Introduction
Mono-adenosine diphosphate (ADP)-ribosyltransferases
(ARTs) have been noted to catalyze ADP-ribosylation, which
transfers an ADP-ribose from nicotinamide adenine dinucleotide
(NAD+) to various amino acid residues, thus altering
protein function and playing an important role in physiological and
pathological processes such as signal transduction, cellular
differentiation and proliferation, protein secretion and transport,
cellular necrosis and apoptosis (1). Seven mono-ADP-ribosyltransferases
(ART1-ART7) have previously been found in mammalians, and five
members (ART1, ART2, ART5, ART6 and ART7) are specific to arginine
(2,3). ART1 was first discovered in
bacteria, viruses and eukaryotic cells (4), and it was subsequently found in
human skeletal muscles, cardiac muscle cells, white blood cells and
airway epithelial cells (5–7).
The gene which codes for ART1 is located on human chromosome 11P15,
and its target proteins include integrin α7, αL (CD11a), β2 (CD18),
human neutrophil peptide-1 (HNP-1), basic fibroblast growth factor
(bFGF) and platelet-derived growth factor (PDGF)-BB (8–12).
In non-neoplastic diseases, ARTs are related to the
occurrence of acute inflammation of the intestinal mucosa (13). When mono-ADP-ribosylation is
inhibited, the release of inflammatory mediators by monocytes and
macrophages is inhibited, suggesting that ARTs promote inflammatory
development (14). Furthermore,
ART1 specifically alters HNP-1 activity, which reduces
antibacterial and antitoxic ability of HNP-1 (15). When T lymphocytes are transfected
with ART1 in the presence of NAD+, the function of the T
cell receptor is altered by the lymphocyte function-associated
factor, thus affecting lymphocyte function and suggesting that ART1
acts in an extensive capacity (9,16).
Furthermore, gastric cancer researchers have found that
Helicobacter pylori modulates ART activity, leading to
modifications in membrane conformation and possibly promoting the
development of gastric cancer (17). In the lung cancer cell line A549,
ART1 is highly expressed; ART1 and ART4 are upregulated upon human
Toll-like receptor (hTLR) stimulation, thus suggesting that ARTs
are important in the development of lung cancer (18).
Previously we demonstrated that ART1 was highly
expressed in human colorectal cancer (CRC), and that its expression
was positively correlated with the expression of angiogenic factors
such as integrin αVβ3, vascular endothelial growth factor (VEGF)
and bFGF (19). It has previously
been suggested that ART1 has the ability to promote angiogenesis in
CRC. Several previous studies have reported that the
phosphoinositide 3-kinase (PI3K)/Akt pathway not only promotes
tumor cell proliferation and inhibits apoptosis, but is also
closely related to tumor angiogenesis. This mechanism may be
involved in the activation of the PI3K/Akt pathway, which
subsequently upregulates the expression of hypoxia-inducible factor
1-α (HIF-1α) and promotes transcription and secretion of VEGF, and
eventually leads to promote tumor angiogenesis through VEGF binding
to vascular endothelial cell surface receptors (20,21). It is unclear whether ART1 is
involved in VEGF, bFGF and HIF-1α expression, which is regulated by
the PI3K/Akt pathway, in CRC cells thus far.
In the present study, LoVo cells transfected with
ART1-cDNA or with ART1-shRNA were co-cultured with human umbilical
vein endothelial cells (HUVECs), and subsequently HUVEC
proliferation, migration and vascular-forming abilities were all
examined. Additionally, the effect of ART1 on tumor microvessel
density (MVD) was examined in vivo. The expression levels of
total (t-)Akt, phosphorylated (p-)Akt, HIF-1α, VEGF and bFGF were
further detected in order to characterize the molecular mechanisms
of ART1 during tumor angiogenesis.
Materials and methods
Cell and lentiviral transfection
The LoVo cell line, a human colon cancer cell line,
and HUVECs were kindly provided by Professor Wei-Xue Tang,
Chongqing Medical University (Chongqing, China). The mouse colon
cancer cell line CT26 was kindly provided by Professor Yu-Quan Wei,
Sichuan University (Chengdu, China). ART1 was cloned into a
pGCSIL-GFP lentiviral vector in order in order to generate both
pGCSIL-GFP-vshART1 and pGCSIL-GFP-ART1-cDNA. The ART1-shRNA
interference sequence was GCCAACAAAGTATACGCGGAT. The primers of
ART1-cDNA used were as follows: primer 1, GAGGAT
CCCCGGGTACCGGTCGCCACCATGAAGATTCCTGCTA TGATG; primer 2,
TCACCATGGTGGCGACCGGACA TCGGGTAAGTTGCTG. The pGCL-GFP-vshART1
lentiviral vector, ART1-cDNA lentiviral vector and empty lentiviral
vector were purchased from GeneChem, Inc. (Shanghai, China). The
human CRC cell line LoVo was cultured and transfected according to
the manufacturer's instructions (GeneChem, Inc.). LoVo cells were
seeded in 24-well plates at a density of 3×104
cells/well, transfected with 10 µl lentiviral vector and
observed under a fluorescence microscope 96 h post-transfection
until the infection rate reached 80%. Transfection efficiency was
detected under a fluorescence microscope (Leica DMI4000B; Leica
Microsystems GmbH, Wetzlar, Germany) after 96 h. The efficiency of
the ART1-cDNA and ART1-shRNA lentivirus transfected into LoVo cells
was detected by reverse transcription (RT)-PCR and western blot
analysis. The murine CRC cell line CT26 was cultured and
transfected as previously described by Tang et al (22). The group which were transfected
with ART1-shRNA lentiviral and the group transfected with ART1-cDNA
lentiviral vector were set as experimental groups. The empty
lentiviral vector transfected group (LV-control group) and
untransfected group were set as control groups.
RT-PCR
Total RNA was separately extracted from
untransfected LoVo cells, LV-control LoVo cells, ART1-cDNA- and
ART1-shRNA-transfected LoVo cells with TRIzol reagent (Takara,
Dalian, China) and reverse transcribed into DNA. ART1 gene
expression was detected using oligonucleotide primers which were
designed and produced by Sangon Biotech Co. (Shanghai, China):
ART1, 5′-ACCTTCTTCGGTATCTGGACCT-3′ (F1) and 5′-TAAGTTGCTGGAGACCTG
GATT-3′ (R1); and β-actin acted as the internal control gene,
5′-ATATCGCTGCGCTGGTC GTC-3′ (F1) and 5′-AGGATGG CGTGAGGGAGAGC-3′
(R1). Using a one-step RT-PCR kit (Takara), in total, 30 PCR cycles
were used for the amplification of RT products (94°C for 30 sec,
60°C for 30 sec and 72°C for 25 sec). Finally, the amplifiation
products were electrophoresed on 2% agarose gels (Genview,
Tallahasse, FL, USA). The above experiment was performed in
triplicate
Analysis of HUVEC proliferation
In the present study, HUVEC proliferation was
detected as previously described (23). The single-cell suspensions of
HUVECs were cultured in a 96-well culture plate at a final
concentration of 2×103 cells/well in Dulbecco's modified
Eagle's medium with 20% fetal bovine serum (both purchased from
Gibco-BRL, Gaithersburg, MD, USA) for 24 h. HUVECs were
subsequently co-cultured with supernatant fluid of LoVo cells in
ratios of 0, 20, 40, 60, 80 and 100% 48 h later. This was followed
by the addition of 10 µl of Cell Counting kit (CCK-8; KeyGen
Biotechnology, Nanjing, China) reagent to each well, incubation for
4 h and detection at an OD of 450 nm by universal microplate reader
(Bio-Tek Instruments Inc., Winooski, VT, USA). The assays were
repeated in triplicate.
