Introduction
Osteoporosis and falls are related to fractures,
which can lead to increased morbidity and mortality, as well as
decreased functional ability. The mortality of patients with hip
fractures within 1 year is 20%, and only one-third of these
patients recover their original functions (1). Diabetes mellitus (DM) is an
endocrine metabolic dysfunctional disease, which is frequently
accompanied by osteoporosis (2).
DM results in hyperglycemia due to an absolute insulin
insufficiency or insulin resistance, which leads to the development
of a number of complications, both micro-vascular and macrovascular
pathological changes (3).
Clinical surveys have indicated that DM enhances the risk of hip
fractures, vertebral fractures, proximal humeral fractures, tibia
fractures, and wrist and ankle fractures, independent of bone
mineral density (BMD) (4,5). Moreover, DM has been shown to be
associated with the delayed healing of fractures and bone
deteriorations in animal models (6,7).
In the KK-Ay mouse, a classic animal model of obesity with T2DM,
high insulin levels increased cortical bone mass and impaired the
trabecular microstructure by upregulating osteoblast-and
osteoclast-related gene expression (8). In vitro, high glucose (16.5
mM) has been shown to inhibit the osteogenic differentiation
potential and proliferation of rat osteoblasts; moreover, high
glucose significantly impairs bone formation by inhibiting
osteoblast proliferation and differentiation (9,10).
Although the in vitro and in vivo contribution of
glucose to bone metabolism has been investigated, the signaling
mechanisms responsible for high glucose-induced bone deteriorations
have not yet been well characterized.
Bone is remodeled constantly throughout life by
bone-resorbing osteoclasts and bone-forming osteoblasts. To
maintain bone volume and quality, the differentiation of
osteoclasts and osteoblasts is tightly regulated through
communication between and within these two cell lineages (11). Bone cells, such as chondrocytes,
osteoblasts, osteocytes and osteoclasts, express a variety of
Ephrin ligands and Eph receptors (12,13). Cultured osteoclasts stimulated
with receptor activator of nuclear factor-κB ligand (RANKL) have
been shown to dynamically express Ephrin ligands (A2, B1 and B2)
and Eph receptors (A1, A2 and A4), as revealed by reverse
transcription-polymerase chain reaction (RT-PCR) (11,14). The protein expression of Ephrin
ligands and Eph receptors was also identified in human mesenchymal
stem cells (MSCs) by western blot analysis and immunohistochemistry
(15). Developmental deficiencies
in the EphB/EphrinB signaling pathway can lead to skeletal
abnormalities, which includes the defective development of the
somite (16), craniofacial
development (17), limb
development (18), and other bone
abnormalities in mutant mice, and in individuals harboring EphrinB1
mutations that cause the X-linked developmental disorder
craniofrontonasal syndrome (19).
The osteoclasts of EphA4 null mice are also larger, and express
higher mRNA levels of matrix metalloproteinase (MMP)3 and MMP9, and
exhibit greater bone resorption activity than wild-type osteoclasts
in vitro (20). A
microarray analysis of bone marrow-derived MSCs (BMSCs) identified
EphA5 as an inhibitory factor candidate, which may be a negative
regulator of bone formation (21). Moreover, cell-cell interactions
mediated by the EphrinB2 ligand on osteoclasts have been shown to
inhibit osteoclast-induced bone resorption, and the EphB4 receptor
on osteoblasts can promote osteoblast-driven bone formation
(22). By contrast, bidirectional
EphrinA2/EphA2 signaling regulates bone remodeling at the
initiation phase through the enhancement of osteoclasto-genesis and
the suppression osteoblastogenesis (11). Although the effects of the
Eph/Ephrin signaling pathway can lead to skeletal abnormalities,
the role of Eph/Ephrin in the process of bone deteriorations
induced by hyperglycemia remains largely unknown.
Previous studies have shown that kininogen
deficiency and bradykinin receptor loss-of-function provide
evidence of an important role of kinin peptides in the regulation
of blood pressure and sodium homeostasis, renal responses to
vasopressin and insulin sensitivity in rodents (23,24). Bradykinin receptor B2 (BK2R) gene
knockout can induce cardiomyopathy in mice, and mice with tissue
kallikrein gene knockout develop impaired flow-induced
vasodilatation (24). In mice,
the knockdown of bradykinin receptor B1 (BK1R; BK1R−/−
mice), has been shown to result in increased bone loss; bone marrow
cells obtained from BK1R−/− mice displayed an enhanced
differentiation ability into functional osteoclasts with consistent
artificial calcium phosphate degradation (25). Intriguingly, the lack of BK1R and
BK2R exacerbates diabetes-associated complications, such as renal
injury and bone mineral loss, and bradykinin receptors play a
protective role in diabetes-related osteoporosis (26).
The above-mentioned data suggest that Eph-Ephrin
bidirectional signaling provides an intriguing explanation for the
cellular and molecular mechanisms which are responsible for
osteoblast-osteoclast coupling and may prove to be a potential
target for the treatment of osteoporosis. Thus, the aim of the
present study was to examine the involvement of the skeletal
Eph/Ephrin signaling pathway and bradykinin receptors in bone
deteriorations induced by hyperglycemia in vitro, as well as
in vivo using an animal model of streptozotocin
(STZ)-induced diabetes.
Materials and methods
Culture of MC3T3 osteoblasts
MC3T3 osteoblasts were obtained from the Chinese
Academy of Sciences (Institute of Shanghai Cell Biology and Chinese
Type Culture Collection, Shanghai, China), and the cells were
maintained in Dulbecco's modified Eagle's medium (DMEM; Invitrogen,
Carlsbad, CA, USA), supplemented with 10% fetal bovine serum (FBS;
HyClone, Logan, UT, USA), 100 µg/ml penicillin and 100
µg/ml streptomycin (Invitrogen) at 37°C in a humidified, 5%
CO2, 95% air atmosphere. The medium was changed to fresh
medium initially after 24 h and then every 2 days.
Animal treatment
Six-week-old male C57BL/6J mice (from the Animal
Center of Xi'an Jiaotong University, Xi'an, China) were allowed to
acclimatize to their environment for 1 week. All experimental
procedures were carried out in accordance with the guidelines of
Xi'an Jiaotong University on Animal Care. All chemicals and
reagents were purchased from Sigma (Oakville, ON, Canada), unless
otherwise stated. The mice were randomly divided into 2 groups as
follows: i) the control group (Cont, n=10); ii) the STZ group
(n=10), where the animals were rendered hyperglycemic by an
intraperitoneal injection of STZ, dissolved in citrate buffer (0.1
M at pH 4.2), at 35 mg/kg body weight for 5 consecutive days. The
mice in the control group was treated with citrate buffer only. All
the mice were sacrificed by cardiac exsanguination under light
ether anesthesia 8 weeks after the STZ injection. The fasting blood
glucose (FBG) levels were measured using a blood glucose monitoring
system (Roche Diagnostics, Manheim, Germany), and the body weight
of the mice was recorded every 2 weeks during the experimental
period.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
The osteoblasts were cultured in 96-well plates (at
103 cells/well). An MTT assay was conducted after 1, 4
or 7 days of exposure to high glucose (16.5 mM) using a cell
proliferation kit (Sigma, Oakville, ON, Canada) according to the
manufacturer's instructions. After 24 h, the culture medium was
replaced with 100 ml of MTT (0.5 mg/ml). The black crystals that
formed after 2–3 h were dissolved with acidified isopropanol and
the absorbance was measured at 570 nm using a microplate reader (MD
SpectraMax M5; Molecular Devices, Sunnyvale, CA, USA).
Quantification of apoptosis by flow
cytometry
Apoptosis was assessed using Annexin V, a protein
that binds to phosphatidylserine (PS) residues which are exposed on
the cell surface of apoptotic cells. The osteoblasts were treated
with the vehicle (normal glucose; 5.5 mM) or high glucose for 4
days. Following treatment, the cells were washed twice with
phosphate-buffered saline (PBS; pH 7.4), and re-suspended in
staining buffer containing 1 µg/ml propidium iodide (PI) and
0.025 µg/ml Annexin V-FITC. Double-labeling was performed at
room temperature for 10 min in the dark prior to flow cytometric
analysis. The osteoblasts were immediately analyzed using FACScan
and the CellQuest program. The quantitative assessment of apoptotic
cells was also assessed by the terminal deoxynucleotidyl
transferase-mediated deoxyuridine triphosphate nick-end labeling
(TUNEL) method, which examines DNA-strand breaks during apoptosis
using a BD ApoAlert™ DNA Fragmentation assay kit. The stained cells
were then analyzed using a flow cytometer (FC500; Beckman Coulter,
Miami, FL, USA).
