Introduction
Streptococcus pneumoniae (S. pneumoniae),
which is a major pathogen found in infants, the elderly and the
immunocompromised, is responsible for more than one third of the
two million global annual deaths of children following acute
respiratory infections. The currently available
polysaccharide-based vaccines provide an antibody-dependent
protection (1) and significantly
reduce morbidity and mortality (2). The pneumococcal polysaccharide
vaccine Pneumovax 23 (PPV23), consists of 23 pneumococcal
polysaccharides from the most common disease-causing serotypes.
However, PPV23 does not provide long-lasting protection and is
ineffective in infants. The polysaccharide-protein conjugate
vaccine, which includes up to 13 pneumococcal polysaccharides,
elicits long-term protection, but provides only a limited coverage,
as the elicited immune response is serotype-specific (1). These inherent disadvantages have
motivated the search for other bacterial immunogens, such as
bacterial proteins, capable of eliciting a protective immune
response against S. pneumoniae (3,4).
Probably the most important concern today in the
field of vaccine development is our inadequate understanding
(3) of how to induce a specific,
potent and durable protective immune response (5). The majority of available vaccines
confer protection through antibodies, the levels of which are
defined as correlates of protection. Although the correlates of
immune response to S. pneumoniae proteins are emerging,
studies have found that the protection elicited by carriage or
immunization with whole-cell vaccines is CD4+ T-cell
dependent and antibody-independent (6,7),
and has been suggested to involve mainly the Th17 arm (8). However, other studies have shown
that both the Th17 and the Th1 arms are necessary for the
development of the protective immune response, whereas the Th2
response is negligible (9,10).
A variety of proteins displayed on the cell wall of
S. pneumoniae have been found to confer protection against
pneumococcal infection. Some of these proteins are adhesins, e.g.,
choline binding protein A (C bpA) (11,12), pneumococcal choline binding
protein A (PcpA) (13),
pneumococcal histidine triad protein D (PhtD) (3), pneumococcal surface adhesin A (PsaA)
(14) and pneumococcal serinerich
repeat protein (PsrP) (15),
while others are involved in various other bacterial functions
e.g., pneumococcal surface protein A (PspA), which reduces
complement deposition on the bacterium (16), protein required for cell wall
separation of group B streptococcus (PcsB), which is involved in
bacterial cell division; or serine/threonine protein kinase (StkP),
which is involved in bacterial morphogenesis (10,17).
Fructose-1, 6-bisphosphate aldolase (FBA) is a
glycolytic enzyme that is highly conserved (99%) among S.
pneumoniae strains. FBA has also been found to localize to the
surface of different pathogens, including Gram-positive bacteria,
Gram-negative bacteria and parasites (18–21). In S. pneumoniae, FBA
surface localization was demonstrated by biochemical fractionation
of the cell wall and by the flow cytometry of live bacteria
immunostained with mouse antisera against recombinant (r)FBA
(22,23). FBA has also been shown to function
as an adhesin in S. pneumoniae and in Neisseria
meningitidis (19,24). Furthermore, Flamingo cadherin
receptor (FCR), a cell adhesion molecule, was found to be a S.
pneumoniae FBA target receptor. Additionally, a peptide derived
from FCR was found to inhibit S. pneumoniae adhesion in
vitro and prevent disease development in vivo in a mouse
model (24). It has been found
that FBA elicits an age-dependent antigenicity in infants and that
it is immunogenic in mice (22,23). Finally, immunizing mice with rFBA
in the presence of alum has been shown to elicit a protective
immune response against a lethal challenge with S.
pneumoniae (22). Thus, rFBA
emerges as a promising vaccine candidate against S.
pneumoniae.
Aluminum-based compounds are the major adjuvants
used, to date, in human vaccination. Although they induce a good
Th2 antibody response to the immunogen, they also inhibit humoral
and cellular Th1 and Th17 responses. Alum salts are potent
inhibitors of Toll-like receptor (TLR) agonist-induced interleukin
(IL)-12 secretion, an effect that is mediated by alum-driven
phosphoinositide (PI)3 kinase signaling. Moreover, alum salts have
been shown to reduce the ability of the immune system to promote
Th17 responses due to their effect on the production of IL-23
(25). Thus, we hypothesized that
the protective effects elicited by alum salts are not
unsurpassed.
