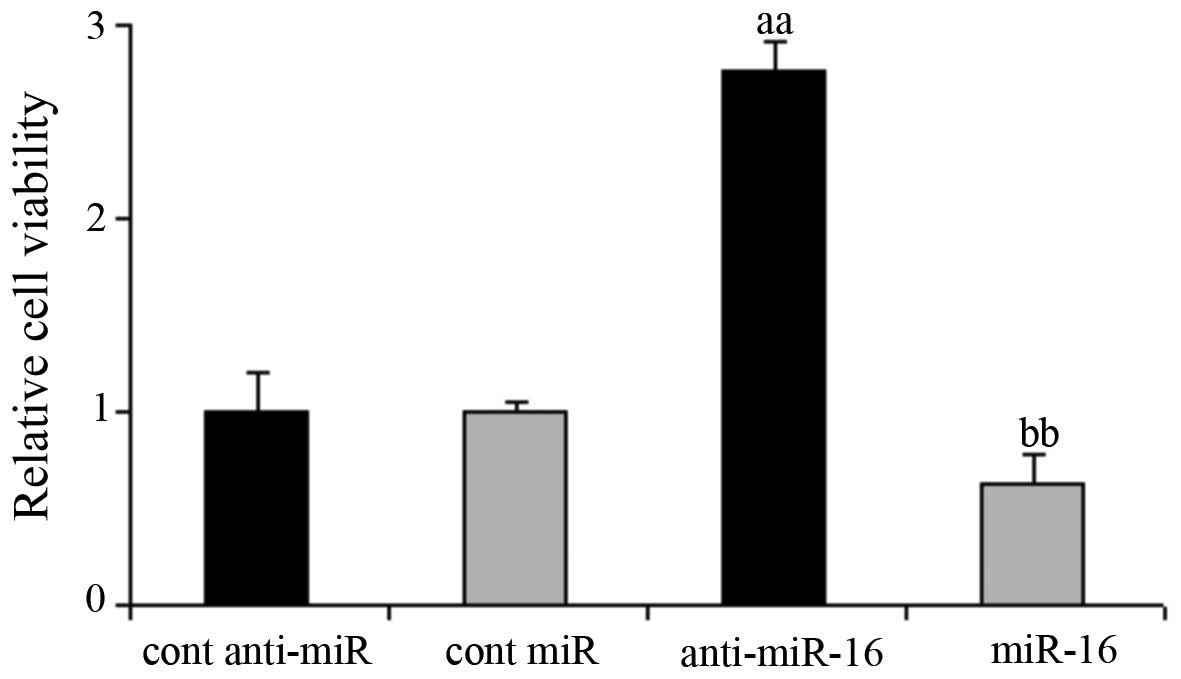
Effect of miR-16 on HepG2 cell
viability
The HepG2 cells were transfected with the miR-16
mimic and miR-16 inhibitor, as well as the respective negative. The
results of MTT assay revealed that compared with the negative
control-transfected cells, the knockdown of miR-16 significantly
increased HepG2 cell viability, whereas the overexpression of
miR-16 inhibited HepG2 cell viability (P<0.05; Fig. 2).
Effect of miR-16 on HepG2 cell
apoptosis
The results of Annexin V/PI flow cytometry revealed
that compared with the negative control-transfected cells, the
impact of miR-16 knockdown on HepG2 cell apoptosis was not
significant. However, the overexpression of miR-16 significantly
promoted HepG2 cell apoptosis (P<0.05; Table I).
 | Table IEffect of miR-16 on HepG2 cell
apoptosis. |
Table I
Effect of miR-16 on HepG2 cell
apoptosis.
| Stage of
apoptosis | Cont anti-miR | Cont miR | Anti-miR-16 | miR-16 |
|---|
| Early apoptosis
(%) | 3.26±0.22 | 0.84±0.13 | 3.98±0.75 | 17.24±2.56a |
| Late apoptosis
(%) | 0.7±0.16 | 0.24±0.05 | 1.76±0.58 | 14.84±1.59a |
Effect of miR-16 on the migratory and
invasive ability of HepG2 cells
The results of the migration and invasion assays
revealed that the overexpression of miR-16 significantly reduced
the number of HepG2 cells migrating through and invading the filter
membrane (P<0.05). The impact of miR-16 knockdown on the
migratory and invasive ability of the HepG2 cells was not
significant (Table II).
| Table IIEffect of miR-16 on the migration and
invasion of HepG2 cells. |
Table II
Effect of miR-16 on the migration and
invasion of HepG2 cells.