HUVECs migration analysis
HUVEC migration was detected using 8.0-mm pore size
Transwell inserts (Costar, Milpitas, CA, USA) as previously
described (24). The Transwell
semipermeable membrane was coated with diluted Matrigel (Sigma,
Ronkonkoma, NY, USA) incubated at 37°C for 30 min to solidify. LoVo
single-cell suspension was seeded into the lower chamber at a
concentration of 1×105 cells/well and incubated
overnight in RPMI-1640 with 10% FBS (both from Gibco-BRL,
Gaithersburg, MD, USA). HUVEC single-cell suspension was seeded in
the upper chamber at a concentration of 1×105 cells/well
and incubated under conventional culture conditions. At 6 and 12 h
later, the cells on the upper surface of the membrane were
carefully scraped off and the migrant cells which adhered to the
lower surface were fixed with 4% methanol for 15 min at room
temperature, and stained with 0.1% crystal violet for 3 min. The
number of cells on the lower surface of the membrane was counted in
five different fields using an inverted microscope at a
magnification of ×200. The experiment was repeated in
triplicate.
Analysis of HUVEC lumen-forming
ability
A 3D collagen cell culture system (Millipore, Inc.,
Billerica, MA, USA) was utilized; we proportionally combined
collagen solution, 5X medium and neutralization solution, with 150
µl of the mixture added to each well of a 24-well plate. The
plate was then incubated at 37°C for 60 min in order to solidify.
The HUVEC single cell suspension was then seeded into each well at
a concentration of 5×105 cells/well and mixed with
different ratios (0, 10, 20 and 50%) of supernatant from
transfected LoVo cells, cultured for 72 h and observed under the
microscope, with the total length randomly selected for 5 lumen,
and the total length was measured (25). The experiment was repeated in
triplicate.
Transplanted tumor mouse model
Sixteen BALB/c female mice (Chongqing Medical
University Laboratory Animal Center, Chongqing, China), 6–8 weeks
old and weighing 18–22 g, were randomly divided into four groups:
the group transfected with ART1-shRNA lentiviral vector, the group
transfected with ART1-cDNA lentiviral vector, a group transfected
with an empty lentiviral and the unransfected group. Mice were
inoculated as previously described (26). Following anesthesia an
intraperitoneal injection of 2% chloral hydrate (Beijing DingGuo
Biological Technology Co., Ltd., Beijing, China), the mice were
fixed in a supine position, and an incision was made into the
abdominal cavity from the left upper quadrant axillary midline to
expose the spleen. The mice were administered the CT26 cells
(5×106) in single suspension/mouse via a subcapsular
injection into the spleen. Fourteen days later, the mice were
sacrificed by cervical dislocation and the metastatic spleen
tissues were removed and fixed with 4% paraformaldehyde, and
paraffin-embedded tissues were cut into slices for H&E staining
and immunohistochemistry.
All animal experiments were performed after
obtaining approval from the Ethics Committee of Chongqing Medical
University. Appropriate care was taken to minimize animal
suffering. The disposition of the animals at the end of the study,
euthanisa criteria and all experimental methods involving animals
were in accordance with the code of practice for the care and use
of animals for scientific purposes.
H&E staining and CD34
immunohistochemical assay
The tissue sections were dewaxed with xylene, and
the slides were then soaked in graded concentrations of alcohol
(100, 95, 85 and 70%) for 5 min, stained with hematoxylin for 3 min
and eosin for 1 min (both from Beijing DingGuo Biological
Technology Co., Ltd.) washing with water. Vascular endothelial
cells were marked with CD34 (Wuhan Boster Biological Technology,
Ltd., Wuhan, China) using an EnVision™ detection kit
(DakoCytomation, Genetech, Beijing, China) following the
manufacturer's protocols, as previously described (27). According to the method put forward
by Weidner et al (27), areas
with the most intense coloring were selected as 'hot spots', and
the microvessels were counted under the microscope and imaged, with
3 'hot spots' counted per slice and averaged to obtain the slide
MVD.
Western blot analysis
LoVo cells from each transfection group were
collected; total protein was extracted according to protein
extraction protocols (Beyotime P0013; Beyotime Biotechnology Co.,
Ltd., Shanghai, China). Protein concentrations were determined
using a BCA assay kit (Beyotime Biotechnology Co., Ltd.) and equal
amounts of total protein were separated via SDS-PAGE and
transferred onto a PVDF membrane. The membrane was blocked at room
temperature for 1 h, and incubated with the primary antibody
overnight at 4°C. The primary antibodies, ART1 (AP2311a; Abgent
Biotechnology, Inc. San Diego, CA, USA), HIF-1α (20960-1-AP;
Proteintech Group, Inc., Chicago, IL, USA), Akt, p-Akt
Thr308 (BS2987 and BS4009, respectively; Bioworld
Technology, Co., Ltd., St. Louis Park, MN, USA), VEGF, bFGF and
β-actin (PB0084, BA0259 and BA2305, respectively; Wuhan Boster
Biological Technology, Ltd.), were diluted to 1:500, 1:1,000,
1:800, 1:600, 1:1,000, 1:1,000 and 1:1,000, respectively. The
membrane was washed with TBST and incubated with the secondary
antibody (peroxidase-conjugated goat anti-rabbit IgG,
peroxidase-conjugated rabbit anti-goat IgG or peroxidase-conjugated
goat anti-mouse IgG; ZDR-5306, ZDR-5308 and ZDR-5307, respectively;
Beijing ZSGB Biotechnology Co., Ltd.) at a dilution of 1:1000 for 2
h. The image was captured and imaged using a ChemiDoc XRS system
(Bio-Rad, Hercules, CA, USA) using chemiluminescence (BeyoECL Plus;
Beyotime Biotechnology Co., Ltd.) and analyzed using Quantity one
software. The experiment was repeated in triplicate.
Statistical analysis
Statistical analysis was performed using SPSS 18.0
statistical software, and the data is expressed as the means ± SD;
single factor compared analysis was used for analysis between
groups, and the LSD method was used between two groups. A P-value
<0.05 was considered to indicate a statistically significant
difference.