In the present study, we explored the ability of
rFBA to protect mice against an S. pneumoniae challenge and
investigated the cytokine profiles elicited in mice immunized with
rFBA. In addition, we evaluated the production of functional
protective antibodies capable of protecting the host against an
S. pneumoniae challenge.
Materials and methods
Reagents
Unless otherwise stated, all chemicals and
biochemicals were of the highest purity available and were
purchased from Sigma-Aldrich (St. Louis, MS, USA).
Bacterial strains and growth
conditions
The encapsulated S. pneumoniae serotype 3
strain WU2 (25) was grown as
previously described (26).
Briefly, pneumococci were grown in Todd-Hewitt broth supplemented
with 0.5% yeast extract (THY) or on blood agar plates.
Mice
Seven-week-old female BALB/cOlaHsd mice (n=121;
thereafter referred to as BALB/c mice; Harlan Laboratories,
Rehovot, Israel) and 7-week-old CBA/CaHN-Btkxid
mice (CBA/Nxid; Jackson Laboratories, Bar Harbor,
ME, USA) were used in this study. The experiments were performed at
one of two locations: i) at the animal facility of Ben-Gurion
University of the Negev, Beer-Sheva, Israel, in accordance with the
guidelines and approval of its Institutional Animal Care and Use
Committee (IACUC) (permit no. 53.08.08); or ii) at the University
of Mississippi Medical Center, Jackson, MS, USA, in accordance with
the guidelines and approval of the relevant IACUC (permit no.
00163).
In all the experiments, the mice were euthanized
with CO2. In the survival experiments, the mice were
monitored for fur appearance, social involvement, and their eating
and drinking ability. Any mouse demonstrating an apparent illness,
manifested by the appearance of bristled fur, social disengagement,
or an inability to eat or drink, was euthanized with
CO2.
Cloning, expression and purification of
rFBA
HAT-tagged rFBA was cloned as previously described
(22). To prevent any alterations
in the immune profile, an untagged protein was cloned, purified and
then used for examining the cytokine profile elicited by rFBA and
for the analysis of the IgG1/IgG2a experiments. The gene from the
locus tag SP_RS02975 that codes for FBA was amplified from the
TIGR4 strain with the following primers: forward,
5′-ATGGGATCCATATGGCGATTGTGTCTGCAGAA-3′ (Nde1); and reverse,
5′-CATGGAGCTCGCTCAGCTTATTACGCTTTGCCTTCG-3′ (Bpu11021). The
amplified product was inserted into the pET32a+ vector and
transformed in DH5α UltraMAX ultracompetent Escherichia coli
(E. coli) cells (Invitrogen, San Diego, CA, USA).
Ampicillin-resistant transformants were selected and the vector was
purified with a Qiagen HiSpeed Plasmid Maxi kit (Qiagen GmbH,
Hilden, Germany). The pET32a+fba vector was transformed into E.
coli host expression strain BL21(DE3) pLysS (Novagen, Jaffrey,
NH, USA). The identity of the inserts was confirmed by sequencing.
The protein was prepared under denaturing conditions and refolded
under physiological conditions. rFBA was purified by anion-exchange
and size-exclusion chromatography. rFBA identity and purity were
confirmed by matrix assisted laser desorption/ionization
time-of-flight (MALDI-TOF) mass spectrometry. The
lipopolysaccharide (LPS) concentration in the rFBA protein was
<8.9 EU/mg.
Preparation of DNA vaccine
The gene SP_0605 coding for FBA was amplified from
the TIGR4 strain and the following primers were used: forward,
5′-TCGGATCCATGGCAATCGTTTCAGCAGA-3′ (BamH1); and reverse,
5′-TCGAATTCTGCTTTACCTTCTGAACCGA-3′ (EcoR1). The amplified
and BamH1- and EcoR1- (Takara Bio Inc., Otsu, Japan)
digested DNA fragments were cloned into the pVAC plasmid, and the
pVACfba was transformed in DH5α UltraMAX
ultracompetent E. coli cells (both from Invitrogen).
