Introduction
Osteocalcin is a non-collagenous vitamin K-dependent
protein, which is secreted particularly by osteoblasts, and its
forms include carboxylated osteocalcin and uncarboxylated
osteocalcin (1). The role played
by osteocalcin in the skeleton has not yet been fully elucidated.
Hoang et al indicated that carboxylated osteocalcin bound
with high affinity to the mineral component of bone,
hydroxyapatite, and regulated mineralization (2). Certain studies have also suggested
that carboxylated osteocalcin has no effect on mineralization
(3–5). Others have provided evidence that
carboxylated osteocalcin may play a role in bone turnover (6,7).
Moreover, serum concentrations of osteocalcin have been established
as the main marker of bone turnover (8). However, it remains unknown as to
whether uncarboxylated osteocalcin influences osteoblast
proliferation and differentiation.
Previous research has demonstrated that
uncarboxylated osteocalcin is involved in energy metabolism.
Fukumoto and Martin revealed that bone, as an endocrine organ, used
the osteoblast-specific secreted molecule, osteocalcin, to promote
glucose homeostasis (9).
Subsequently, Ferron et al provided evidence that daily
injections of uncarboxylated osteocalcin improved glucose handling
and prevented the development of type 2 diabetes in mice (10). In vitro studies have also
demonstrated that uncarboxylated osteocalcin regulates glucose
homeostasis by increasing insulin secretion and sensitivity
(11,12). In another study, Pi et al
reported that uncarboxylated osteocalcin mediated insulin secretion
by β-cells through G protein-coupled receptor, class C, group 6,
member A (GPRC6A) and regulated glucose metabolism (12). GPRC6A, a G protein-coupled
receptor, is an osteocalcin receptor which exists in osteoblasts
(13,14). In addition, GPRC6A is widely
expressed in brain and peripheral tissues with highest levels being
noted in the kidneys, skeletal muscle, testes and leucocytes
(14).
Hyperglycemia, as a characteristic of diabetes
mellitus, has been suggested to be a potential contributor to
diabetes-related osteoporosis (15,16). Epidemiological studies have
theorized that patients with diabetes have an increased risk of
developing osteoporosis or osteoporotic fractures (17,18). Evidence from rodent models and
clinical research has indicated that dysfunctions in glucose
metabolism are associated with a higher risk of bone loss and
fractures (19). In addition,
hyperglycemia impairs bone formation and function in both types 1
(20) and 2 diabetes (21). The results of our previous studies
demonstrated that high glucose levels inhibited osteogenic
differentiation and promoted adipogenic differentiation in primary
osteoblasts (22) and MG-63 cells
(23). In clinical studies, it
was noted that the serum osteocalcin concentrations were reduced in
patients with diabetes (24,25). However, to the best of our
knowledge, whether uncarboxylated osteocalcin promotes the
proliferation and differentiation of osteoblasts under high glucose
conditions has not been investigated to date.
In the present study, we examined the effects of
uncarboxylated osteocalcin on the proliferation and differentiation
of osteoblasts under high glucose conditions. Our results revealed
that uncarboxylated osteocalcin prevented the activation of
PI3K/Akt signaling, inhibited high glucose induced-reactive oxygen
species (ROS) production and stimulated the osteoblastic
differentiation of MC3T3-E1 cells. However, we found that
uncarboxylated osteocalcin had no effect on osteoblastic
differentiation under normal conditions.
Materials and methods
Preparation of recombinant
osteocalcin
The bacterial expression and purification of
recombinant mouse uncarboxylated osteocalcin were performed as
previously described (26).
Briefly, His-tagged osteocalcin (GE Healthcare, Uppsala, Sweden)
fusion protein was bacterially produced and purified on
His-sepharose (GE Healthcare) according to standard procedures. The
purity (>95%) of the osteocalcin preparation was assessed by 15%
SDS-PAGE with Coomassie blue staining. The concentration of the
recombinant osteocalcin protein was precisely determined using
mouse osteocalcin enzyme-linked immunosorbent assay (ELISA)
according to manufacturer's instructions (Immunotopics, San
Clemente, CA, USA).
Cell culture
MC3T3-E1 cells were purchased from The Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). The MC3T3-E1 cells were cultured in α-MEM
(HyClone, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco, Grand Island, NY, USA) and 1% penicillin-streptomycin
in 5% CO2 at 37°C.
Cell proliferation assay
In the present study, a Cell Counting Kit-8 (CCK-8)
was used to examine the effects of uncarboxylated osteocalcin on
the proliferation of osteoblasts under high glucose conditions.
Briefly, the cells were plated in 96-well plates (3,000 cells/well)
in medium containing 10% serum for 24 h and then replaced with 4%
serum medium containing normal (5.5 mM) or high levels of glucose
(25.5 mM) and treated with various concentrations of uncarboxylated
osteocalcin (0.3, 3, 10 and 30 ng/ml) for 1, 2 or 3 days. The
absorbance values were read at a 450 nm wavelength using an
automated micro-plate reader (Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
Measurement of ROS production
The production of ROS was quantified using the
peroxide-dependent oxidation of 2′,7′-dichlorofluorescein diacetate
(DCFH-DA) (Beyotime Institute of Biotechnology, China).
Intracellular esterases hydrolyze DCFH-DA and form non-fluorescent
DCFH, which is further oxidized by ROS to form the fluorescent
compound, DCF. The MC3T3-E1 cells were plated in 24-well plates
with 10% FBS. After the cells had attached, the medium was replaced
with new medium containing 4% FBS and normal (5.5 mM) or high (25.5
mM) amounts of glucose, as well as with or without the antioxidant,
N-acetyl-L-cysteine (NAC), and uncarboxylated osteocalcin (3 ng/ml)
for 48 h. Subsequently, DCFH-DA (final concentration of 10 mM) was
added to the cells followed by incubation at 37°C for 30 min.
Finally, the cells were digested for the detection of ROS levels
using a BioTek Synergy HT microplate reader with the excitation set
at 485 nm and the emission at 528 nm (BioTek Instruments Inc.,
Winooski, VT, USA.
Assay of mineralization
The MC3T3-E1 cells were seeded at a density of
1×106 cells/well in 6-well plates with α-MEM containing
10% FBS and grown until they reached confluence. The medium was
then replaced with mineralizion medium (α-MEM, 4% FBS, 50
µg/ml ascorbic acid, 10 mM β-glycerophosphate phosphate and
1×10−8 M dexamethasone; Sigma, St. Louis, MO, USA)
containing normal (5.5 mM) or high (25.5 mM) amounts of glucose and
either with or without the antioxidant, NAC, and uncarboxylated
osteocalcin (3 ng/ml). The mineralizing medium was replaced every
day. Calcified nodule formation was detected by Alizarin red S
staining on day 21. Briefly, the cells were washed twice with PBS
and were then fixed with 10% formaldehyde for 30 min at room
temperature. The cells were then stained with 2% Alizarin Red S (pH
4.2; Sigma) for 20 min at room temperature and thoroughly rinsed
with water. For quantification, the bound stain was eluted with 10%
wt/vol) cetylpyridinium chloride, and the absorbance of
supernatants was measured at 550 nm using a microplate reader
(Bio-Rad Laboratories, Inc.).