Results
LoVo Cells transfected with
lentivirus
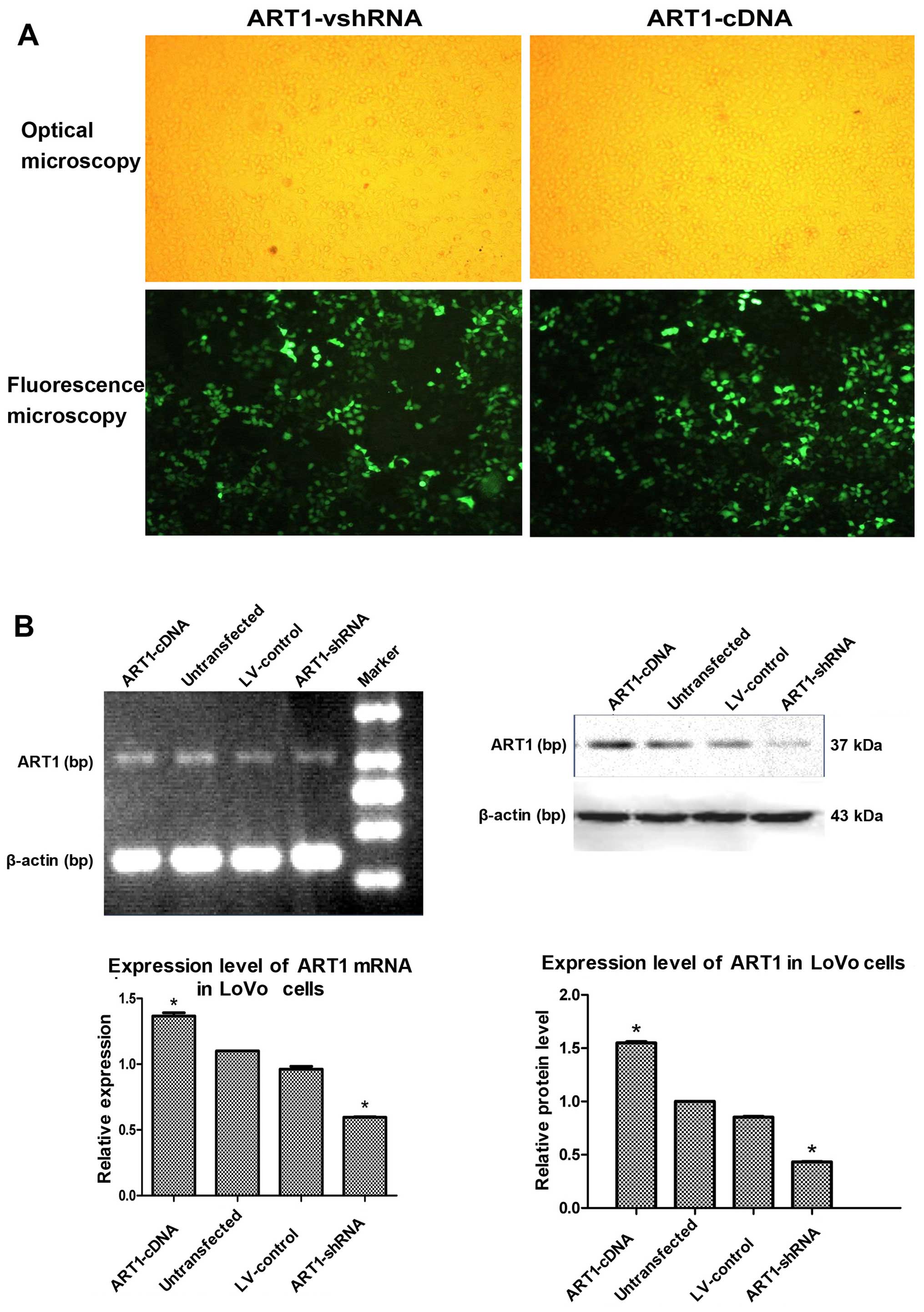
LoVo cells were transfected with ART1-shRNA or
ART1-cDNA lentiviral vector, and cells exhibited 80% fluorescence
on the third and fifth days, respectively (Fig. 1A). ART1 expression in LoVo cells
was detected by RT-PCR and western blot analysis. Compared with the
untransfected and LV-control-transfected groups, ART1 mRNA and
protein expression was significantly lower in the ART1-silenced
groups and significantly higher in the ART1-cDNA-transfected group
(P<0.05). However, there was no significant difference in ART1
mRNA and protein expression between un-transfected LoVo cells and
LV-control-transfected LoVo cells (P>0.05) (Fig. 1B).
Effects of ART1-shRNA-transfected LoVo
cells and ART1-cDNA-transfected LoVo cells on HUVEC
proliferation
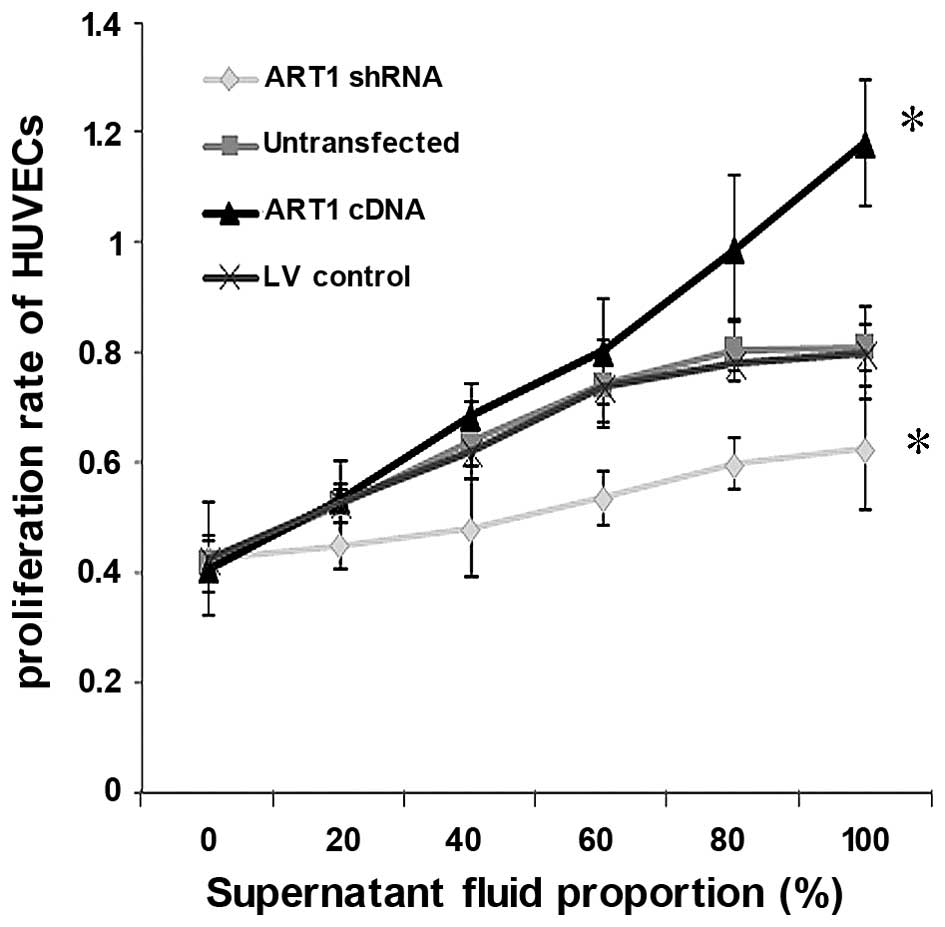
The proliferation capacity of HUVECs co-cultured
with the supernatant fluid of ART1-shRNA LoVo cells was
significantly decreased relative to the control group and
untransfected group (P<0.05). By contrast, the proliferation
capacity of HUVECs co-cultured with the supernatant fluid of
ART1-cDNA LoVo cells was significantly higher (P<0.05), and the
proliferation capacity increased with the increasing concentration
of the LoVo supernatant. Furthermore, no significant difference
between HUVECs co-cultured with the supernatant fluid of
untransfected LoVo cells and LV-control LoVo cells was noted
(P>0.05) (Fig. 2).