Zeocin™-resistant transformants were cultured and the vector was
purified from the DH5α cells with a Qiagen HiSpeed Plasmid Maxi kit
(Qiagen GmbH).
Immunization of mice with rFBA for the
survival assay
The BALB/c mice were immunized in one of three ways:
i) subcutaneously (SC) with 25 µg of HAT-rFBA plus complete
Freund's adjuvant (CFA) for the first immunization (day 0) and with
incomplete Freund's adjuvant (IFA) in two subsequent booster
immunizations (days 14 and 28; n=25); ii) intramuscularly (IM) with
50 µg pVACfba (n=21); or iii)
intraperitoneally (IP) with 25 µg of HAT-rFBA and 75
µl of the Imject Alum adjuvant (Pierce Biotechnology, Inc.,
Rockford, IL, USA; n=15) on days 0, 14 and 28. The control mice
were immunized with the respective adjuvant (CFA/IFA/IFA n=34,
pVAC, empty plasmid n=14, Imject alum, n=6). The results shown are
composed of the three different experiments performed at different
times; in each experiment, all three adjuvants were tested.
Immunization of mice with rFBA for
cytokine profile determination
The BALB/c mice were immunized on day 0 with 10
µg of untagged rFBA alone or with untagged rFBA plus CFA.
Booster immunizations of rFBA alone or of rFBA with IFA were
administered on days 7 and 21. The control mice were immunized with
CFA/IFA alone. All mice were injected SC on both inner thighs, 25
µl/thigh.
Inoculation of immunized mice
The immunized BALB/c mice were inoculated
intranasally with a lethal dose of 108 CFU of WU2.
Survival was monitored daily.
Preparation of antigen-presenting cells
(APCs)
The mononuclear cells obtained from the spleens of
naïve BALB/c mice were treated as previously described (27). Briefly, they were treated with rat
anti-mouse monoclonal anti-CD4 (GK1.5), anti-CD8 (YTS169) and
anti-Thy1.2 (30-H-12) antibodies, followed by the addition of a
low-toxicity rabbit serum, as a source of complement. The
monoclonal antibodies and the hybridomas were kindly provided by
Professor L. Eizenbach from the Weizmann Institute of Science,
Rehovot, Israel. The cells were treated with mitomycin C. The
verification of CD4 and CD8 cell depletion and of the purity of the
APCs was performed by flow cytometry.
Enrichment of CD4+ T-cells by
positive selection
CD4+ T-cells from the inguinal lymph
nodes, popliteal lymph nodes and from the spleen were obtained from
the mice in all immunization groups. The mice were euthanized and
the lymph nodes and spleens were mechanically disrupted.
CD4+ T-cells were enriched using magnetic beads
conjugated to rat anti-mouse CD4+ (L3T4) (Mini MACS
system; Miltenyi Biotec, Bisley, UK). CD4+ enrichment
(95%) was verified by flow cytometry.
Bone marrow-derived dendritic cells
(BMDCs)
The BMDCs were generated from bone marrow cells
obtained from BALB/c mice as described previously (28). Briefly, BMDCs were generated from
bone marrow, which was obtained from BALB/c mice by flushing
femoral cells with a 23-gauge needle with RPMI. Low-density
mononuclear bone marrow cells were isolated using red blood cell
lysing buffer (Sigma-Aldrich) and centrifuged. Cells
(5×106) were cultured in 10-cm tissue dishes with the
fresh complete Iscove's Modified Dulbecco's Medium (cIMDM) with
recombinant GM-CSF (100 ng/ml) or 20% lymphocyte conditioned medium
(LCM), respectively. The medium was changed every 2–3 days and
replaced with fresh medium supplemented with GM-CSF (100 ng/ml) or
20% LCM respectively. The following modification to the original
protocol is: the cells were grown in a complete RPMI medium and
were allowed to differentiate for 9 days prior to co-culture with
CD4+ T-cells.