Lipid droplet assay
Oil Red O staining (Sigma) was used to analyze the
formation of lipid droplets. The MC3T3-E1 cells were plated at a
density of 1×106 cells in 6-well plates with α-MEM
containing 10% FBS and then treated with adipogenic differentiation
medium (α-MEM, 4% FBS, 10 µg/ml insulin and
1×10−7 M dexamethasone) containing normal (5.5 mM) or
high (25.5 mM) levels of glucose, and either with or without the
antioxidant, NAC, and uncarboxylated osteocalcin (3 ng/ml).
Fourteen days later, the formation of lipid droplets was determined
by staining with freshly diluted Oil Red O solution (0.1% Oil Red
O:water, 3:2) for 10 min. Images were observed under an inverted
phase contrast microscope (IMT-2-21; Olympus, Tokyo, Japan). The
optical density at 510 nm was measured to quantify lipid
accumulation in osteoblasts.
Alkaline phosphatase (ALP) assay
The MC3T3-E1 cells were cultured in α-MEM with 10%
FBS and the medium was then changed to new medium with 4% FBS,
which contained normal (5.5 mM) or high (25.5 mM) levels of glucose
and inhibitors [LY294002 (Sigma); PI3K inhibitor], as well as with
or without the antioxidant, NAC, and uncarboxylated osteocalcin (3
ng/ml). ALP activity in the culture supernatants was measured using
an ALP assay kit (Nanjing Jiancheng Bioengineering Institute,
Nanjing, China) according to the manufacturer's instructions.
Enzyme-linked immunosorbent assay for
type I collagen secretion
The cultured cells were treated as above, and
collagen I (Col I) secretion was quantified using commercial ELISA
kits (Boster Biological Technology, Wuhan, China) according to the
manufacturer's instructions.
Reverse transcription-quantitative PCR
(RT-qPCR)
To detect the expression of osteogenic [runt-related
transcription factor 2 (Runx2), osterix and osteocalcin] and
adipogenic markers [peroxisome proliferator-activated receptor γ
(PPARγ), adipocyte fatty acid-binding protein (adipocyte Protein 2;
aP2) and fatty acid synthase (FAS)], total RNA was extracted using
an RNeasy mini spin column (Qiagen, Inc., Valencia, CA, USA), and 2
µg RNA was reverse transcribed into cDNA using the SuperScript
First-Stand Synthesis system for RT-PCR (Invitrogen, Carlsbad, CA,
USA). Quantitative PCR (qPCR) was performed using 2 µl of
cDNA in a 10 µl reaction volume with an ABI Gene Amp 5700
Sequence Detection System and QuantiTect SYBR-Green PCR Master mix
(Qiagen, Inc.). The forward and reverse primer sequences are listed
in Table I. The relative mRNA
levels were normalized using β-actin as a housekeeping gene. The
cycling conditions included incubation at 95°C for 5 min, 40 cycles
of denaturation at 94°C for 30 sec, annealing at 56°C for 45 sec
and a final extension at 72°C for 30 sec. We compared each gene
sample level using ABI Gene Amp 5700 SDS software.
 | Table IPrimer sequences used for
RT-qPCR. |
Table I
Primer sequences used for
RT-qPCR.
| Gene | Forward primers
(5′→3′) | Reverse primers
(5′→3′) |
|---|
| Runx2 |
GCCTTCAAGGTGGTAGCCC |
CGTTACCCGCCATGACAGTA |
| Osx |
ACTGGCTAGGTGGTGGTCAG |
GGTAGGGAGCTGGGTTAAGG |
| OC |
GCAATAAGGTAGTGAACAGACTCC |
AGCAGGGTTAAGCTCACACTG |
| PPARγ |
GCATGGTGCCTTCGCTGA |
TGGCATCTCTGTGTCAACCATG |
| aP2 |
ACACCGAGATTTCCTTCAAACTG |
CCATCTAGGGTTATGATGCTCTTC |
| FAS |
GGCTGCAGTGAATGAATTTG |
TTCGTACCTCCTTGGCAAAC |
| β-actin |
GCTCTTTTCCAGCCTTCCTT |
AGGTCTTTACGGATGTCAACG |
Western blot analysis
The cells were washed twice with ice-cold PBS and
scraped on ice into RIPA lysis buffer. The lysates were cleared by
centrifugation (12,000 rpm) at 4°C for 15 min. An equivalent amount
of protein was electrophoresed on a 10% SDS-PAGE gel and
transferred onto PVDF membranes (Millipore Corp., Bedford, MA,
USA). The membranes were blocked in TBS buffer containing 5% skim
milk for 1 h at room temperature, and the membranes were then
incubated overnight at 4 °C with a primary monoclonal antibody
against p-Akt (Ser473) or Akt (1:1,000 dilution) (Cell Signaling
Technology, Beverly, MA, USA), followed by a horseradish peroxidase
(HRP) conjugated secondary antibody (1:5,000 dilution) (Cell
Signaling Technology) for 1 h at room temperature after being
washed 3 times with TBST for 10 min each. Subsequently, the
membranes were washed 3 times with TBST, and the immunoreactive
bands were visualized using an enhanced chemiluminescence (ECL) kit
(Biomiga, Inc., San Diego, CA, USA). The intensity of the protein
bands was quantified by densitometric scanning using ImageJ
software and normalized to β-actin.
Statistical analysis
All experiments were performed at least 3 times.
Data are presented as the means ± SD. The results were analyzed by
one way analysis of variance (ANOVA) followed by Bonferroni's
multiple comparison tests using statistical software SPSS 13.0. A
p-value <0.05 was considered to indicate a statistically
significant difference.
Results
High glucose inhibits osteoblast
proliferation but this effect is reversed by treatment with
uncarboxylated osteocalcin in MC3T3-E1 cells
In order to determine whether uncarboxylated
osteocalcin influences the proliferation of MC3T3-E1 cells under
high glucose conditions, various concentrations of uncarboxylated
osteocalcin (0.3, 3, 10 and 30 ng/ml) were added to the culture
medium containing high levels of glucose (25 mM). CCK-8 assay was
used to examine cell viability. As shown in Fig. 1A, high glucose levels inhibited
the growth of osteoblasts, whereas treatment with various
concentrations of uncarboxylated osteocalcin reversed this effect.
Our results indicated that uncarboxylated osteocalcin attenuated
the high glucose-induced inhibition of osteoblast proliferation. In
addition, we selected the concentration of 3 ng/ml uncarboxylated
osteocalcin as the treatment dose for subsequent experiments.