Effects of ART1-shRNA-transfected LoVo
cells and ART1-cDNA-transfected LoVo cells on HUVEC migration
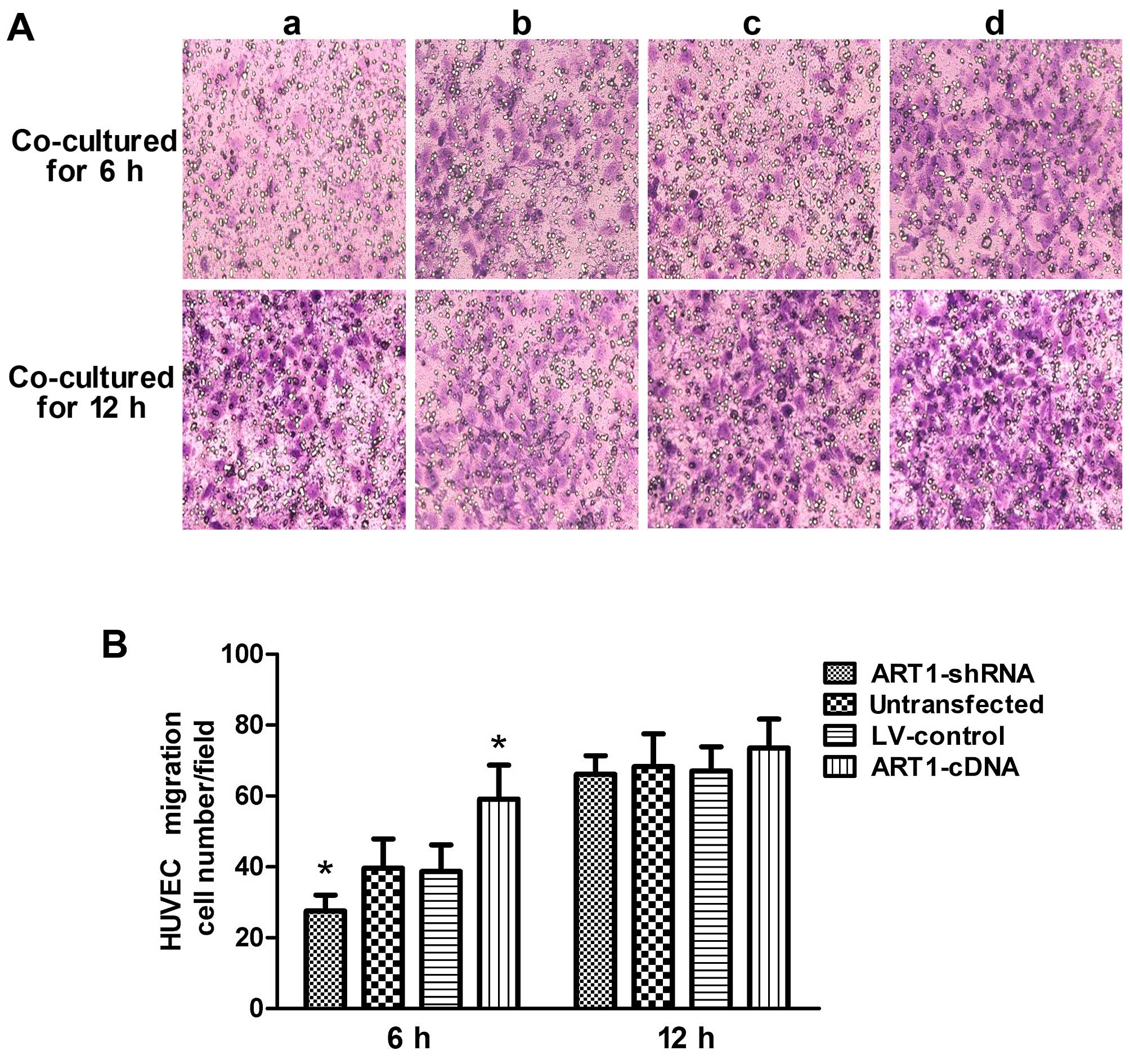
Compared with the control group, the migratory
ability of HUVECs co-cultured with the ART1-shRNA-transfected cells
was significantly decreased (P<0.05). By contrast, HUVEC
migration increased after co-culture with ART1-cDAN LoVo cells
(P<0.05). The difference between HUVECs co-cultured with
untransfected LoVo cells and with LV-control LoVo cells was not
significant (P>0.05). Moreover, after 12 h, the majority of the
HUVECs had passed through the Transwell membrane, and the
difference was indistinguishable from the HUVECs co-cultured with
the control LoVo cells (P>0.05) (Fig. 3).
Effects of ART1-shRNA-transfected LoVo
cells and ART1-cDNA-transfected LoVo cells on the blood vessel
forming ability of HUVECs
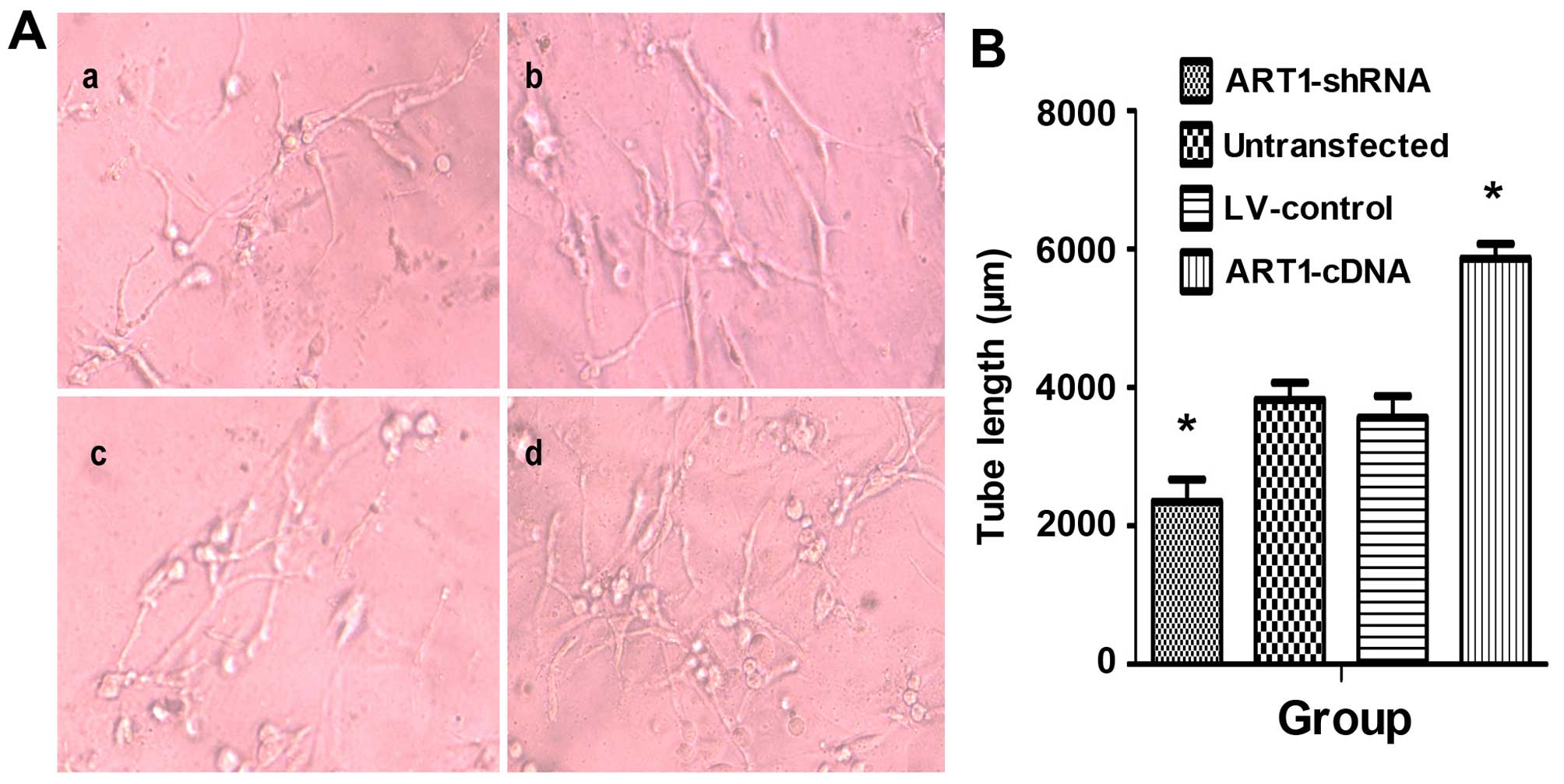
HUVEC angiogenic abilities were significantly
enhanced after incubation with the supernatant of
ART1-cDNA-transfected LoVo cells. Both the number and length of
vessels were significantly higher than those in the control group
and untransfected group (P<0.05). Furthermore, the blood vessel
lumens were mutually staggered. Moreover, HUVEC vascular forming
ability was significantly reduced when co-cultured with the
supernatant of ART1-silenced cells (P<0.05). There was no
significant difference between HUVECs incubated with the
supernatant of untransfected LoVo cells and LV-control LoVo cells
(P>0.05) (Fig. 4).