Concentrations of calcium (Ca) and
creatinine (Cr) in serum and urine
The concentrations of Ca and Cr in the serum and
urine of the mice were measured using standard colorimetric methods
with a microplate reader (Bio-Tek, Winooski, VT, USA). Serum was
collected by cardiac exsanguination under light ether anesthesia.
After 24 h urine was collected by metabolic cages. The level of Ca
in the urine was corrected by the concentration of urine Cr levels.
The serum levels of tartrate-resistant acid phosphatase-5b
(TRACP-5b) and osteocalcin (OCN) were detected using a mouse
bioactive enzyme-linked immunosorbent assay (ELISA; Immutopics,
Inc., San Clemente, CA, USA) with an ELISA reader (MD SpectraMax
M5; Molecular Devices).
Gene silencing by small interfering RNA
(siRNA)
For siRNA experiments, RNA primers complementary to
BK2R were designed and synthesized by the Shanghai Invitrogen
Biotechnology Co. (Shanghai, China). The osteoblasts were
transfected with the annealed RNA primer pair using Lipo fectamine
2000 (Invitrogen) in accordance with the instructions provided by
the manufacturer. Cells transfected with scrambled siRNA served as
controls. The siRNA had the following sequences: BK2R siRNA,
5′-ACGGCCCACCAGUCGCCCCAG-3′; and scrambled siRNA,
5′-UUCGACGGUCAGCACGCGAGC-3′.
Mineralization assay
Mineralized matrix formation was assessed by
Alizarin Red S staining and the Ca45 intake method. The osteoblasts
were fixed in 4% paraformaldehyde and stained with 40 mmol/l
Alizarin Red S (pH 4.2; Sigma-Aldrich, Munich, Germany) for 30 min
at room temperature. The excess dye was removed by washing the
plates with distilled water. The incorporated calcium was eluted
with 100 mmol/l cetylpyridinium chloride and visualized under a
microscope (Leica DM 2500; Leica Microsystems GmbH, Wetzlar,
Germany).
Bone histomorphology
Following fixation in 4% formaldehyde solution for 7
days, the tibias were decalcified in 0.5 M EDTA (pH 8.0) and then
embedded in paraffin using standard histological procedures. After
cardiac exsanguination, the tibias were collected with all soft
tissue removed and stored at −80°C. Sections (4-µm-thick)
were cut and stained with hematoxylin and eosin (H&E), and
visualized under a microscope (Leica DM 2500). TRAP staining was
used for the identification of the osteoclasts according to the
manufacturer's instructions (387-A; Sigma-Aldrich, St. Louis, CA,
USA). The number of osteoclasts was quantified below the growth
plates in the proximal tibial metaphysis, and visualized under a
microscope (Leica DM 2500).
RT-PCR
RNA extraction was performed using TRIzol reagent
according to the manufacturer's instructions (Invitrogen). cDNA
synthesis was performed by reverse transcription reactions with 2
µg of total RNA using moloney murine leukemia virus reverse
transcriptase (Invitrogen) with oligo(dT)15 primers (Fermentas,
Burlington, ON, Canada) as described by the manufacturer. First
strand cDNA served as the template for the regular PCR performed
using a DNA engine (ABI 7300). Glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) as an internal control was used to normalize
the data to determine the relative expression levels of the target
genes. The reaction conditions were set according to the
instructions provided with the kit. The PCR primers used in this
study were as follows: runt-related transcription factor (Runx2)
forward, 5′-CCTGACTCTGCACCAAGTC-3′ and reverse,
5′-GAGGTGGCAGTGTCATCATC-3′; alkaline phosphatase (ALP) forward,
5′-GCTGAACAGGAACAACGTGA-3′ and reverse, 5′-AGACTGCGCCTGGTAGTTGT-3′;
MMP9 forward, 5′-GGTCGGTTCTGACCTTTTGT-3′ and reverse,
5′-TGGTGTCCTCCGATGTAAGA-3′; carbonic anhydrase II (CAII) forward,
5′-TGGTTCACTGGAACACCAAA-3′ and reverse, 5′-AGCAAGGGTCGAAGTTAGCA-3′;
osteoprotegerin (OPG) forward, 5′-TGCTCCTGGCACCTACCTA-3′ and
reverse, 5′-ACTCCTGCTTCACGGACTG-3′; RANKL forward,
5′-GGAAGCGTACCTACAGACTA-3′ and reverse, 5′-AGTACGTCGCATCTTGATCC-3′;
EphB4 forward, 5′-AGTGGCTTCGAGCCATCAAGA-3′ and reverse,
5′-CTCCTGGCTTAGCTTGGGACTTC-3′; EphrinB2 forward,
5′-ACGGTCCAACAAGACGTCCA-3′ and reverse, 5′-GCTGTTGCCATCGGTGCTA-3′;
BK1R forward, 5′-TGGAACCAGTTTAACTGGCC-3′ and reverse,
5′-ATGAAGTCCTCCCAAAAGCA-3′; BK2R forward,
5′-TGTTCGTGAGGACTCCGTGC-3′ and reverse, 5′-GCCAGGATCAGGTCTGCTG-3′;
and GAPDH forward, 5′-CACCATGGAGAAGGCCGGGG-3′ and reverse,
5′-GACGGACACATTGGGGGTAG-3′.
Immunoprecipitation and western blot
analysis
Cell lysates were prepared as previously described
(27). The lysates were then
incubated with the indicated antibodies for 1 h at 4°C. The immune
complexes were precipitated with protein A/G agarose (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) for 1 h at 4°C, washed
extensively with lysis buffer, resolved in 4–20% gradient sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and
analyzed by western blotting. All blots were developed by enhanced
chemiluminesence (Amersham Pharmacia Biotech, Piscataway, NJ, USA).
The cells and tibias were homogenized and extracted in NP-40
buffer, followed by 5–10 min of boiling and centrifugation (12,000
× g, for 15 min at 4°C) to obtain the supernatant. Samples
containing 60 µg of protein were separated on a 10% SDS-PAGE
gel and transferred onto nitrocellulose membranes (Bio-Rad
Laboratories, Hercules, CA, USA). Following saturation with 5%
(w/v) non-fat dry milk in TBS and 0.1% (w/v) Tween-20 (TBST), the
membranes were incubated with the following antibodies: Bcl-2
(sc-7382), BAX (sc-20067), caspase-3 (sc-392736), caspase-8
(sc-81656), BK1R (sc-293196), BK2R (sc-136216), EphB4 (sc-130081)
and EphrinB2 (sc-398735) (Santa Cruz Biotechnology, Inc.) at
dilutions ranging from 1:500 to 1:2,000 at 4°C overnight. Following
3 washes with TBST, the membranes were incubated with secondary
immunoglobulins (Igs; sc-2002) conjugated to IRDye 800CW Infrared
Dye (LI-COR Biotechnology, Lincoln, NE, USA), including donkey
anti-goat IgG and donkey anti-mouse IgG at a dilution of
1:10,000–1:20,000. Following 1 h of incubation at 37°C, the
membranes were washed 3 times with TBST. The blots were visualized
using the Odyssey Infrared Imaging System (LI-COR Biotechnology).
The signals were densitometrically assessed (Odyssey Application
software version 3.0) and normalized to the mouse monoclonal
anti-β-actin antibody (AP0733; Bioworld Technology, Inc., St. Louis
Park, MN, USA).