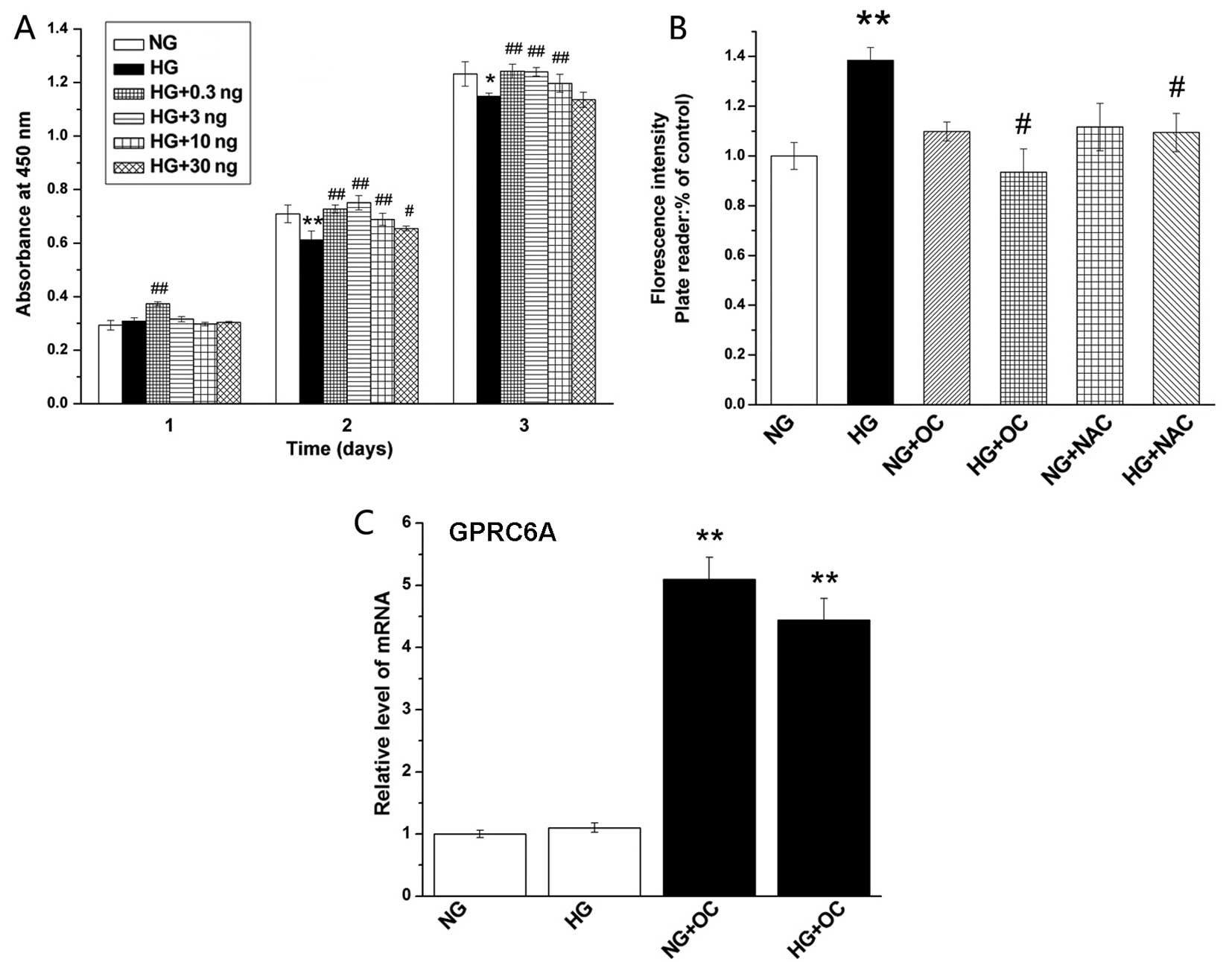 | Figure 1Uncarboxylated osteocalcin increases
osteoblast proliferation and reduces high glucose-induced reactive
oxygen species (ROS) production in MC3T3-E1 cells. (A)
Uncarboxylated osteocalcin increased the proliferation of MC3T3-E1
cells under high glucose conditions. Cells were exposed to normal
glucose (5.5 mM) or high glucose (25.5 mM) medium containing
various concentrations of uncarboxylated osteocalcin (0.3, 3, 10
and 30 ng/ml) for 1, 2 or 3 days, and cell growth was measured by
CCK-8 assay. (B) Cells were cultured in the presence of 5.5 mM
(normal glucose, NG), 25.5 mM glucose (high glucose, HG) as well as
with or without the antioxidant, N-acetyl-L-cysteine (NAC), and
uncarboxylated osteocalcin (3 ng/ml), and ROS levels were detected
by DCFH-DA staining. (C) Uncarboxylated osteocalcin stimulates the
expression of G protein-coupled receptor, class C, group 6, member
A (GPRC6A). Results are presented as the means ± SD, n=3
experiments, *p<0.05, **p<0.01, vs. NG
and #p<0.05, ##p<0.01 vs. HG. |
Uncarboxylated osteocalcin inhibits high
glucose-induced ROS production and stimulates the osteoblast
differentiation, but inhibits the adipogenic differentiation of
MC3T3-E1 cells Uncarboxylated osteocalcin inhibits high
glucose-induced ROS production
It has previously been noted that high
glucose-induced ROS production inhibits the osteogenic
differentiation and promotes the adipogenic differentiation of
primary osteoblasts (22). In
this study, in order to determine whether uncarboxylated
osteocalcin reverses the effects of ROS (induced by high glucose)
on osteoblast differentiation, we measured intracellular ROS levels
by DCFH-DA staining. Our results revealed that the intracellular
ROS level was significantly reduced by treatment with
uncarboxylated osteocalcin (3 ng/ml) or NAC (ROS scavenger) in the
cells exposed to high glucose (25.5 mM) compared to the cells
treated only with high glucose (Fig.
1B). These results suggest that uncarboxylated osteocalcin
reduces ROS levels induced by high glucose.
Uncarboxylated osteocalcin increases the
expression of GPRC6A
Osteoblasts contain the osteocalcin receptor GPRC6A.
To determine the expression of GPRC6A, RT-qPCR was performed. The
results revealed that the expression level of GPRC6A was
significantly enhanced by uncarboxylated osteocalcin (Fig. 1C).
High glucose-induced ROS production
inhibits mineralization and facilitates lipid droplet formation,
but these effects are reversed by treatment with uncarboxylated
osteocalcin
Both the images and the quantification of the
Alizarin red staining demonstrated that high glucose-induced ROS
production significantly inhibited the formation of mineralized
nodules in MC3T3-E1 cells under high glucose conditions. However,
these effects were reversed by treatment with NAC, as well as
uncarboxylated osteocalcin (Fig. 2A
and C). In addition, Oil Red O staining demonstrated that lipid
droplet accumulation was significantly increased by high
glucose-induced ROS production, and this effect was also reversed
by treatment with NAC or uncarboxylated osteocalcin (Fig. 2B and D). However, we also noted
that uncarboxylated osteocalcin had no effect on cells cultured
under normal glucose conditions. These results demonstrated that
uncarboxylated osteocalcin promoted the formation of mineralized
nodules and decreased lipid droplet accumulation in MC3T3-E1 cells
under high glucose conditions.