|
Migration/invasion | Cont anti-miR | Cont miR | Anti-miR-16 | miR-16 |
|---|
| Migration | 99.10±9.01 | 99.23±7.93 | 98.67±3.78 | 66.76±6.39a |
| Invasion | 99.00±8.02 | 99.09±9.26 | 99.10±2.52 | 67.35±5.73a |
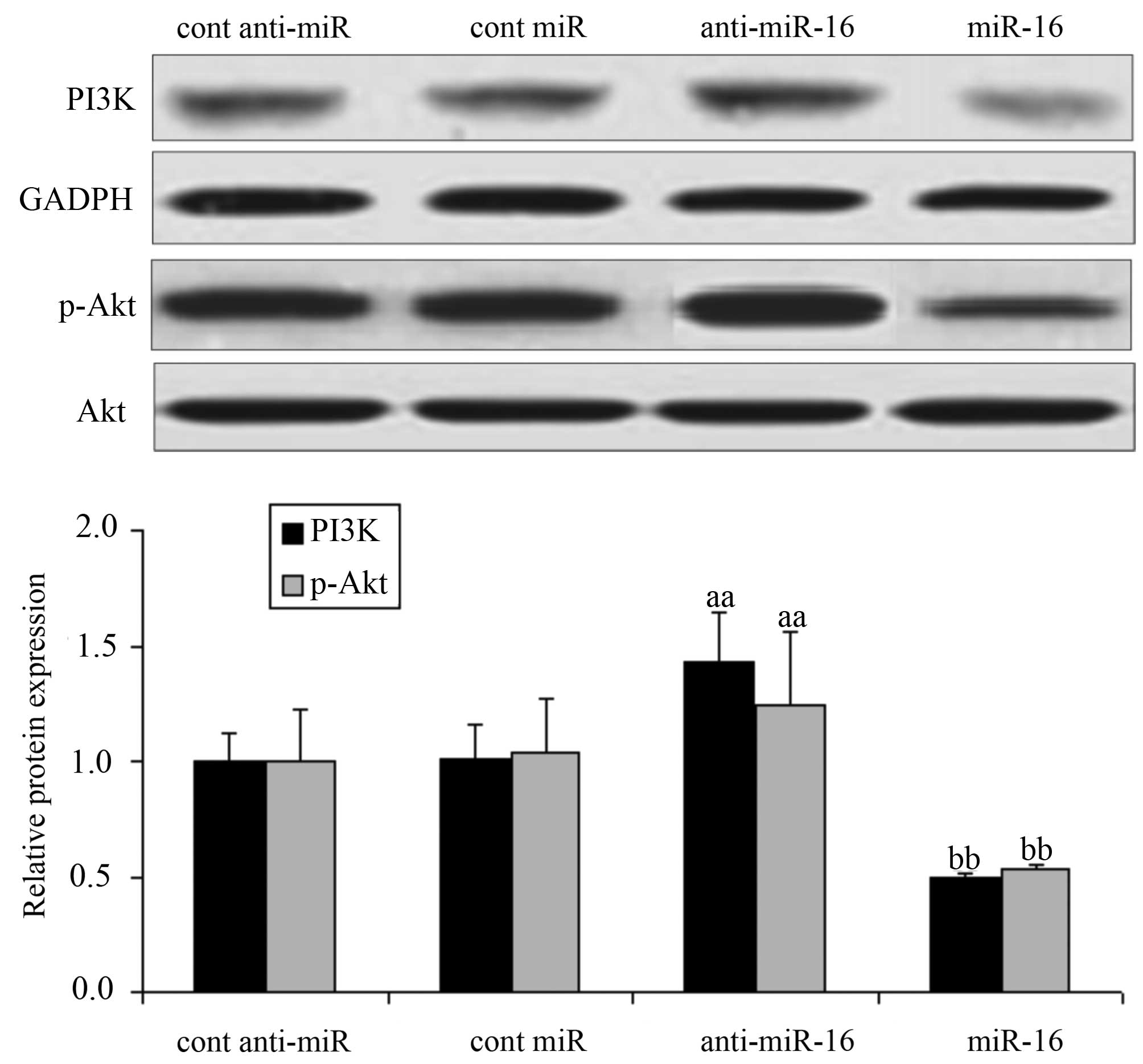
Effect of miR-16 on proteins related to
the PI3K/Akt signaling pathway in HepG2 cells
The expression of PI3K/Akt pathway-realted proteins
in HepG2 cells was examined by western blot analysis. The
expression level of PI3K and the Akt phosphorylation level in the
cells transfected with the miR-16 inhibitor (anti-miR-16) were
significantly increased compared with the levels in the negative
control-transfected cells (P<0.05). Conversely, the expression
level of PI3K and the Akt phosphorylation level were significantly
decreased in the miR-16 mimic-transfected cells (P<0.05;
Fig. 3).