Statistical analysis
The data from our experiments are reported as the
means ± standard errors of mean (SEM) for each group. All
statistical analyses were performed using GraphPad Prism version
5.0 (GraphPad Software, La Jolla, CA, USA). Inter-group differences
were analyzed by one-way ANOVA, followed by Tukey's multiple
comparison test as a post-test to compare the group means if
overall the P-value was <0.05. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
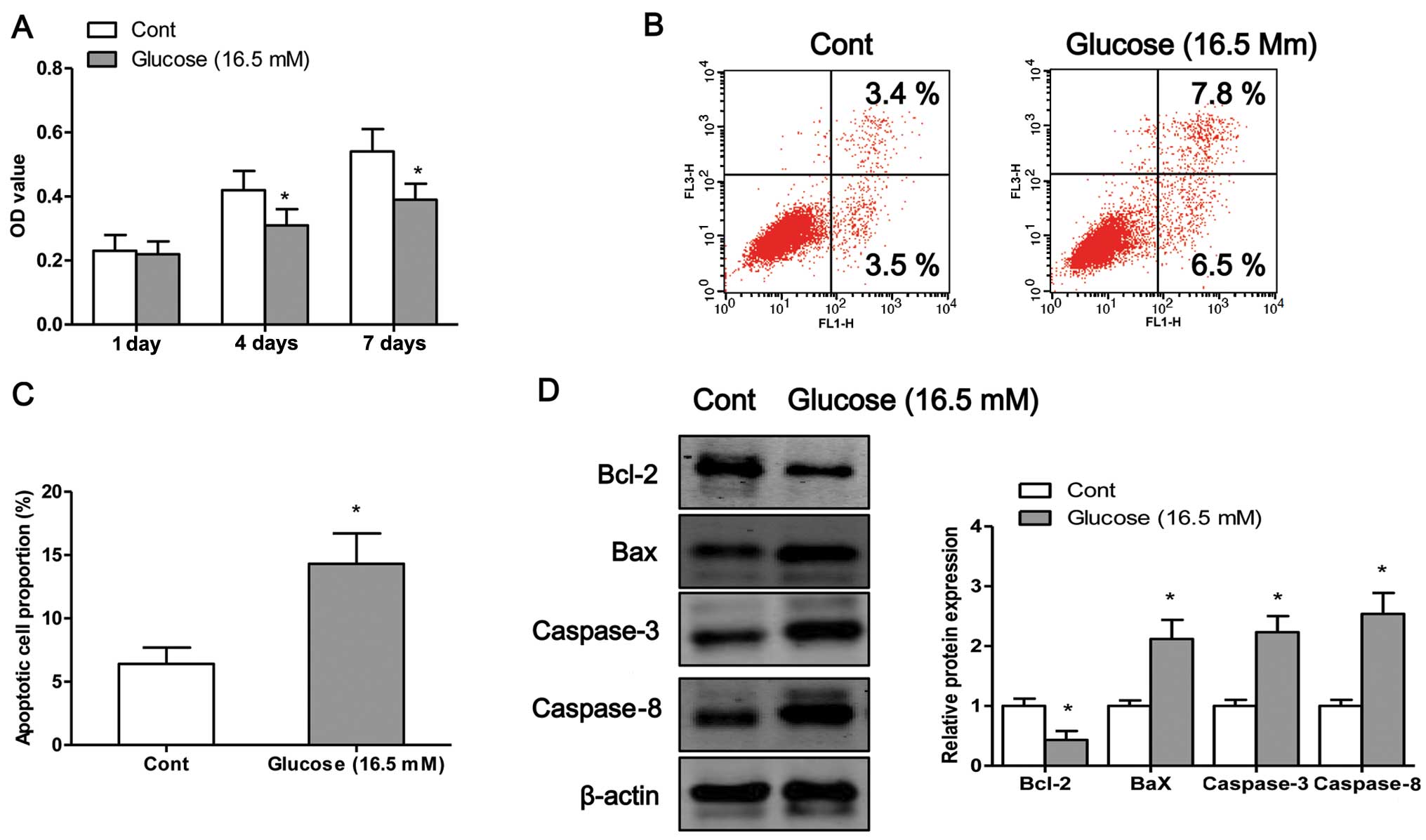
High glucose induces osteoblast
apoptosis
In order to examine the detrimental effects of high
glucose on osteo-blasts, we exposed the osteoblasts to high glucose
(16.5 mM) for different periods of time. We first examined the
viability of the osteoblasts after 1, 4 or 7 days of exposure to
high glucose by MTT assay. The results revealed that exposure to
high glucose induced osteoblast cell death in a time-dependent
manner (Fig. 1A). To determine
whether high glucose induced cell death through an apoptotic
mechanism, Annexin V-PI double-labeling was used for the detection
of the proportion of apoptotic osteoblasts. The apoptotic rate was
significantly increased in the osteoblasts following exposure to
high glucose for 4 days (P<0.05; Fig. 1B and C). Finally, we measured the
expression levels of apoptosis-related proteins. Exposure to high
glucose significantly decreased the protein expression levels of
Bcl-2 and increased the protein expression of Bax, caspase-3 and
caspase-8 compared to the control cells (exposure to normal
glucose) (Fig. 1D).
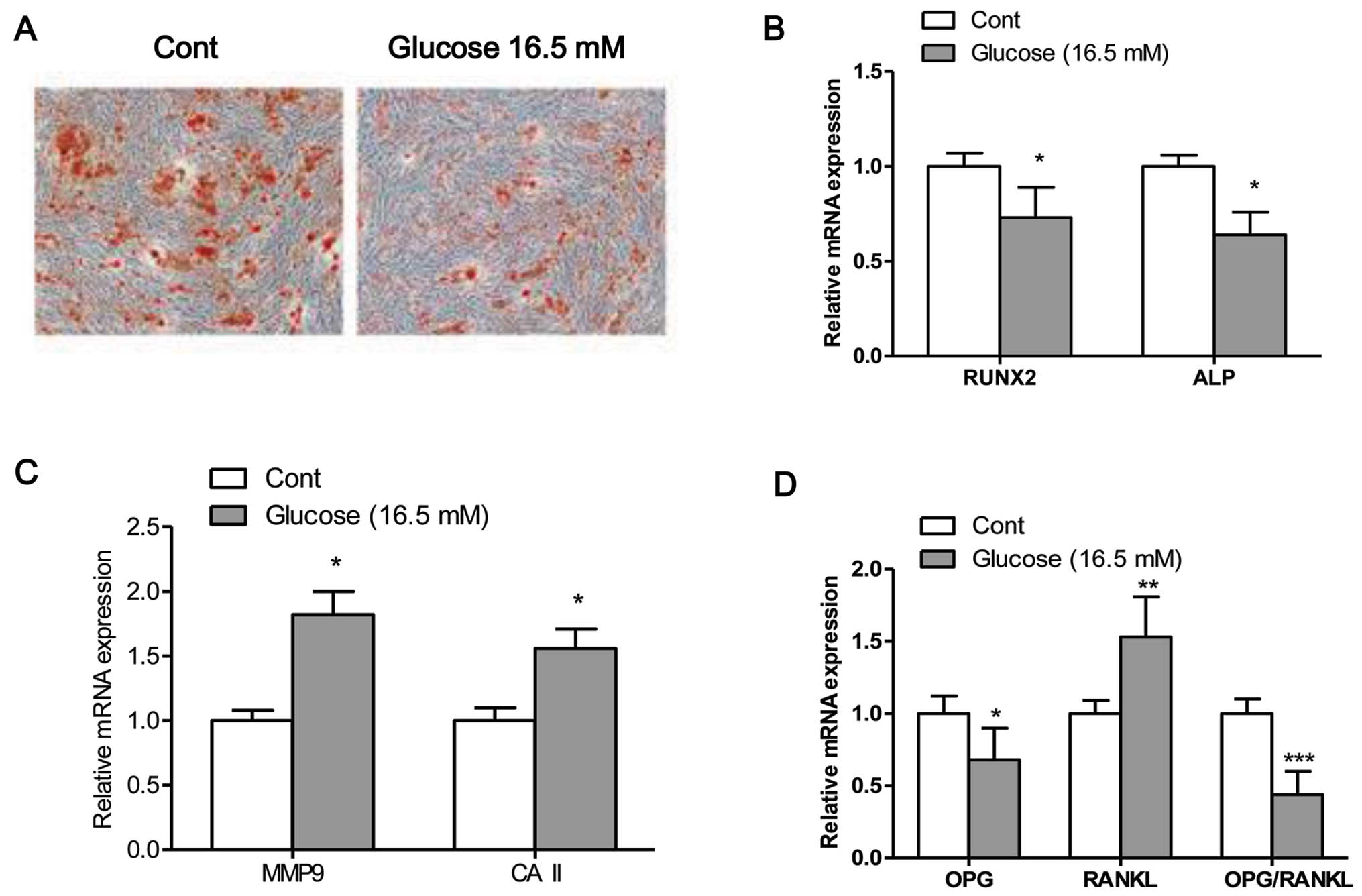
High glucose inhibits osteogenic
differentiation
The mineralization capacity of the osteoblasts was
assessed by Alizarin Red S staining (Fig. 2A), which stains deposited calcium
red. Osteogenic transdifferentiation was inhibited when the
osteo-blasts were exposed to high glucose. Moreover, exposure of
the cells to high glucose significantly decreased the mRNA
expression levels of Runx2 and ALP, and increased the mRNA
expression levels of MMP9 and CAII compared to the control cells
(exposed to normal glucose) (Fig. 2B
and C). The maturation and formation of osteoclasts are mainly
regulated by the balance of extracellular OPG and RANKL, which are
secreted by osteoblasts. Thus, in this study, we measured the
expression levels of OPG and RANKL, and the ratio of OPG/RANKL in
osteoblasts (28). The results
from RT-PCR revealed that the mRNA expression levels of OPG and the
ratio of OPG/RANKL were significantly decreased in the osteoblasts
exposed to high glucose (16.5 mM) compared to the control cells
(P<0.05; Fig. 2D). By
contrast, the mRNA expression of RANKL was significantly higher in
the cells exposed to high glucose compared to the control cells
(P<0.05; Fig. 2D).