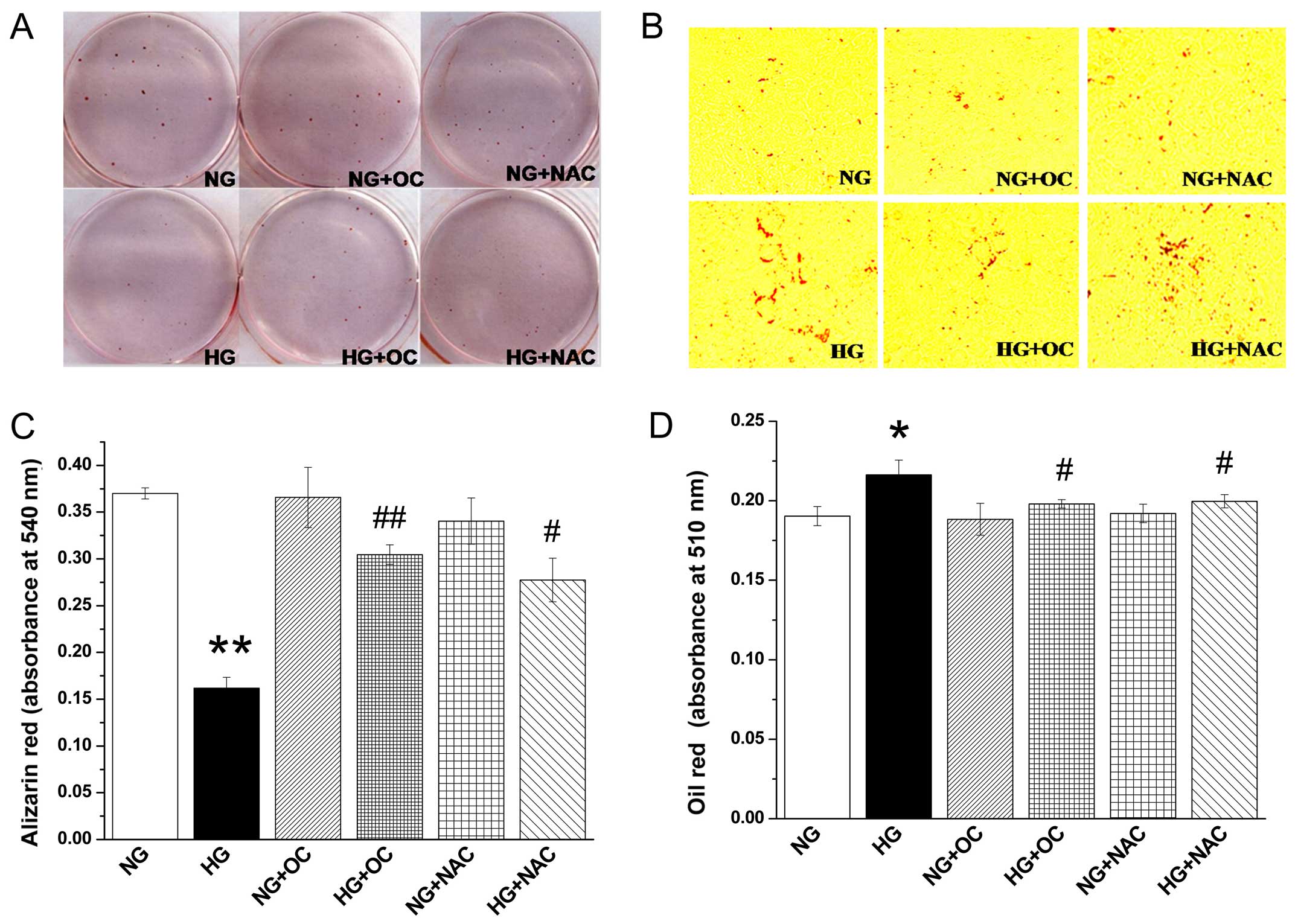 | Figure 2Uncarboxylated osteocalcin reduces
high glucose-induced reactive oxygen species (ROS) production to
increase mineralization and inhibit lipid droplet accumulation in
MC3T3-E1 cells. Cells were cultured in inductive medium containing
normal levels of glucose (5.5 mM, NG) or high glucose (25.5 mM,
HG), with or without the antioxidant, N-acetyl-L-cysteine (NAC),
and uncarboxylated osteocalcin (3 ng/ml). (A) Alizarin red staining
and (C) quantification showed that high glucose inhibited
mineralized bone nodules. (B) Oil Red O staining and (D)
quantification showed that high glucose-induced ROS promoted lipid
droplet formation. Results are presented as the means ± SD, n=3
experiments, *p<0.05, **p<0.01, vs. NG
and #p<0.05, ##p<0.01, vs. HG. |
High glucose-induced ROS production
inhibits the expression of osteogenic genes and promotes the
expression of adipogenic genes, but these effects are reversed by
treatment with uncarboxylated osteocalcin
RT-qPCR was performed to further characterize the
gene expression levels of the osteogenic markers, Runx2, osterix
and osteocalcin, and the gene expression of the adipogenic markers,
PPARγ, aP2, and FAS in MC3T3-E1 cells under high glucose
conditions. The results revealed that high glucose-induced ROS
production increased the mRNA expression of adipogenic markers and
inhibited the expression of osteogenic markers in MC3T3-E1 cells
under high glucose conditions; these effects were reversed by
treatment with NAC and uncarboxylated osteocalcin. However,
uncarboxylated osteocalcin did not have such a marked effect on
cells cultured under normal glucose conditions (Fig. 3). The above results indicated that
uncarboxylated osteocalcin promoted the expression of osteogenic