Enrichment of CD4+ T-cells by
negative selection
CD4+ T-cells were harvested from the
spleens and from the inguinal and popliteal lymph nodes of mice
immunized with rFBA plus adjuvant, or with phosphate-buffered
saline (PBS) plus adjuvant. The mononuclear cells were first
separated with a standard Ficoll separation protocol.
CD4+ T-cells were negatively selected using the EasySep
CD4+ T-cell enrichment kit (StemCell Technologies, Inc.,
Vancouver, BC, Canada). The enrichment of CD4+ T-cells
(87–90% CD4+ cells out of 23,609 events) was verified by
flow cytometry.
Antibodies used for flow cytometry
In the present study, phycoerythrin (PE)-conjugated
rat anti-mouse CD11c, fluorescein isothiocyanate (FITC)-conjugated
rat anti-mouse B220, FITC-conjugated anti-mouse CD11b and
allophycocyanin-conjugated rat anti-mouse major histocompatibility
complex (MHC II) (IAbdq, I-Edk), all purchased from Pharmingen (San
Diego, CA, USA), constituted the antibodies used for the purity
verification of the APCs. The depletion of CD4+ and of
CD8+ T-cells was verified with a FITC-conjugated rat
anti-mouse CD4 and rat anti-mouse CD8 (YTS169) respectively,
detected using a FITC-conjugated AffiniPure F(ab′)2 fragment of
mouse anti-rat IgG (H+L) (Jackson ImmunoResearch Laboratories,
Inc., West Grove, PA, USA). The negative selection of
CD4+ T-cells was verified using PE-conjugated rat
anti-mouse CD4 and allophycocyanin-conjugated rat anti-mouse CD45
(eBioscience, San Diego, CA, USA). Stained cells were analyzed
using either FACSCalibur or FACS Canto II (Becton-Dickinson,
Mountain View, CA). The data were then analyzed using CELLQuest™
software (version 3.3; BD Biosciences, San Jose, CA, USA) or with
FlowJo software (version 6.3.4; Tree Star Inc., Ashland, OR, USA).
Fluorescence data were acquired using logarithmic amplification and
the reported fluorescence intensity units represent conversion of
channel values according to the logarithmic scale.
In vitro stimulation of CD4+
T-cells
CD4+ T-cell
proliferation
The APCs (1 × 105 cells) treated with
mitomycin C were incubated with increasing concentrations of rFBA
(0–2 µg/ml) for 30 min in RPMI supplemented with 10%
heat-inactivated fetal calf serum (FCS). An enriched
CD4+ T-cell population was added to the rFBA-pulsed APCs
(1:1) and incubated for 72 h at 37°C. CD4+ T-cell
proliferation was evaluated as previously described (27), with a slight modification, namely,
the extent of proliferation was determined using the BrdU ELISA kit
(Hoffman-La Roche, Nutley, NJ, USA) according to the manufacturer's
instructions. In these experiments we used 3 mice in each group and
the results are a summary of 2 independent experiments, thus, 12
mice were used to perform these experiments.
Cytokine secretion
BMDCs, differentiated as previously described
(28), were matured on cRPMI
(penicillin 100 U/ml, streptomycin 100 µg/ml, L-glutamine 2
mM, 2-mercaptoethanol 50 µm, FCS 10%) containing 100 ng/ml
LPS for 5 h, and then pulsed with 10 ng/ml rFBA for 24 h at 37°C.
The enriched CD4+ T-cells, obtained from rFBA-immunized
and PBS-immunized mice (5 × 106 cells), were co-cultured
with 10-day-stimulated BMDCs (1 × 106 cells) in
triplicate wells. The supernatants were collected at the denoted
times and were frozen at −20°C for later analysis. The cytokine
concentrations were determined with a Bio-Plex cytokine (11-plex)
assay (Bio-Rad Laboratories, Berkeley, CA, USA) according to the
manufacturer's instructions. In addition, the concentration of
interferon-γ (IFN-γ) was determined using an ELISA kit
(Becton-Dickinson) according to the instructions provided by the
manufacturer. In these experiments we used 3 mice in each group and
the results are a summary of 3 independent experiments, thus, 18
mice were used to perform these experiments.