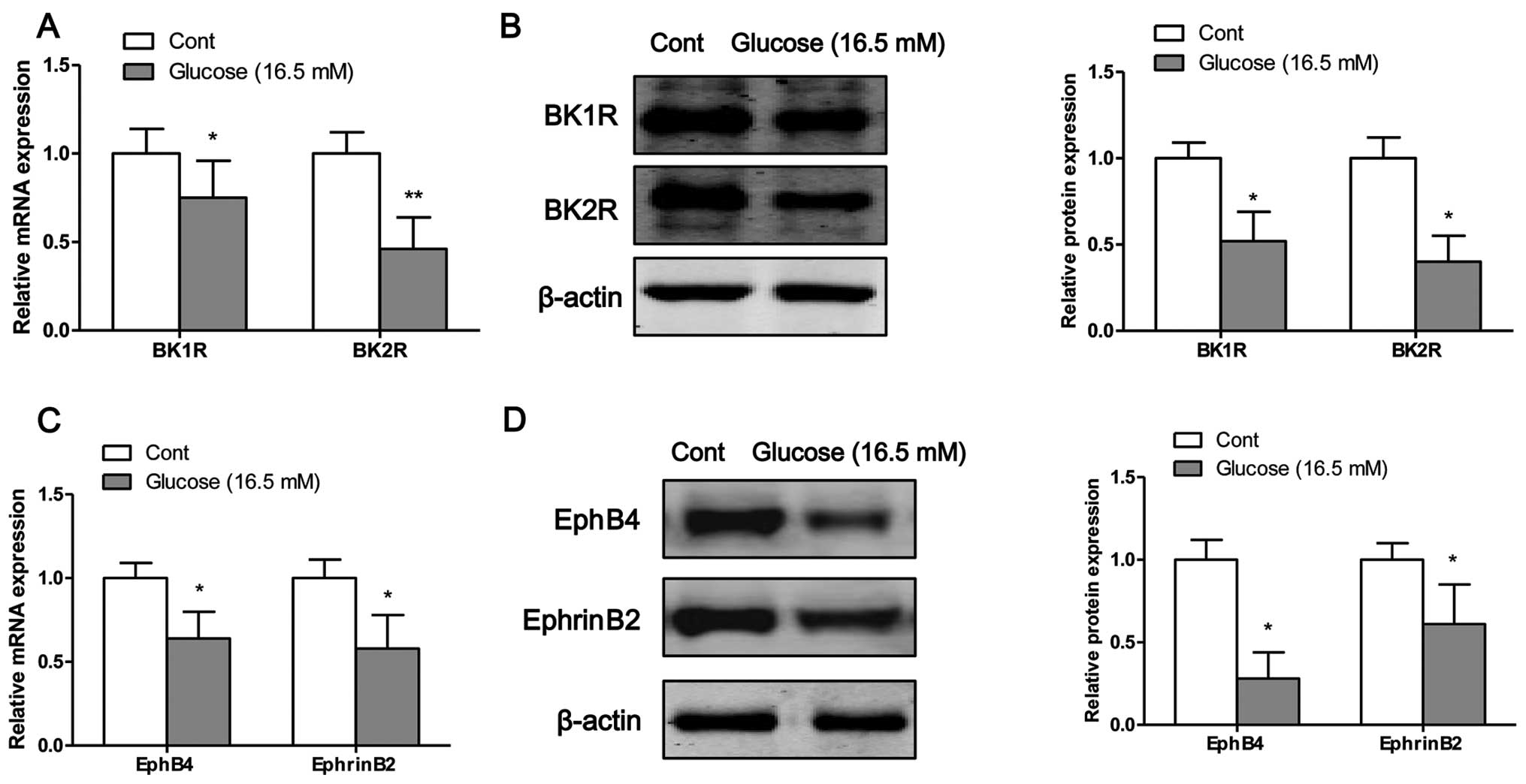
High glucose inhibits the mRNA and
protein expression of bradykinin receptors and EphB4/EphrinB2
The mRNA and protein expression levels of bradykinin
receptors and EphB4/EphrinB2 were measured by RT-PCR and western
blot analysis, respectively. Following exposure of the osteoblasts
to high glucose for 4 days, the mRNA (Fig. 3A) and protein (Fig. 3B) expression levels of BK1R and
BK2R were significantly decreased compared to the control cells
(exposure to normal glucose). Moreover, the mRNA and protein
expression levels of EphB4 and EphrinB2 were significantly
decreased following the exposure of the osteoblasts to high glucose
on the fourth day (Fig. 3C and
D).
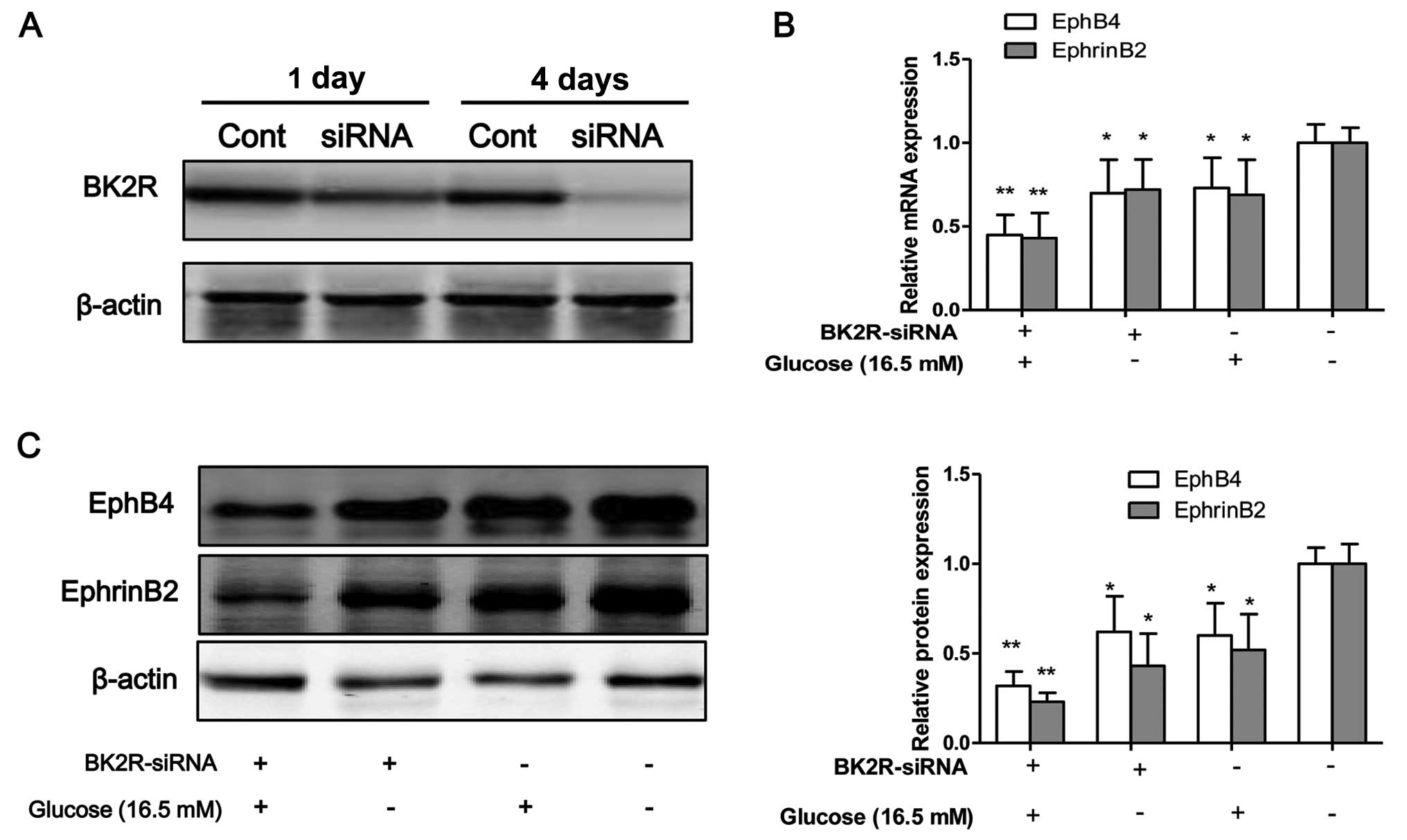
BK2R loss-of-function influences the
expression of EphB4/EphrinB2
To inhibit the function of BK2R, the osteo-blasts
were transfected with siRNA against BK2R. Western blot analysis
revealed that the protein expression levels of BK2R were markedly
decreased following transfection with BK2R siRNA in a
time-dependent manner (Fig. 4A).