genes and inhibited the expression of adipogenic genes.
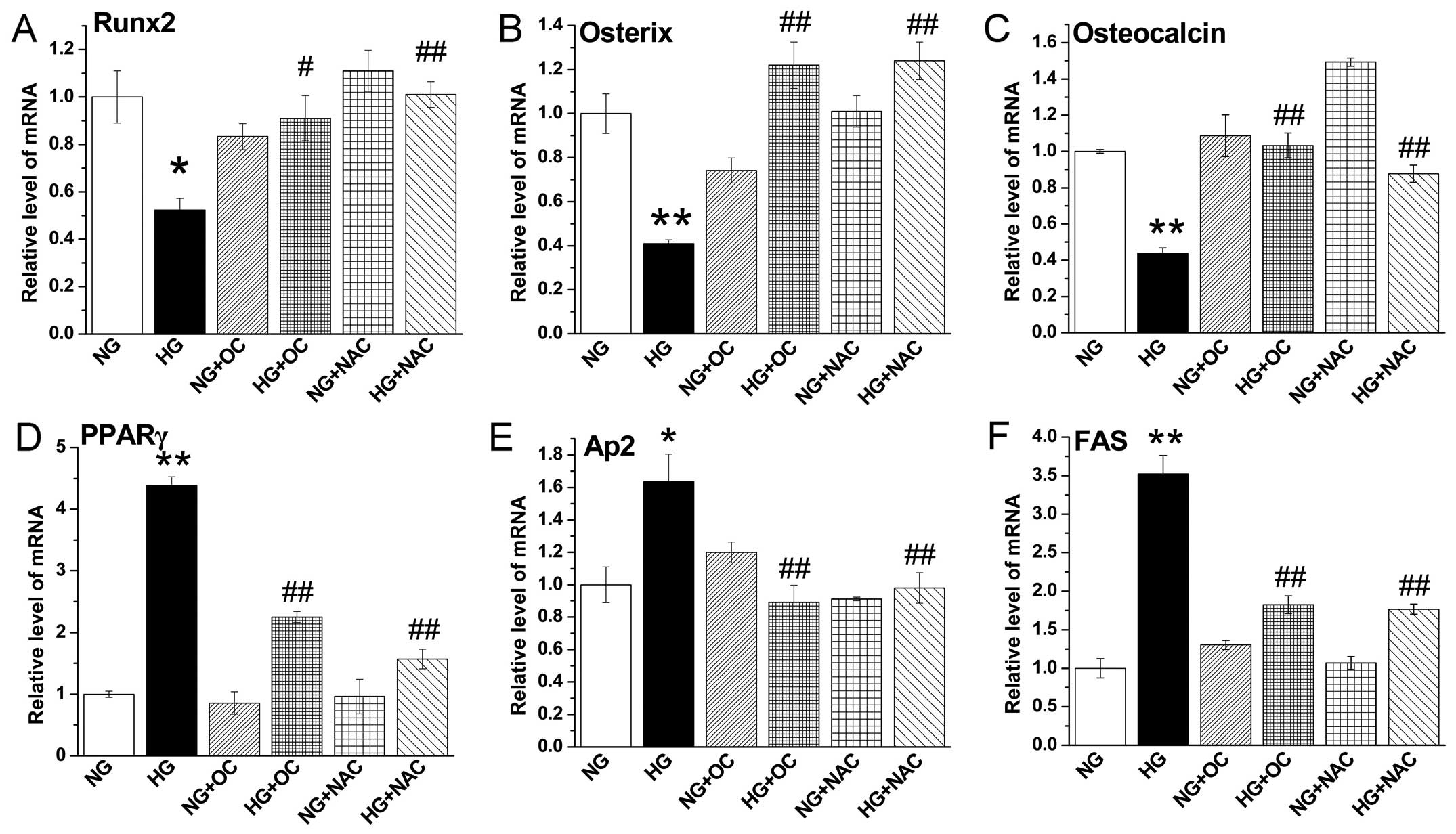 | Figure 3Uncarboxylated osteocalcin reduces
high glucose-induced reactive oxygen species (ROS) production to
increase the expression of osteogenic genes and inhibit the
expression of adipogenic genes. RT-qPCR was performed to quantify
the expression of the osteogenic genes, (A) runt-related
transcription factor 2 (Runx2), (B) osterix and (C) osteocalcin,
and the adipogenic genes, (D) peroxisome proliferator-activated
receptor γ (PPARγ), (E) adipocyte protein 2 (aP2) and (F) fatty
acid synthase (FAS). The expression of target genes was normalized
to β-actin gene expression. Results are presented as the means ±
SD, n=3 experiments, *p<0.05, **p<0.01
vs. normal glucose (NG), and #p<0.05,
##p<0.01 vs. high glucose (HG). |
High glucose-induced ROS production
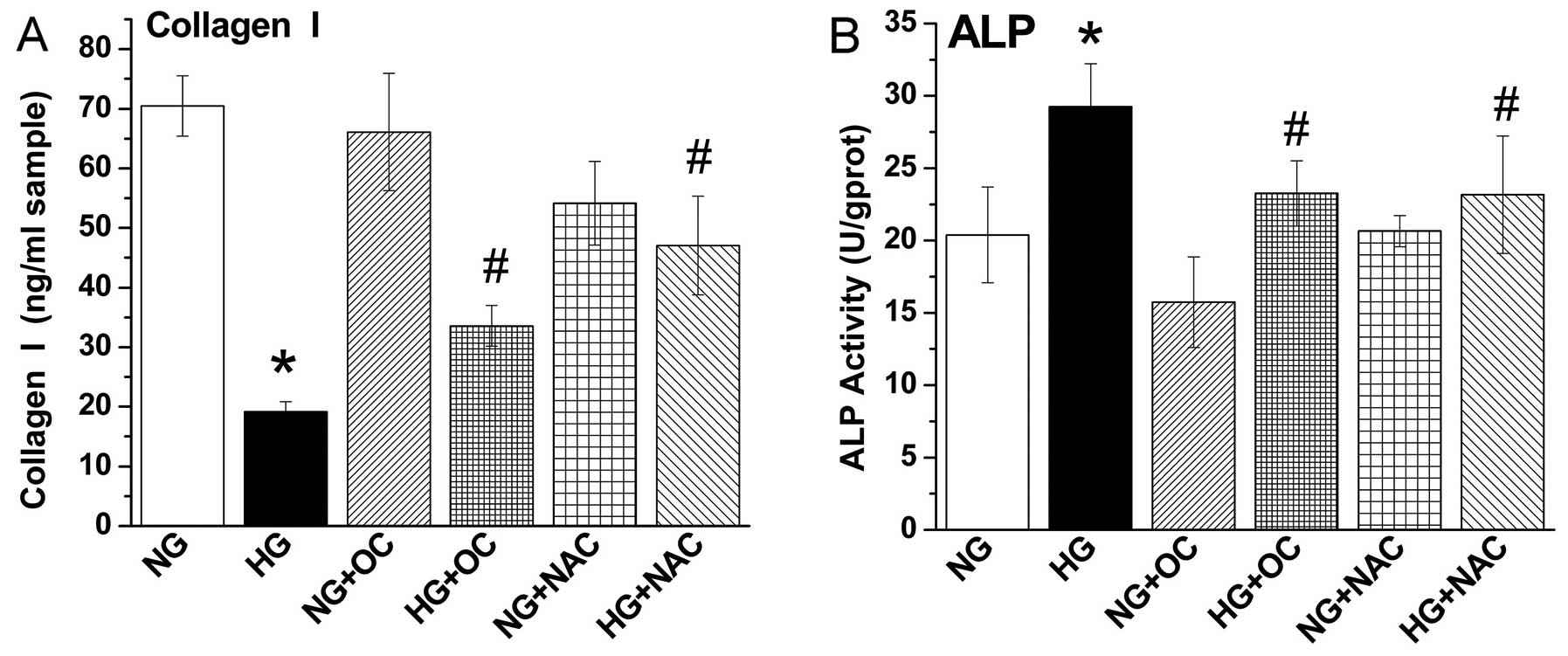
decreases (Col I) protein levels and increases ALP activity, but
these effects are reversed by treatment with uncarboxylated
osteocalcin
Subsequently, we examined the effects of
uncarboxylated osteocalcin on the Col I protein levels and ALP
activity in MC3T3-E1 cells cultured under high glucose conditions.
The results revealed that high glucose levels increased ALP
activity and decreased the protein levels of Col I, and treatment
with NAC or uncarboxylated osteocalcin also reversed these effects
(Fig. 4). No such phenomenon was
observed in the MC3T3-E1 cells cultured under normal glucose
conditions. Taken together, our results indicated that
uncarboxylated osteocalcin reduced ROS levels and promoted the
osteogenic differentiation and inhibited the adipogenic
differentiation of MC3T3-E1 cells under high glucose
conditions.