Quantitative analysis of IgG1 and
IgG2a by ELISA
Two weeks after the final immunization, the BALB/c
mice were euthanized and individually bled from the heart. ELISA
was carried out in a 96-well plate coated with rFBA. Individual
samples of mouse serum were added to the rFBA-coated plates. The
concentrations of anti-rFBA mouse IgG1 and anti-mouse IgG2a were
determined with biotin-conjugated goat anti-mouse IgG1 and
biotin-conjugated goat anti-mouse IgG2a (Southern Biotechnology
Associates, Birmingham, AL, USA), respectively, together with
streptavidin-horseradish peroxide (HRP; Jackson ImmunoResearch
Laboratories, Inc.). Standard curves were generated for mouse IgG1
and IgG2a concentrations (Biolegends, San Diego, CA, USA). In these
experiments we used 3 mice in each group and the results are a
summary of 3 independent experiments, thus, 18 mice were used to
perform these experiments.
Immunization of rabbits with rFBA for
neutralization assay
Three-month-old New Zealand white (NZW) rabbits
(Harlan Laboratories) were immunized as previously described
(26). Briefly, 3-month-old white
albino rabbits (Harlan Laboratories) were immunized intramuscularly
(IM) with 200 µg HAT-rPtsA emulsified with CFA (1:1) in the
first immunization or with IFA in booster immunizations in 2-week
intervals. Two weeks after their final immunization, the rabbits
were exsanguinated, and sera were prepared. The specificity of the
antisera was confirmed by immunoblotting and ELISA.
Ex vivo neutralization of S.
pneumoniae with anti-rFBA antisera
The WU2 strain was incubated at 37°C for 1 h either
with diluted rabbit pre-immune or rabbit anti-rFBA antiserum.
Subsequently, 500 CFU of the bacteria were IP-inoculated into the
BALB/c mice. Survival was monitored daily. In this experiment 10
mice were inoculated with S. pneumoniae treated ex
vivo with sera obtained from rFBA/CFA/IFA immunized rabbits and
10 mice were inoculated with S. pneumoniae treated ex
vivo with sera obtained from pre-immune sera from rabbits.
The WU2 strain was incubated at 37°C for 1 h either
with 1:10 diluted CBA/Nxid mice pre-immune serum
or with 1:25 diluted serum obtained from rFBA-immunized
CBA/Nxid mice. Subsequently, 104 CFU
of bacteria were IV-inoculated into the BALB/c mice. Survival was
monitored continuously. In these experiments 10
CBA/Nxid were immunized with rFBA and 10 were
immunized with the adjuvant only as described above. Sera were
drawn and 10 mice were inoculated with bacteria treated ex
vivo with sera obtained from rFBA immunized mice and 10 mice
were inoculated with bacteria treated ex vivo with sera
obtained from adjuvant immunized mice.
Statistical analysis
Log-rank and Wilcoxon signed-rank test, as
determined by the GraphPad 6 software, were used to evaluate
survival. Differences in cytokine levels or cell proliferation
rates between two groups were analyzed with a two-tailed Student's
t-test. Values of p<0.05 were considered to indicate a
statistically significant difference.