Thus, our data suggested that the siRNA experiments were
successfully performed to inhibit the expression of BK2R. BK2R
loss-of-function significantly decreased the mRNA (P<0.05) and
protein (P<0.05) expression of EphB4 and EphrinB2 (Fig. 4B–D). Moreover, BK2R
loss-of-function combined with exposure to high glucose (16.5 mM)
markedly decreased the mRNA (P<0.05) and protein (P<0.05)
expression of EphB4 and EphrinB4 in the osteoblasts compared to the
control cells. We found that the mRNA and protein expression levels
of both EphB4/EphrinB2 and BK2R, were significantly decreased in
the osteoblasts exposed toh high glucose (16.5 mM) in vitro.
To examine whether the interaction between EphB4/EphrinB2 and BK2R
is dependent on high glucose, cell lysates were prepared from high
glucose-exposed cells and immunoprecipitated with anti-BK2R
antibody. The results revealed that the binding capacity between
EphB4/EphrinB2 and BK2R was strong in the control cells, but the
binding capacity was weak in the osteoblasts exposed to high
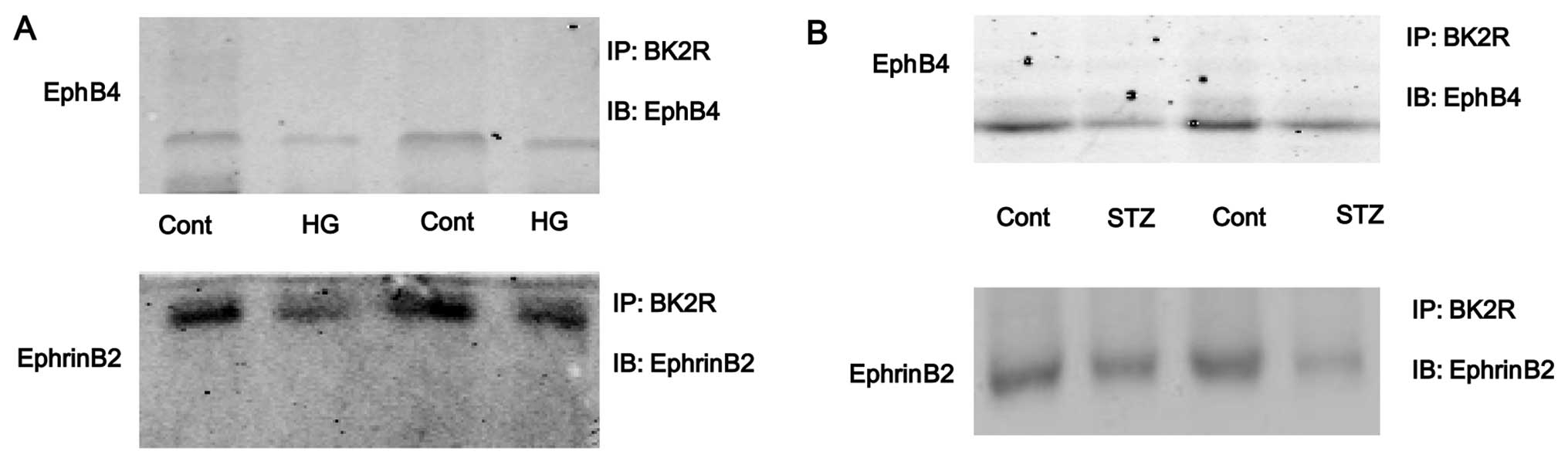
glucose (HG, Fig. 5A). The
interaction between EphB4/EphrinB2 and BK2R was also detected in
the tibias of mice. The results revealed that the binding capacity
between EphB4/EphrinB2 and BK2R was significantly lower in the STZ
group compared with the control group (Fig. 5B). These results suggest that the
interaction between EphB4/EphrinB2 and BK2R may be involved in
STZ-induced diabetes-related osteoporosis.
Basic parameters and biomarkers in serum
and urine of mice
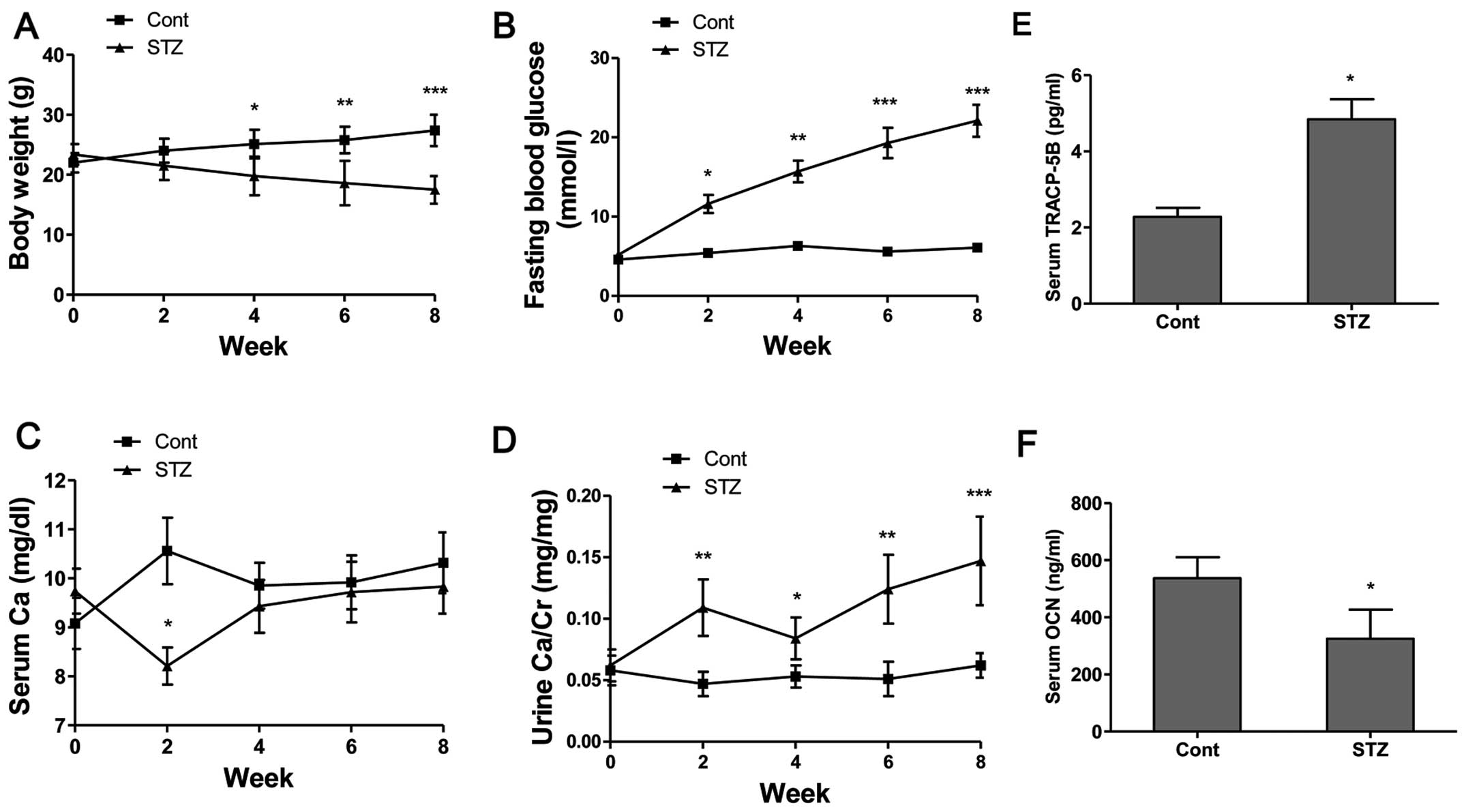
The body weights of the non-diabetic control mice
steadily increased and the FBG levels were maintained within the
normal range of 4–6.1 mmol/l during the experimental period
(Fig. 6A and B). The body weights
of the diabetic mice continued to decrease and the FBG levels
increased following the administration of STZ (Fig. 6A and B). The FBG levels of the
diabetic mice increased from 11.4 mmol/l at week 2 to 21.6 mmol/l
at week 8, indicating a significant increase compared to the
control group (Fig. 6B). The
administration of STZ decreased the serum Ca levels at week 2
(P<0.05); however, the serum Ca level showed no obvious
difference after week 4 compared to the control group (Fig. 6C). Additionally, our results
revealed that the urine Ca level (Fig. 6D) in the STZ group was
significantly increased at week 2 (P<0.01), week 4 (P<0.05),
week 6 (P<0.01) and week 8 (P<0.001) compared to the
non-diabetic control mice. From these calcium metabolic data, it
was demonstrated that hyperglycemia enhanced Ca excretion in mice
with STZ-induced diabetes-related osteoporosis. The serum
concentrations of bone turnover markers, such as TRACP-5b, as a
bone resorption marker, and OCN, as a bone formation marker, were
determined. The results revealed that the serum TRACP-5b levels
(Fig. 6E) in the STZ group were
significantly increased, and the serum OCN levels (Fig. 6F) were significantly decreased
compared to those of the control group.