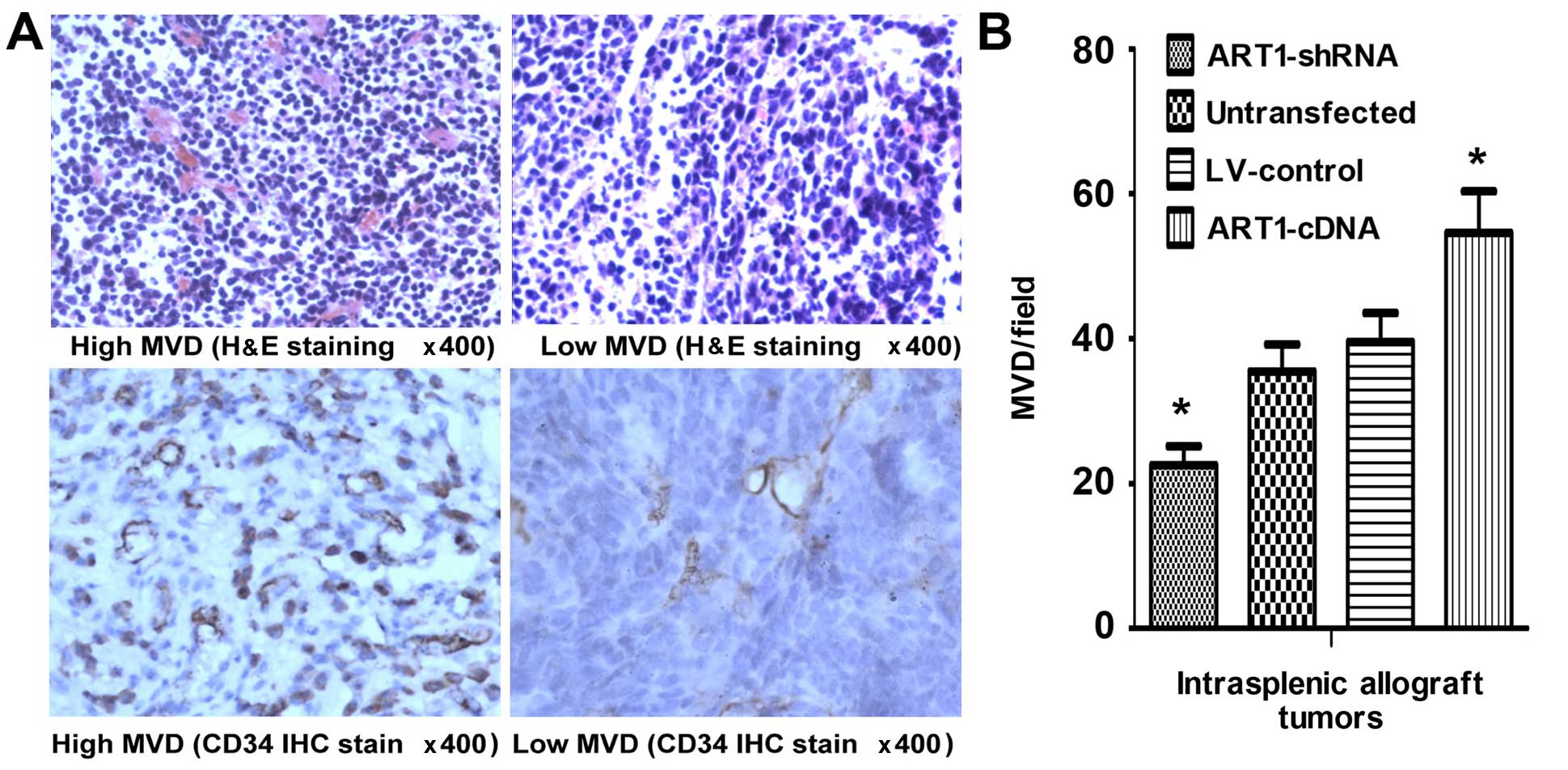
MVD in mice with transplanted tumor
tissues
We noted that, compared with the control group, the
MVD of tumors was significantly increased in mice which had
undergone intrasplenic ART1-cDNA-transfection, whereas it was
decreased in mice which had undergone intrasplenic transplantation
with ART1-shRNA-transfected CT26 cells (P<0.05). However, the
difference was not significant in the mice which were infected with
untransfected CT26 cells and LV-control-transfected CT26 cells
(P>0.05) (Fig. 5).
Effects of ART1 on the expression of
t-Akt, p-Akt, HIF-1α, VEGF and bFGF
In comparison with the control and also the
untransfected groups, we noted that p-Akt, HIF-1α, VEGF and bFGF
expression were increased in the ART1-cDNA-transfected group, while
expression was decreased in the ART1-shRNA-transfected group
(P<0.05). The expression of t-Akt did not differ significantly
between the experimental groups and the control groups (P>0.05).
Furthermore, p-Akt, HIF-1α, VEGF and bFGF expression did not differ
significantly between the untransfected LoVo cells and
LV-control-transfected LoVo cells (Fig. 6).