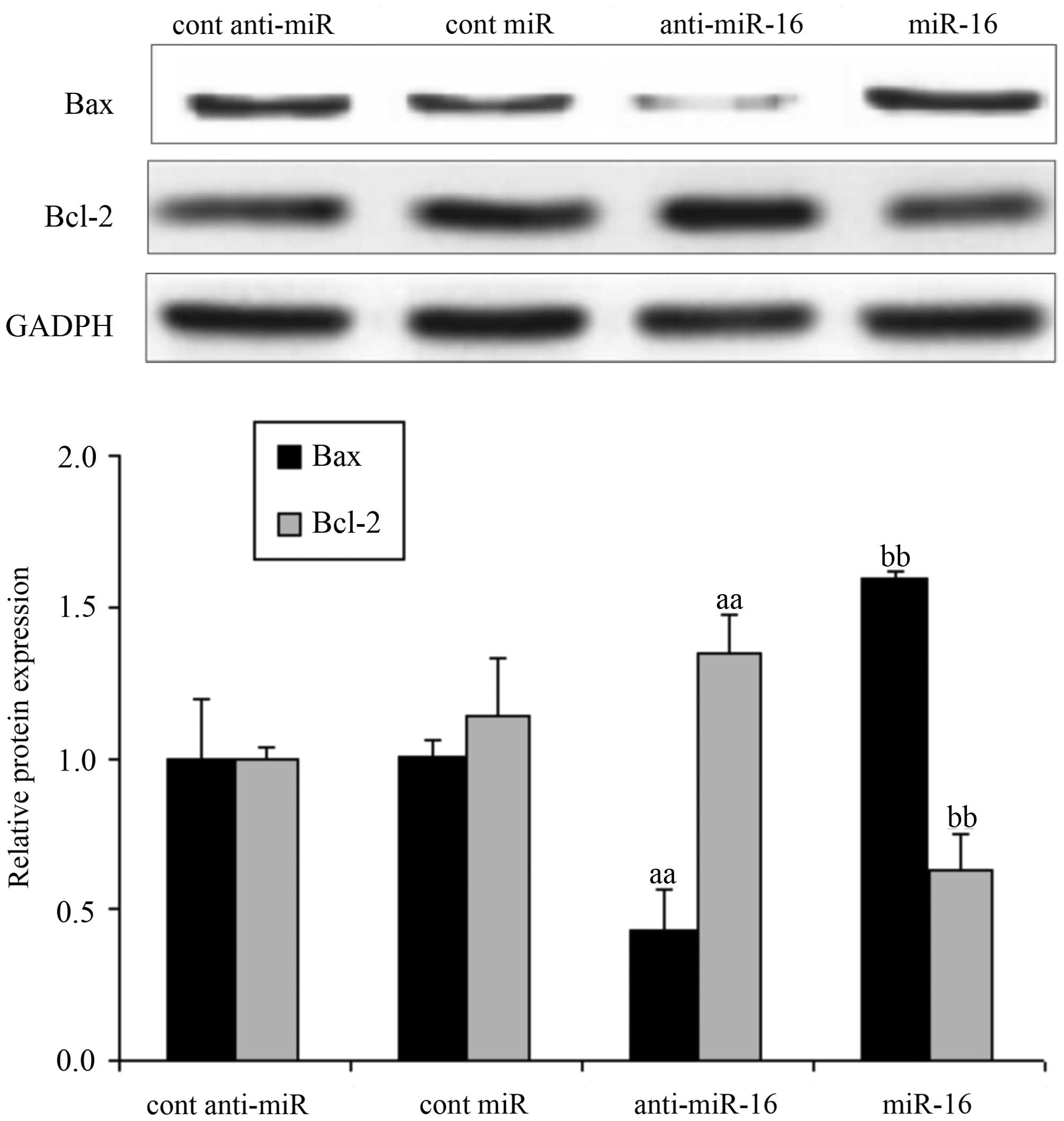
Effect of miR-16 on proteins related to
apoptosis in HepG2 cells
Following transfection of the HepG2 cells with
miR-16 mimic or miR-16 inhibitor, or the respective negative
controls, we measured the expression levels of Bax and Bcl-2.
Compared with the negative control-transfected cells, the
expression level of Bax significantly decreased and that of Bcl-2
increased in the cells transfected with the miR-16 inhibitor
(anti-miR-16) (P<0.05); however, the expression level of Bax
significantly increased and that of Bcl-2 decreased in the cells
transfected with the miR-16 mimic (P<0.05; Fig. 4).