Bone histology and TRAP staining
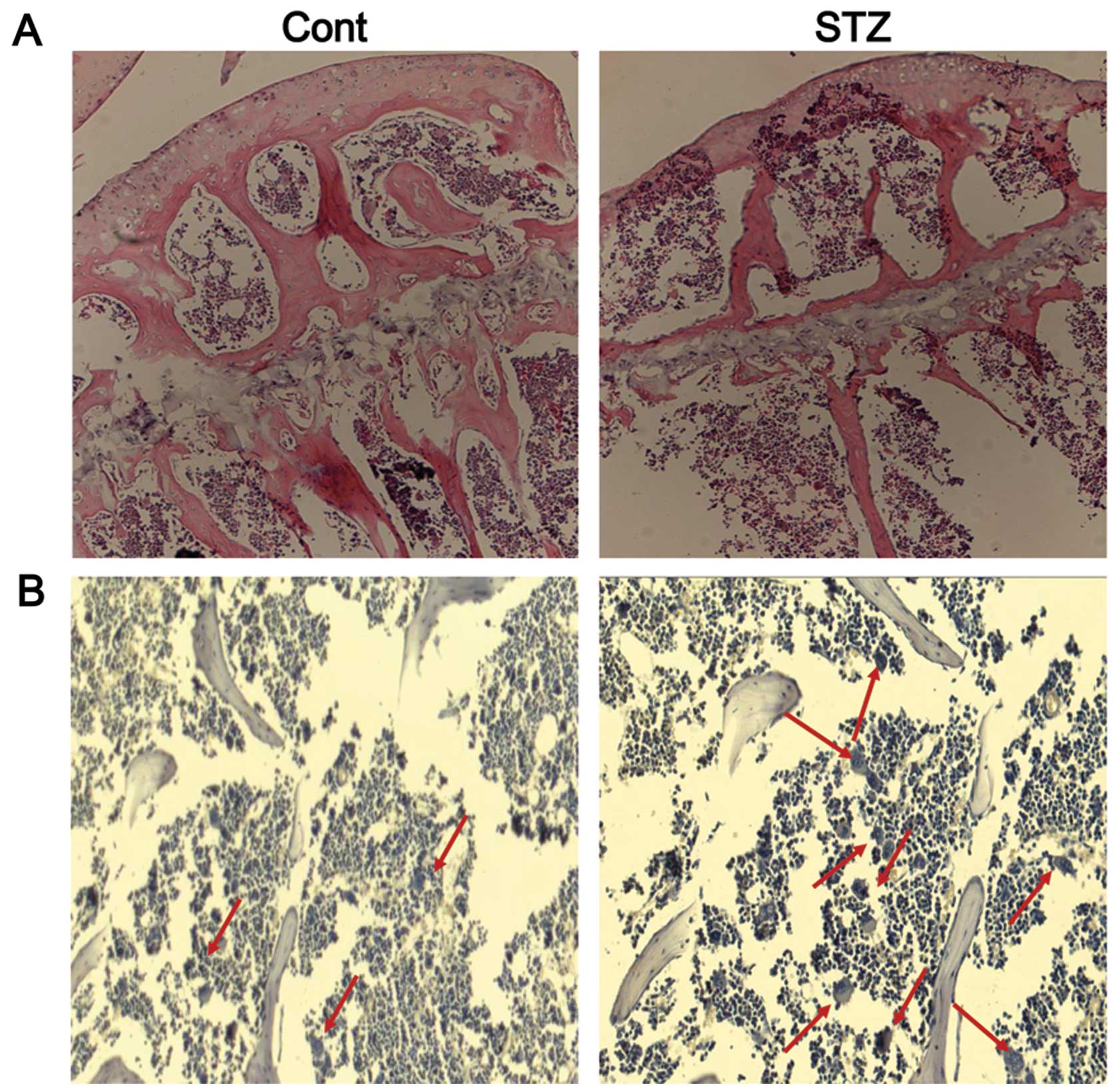
Histological analysis of trabecular bone in the
proximal tibia of mice was performed by H&E staining. The
results revealed the increased disconnections and separation among
the growth plate and trabecular bone network, as well as the
reduction of trabecular bone mass of the primary and secondary
spongiosa throughout the proximal tibial epiphysis and proximal
tibial metaphysis in the STZ group (Fig. 7A). TRAP staining revealed very few
mature osteoclasts in the trabecular bone area below the growth
plate in the control group, whereas the number of osteoclasts was
significantly increased in this area in the STZ group (Fig. 7B).