High glucose-induced ROS production
inhibits osteoblast differentiation and increases adipogenic
differentiation via the activation of the PI3K/Akt signal pathway,
but these effects are reversed by treatment with uncarboxylated
osteocalcin in MC3T3-E1 cells Uncarboxylated osteocalcin inhibits
the activation of the PI3K/Akt signaling pathway by high
glucose-induced ROS production
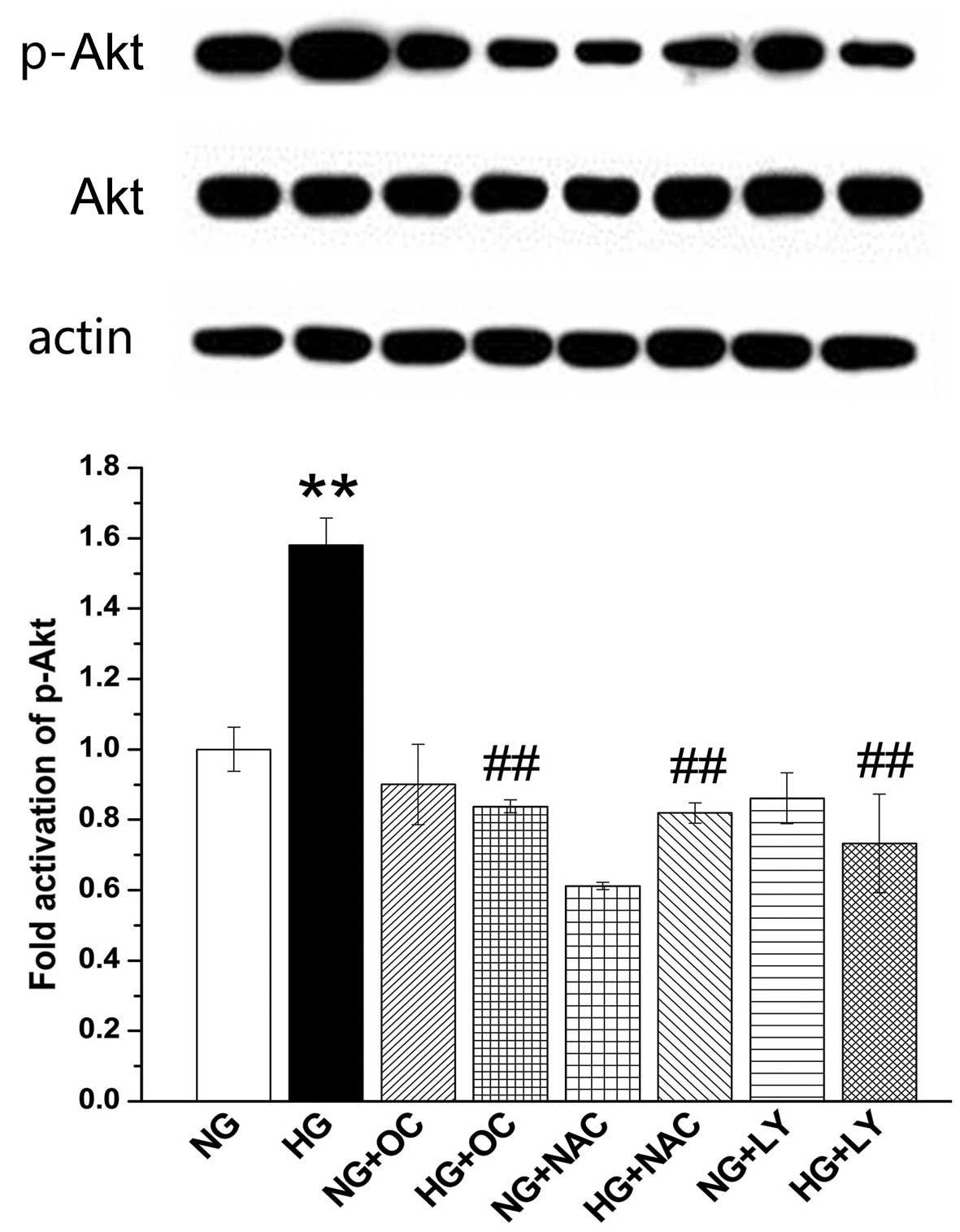
In order to further determine whether uncarboxylated
osteocalcin inhibits ROS-induced PI3K/Akt signaling, we measured
the phosphorylation level of PI3K/Akt with or without LY-294002 (a
PI3K/Akt specific inhibitor), NAC, as well as uncarboxylated
osteocalcin. As shown in Fig. 5,
the intracellular p-Akt level increased significantly under high
glucose conditions but was reduced by the addition of LY294002, NAC
and uncarboxylated osteocalcin. These results suggest that
uncarboxylated osteocalcin inhibits PI3K/Akt signaling by reducing
ROS levels.
Uncarboxylated osteocalcin inhibits the
PI3K/Akt signaling pathway to promote the expression of osteogenic
genes and inhibit the expression of adipogenic genes
We measured the expression levels of osteogenic
differentiation- and adipogenic differentiation-related genes. Our
results demonstrated that LY294002 or uncarboxylated osteocalcin
also reversed the effects of high glucose on the expression of
marker genes of osteogenic and adipogenic differentiation in
MC3T3-E1 cells. We did not observe the same phenomenon in the cells
cultured under normal glucose conditions (Fig. 6).
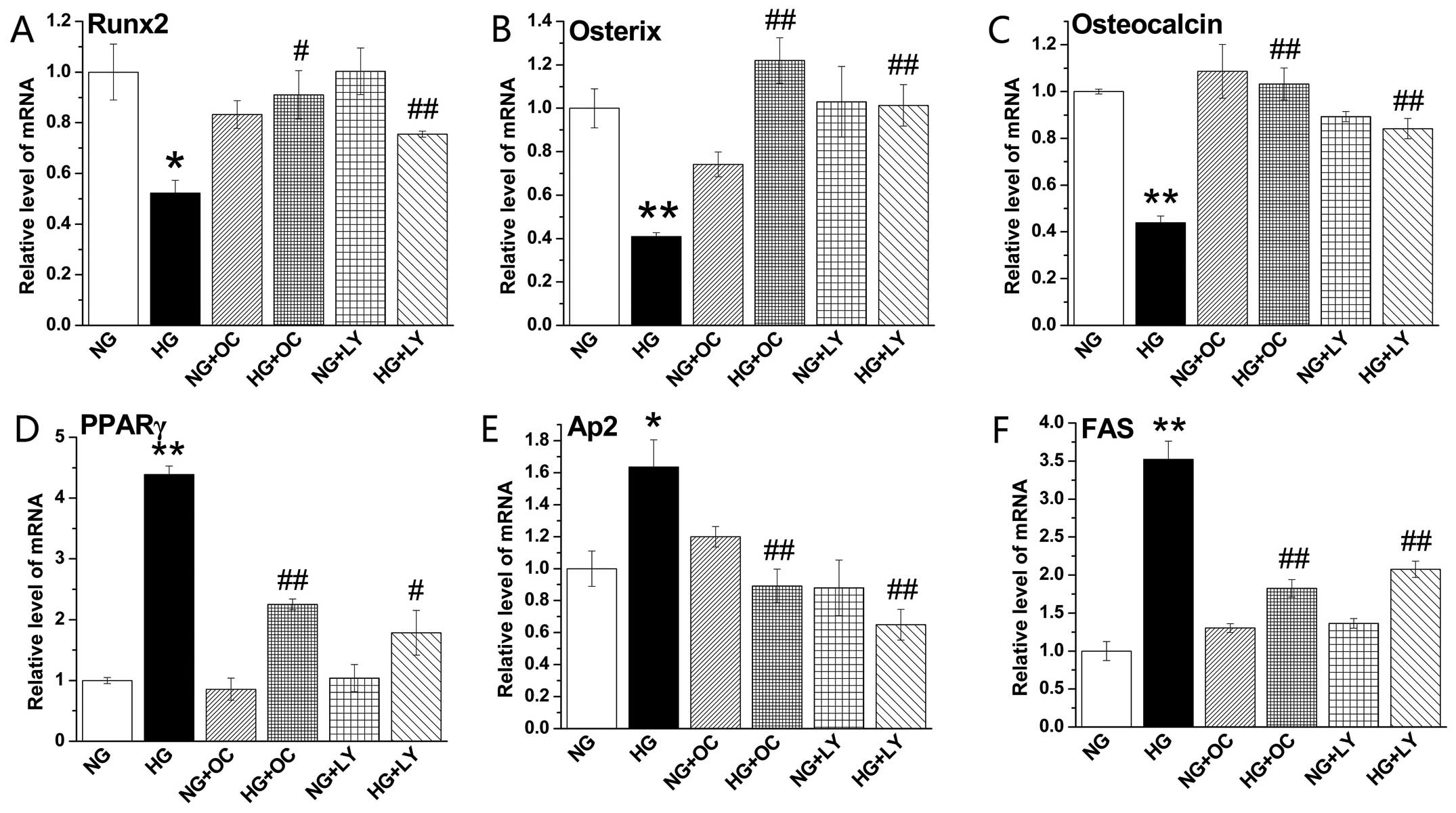 | Figure 6Uncarboxylated osteocalcin increases
the expression of osteogenic genes and inhibits the expression of
adipogenic genes by blocking the activation of the PI3K/Akt pathway
induced by reactive oxygen species (ROS) production in MC3T3-E1
cells under high glucose conditions. Cells were cultured in normal
and high glucose medium with or without the specific PI3K
inhibitor, LY294002, and N-acetyl-L-cysteine (NAC) as well as
uncarboxylated osteocalcin (3 ng/ml). RT-qPCR was performed to
quantify the expression of the osteogenic genes, (A) runt-related
transcription factor 2 (Runx2), (B) osterix and (C) osteocalcin,
and the adipogenic genes, (D) peroxisome proliferator-activated
receptor γ (PPARγ), (E) adipocyte protein 2 (aP2) and (F) fatty
acid synthase (FAS). The expression of target genes was normalized
to β-actin gene expression. Results are presented as the means ±
SD, n=3 experiments, *p<0.05, **p<0.01
vs. normal glucose (NG), and #p<0.05,
##p<0.01 vs. high glucose (HG). |
Uncarboxylated osteocalcin inhibits the
PI3K/Akt signaling pathway to increase Col I protein levels and
decrease ALP activity
Subsequently, we examined ALP activity and Col I
levels. The results revealed that the increase in ALP activity and
the decrease in Col I levels in the cells cultured with high
concentrations of glucose (25.5 mM) were reversed by the addition
of LY294002 or uncarboxylated osteocalcin. By contrast, this did
not occur in the cells cultured with normal levels of glucose (5.5
mM) (Fig. 7). These results
suggest that uncarboxylated osteocalcin facilitates osteogenic
differentiation and inhibits adipogenic differentiation by blocking
ROS-stimulated PI3K/Akt signaling in MC3T3-E1 cells under high
glucose conditions.