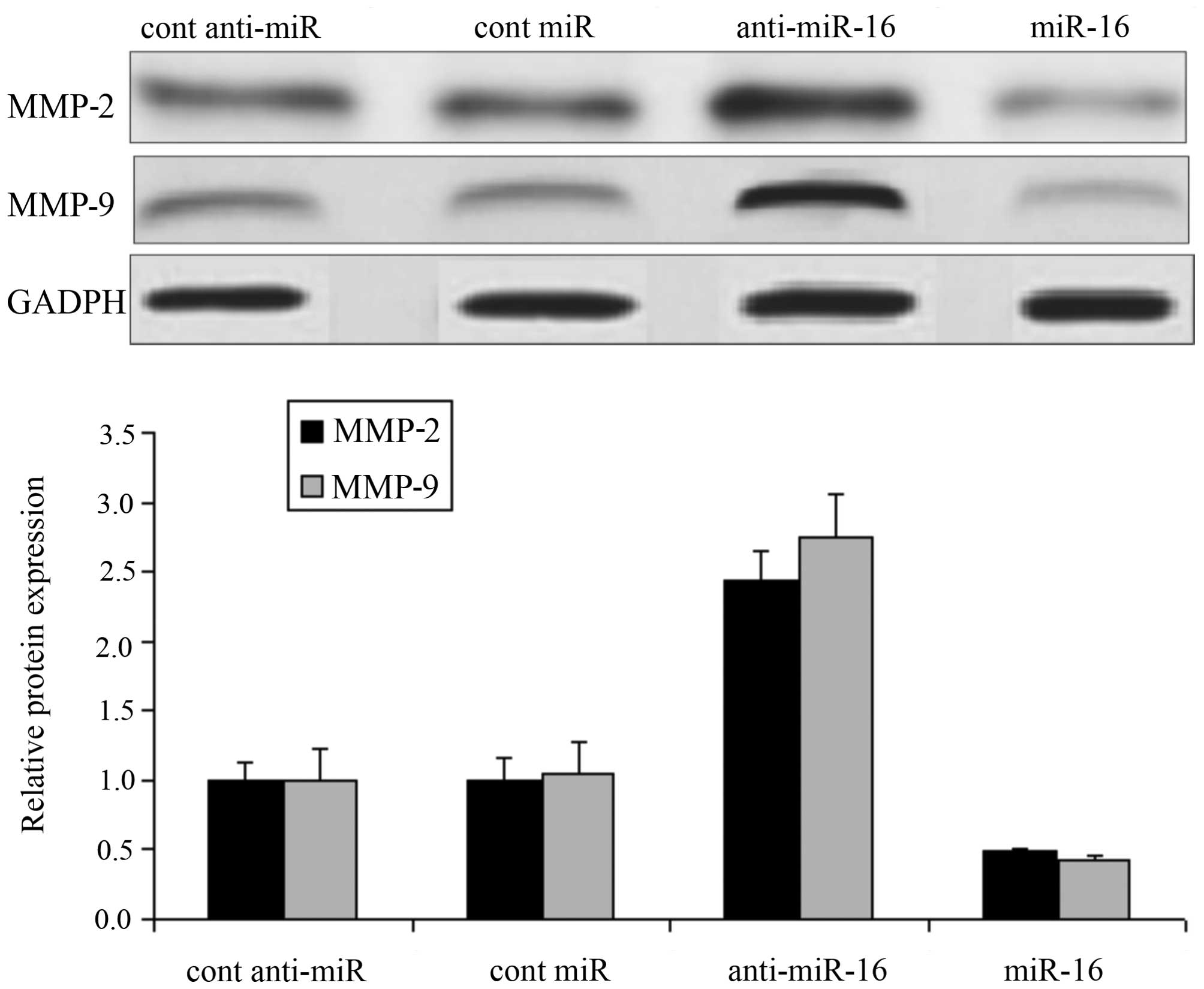
Effect of miR-16 on MMP protein
expression in HepG2 cells
Following transfection of the HepG2 cells with
miR-16 mimic or miR-16 inhibitor, or the respective negative
controls, we measured the expression levels of MMP-2 and MMP-9 in
the HepG2 cells. Compared with the negative control-transfected
cells, the expression levels of MMP-2 and MMP-9 in the cells
transfected with the miR-16 inhibitor (anti-miR-16) were
significantly increased; however, the MMP-2 and MMP-9 expression
levels were also markedly inhibited in the cells transfected with
the miR-16 mimic (P<0.05; Fig.
5).
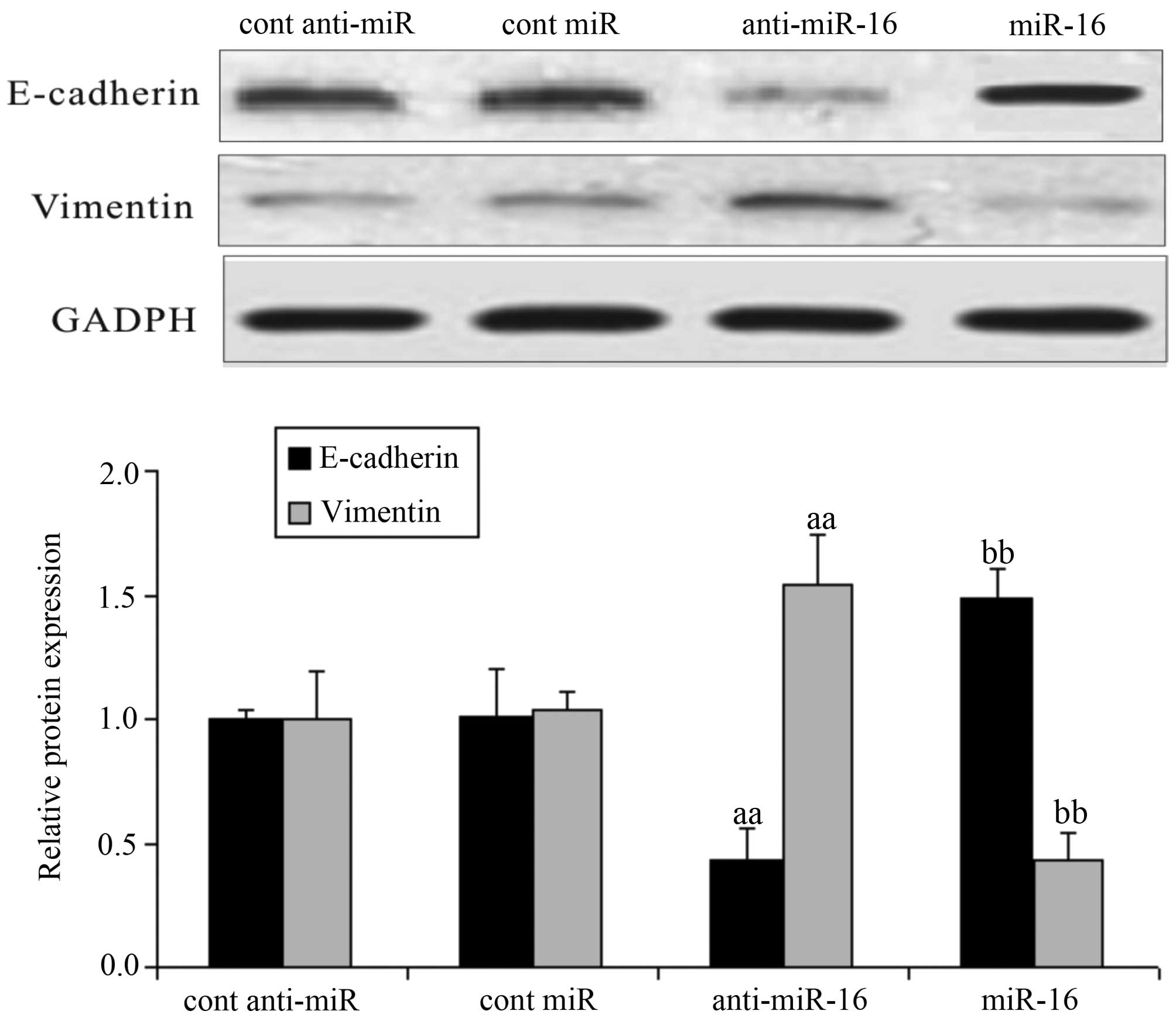
Effect of miR-16 on EMT-related proteins
in HepG2 cells
Following transfection of the HepG2 cells with
miR-16 mimics or miR-16 inhibitor, or the respective negative
controls, we measured the expression levels of E-cadherin and
vimentin in the HepG2 cells. Compared with the negative
control-transfected cells, the expression of E-cadherin was
downregulated and that of vimentin was upregulated in the cells
transfected with the miR-16 inhibitor (anti-miR-16) (P<0.05). In
the cells transfected with the miR-16 mimic, the expression of
E-cadherin was upregulated and that of vimentin was down-regulated
(P<0.05; Fig. 6).
Discussion
miR-16 is an miRNA which exerts an antitumor effect
and is closely associated with cancer cell proliferation and the
development of malignant tumors. The results of the present study,
revealed moderate mRNA expression levels of miR-16 in HepG2 cells,
and thus this cell line was selected for use in follow-up
experiments. We also observed the effects of miR-16 on the
proliferation, invasion and metastasis of HepG2 HCC cells.
Bax and Bcl-2 are genes known to play a role in the
regulation of apoptosis. Bax expression is downregulated in
multiple tumors, including HCC tissue (18). Bcl-2 overexpres-sion is clearly
involved in the occurrence, development and metastasis of HCC cells