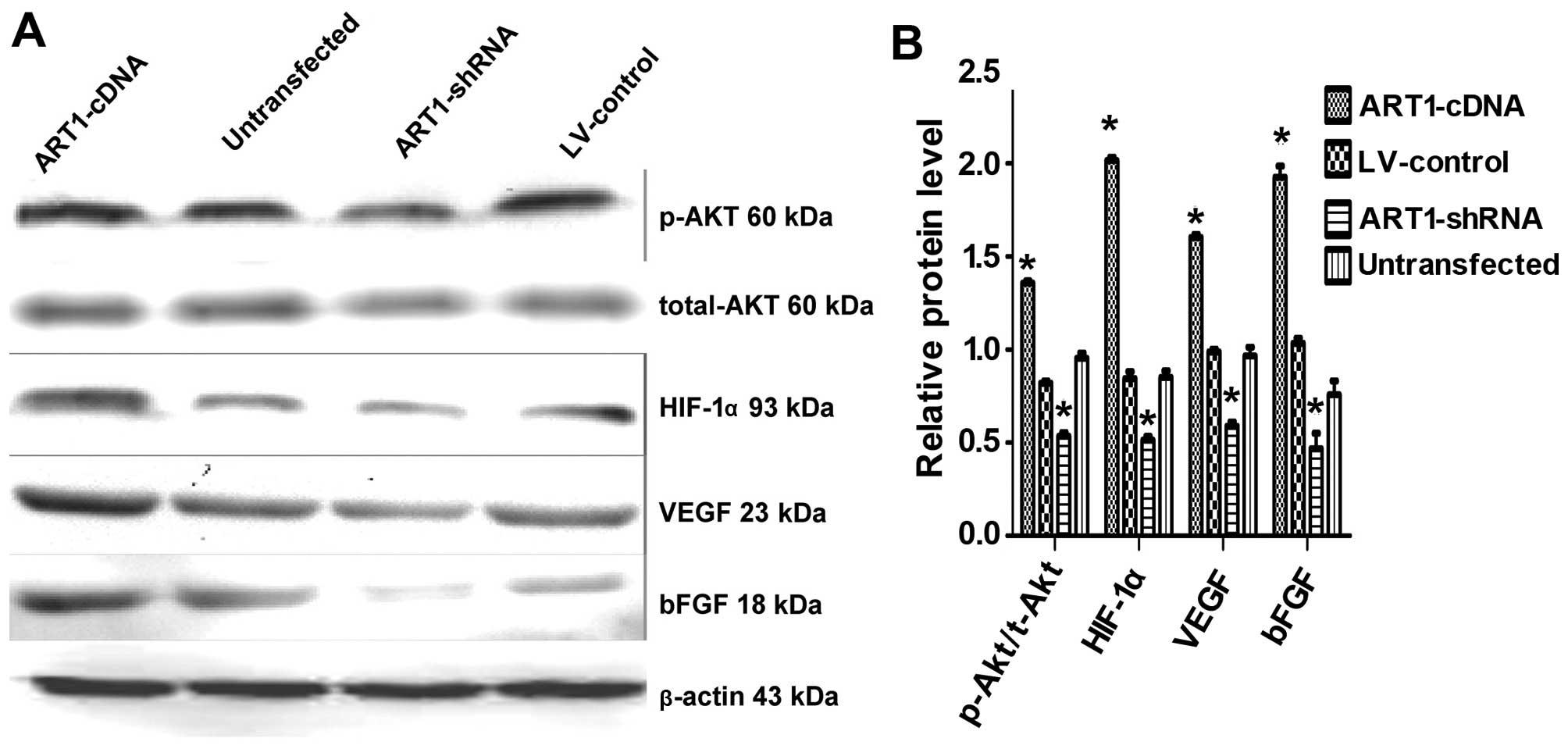 | Figure 6Arginine-specific adenosine
diphosphate (ADP)-ribosyltransferase 1 (ART1) affects total-Akt,
p-Akt, HIF-1α, VEGF and bFGF expression in LoVo cells. (A) The
expression of total-Akt, p-Akt, hypoxia-inducible factor 1-α
(HIF-1-α), vascular endothelial growth factor (VEGF) and basic
fibroblast growth factor (bFGF) in ART1-mediated LoVo cells. (B)
Comparison of Akt, p-Akt, HIF-1α, VEGF and bFGF expression in
ART1-cDNA-transfected LoVo cells with that of
ART1-shRNA-transfected LoVo cells, LV-control-transfected LoVo
cells and untransfected LoVo cells. *P<0.05 vs.
untransfected LoVo cells; n=3 experiments. |
Discussion
ART1, a representative ecto-ART family protein,
catalyzes ADP-ribosylation, which transfers an ADP-ribose from
NAD+ to an amino acid residue, and thus plays an
important role in the protein modification mechanism. Pioneering
research has found that certain growth factors are directly
regulated by ART1, and ART1 modulates the binding of PDGF-BB to its
receptor and regulates its signal transduction capabilities
(12). Our earlier studies have
demonstrated that ART1 is related to apoptosis, proliferation and
migration of mouse CT26 cells (28–30), suggesting that ART1 is involved in
tumor growth, invasion and metastasis. It is widely known that
angiogenesis plays an important role in malignant tumor occurrence,
development, invasion and metastasis, but as yet there have been no
studies regarding the role of ART1 in angiogenesis during CRC.
We found that ART1 was highly expressed in a human
CRC and was positively correlated with the expression of antigenic
factors (19). In order to
further confirm the association between ART1 and angiogenesis, in
the present study, ART1 was stably expressed at a high level in the
LoVo cell line and silenced using lentiviral vectors. Additionally,
LoVo cells were co-cultured with HUVECs to determine the effects of
high expression of ART1 or ART1 silencing on HUVEC proliferation,
migration and angiogenesis. Our data demonstrated that in LoVo
cells with high ART1 expression, HUVEC proliferation, migration and
angiogenesis was increased relative to the control. By contrast,
HUVEC proliferation, migration and angiogenesis were significantly
reduced following co-culture with ART1-silenced LoVo cells. These
data suggest that ART1 promotes the proliferation and migration of
vascular endothelial cells, thereby inducing tumor
angiogenesis.
VEGF is a crucial mitogen for vascular endothelial
cells in vitro and in vivo, and plays an important
role in inducing endothelial cell proliferation and angiogenesis.
VEGF has been shown to correlate with angiogenesis in a number of
human cancers, as evidenced by MVD (31,32). bFGF is the strongest growth factor
to induce cell generation, and increases the expression of VEGF in
endothelial cells while directly contributing to the proliferation
of vascular endothelial cells, exerting a synergistic effect with
VEGF (33). VEGF/bFGF-induced
angiogenesis is a crucial step in tumor progression and metastasis.
The data in our present study showed that compared with the control
lentivirus-transfected group and untransfected group, VEGF and bFGF
expression was upregulated in the group transfected with ART1-cDNA,
while expression was downregulated in silenced ART1 LoVo cells. MVD
was significantly higher in tumor tissues of the mice which had
been infected with CT26 cells transfected with ART1-cDNA, whereas
angiogenesis was inhibited when ART1 was silenced in the
intrasplenic implantation model. These results suggest that ART1
promotes the expression of VEGF and bFGF, but its mechanism is not
yet clear.
Our previous research has shown that the
ART1-specific inhibitor MIBG inhibits the expression of the
integrin α7β1, focal adhesion kinase (FAK) and PI3K, thereby
inhibiting HepG2 cell growth and invasion (34). Another study has shown that ARTs
directly regulate phosphodiesterase in the retina (35), suggesting that ARTs affects
protein functions by directly regulating ADP-ribosylation, thus
modulating signal transduction. It has previously been deonstrated
that multiple signaling networks are involved in the regulation of
tumor angiogenesis, and the PI3K/Akt pathway has been shown to be
of significant importance (36).