Discussion
Previous research in the field of skeletal biology
has revealed that the differentiation and functions of the two
bone-specific cell types, osteoblasts and osteoclasts, are
determined by secreted molecules that can either be cytokines
acting locally, or hormones acting systemically (27,28). In the present study, we
demonstrated that uncarboxylated osteocalcin promoted osteogenic
differentiation and inhibited adipogenic differentiation by
blocking the ROS-induced activation of the PI3K/Akt pathway in
MC3T3-E1 cells under high glucose conditions, manifested by an
increased expression of the osteogenic markers, Runx2, osterix and
osteocalcin, and the formation of mineralized nodules, as well as
the decreased mRNA expression of the adipogenic markers, PPARγ, aP2
and FAS, and decreased lipid drop accumulation.
Uncarboxylated osteocalcin and carboxylated
osteocalcin are two different forms of osteocalcin. Previous
research has demonstrated that in osteocalcin-deficient mice, lower
carbonate:phosphate ratios were noted in variectomized knockout
cortices than wild-type ones, and crystallite size and perfection
resembled that in wild-type trabeculae, providing evidence that
osteocalcin regulates bone mineralization (29). Recently, Poundarik et al
confirmed that osteocalcin−/− mice had lower crystal
thickness than wild-type mice, in a small angle X-ray scattering
(SAXS) study (30). However, as
shown in our study in Figs.
1Figure 2Figure 3–4, our results proved that uncarboxylated
osteocalcin did not influence the proliferation, differentiation
and mineralization of MC3T3-E1 cells cultured under normal glucose
conditions.
Previous research has demonstrated that high glucose
not only inhibits the proliferation and function (31), but also decreases the osteogenic
differentiation and mineralization (32) of osteoblasts. Our previous studies
also found that high amounts of glucose inhibited the osteogenic
differentiation of MG-63 cells (23) and high glucose-induced ROS
production inhibited the osteogenic, but promoted the adipogenic
differentiation of rat primary osteoblasts (22). Oxidative stress has been
acknowledged as a major regulator of mineral tissue homeostasis and
the promotion of bone resorption (33–36). It has previously been suggested
that oxidative stress leads to bone pathogenesis, including
osteoporosis, bone tumor development and diabetes-induced bone
complications (37). In addition,
Liu et al demonstrated that the presence of metallothionein,
a ROS inhibitor, restored cell differentiation, suggesting that
oxidative stress plays a crucial role in the inhibition of
osteoblastic differentiation (38). In the present study, our results
revealed that uncarboxylated osteocalcin reduced high
glucose-induced ROS production and increased the osteogenic
differentiation, but decreased the adipogenic differentiation of
MC3T3-E1 cells under high glucose conditions.
However, the molecular mechanisms responsible for
the effects of uncarboxylated osteocalcin on the differentiation of
osteoblasts under high glucose conditions have yet to be
elucidated. It has previously been shown that high glucose
increases PPARγ expression and PI3K activity and the
phosphorylation of its downstream effector Akt during adipogenesis
(39). In this study, we also
proved that high glucose-induced ROS production accelerated the
adipogenic and weakened the osteogenic differentiation of primary
osteoblasts through the PI3K-Akt pathway. As shown in Fig. 5, high glucose increased the
phosphorylation level of PI3K/Akt, which was reversed by
uncarboxylated osteocalcin, LY-294002, as well as NAC in MC3T3-E1
cells under high glucose conditions. These results suggested that
uncarboxylated osteocalcin blocked PI3K-Akt signaling induced by
ROS and upregulated the expression of osteogenic markers, increased
Col I protein levels, as well as mineralization, but downregulated
the expression of adipogenic markers and lipid droplet
formation.
Overall, the findings of our study demonstrated that
uncarboxylated osteocalcin inhibited high glucose induced-ROS
production and stimulated osteoblast differentiation by preventing
the activation of PI3K/Akt in MC3T3-E1 cells. These results provide
new insight into the molecular mechanisms involved in the
regulation of glucose metabolism by uncarboxylated osteocalcin.
These findings suggest that uncarboxylated osteocalcin may be a
potential therapeutic agent for use in the treatment of
diabetes-related osteoporosis.
Acknowledgments
The present study was supported by the Knowledge
Innovation Program of the Chinese Academy of Sciences (nos.
KSCX2-EW-J-29 and Y129015EA2).
References
|
1
|
Neve A, Corrado A and Cantatore FP:
Osteocalcin: skeletal and extra-skeletal effects. J Cell Physiol.
228:1149–1153. 2013. View Article : Google Scholar
|
|
2
|
Hoang QQ, Sicheri F, Howard AJ and Yang
DSC: Bone recognition mechanism of porcine osteocalcin from crystal
structure. Nature. 425:977–980. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Karsenty G, Kronenberg HM and Settembre C:
Genetic control of bone formation. Annu Rev Cell Dev Biol.
25:629–648. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ducy P, Desbois C, Boyce B, Pinero G,
Story B, Dunstan C, Smith E, Bonadio J, Goldstein S, Gundberg C, et
al: Increased bone formation in osteocalcin-deficient mice. Nature.
382:448–452. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Murshed M, Schinke T, McKee MD and
Karsenty G: Extracellular matrix mineralization is regulated
locally; different roles of two gla-containing proteins. J Cell
Biol. 165:625–630. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dubois-Ferrière V, Brennan TC, Dayer R,
Rizzoli R and Ammann P: Calcitropic hormones and IGF-I are
influenced by dietary protein. Endocrinology. 152:1839–1847. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rutter MM, Markoff E, Clayton L, Akeno N,
Zhao G, Clemens TL and Chernausek SD: Osteoblast-specific
expression of insulin-like growth factor-1 in bone of transgenic
mice induces insulin-like growth factor binding protein-5. Bone.