Results
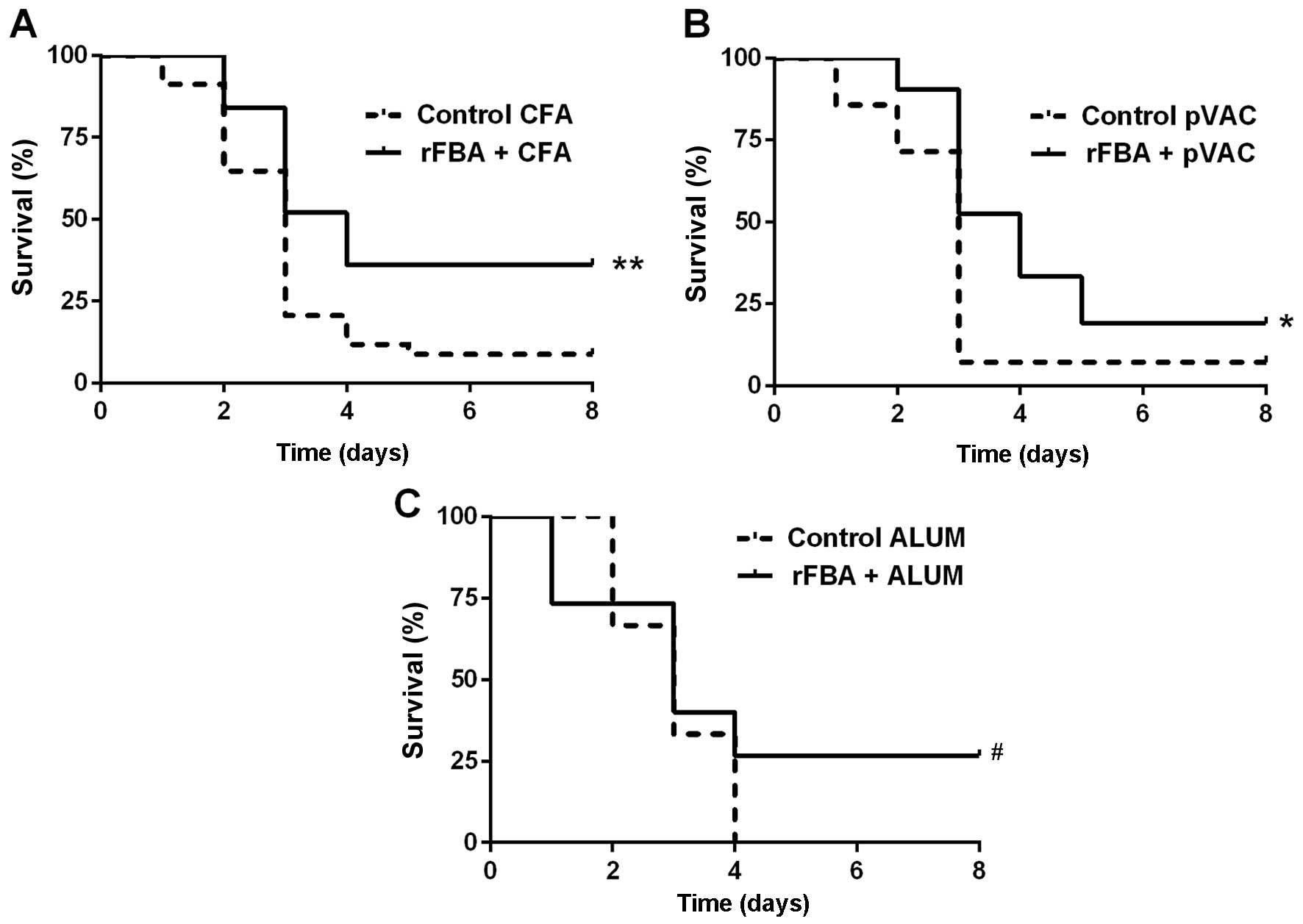
rFBA as a potential vaccine
The BALB/c mice immunized with rFBA + CFA/IFA/IFA
[namely, with CFA and then with two IFA boosters] or with the
adjuvants only (control) were challenged intranasally with a lethal
dose of encapsulated S. pneumoniae serotype 3 strain WU2.
The survival rates of the mice immunized with rFBA + CFA/IFA/IFA
were significantly higher than those of the control mice (Fig. 1A; log-rank test p<0.01). The
survival rates of the mice immunized with a DNA vaccine (namely,
with fba inserted into the pVAC plasmid;
pVACfba) were also significantly higher than
those of the mice immunized with the respective adjuvant alone
(Fig. 1B; log-rank test
p<0.05); however, the DNA vaccine was less protective (19%
survivors) than immunization with FBA + CFA/IFA/IFA (36%
survivors). The mice immunized with rFBA + alum demonstrated 26%
survival, in comparison with 0% survival in the mice immunized with
alum alone (Fig. 1C; Wilcoxon,
p<0.05).