Akt is a major downstream effector molecule of PI3K
(37), and increased
AktThr308 phosphorylation implies the augmentation of
the PI3K pathway (38); it can
directly phosphorylate various transcription factors such as NF-κB
and mTOR and is involved in the regulation of various biological
activities (39). PI3K activation
induces Akt phosphorylation, thus promoting GSK-3 phosphorylation,
inhibiting HIF-1α degradation (40), activating the downstream gene mTOR
and promoting HIF-1α gene transcription (41). HIF-1α has been revealed to play a
central role in regulating the metabolic switch in cancer (42). Overexpression of HIF-1α is related
to increased mortality in patients with various types of tumor
(43,44) HIF-1α is activated by PI3K-Akt-mTOR
pathways and hyper-activity of RAS/MAPK (45). It is also activated by oncogenic
mutation of PTEN and VHL (46,47). Activated HIF-1α binds to the
hypoxia response element (HREs), promoting the transcription of a
variety of pro-angiogenic genes in the cell and cellular matrix and
upregulating VEGF, bFGF expression and that of other
angiogenesis-related factors in tumor tissues (48). HIF-1α regulates the expression of
VEGF in cancer cells, is activated by growth factors, and induces
formation of the proangiogenic microenvironment in gastric cancer
(45). The CD31-positive vessels
and tumor growth both decreased in a subcutaneous implantation
model when HIF-1α activation was blocked in a gastric cancer cell
line (49). These results suggest
that suppression of Thr PI3K/Akt pathway leads to decreased
expression of HIF-1α and inhibits tumor angiogenesis.
Based on these studies, in the present study we
detected the expression of t-Akt, p-Akt and HIF-1α, and the data
showed that compared with the control groups, p-Akt, and HIF-1α
expression was upregulated in LoVo cells transfected with
ART1-cDNA, whereas expression was downregulated in LoVo cells
transfected with ART1-shRNA. We speculate that ART1 regulates
HIF-1α expression through the PI3K/Akt pathway. Similar to the
effects of ART1 silencing, the PI3K/Akt pathway inhibitor LY294002
inhibited Akt phosphorylation but failed to bring about significant
changes in ART1 expression (28),
and this confirms that the Akt pathway is located downstream of
ART1. These data suggest that ART1 promotes Akt phosphorylation and
subsequent HIF-1α expression and activation, increases VEGF and
bFGF expression of cancer cells, and eventually leads to an
increase in tumor angiogenesis in tumor tissue.
In brief, these data suggest that the mechanism of
ART1 in tumor angiogenesis is quite complex and is involved in
multiple signaling pathways. The data from the present study
demonstrate that ART1 upregulates HIF-1α through the PI3K/Akt
signaling pathway to promote the expression of angiogenic factors
such as VEGF and bFGF and induce angiogenesis in cancer tissue.
Furthermore, its exact mechanisms require further in-depth
study.
Acknowledgments
This study was supported by the Ministry of
Education Specialized Research Fund for the Doctoral Program of
Higher Education (Grant no. 20105503110009), the Science and
Technology Project of Chongqing Municipal Education Commission
(Grant no. KJ110322) and the National Nature Science Foundation of
China (NSFC: 30870946). We would like to thank Tang Yi and Xiao
Ming for preparing the CT26 murine CRC cells, and we also thank
Kuang Jing, and Song Guang-Lin, postgraduate students of Chongqing
Medical and Science University, for their assistance in this
study.
References
|
1
|
Bergers G, Brekken R, McMahon G, Vu TH,
Itoh T, Tamaki K, Tanzawa K, Thorpe P, Itohara S, Werb Z and
Hanahan D: Matrix metalloproteinase-9 triggers the angiogenic
switch during carcinogenesis. Nat Cell Biol. 2:737–744. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Seman M, Adriouch S, Haag F and Koch-Nolte
F: Ecto-ADP-ribosyltransferases (ARTs): Emerging actors in cell
communication and signaling. Curr Med Chem. 11:857–872. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Braren R, Glowacki G, Nissen M, Haag F and
Koch-Nolte F: Molecular characterization and expression of the gene
for mouse NAD+: arginine
ecto-mono(ADP-ribosyl)transferase, Art1. Biochem J. 336:561–568.
1998. View Article : Google Scholar
|
|
4
|
Corda D and Di Girolamo M: Functional
aspects of protein mono-ADP-ribosylation. EMBO J. 22:1953–1958.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Okazaki IJ, Zolkiewska A, Nightingale MS
and Moss J: Immunological and structural conservation of mammalian
skeletal muscle glycosylphosphatidylinositol-linked
ADP-ribosyltransferases. Biochemistry. 33:12828–12836. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kefalas P, Allport JR, Donnelly LE,
Rendell NB, Murray S, Taylor GW, Lo G, Yadollahi-Farsani M and
MacDermot J: Arginine-specific mono(ADP-ribosyl)transferase
activity in human neutrophil polymorphs. A possible link with the
assembly of filamentous actin and chemotaxis. Adv Exp Med Biol.
419:241–244. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Balducci E, Horiba K, Usuki J, Park M,
Ferrans VJ and Moss J: Selective expression of RT6 superfamily in
human bronchial epithelial cells. Am J Respir Cell Mol Biol.
21:337–346. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zolkiewska A and Moss J: Integrin alpha 7
as substrate for a glycosylphosphatidylinositol-anchored
ADP-ribosyltransferase on the surface of skeletal muscle cells. J
Biol Chem. 268:25273–25276. 1993.PubMed/NCBI
|
|
9
|
Nemoto E, Yu Y and Dennert G: Cell surface
ADP-ribosyltransferase regulates lymphocyte function-associated
molecule-1 (LFA-1) function in T cells. J Immunol. 157:3341–3349.
1996.PubMed/NCBI
|
|
10
|
Wang J, Nemoto E and Dennert G: Regulation
of CTL by ecto-nictinamide adenine dinucleotide (NAD) involves
ADP-ribosylation of a p56lck-associated protein. J Immunol.
156:2819–2827. 1996.PubMed/NCBI
|
|
11
|
Laing S, Unger M, Koch-Nolte F and Haag F:
ADP-ribosylation of arginine. Amino Acids. 41:257–269. 2011.
View Article : Google Scholar :
|
|
12
|
Saxty BA, Yadollahi-Farsani M, Upton PD,
Johnstone SR and MacDermot J: Inactivation of platelet-derived
growth factor-BB following modification by ADP-ribosyltransferase.
Br J Pharmacol. 133:1219–1226. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kato J, Zhu J, Liu C and Moss J: Enhanced
sensitivity to cholera toxin in ADP-ribosylarginine
hydrolase-deficient mice. Mol Cell Biol. 27:5534–5543. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Del Vecchio M and Balducci E: Mono
ADP-ribosylation inhibitors prevent inflammatory cytokine release
in alveolar epithelial cells. Mol Cell Biochem. 310:77–83. 2008.
View Article : Google Scholar
|
|
15
|
Stevens LA, Levine RL, Gochuico BR and
Moss J: ADP-ribosylation of human defensin HNP-1 results in the
replacement of the modified arginine with the noncoded amino acid
ornithine. Proc Natl Acad Sci USA. 106:19796–19800. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu ZX, Yu Y and Dennert G: A cell surface
ADP-ribosyltransferase modulates T cell receptor association and
signaling. J Biol Chem. 274:17399–17401. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akai T, Nabeya Y, Yahiro K, Morinaga N,
Mitsuhashi K, Inoue M, Sakamoto A, Ochiai T and Noda M:
Helicobacter pylori induces mono-(adenosine
5′-diphosphate)-ribosylation in human gastric adenocarcinoma. Int J
Oncol. 29:965–972. 2006.PubMed/NCBI
|
|
18
|
Balducci E, Micossi LG, Soldaini E and
Rappuoli R: Expression and selective up-regulation of toxin-related
mono ADP-ribosyltransferases by pathogen-associated molecular
patterns in alveolar epithelial cells. FEBS Lett. 581:4199–4204.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang L, Wang YL, Sheng YT, Xiong W, Xu JX,
Tang Y and Li X: The correlation of ART1 expression with
angiogenesis in colorectal carcinoma and its relationship with VEGF
and integrin αVβ3 expressions. J Basic Clin Med. 32:1065–1069.