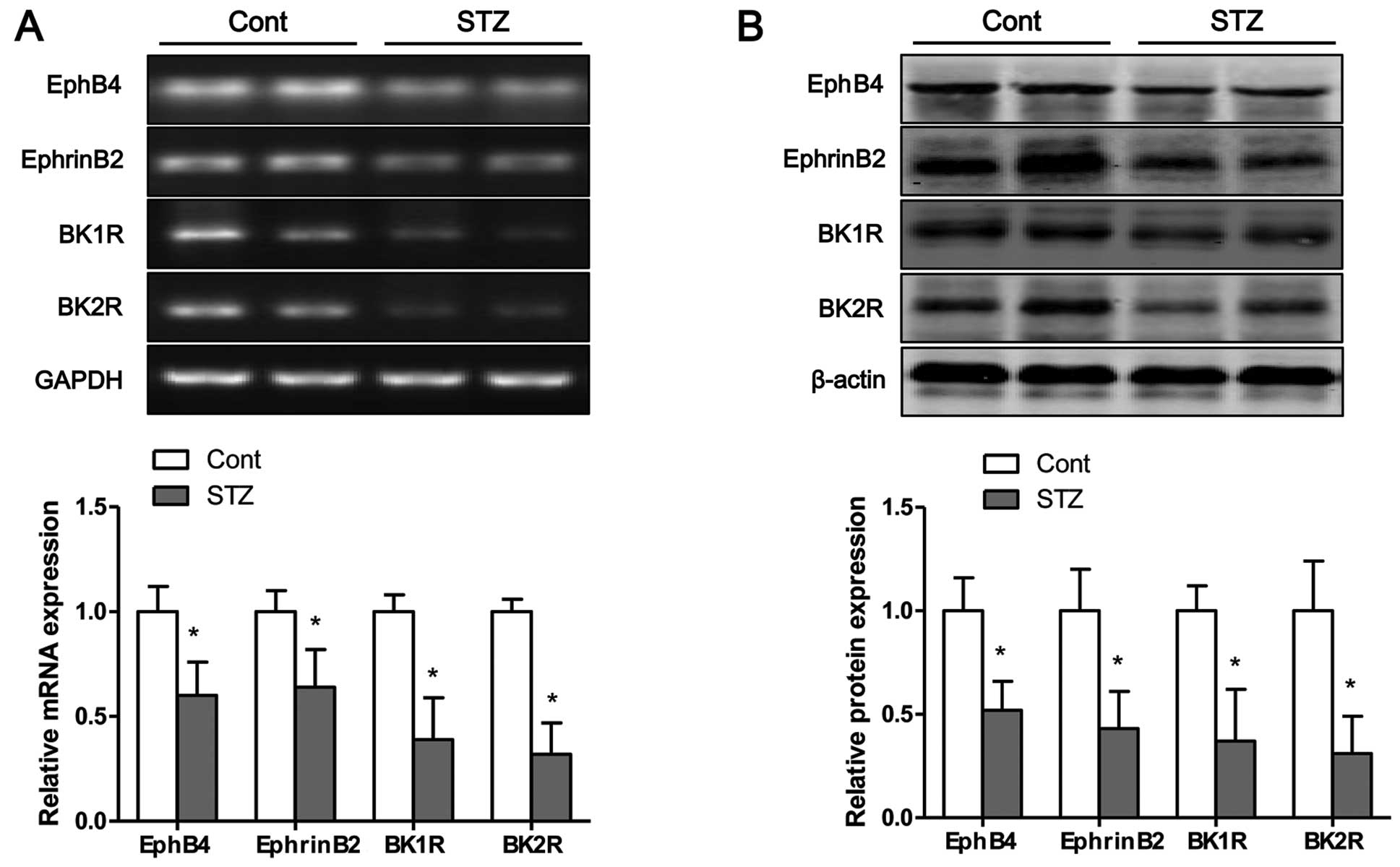
Hyperglycemia inhibits the mRNA and
protein expression of bradykinin receptors and EphB4/EphrinB2 in
tibias of mice
The mRNA and protein expression levels of bradykinin
receptors and EphB4/EphrinB2 in the tibias of mice were measured by
RT-PCR and western blot analysis, respectively. Eight weeks after
the STZ injection, the mRNA (Fig.
8A) and protein (Fig. 8B)
expression levels of BK1R and BK2R in the tibias of mice were
significantly decreased compared to those of the control group.
Moreover, the mRNA and protein expression levels of EphB4 and
EphrinB2 were significantly lower in the STZ group than those in
the control group (Fig. 8).
Discussion
In this study, we investigated the
physiopathological roles of skeletal EphB4/EphrinB2 and bradykinin
receptors in mice with diabetes-related osteoporosis.
EphB4/EphrinB2 signaling links two major molecular mechanisms of
cell differentiation, EphB4 in osteoblasts enhancing osteogenic
differentiation and EphrinB2 in osteoclast precursors suppressing
osteoclast differentiation, thereby maintaining bone homeostasis
(26). In myeloma cells,
EphB4/EphrinB2 is expressed at very low levels to be detectable,
and its activation by EphB4-Fc inhibits myeloma growth and bone
disease (29). In the present
study, we made several important observations. First, the high
glucose-induced increase in osteoblast apoptosis was confirmed by
flow cytometric analysis and the analysis of apoptosis-related
protein expression. Next, the EphB4/EphrinB2 axis and brady-kinin
receptors were found to be dysregulated in osteoblasts following
expesure to high glucose, and BK2R loss-of-function downregulated
the levels of EphB4/EphrinB2 in vitro. Further experiments
demonstrated that high glucose inhibited the interaction between
BK2R and EphB4/EphrinB2, which was identified by
co-immunoprecipitation. Furthermore, the dysreg-ulated expression
of bradykinin receptors and EphB4/EphrinB2 was confirmed in mice
with diabetes-related osteoporosis, and the binding capacity
between EphB4/EphrinB2 and BK2R was significantly weaker in the
mice with diabetes-related osteoporosis than in the non-diabetic
control mice.
EphB4/EphrinB2 is a research topic of great
scientific value; EphB4/EphrinB2 signaling has been found related
with the occurrence of many diseases. The reduced activity of EphB4
has been shown to accelerate the progression of colorectal cancer,
and the activation of EphB4 inhibits breast cancer cell growth and
migration (30). The
communication between osteo-clasts and osteoblasts is
bidirectional, and forward signaling by EphB4 and possibly other
co-expressed Eph receptors promotes the differentiation of
osteoblasts, which deposit new bone at sites of resorption by
osteoclasts (26). Several Eph
receptors present in osteoblasts are regulated by EphrinB reverse
signaling, which suppresses osteoblast differentiation through a
negative feedback loop that inhibits the expresion of bone
sialoprotein (16,31). In this study, we found that
exposure to high glucose inhibited osteoblast mineralization and
induced bone deterioration, including trabecular bone network
degradation and the reduction of the trabecular bone mass of the
primary and secondary spongiosa through the suppression of the the
expresion of bradykinin recptors and EphB4/EphrinB2 in vitro
and in vivo. Previous studies have demonstrated that
bradykinin recptors or the EphB4/EphrinB2 signaling pathway
maintain bone homeostasis (13,26). In the present study, our data
suggested that the interaction between EphB4/EphrinB2 and BK2R
plays a vital role in osteogenic differentiation. Moreover, BK2R
loss-of-function significantly decreased the mRNA and protein
expression levels of EphB4 and EphrinB2.
As previously demonstrated, in Akita diabetic and
non-diabetic mice, the lack of both bradykinin receptors results in
a severe reduction in bone mineral density, demonstrating the
importance of the bradykinin receptors in bone mineralization
(14). In this study, the BK1R
and BK2R levels were decreased following exposure to high glucose
in vitro or downregulated by hyperglycemia in vivo.
However, although osteoblasts express both BK1R and BK2R, their
stimulation with bradykinin increases the expression of RANKL,
which is known to be involved in osteoclastogenesis and to enhance
bone resorption (32). The
pro-inflammatory cytokines, interleukin (IL)-1β and tumor necrosis
factor (TNF)-α can enhance bradykinin receptor expression in
osteoblasts, which may help to explain the enhanced bone resorption
associated with inflammatory disorders (33). As a key active peptide in the
kallikrein-kinin system (KKS), bradykinin can elicit the BK1R and
BK2R-mediated activation of ERK1/2 and Akt pathways, which finally
leads to the activation of RANKL and decreases the differentiation
of osteoblasts with a concomitant increase in osteoclast formation
(34). Based on the
above-mentioned data, we concluded that bradykinin receptor
loss-of-function or gain-of-function can influence bone metabolism
in vitro and in vivo. Intriguingly, both BK1R and
BK2R loss-of-function or BK2R loss-of-function can lead to
insulin-resistant or inhibit insulin levels in mice, and B1R and
B2R mediate the development of complications in diabetic mice, and
the activation of KKS may be beneficial in reducing the severity of
complications in diabetic mice (14,35).
In conclusion, in the present study, we demonstrated
that high glucose or hyperglycemia inhibited the expression of
bradykinin receptors and EphB4/EphrinB2 in vitro or in
vivo, respectively. Subsequently, the interaction between
bradykinin receptors and EphB4/EphrinB2 was confirmed in
osteoblasts, and bradykinin receptor loss-of-function significantly
downregulated the levels of EphB4/EphrinB2. These results
demonstrated that bradykinin receptors and EphB4/EphrinB2 mediate
the development of complications in diabetic mice and suggest that
the inactivation of bradykinin receptors and EphB4/EphrinB2 may
enhance the severity of complications in mice with diabetes-related
osteoporosis.