(19). Studies have demonstrated
that miRNAs are involved in cancer cell differentiation, apoptosis
and proliferation, functioning as either tumor suppressor genes or
proto-oncogenes (19,20). miR-16 plays an extremely important
biological role in the occurrence and development of tumors
(21,22). Bhattacharya et al reported
that miR-16 and -15a control Bmi-1 expression in ovarian cancer and
that miR-15a and miR-16 overexpression inhibited the proliferation
of ovarian carcinoma cells (23).
Tang et al reported that miR-16 regulates the proliferation
and apoptosis of ovarian epithelial carcinoma cells (24). In a previous study, microbubbles
with rick miR-16 were subcutaneously injected into nude mice with
prostate cancer and it was found that that miR-16 entered the
cancer cells and inhibited the expression of Bcl-2 with fluorescent
reporter gene expression (25).
Our study also confirmed that miR-16 overexpression increased Bax
expression and decreased Bcl-2 expression in HepG2 cells.
MMPs are a group of proteases, which play a key role
in the degradation of the extracellular matrix, and in the invasion
and metastasis of cancer cells. It has previously been demonstrated
that MMPs play a crucial role in the process of HepG2 cell invasion
(26). High expression levels of
MMP-2 and MMP-9 have been shown to correlate with tumor size and
the occurrence of lymphatic metastasis in HCC (27). Renjie and Haiqian (10) and Wang et al (12) reported that miR-16 overexpression
inhibited the migration and invasion of glioma and pituitary cells.
Yang et al that miR-16 inhibits glioma cell growth and
invasion by suppressing Bcl-2 and the nuclear factor-κB1/MMP-9
pathway (25). In the present
study, following transfection of HepG2 cells with miR-16 mimic or
miR-16 inhibitor, we found that miR-16 overexpression decreased the
expression levels of MMP-2 and MMP-9 and thus inhibited cell
migration and invasion.