2012.
|
|
20
|
Fang J, Ding M, Yang L, Liu LZ and Jiang
BH: PI3K/PTEN/AKT signaling regulates prostate tumor angiogenesis.
Cell Signal. 19:2487–2497. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gray MJ, Zhang J, Ellis LM, Semenza GL,
Evans DB, Watowich SS and Gallick GE: HIF-1alpha, STAT3, CBP/p300
and Ref-1/APE are components of a transcriptional complex that
regulates Src-dependent hypoxia-induced expression of VEGF in
pancreatic and prostate carcinomas. Oncongene. 24:3110–3120. 2005.
View Article : Google Scholar
|
|
22
|
Tang Y, Wang Y-L, Yang L, Xu J-X, Xiong W,
Xiao M and Li M: Inhibition of arginine ADP-ribosyltransferase 1
reduces the expression of poly(ADP-ribose) polymerase-1 in colon
carcinoma. Int J Mol Med. 32:130–136. 2013.PubMed/NCBI
|
|
23
|
Yang LP, Cheng P, Peng XC, Shi HS, He WH,
Cui FY, Luo ST, Wei YQ and Yang L: Anti-tumor effect of
adenovirus-mediated gene transfer of pigment epithelium-derived
factor on mouse B16-F10 melanoma. J Exp Clin Cancer Res. 28:75–78.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang C, Yuan X, Li Z, Tian Z, Zhan X,
Zhang J and Li X: VE-statin/Egfl7 siRNA inhibits angiogenesis in
malignant glioma in vitro. Int J Clin Exp Pathol. 7:1077–1084.
2014.PubMed/NCBI
|
|
25
|
Rajesh M, Mukhopadhyay P, Godlewski G,
Bátkai S, Haskó G, Liaudet L and Pacher P:
Poly(ADP-ribose)polymerase inhibition decreases angiogenesis.
Biochem Biophys Res Commun. 350:1056–1062. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Neo JH, Malcontenti-Wilson C, Muralidharan
V and Christophi C: Effect of ACE inhibitors and angiotensin II
receptor antagonists in a mouse model of colorectal cancer liver
metastases. J Gastroenterol Hepatol. 22:577–584. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiao M, Tang Y, Wang YL, Yang L, Li X,
Kuang J and Song GL: ART1 silencing enhances apoptosis of mouse
CT26 cells via the PI3K/Akt/NF-κB pathway. Cell Physiol Biochem.
32:1587–1599. 2013.
|
|
29
|
Xu JX, Wang YL, Tang Y and Xiong W: RNA
interference of ART1 on colon cancer cell proliferation in mice and
its mechanism. Tumor. 32:949–954. 2012.
|
|
30
|
Wei X, Tang Y, Wang Y and Xu J-X: Effects
of ART1 gene silencing on the ability of CT26 cellular matrix
adhesion and migration. Fudan Univ J Med Sci. 40:328–334. 2013.
|
|
31
|
Imura S, Miyake H, Izumi K, Tashiro S and
Uehara H: Correlation of vascular endothelial cell proliferation
with microvessel density and expression of vascular endothelial
growth factor and basic fibroblast growth factor in hepatocellular
carcinoma. J Med Invest. 51:202–209. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Raspollini MR, Castiglione F, Garbini F,
Villanucci A, Amunni G, Baroni G, Boddi V and Taddei GL:
Correlation of epidermal growth factor receptor expression with
tumor microdensity vessels and with vascular endothelial growth
factor expression in ovarian carcinoma. Int J Surg Pathol.
13:135–142. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Eriksson K, Magnusson P, Dixelius J,
Claesson-Welsh L and Cross MJ: Angiostatin and endostatin inhibit
endothelial cell migration in response to FGF and VEGF without
interfering with specific intracellular signal transduction
pathways. FEBS Lett. 536:192242003. View Article : Google Scholar
|
|
34
|
Su Y, Guan XQ, Liu FQ and Wang YL: The
effects of MIBG on the invasive properties of HepG2 hepatocellular
carcinoma cells. Int J Mol Med. 34:842–848. 2014.PubMed/NCBI
|
|
35
|
Bondarenko VA and Yamazaki A:
Characterization of argininespecific mono-ADP ribosyltransferase
isolated from frog retina and its function in signal transduction.
Invest Ophthalmol Vis Sci. 50:5440–A391. 2009.
|
|
36
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Montalto G, Cervello M, Nicoletti F, Fagone P, Malaponte
G, Mazzarino MC, et al: Mutations and deregulation of
Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascades which alter therapy
response. Oncotarget. 3:954–987. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hart JR and Vogt PK: Phosphorylation of
AKT: a mutational analysis. Oncotarget. 2:467–476. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sheng S, Qiao M and Pardee AB: Metastasis
and AKT activation. J Cell Physiol. 218:451–454. 2009. View Article : Google Scholar
|
|
40
|
Hawkins PT, Anderson KE, Davidson K and
Stephens LR: Signalling through class I PI3Ks in mammalian cells.
Biochem Soc Trans. 34:647–662. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Agani F and Jiang BH: Oxygen-independent
regulation of HIF-1: novel involvement of PI3K/AKT/mTOR pathway in
cancer. Curr Cancer Drug Targets. 13:245–251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Semenza GL: HIF-1: Upstream and downstream
of cancer metabolism. Curr Opin Genet Dev. 20:51–56. 2010.
View Article : Google Scholar :
|
|
43
|
Lu X and Kang Y: Hypoxia and
hypoxia-inducible factors: master regulators of metastasis. Clin
Cancer Res. 16:5928–5935. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rohwer N and Cramer T: HIFs as central
regulators of gastric cancer pathogenesis. Cancer Biol Ther.
10:383–385. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kitajima Y and Miyazaki K: The critical
impact of HIF-1α on gastric cancer biology. Cancers (Basel).
5:15–26. 2013. View Article : Google Scholar
|
|
46
|
Lee BL, Kim WH, Jung J, Cho SJ, Park JW,
Kim J, Chung HY, Chang MS and Nam SY: A hypoxia-independent
up-regulation of hypoxia-inducible factor-1 by AKT contributes to
angiogenesis in human gastric cancer. Carcinogenesis. 29:44–51.
2008. View Article : Google Scholar
|
|
47
|
Chun SY, Johnson C, Washburn JG,
Cruz-Correa MR, Dang DT and Dang LH: Oncogenic KRAS modulates
mitochondrial metabolism in human colon cancer cells by inducing
HIF-1α and HIF-2α target genes. Mol Cancer. 9:2932010. View Article : Google Scholar
|
|
48
|
Pugh CW and Ratcliffe PJ: Regulation of
angiogenesis by hypoxia-inducible factor-1: role of the HIF system.
Nat Med. 9:677–684. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lang SA, Gaumann A, Koehl GE, Seidel U,
Bataille F, Klein D, Ellis LM, Bolder U, Hofstaedter F, Schlitt HJ,
et al: Mammalian target of rapamycin is activated in human gastric
cancer and serves as a target for therapy in an experimental model.
Int J Cancer. 120:1803–1810. 2007. View Article : Google Scholar : PubMed/NCBI
|