36:224–231. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garnero P: Biomarkers for osteoporosis
management: utility in diagnosis, fracture risk prediction and
therapy monitoring. Mol Diagn Ther. 12:157–170. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fukumoto S and Martin TJ: Bone as an
endocrine organ. Trends Endocrinol Metab. 20:230–236. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ferron M, McKee MD, Levine RL, Ducy P and
Karsenty G: Intermittent injections of osteocalcin improve glucose
metabolism and prevent type 2 diabetes in mice. Bone. 50:568–575.
2012. View Article : Google Scholar
|
|
11
|
Lee NK, Sowa H, Hinoi E, Ferron M, Ahn JD,
Confavreux C, Dacquin R, Mee PJ, McKee MD, Jung DY, et al:
Endocrine regulation of energy metabolism by the skeleton. Cell.
130:456–469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pi M, Wu Y and Quarles LD: GPRC6A mediates
responses to osteocalcin in β-cells in vitro and pancreas in vivo.
J Bone Miner Res. 26:1680–1683. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bodine PVN and Komm BS: Evidence that
conditionally immortalized human osteoblasts express an osteocalcin
receptor. Bone. 25:535–543. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wellendorph P and Bräuner-Osborne H:
Molecular cloning, expression, and sequence analysis of GPRC6A, a
novel family C G-protein-coupled receptor. Gene. 335:37–46. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schwartz AV: Diabetes mellitus: does it
affect bone? Calcif Tissue Int. 73:515–519. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McCabe LR: Understanding the pathology and
mechanisms of type I diabetic bone loss. J Cell Biochem.
102:1343–1357. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hamann C, Kirschner S, Günther KP and
Hofbauer LC: Bone, sweet bone - osteoporotic fractures in diabetes
mellitus. Nat Rev Endocrinol. 8:297–305. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hayakawa N and Suzuki A: Diabetes mellitus
and osteoporosis. Effect of antidiabetic medicine on osteoporotic
fracture. Clin Calcium. 22:1383–1390. 2012.In Japanese. PubMed/NCBI
|
|
19
|
Motyl KJ, McCabe LR and Schwartz AV: Bone
and glucose metabolism: a two-way street. Arch Biochem Biophys.
503:2–10. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thrailkill KM, Liu L, Wahl EC, Bunn RC,
Perrien DS, Cockrell GE, Skinner RA, Hogue WR, Carver AA, Fowlkes
JL, et al: Bone formation is impaired in a model of type 1
diabetes. Diabetes. 54:2875–2881. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hamann C, Goettsch C, Mettelsiefen J,
Henkenjohann V, Rauner M, Hempel U, Bernhardt R, Fratzl-Zelman N,
Roschger P, Rammelt S, et al: Delayed bone regeneration and low
bone mass in a rat model of insulin-resistant type 2 diabetes
mellitus is due to impaired osteoblast function. Am J Physiol
Endocrinol Metab. 301:E1220–E1228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y and Yang JH: Activation of the
PI3K/Akt pathway by oxidative stress mediates high glucose-induced
increase of adipogenic differentiation in primary rat osteoblasts.
J Cell Biochem. 114:2595–2602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang W, Zhang X, Zheng J and Yang J: High
glucose stimulates adipogenic and inhibits osteogenic
differentiation in MG-63 cells through cAMP/protein kinase
A/extracellular signal-regulated kinase pathway. Mol Cell Biochem.
338:115–122. 2010. View Article : Google Scholar
|
|
24
|
Movahed A, Larijani B, Nabipour I,
Kalantarhormozi M, Asadipooya K, Vahdat K, Akbarzadeh S, Farrokhnia
M, Assadi M, Amirinejad R, et al: Reduced serum osteocalcin
concentrations are associated with type 2 diabetes mellitus and the
metabolic syndrome components in postmenopausal women: The
crosstalk between bone and energy metabolism. J Bone Miner Metab.
30:683–691. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou M, Ma X, Li H, Pan X, Tang J, Gao Y,
Hou X, Lu H, Bao Y and Jia W: Serum osteocalcin concentrations in
relation to glucose and lipid metabolism in Chinese individuals.
Eur J Endocrinol. 161:723–729. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim JH, Park S, Kim HW and Jang JH:
Recombinant expression of mouse osteocalcin protein in Escherichia
coli. Biotechnol Lett. 29:1631–1635. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Harada S and Rodan GA: Control of
osteoblast function and regulation of bone mass. Nature.
423:349–355. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Teitelbaum SL and Ross FP: Genetic
regulation of osteoclast development and function. Nat Rev Genet.
4:638–649. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Boskey AL, Gadaleta S, Gundberg C, Doty
SB, Ducy P and Karsenty G: Fourier transform infrared
microspectroscopic analysis of bones of osteocalcin-deficient mice
provides insight into the function of osteocalcin. Bone.
23:187–196. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Poundarik A, Gundberg C and Vashishth D:
Non-collageneous proteins influence bone mineral size, shape and
orientation: a SAXS study. J Bone Miner Res. 26(Suppl):
S362011.
|
|
31
|
Zhen D, Chen Y and Tang X: Metformin
reverses the deleterious effects of high glucose on osteoblast
function. J Diabetes Complications. 24:334–344. 2010. View Article : Google Scholar
|
|
32
|
Fujimori S, Osawa M, Iemata M, Hinoi E and
Yoneda Y: Increased GABA transport activity in rat calvarial
osteoblasts cultured under hyperglycemic conditions. Biol Pharm
Bull. 29:297–301. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koh JM, Lee YS, Kim YS, Kim DJ, Kim HH,
Park JY, Lee KU and Kim GS: Homocysteine enhances bone resorption
by stimulation of osteoclast formation and activity through
increased intracellular ROS generation. J Bone Miner Res.
21:1003–1011. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bai XC, Lu D, Liu AL, Zhang ZM, Li XM, Zou
ZP, Zeng WS, Cheng BL and Luo SQ: Reactive oxygen species
stimulates receptor activator of NF-kappaB ligand expression in
osteoblast. J Biol Chem. 280:17497–17506. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee NK, Choi YG, Baik JY, Han SY, Jeong
DW, Bae YS, Kim N and Lee SY: A crucial role for reactive oxygen
species in RANKL-induced osteoclast differentiation. Blood.
106:852–859. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wittrant Y, Gorin Y, Woodruff K, Horn D,
Abboud HE, Mohan S and Abboud-Werner SL: High d(+)glucose
concentration inhibits RANKL-induced osteoclastogenesis. Bone.
42:1122–1130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wauquier F, Leotoing L, Coxam V, Guicheux
J and Wittrant Y: Oxidative stress in bone remodelling and disease.
Trends Mol Med. 15:468–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu AL, Zhang ZM, Zhu BF, Liao ZH and Liu
Z: Metallothionein protects bone marrow stromal cells against
hydrogen peroxide-induced inhibition of osteoblastic
differentiation. Cell Biol Int. 28:905–911. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chuang CC, Yang RS, Tsai KS, Ho FM and Liu
SH: Hyperglycemia enhances adipogenic induction of lipid
accumulation: involvement of extracellular signal-regulated protein
kinase 1/2, phosphoinositide 3-kinase/Akt, and peroxisome
proliferator-activated receptor gamma signaling. Endocrinology.
148:4267–4275. 2007. View Article : Google Scholar : PubMed/NCBI
|