Previous research has indicated that EMT plays an
important role in tumorigenesis, including local invasion, and
transfer and diffusion through the circulatory system, and is
related to the aggressiveness of cancer cells. The beginning of
metastasis is said to occur when the EMT process commences in
malignant cancer cells (28,29). A marked characteristic of EMT is
the downregulated expression of the intercellular adhesion
molecule, E-cadherin, and the upregulated expression of a series of
mesenchymal markers, including N-cadherin, vimentin and fibronectin
(30,31). In HCC, the lack of E-cadherin
expression is associated with tumor invasion, migration and a poor
prognosis (32). It is considered
that E-cadherin plays a role in the invasion ability of numerous
cancer cells (33). Vimentin
expression contributes to the enhanced invasion and metastasis in
cancer (32,34). Previous studies have reported that
miR-16 influences cancer cell metastasis by affecting the process
of EMT in colorectal cancer and pituitary tumor cells (10,11). In the present study, we noted that
miR-16 overexpression in HepG2 cells downregulated the expression
of vimentin, and upregulated the expression of E-cadherin.
We also noted that miR-16 overexpression in HepG2
cells led to the significantly increased expression of PI3K and
decreased levels of Akt phosphorylation. PI3K belongs to the
phosphatidylinositol-dependent kinase family, and activates Akt
through a series of signal transductions. Akt participates in
various basic cellular processes, including cell proliferation,
apoptosis, migration, invasion and EMT. In relation to HCC tissue,
the abnormal activation of the PI3K/Akt pathway has been proven to
be closely related to the prognosis of patients (35). It has also previously been
demonstrated that PI3K/Akt activity is significantly decreased by
the overexpression of miRNAs in HCC cells by treating HCC cells
with miRNAs, thus inhibiting the proliferation, invasion and
metastasis of HCC cells (36,37).
Previous research has confirmed that mitochondrial
dysfunction and deactivation of Akt may contribute to the
apoptosis-inducing effects of carnosic acid in HepG2 cells
(16). Mice MMP-2 gene knockout
has been shown to affect the secretion of MMPs, which is mediated
through the PI3K/Akt signaling pathway (38). In HepG2 cells, the downregulated
expression of MMP-9, through the PI3K/Akt signaling pathway, has
been shown to inhibit cell invasion (39). In the processes of cancer cell
invasion and metastasis, it has been confirmed that EMT is
associated with multiple signaling pathways. The activation of the
Akt and Erk signaling pathways prompts the EMT process in cancer
cells, providing cancer cells with the ability to survive (40). Wang et al (12) also found that miR-16 regulated
EMT-related proteins in the glioma cell lines, U87 and U25, and
that this involved the Akt signaling pathway; the decreased
expression of E-cadherin and vimentin was also noted (12). Thus, miR-16 may also influence
HepG2 cell proliferation, migration and invasion through the
PI3K/Akt signaling pathway, and affect the expression of related
proteins (12).
It must be noted that the experimental design of our
current study had certain limitations. First, this study was
designed in order to explore the influence of miR-16 on HepG2 cell
proliferation, invasion, metastasis and apoptosisat the the protein
level, not the genetic level. The synthesis of mRNA and protein
should not be examined together as this would affect the results.
Moreover, this study was an in vitro study. No in
vivo experiments using animals were performed to observe the
effects of miR-16 on the proliferation, invasion and metastasis of
cancer cells from transplanted tumors. Thus, further in vivo
animal studies are warranted to confirm our findings.
In conclusion, the findings of the present study
demonstrated that miR-16 overexpression inhibited the
proliferation, invasion and metastasis of HepG2 HCC cells, and that
these effects involved the upregulation of Bax expression, the
downregulation of Bcl-2 expression, and the decrease in the
expression levels of MMP-2 and MMP-9. In addition, E-cadherin
expression was increased and vimentin expression was decreased and
this also involved the inhibition of PI3K expression and Akt
phosphorylation.
References
|
1
|
Ichikawa T, Sano K and Morisaka H:
Diagnosis of pathologically early HCC with EOB-MRI: Experiences and
current consensus. Liver Cancer. 3:97–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wei KR, Yu X, Zheng RS, Peng XB, Zhang SW,
Ji MF, Liang ZH, Ou ZX and Chen WQ: Incidence and mortality of
liver cancer in China, 2010. Chin J Cancer. 33:388–394.
2014.PubMed/NCBI
|
|
3
|
Ringelhan M, O'Connor T, Protzer U and
Heikenwalder M: The direct and indirect roles of HBV in liver
cancer: prospective markers for HCC screening and potential
therapeutic targets. J Pathol. 235:355–367. 2015. View Article : Google Scholar
|
|
4
|
de Lope CR, Tremosini S, Forner A, Reig M
and Bruix J: Management of HCC. J Hepatol. 56(Suppl 1): S75–S87.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Finn RS: Advanced HCC: Emerging molecular
therapies. Minerva Gastroenterol Dietol. 58:25–34. 2012.PubMed/NCBI
|
|
6
|
Brodersen P, Sakvarelidze-Achard L,
Bruun-Rasmussen M, Dunoyer P, Yamamoto YY, Sieburth L and Voinnet
O: Widespread translational inhibition by plant miRNAs and siRNAs.