References
|
1
|
Cummings SR and Melton LJ: Epidemiology
and outcomes of osteoporotic fractures. Lancet. 359:1761–1767.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hamann C, Kirschner S, Günther KP and
Hofbauer LC: Bone, sweet bone - osteoporotic fractures in diabetes
mellitus. Nat Rev Endocrinol. 8:297–305. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Armas LA, Akhter MP, Drincic A and Recker
RR: Trabecular bone histomorphometry in humans with type 1 diabetes
mellitus. Bone. 50:91–96. 2012. View Article : Google Scholar
|
|
4
|
Loureiro MB, Ururahy MA, Freire-Neto FP,
Oliveira GH, Duarte VM, Luchessi AD, Brandão-Neto J, Hirata RD,
Hirata MH, Maciel-Neto JJ, et al: Low bone mineral density is
associated to poor glycemic control and increased OPG expression in
children and adolescents with type 1 diabetes. Diabetes Res Clin
Pract. 103:452–457. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Janghorbani M, Feskanich D, Willett WC and
Hu F: Prospective study of diabetes and risk of hip fracture: The
Nurses' Health Study. Diabetes Care. 29:1573–1578. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Diao TY, Pan H, Gu SS, Chen X, Zhang FY,
Wong MS and Zhang Y: Effects of angiotensin-converting enzyme
inhibitor, captopril, on bone of mice with streptozotocin-induced
type 1 diabetes. J Bone Miner Metab. 32:261–270. 2014. View Article : Google Scholar
|
|
7
|
Rao Sirasanagandla S, Ranganath Pai
Karkala S, Potu BK and Bhat KM: Beneficial effect of Cissus
quadrangularis Linn. on osteopenia associated with
streptozotocin-induced type 1 diabetes mellitus in male Wistar
rats. Adv Pharmacol Sci. 2014:4830512014.PubMed/NCBI
|
|
8
|
Fu C, Zhang X, Ye F and Yang J: High
insulin levels in KK-Ay diabetic mice cause increased cortical bone
mass and impaired trabecular micro-structure. Int J Mol Sci.
16:8213–8226. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guan CC, Yan M, Jiang XQ, Zhang P, Zhang
XL, Li J, Ye DX and Zhang FQ: Sonic hedgehog alleviates the
inhibitory effects of high glucose on the osteoblastic
differentiation of bone marrow stromal cells. Bone. 45:1146–1152.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wittrant Y, Gorin Y, Woodruff K, Horn D,
Abboud HE, Mohan S and Abboud-Werner SL: High d(+)glucose
concentration inhibits RANKL-induced osteoclastogenesis. Bone.
42:1122–1130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Irie N, Takada Y, Watanabe Y, Matsuzaki Y,
Naruse C, Asano M, Iwakura Y, Suda T and Matsuo K: Bidirectional
signaling through ephrinA2-EphA2 enhances osteoclastogenesis and
suppresses osteoblastogenesis. J Biol Chem. 284:14637–14644. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Matsuo K and Otaki N: Bone cell
interactions through Eph/ephrin: Bone modeling, remodeling and
associated diseases. Cell Adhes Migr. 6:148–156. 2012. View Article : Google Scholar
|
|
13
|
Kwan Tat S, Pelletier JP, Amiable N,
Boileau C, Lavigne M and Martel-Pelletier J: Treatment with ephrin
B2 positively impacts the abnormal metabolism of human
osteoarthritic chondrocytes. Arthritis Res Ther. 11:R1192009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cheng S, Zhao SL, Nelson B, Kesavan C, Qin
X, Wergedal J, Mohan S and Xing W: Targeted disruption of ephrin B1
in cells of myeloid lineage increases osteoclast differentiation
and bone resorption in mice. PLoS One. 7:e328872012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arthur A, Zannettino A, Panagopoulos R,
Koblar SA, Sims NA, Stylianou C, Matsuo K and Gronthos S:
EphB/ephrin-B interactions mediate human MSC attachment, migration
and osteochondral differentiation. Bone. 48:533–542. 2011.
View Article : Google Scholar
|
|
16
|
Watanabe T, Sato Y, Saito D, Tadokoro R
and Takahashi Y: EphrinB2 coordinates the formation of a
morphological boundary and cell epithelialization during somite
segmentation. Proc Natl Acad Sci USA. 106:7467–7472. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xing W, Kim J, Wergedal J, Chen ST and
Mohan S: Ephrin B1 regulates bone marrow stromal cell
differentiation and bone formation by influencing TAZ
transactivation via complex formation with NHERF1. Mol Cell Biol.
30:711–721. 2010. View Article : Google Scholar :
|
|
18
|
Davy A, Aubin J and Soriano P: Ephrin-B1
forward and reverse signaling are required during mouse
development. Genes Dev. 18:572–583. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Davy A, Bush JO and Soriano P: Inhibition
of gap junction communication at ectopic Eph/ephrin boundaries
underlies craniofrontonasal syndrome. PLoS Biol. 4:e3152006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stiffel V, Amoui M, Sheng MH, Mohan S and
Lau KH: EphA4 receptor is a novel negative regulator of osteoclast
activity. J Bone Miner Res. 29:804–819. 2014. View Article : Google Scholar
|
|
21
|
Yamada T, Yuasa M, Masaoka T, Taniyama T,
Maehara H, Torigoe I, Yoshii T, Shinomiya K, Okawa A and Sotome S:
After repeated division, bone marrow stromal cells express
inhibitory factors with osteogenic capabilities, and EphA5 is a
primary candidate. Bone. 57:343–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao C, Irie N, Takada Y, Shimoda K,
Miyamoto T, Nishiwaki T, Suda T and Matsuo K: Bidirectional
ephrinB2-EphB4 signaling controls bone homeostasis. Cell Metab.
4:111–121. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Campbell DJ: The renin-angiotensin and the
kallikrein-kinin systems. Int J Biochem Cell Biol. 35:784–791.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Campbell DJ: The kallikrein-kinin system
in humans. Clin Exp Pharmacol Physiol. 28:1060–1065. 2001.
View Article : Google Scholar
|
|
25
|
Gonçalves-Zillo TO, Pugliese LS, Sales VM,
Mori MA, Squaiella-Baptistão CC, Longo-Maugéri IM, Lopes JD, de
Oliveira SM, Monteiro AC and Pesquero JB: Increased bone loss and
amount of osteoclasts in kinin B1 receptor knockout mice. J Clin
Periodontol. 40:653–660. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kakoki M, Sullivan KA, Backus C, Hayes JM,
Oh SS, Hua K, Gasim AM, Tomita H, Grant R, Nossov SB, et al: Lack
of both bradykinin B1 and B2 receptors enhances nephropathy,
neuropathy, and bone mineral loss in Akita diabetic mice. Proc Natl
Acad Sci USA. 107:10190–10195. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsukamoto T, Li X, Morita H, Minowa T,
Aizawa T, Hanagata N and Demura M: Role of S-palmitoylation on
IFITM5 for the interaction with FKBP11 in osteoblast cells. PLoS
One. 8:e758312013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bi Y, Nielsen KL, Kilts TM, Yoon A, A
Karsdal M, Wimer HF, Greenfield EM, Heegaard AM and Young MF:
Biglycan defi-ciency increases osteoclast differentiation and
activity due to defective osteoblasts. Bone. 38:778–786. 2006.
View Article : Google Scholar
|
|
29
|
Pennisi A, Ling W, Li X, Khan S,
Shaughnessy JD Jr, Barlogie B and Yaccoby S: The ephrinB2/EphB4
axis is dysregulated in osteoprogenitors from myeloma patients and
its activation affects myeloma bone disease and tumor growth.
Blood. 114:1803–1812. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pasquale EB: Eph-ephrin bidirectional
signaling in physiology and disease. Cell. 133:38–52. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shimizu E, Tamasi J and Partridge NC:
Alendronate affects osteoblast functions by crosstalk through
EphrinB1-EphB. J Dent Res. 91:268–274. 2012. View Article : Google Scholar :
|
|
32
|
Brechter AB and Lerner UH: Bradykinin
potentiates cytokine-induced prostaglandin biosynthesis in
osteoblasts by enhanced expression of cyclooxygenase 2, resulting
in increased RANKL expression. Arthritis Rheum. 56:910–923. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brechter AB, Persson E, Lundgren I and
Lerner UH: Kinin B1 and B2 receptor expression in osteoblasts and
fibroblasts is enhanced by interleukin-1 and tumour necrosis
factor-alpha. Effects dependent on activation of NF-kappaB and MAP
kinases. Bone. 43:72–83. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Srivastava S, Sharma K, Kumar N and Roy P:
Bradykinin regulates osteoblast differentiation by Akt/ERK/NFκB
signaling axis. J Cell Physiol. 229:2088–2105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Duka I, Shenouda S, Johns C,
Kintsurashvili E, Gavras I and Gavras H: Role of the B(2) receptor
of bradykinin in insulin sensitivity. Hypertension. 38:1355–1360.
2001. View Article : Google Scholar : PubMed/NCBI
|