Science. 320:1185–1190. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Anwar SL and Lehmann U: MicroRNAs:
emerging novel clinical biomarkers for hepatocellular carcinomas. J
Clin Med. 4:1631–1650. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calin GA, Cimmino A, Fabbri M, Ferracin M,
Wojcik SE, Shimizu M, Taccioli C, Zanesi N, Garzon R, Aqeilan RI,
et al: MiR-15a and miR-16-1 cluster functions in human leukemia.
Proc Natl Acad Sci USA. 105:5166–5171. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu G, Yu F, Xiao Z, Xu K, Xu J, Tang W,
Wang J and Song E: Hepatitis B virus X protein downregulates
expression of the miR-16 family in malignant hepatocytes in vitro.
Br J Cancer. 105:146–153. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Renjie W and Haiqian L: MiR-132, miR-15a
and miR-16 syner-gistically inhibit pituitary tumor cell
proliferation, invasion and migration by targeting Sox5. Cancer
Lett. 356B:568–578. 2015. View Article : Google Scholar
|
|
11
|
Shi L, Jackstadt R, Siemens H, Li H,
Kirchner T and Hermeking H: p53-induced miR-15a/16-1 and AP4 form a
double-negative feedback loop to regulate epithelial-mesenchymal
transition and metastasis in colorectal cancer. Cancer Res.
74:532–542. 2014. View Article : Google Scholar
|
|
12
|
Wang Q, Li X, Zhu Y and Yang P:
MicroRNA-16 suppresses epithelial-mesenchymal transition-related
gene expression in human glioma. Mol Med Rep. 10:3310–3314.
2014.PubMed/NCBI
|
|
13
|
Zhang LY, Liu M, Li X and Tang H:
miR-490-3p modulates cell growth and epithelial to mesenchymal
transition of hepatocellular carcinoma cells by targeting
endoplasmic reticulum-Golgi intermediate compartment protein 3
(ERGIC3). J Biol Chem. 288:4035–4047. 2013. View Article : Google Scholar :
|
|
14
|
Dai XF, Ding J, Zhang RG, Ren JH, Ma CM
and Wu G: Radio-sensitivity enhancement of human hepatocellular
carcinoma cell line SMMC-7721 by sorafenib through the MEK/ERK
signal pathway. Int J Radiat Biol. 89:724–731. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tian T, Nan KJ, Guo H, Wang WJ, Ruan ZP,
Wang SH, Liang X and Lu CX: PTEN inhibits the migration and
invasion of HepG2 cells by coordinately decreasing MMP expression
via the PI3K/Akt pathway. Oncol Rep. 23:1593–1600. 2010.PubMed/NCBI
|
|
16
|
Xiang Q, Ma Y, Dong J and Shen R: Carnosic
acid induces apoptosis associated with mitochondrial dysfunction
and Akt inactivation in HepG2 cells. Int J Food Sci Nutr. 66:76–84.
2015. View Article : Google Scholar
|
|
17
|
Tsai JH, Hsu LS, Lin CL, Hong HM, Pan MH,
Way TD and Chen WJ: 3,5,4′-Trimethoxystilbene, a natural
methoxylated analog of resveratrol, inhibits breast cancer cell
invasiveness by downregulation of PI3K/Akt and Wnt/β-catenin
signaling cascades and reversal of epithelial-mesenchymal
transition. Toxicol Appl Pharmacol. 272:746–756. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nagasawa T, Matsushima-Nishiwaki R, Toyoda
H, Matsuura J, Kumada T and Kozawa O: Heat shock protein 20 (HSPB6)
regulates apoptosis in human hepatocellular carcinoma cells: Direct
association with Bax. Oncol Rep. 32:1291–1295. 2014.PubMed/NCBI
|
|
19
|
Zhao N, Sun BC, Zhao XL, Wang Y, Sun HZ,
Dong XY, Meng J and Gu Q: Changes in microRNAs associated with
Twist-1 and Bcl-2 overexpression identify signaling pathways. Exp
Mol Pathol. 99:524–532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chamorro-Jorganes A, Araldi E, Penalva LO,
Sandhu D, Fernández-Hernando C and Suárez Y: MicroRNA-16 and
microRNA-424 regulate cell-autonomous angiogenic functions in
endothelial cells via targeting vascular endothelial growth factor
receptor-2 and fibroblast growth factor receptor-1. Arterioscler
Thromb Vasc Biol. 31:2595–2606. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goretti E, Rolland-Turner M, Léonard F,
Zhang L, Wagner DR and Devaux Y: MicroRNA-16 affects key functions
of human endothelial progenitor cells. J Leukoc Biol. 93:645–655.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bhattacharya R, Nicoloso M, Arvizo R, Wang
E, Cortez A, Rossi S, Calin GA and Mukherjee P: MiR-15a and MiR-16
control Bmi-1 expression in ovarian cancer. Cancer Res.
69:9090–9095. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang R, Cui ZM and Lou YH: MicroRNA-16
regulates the proliferation, invasion and apoptosis of ovarian
epithelial carcinoma cells in vitro. Zhonghua Fu Chan Ke Za Zhi.
47:846–850. 2012.In Chinese.
|
|
25
|
Yang TQ, Lu XJ, Wu TF, Ding DD, Zhao ZH,
Chen GL, Xie XS, Li B, Wei YX, Guo LC, et al: MicroRNA-16 inhibits
glioma cell growth and invasion through suppression of BCL2 and the
nuclear factor-κB1/MMP9 signaling pathway. Cancer Sci. 105:265–271.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ordoñez R, Carbajo-Pescador S,
Prieto-Dominguez N, García-Palomo A, González-Gallego J and Mauriz
JL: Inhibition of matrix metalloproteinase-9 and nuclear factor
kappa B contribute to melatonin prevention of motility and
invasiveness in HepG2 liver cancer cells. J Pineal Res. 56:20–30.
2014. View Article : Google Scholar
|
|
27
|
Zhang Y, Shen Y, Cao B, Yan A and Ji H:
Elevated expression levels of androgen receptors and matrix
metalloproteinase-2 and -9 in 30 cases of hepatocellular carcinoma
compared with adjacent tissues as predictors of cancer invasion and
staging. Exp Ther Med. 9:905–908. 2015.PubMed/NCBI
|
|
28
|
Ogunwobi OO and Liu C: Therapeutic and
prognostic importance of epithelial-mesenchymal transition in liver
cancers: Insights from experimental models. Crit Rev Oncol Hematol.
83:319–328. 2012. View Article : Google Scholar
|
|
29
|
Yang JD, Nakamura I and Roberts LR: The
tumor microenvironment in hepatocellular carcinoma: Current status
and therapeutic targets. Semin Cancer Biol. 21:35–43. 2011.
View Article : Google Scholar :
|
|
30
|
Hou KZ, Fu ZQ and Gong H: Chemokine ligand
20 enhances progression of hepatocellular carcinoma via
epithelial-mesen-chymal transition. World J Gastroenterol.
21:475–483. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamada S, Okumura N, Wei L, Fuchs BC,
Fujii T, Sugimoto H, Nomoto S, Takeda S, Tanabe KK and Kodera Y:
Epithelial to mesenchymal transition is associated with shorter
disease-free survival in hepatocellular carcinoma. Ann Surg Oncol.
21:3882–3890. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhai X, Zhu H, Wang W, Zhang S, Zhang Y
and Mao G: Abnormal expression of EMT-related proteins, S100A4,
vimentin and E-cadherin, is correlated with clinicopathological
features and prognosis in HCC. Med Oncol. 31:9702014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang H and Zhou Q: E-cadherin/beta-catenin
and the invasion and metastasis of lung cancer. Zhongguo Fei Ai Za
Zhi. 13:254–259. 2010.In Chinese. PubMed/NCBI
|
|
34
|
Wang TH, Lin YS, Chen Y, Yeh CT, Huang YL,
Hsieh TH, Shieh TM, Hsueh C and Chen TC: Long non-coding RNA AOC4P
suppresses hepatocellular carcinoma metastasis by enhancing
vimentin degradation and inhibiting epithelial-mesenchymal
transition. Oncotarget. 6:23342–23357. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou Q, Lui VW and Yeo W: Targeting the
PI3K/Akt/mTOR pathway in hepatocellular carcinoma. Future Oncol.
7:1149–1167. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lang Q and Ling C: MiR-124 suppresses cell
proliferation in hepatocellular carcinoma by targeting PIK3CA.
Biochem Biophys Res Commun. 426:247–252. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan H, Wang S, Yu H, Zhu J and Chen C:
Molecular pathways and functional analysis of miRNA expression
associated with paclitaxel-induced apoptosis in hepatocellular
carcinoma cells. Pharmacology. 92:167–174. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chetty C, Lakka SS, Bhoopathi P and Rao
JS: MMP-2 alters VEGF expression via alphaVbeta3 integrin-mediated
PI3K/AKT signaling in A549 lung cancer cells. Int J Cancer.
127:1081–1095. 2010. View Article : Google Scholar :
|
|
39
|
Shan RF, Zhou YF, Peng AF and Jie ZG:
Inhibition of Aurora-B suppresses HepG2 cell invasion and migration
via the PI3K/Akt/NF-κB signaling pathway in vitro. Exp Ther Med.
8:1005–1009. 2014.PubMed/NCBI
|
|
40
|
Xu L, Zhang Y, Wang H, Zhang G, Ding Y and
Zhao L: Tumor suppressor miR-1 restrains epithelial-mesenchymal
transition and metastasis of colorectal carcinoma via the MAPK and
PI3K/AKT pathway. J Transl Med. 12:2442014. View Article : Google Scholar : PubMed/NCBI
|