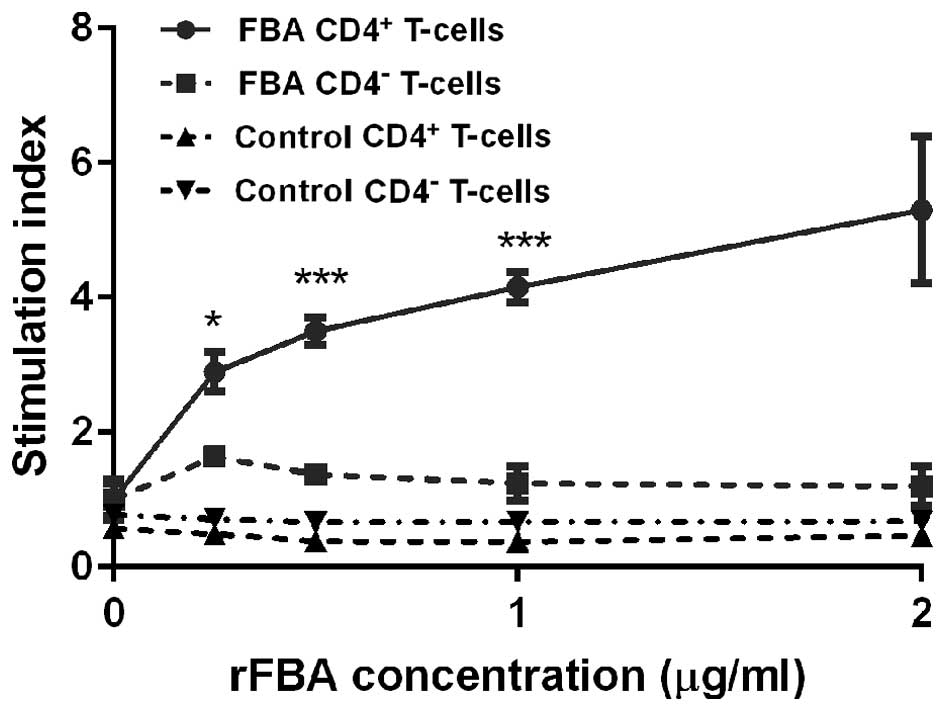
CD4+ T-cell proliferation
To examine the extent of CD4+ T-cell
proliferation in response to rFBA stimulation, CD4+
T-cells obtained from the rFBA + CFA/IFA/IFA-immunized mice were
co-cultured with naïve APCs treated in vitro with rFBA. The
control groups included CD4− T-cells derived from the
mice immunized with rFBA + CFA/IFA/IFA, CD4+ T-cells
obtained from the mice immunized with the adjuvant alone, and CD4
T-cells obtained from the mice immunized with the adjuvant alone.
The CD4+ T-cells derived from the mice immunized with
rFBA + CFA/IFA/IFA showed a dose-dependent increase in the
proliferation response upon re-stimulation with rFBA-pulsed APCs
(Fig. 2; p<0.05). The
CD4+ T-cells that were derived from the mice immunized
with the adjuvant alone and from the CD4+
T-cell-depleted groups did not respond to rFBA-pulsed APCs. These
results demonstrate that rFBA induces an antigen-specific
CD4+ T-cell proliferative response.
Cytokine secretion from memory
CD4+ T-cells in response to re-stimulation with
rFBA
To evaluate the nature of the immune response
following immunization with rFBA, we measured in vitro the
cytokine secretion by CD4+ T-cells in response to
stimulation with rFBA-pulsed BMDCs. Two types of co-cultures were
examined: CD4+ T-cells obtained from the mice immunized
with rFBA + CFA/IFA/IFA ('FBA culture), and CD4+ T-cells
obtained from the mice immunized with the adjuvant alone ('CFA'
culture). Additional controls included cultures with BMDCs only
('DC-only' culture), CD4+ T-cells only, obtained from
rFBA + CFA/IFA/IFA-immunized mice ('CD4-only' culture) and
CD4+ T-cells stimulated with concanavalin A (ConA)
('CD4+ConA' culture).
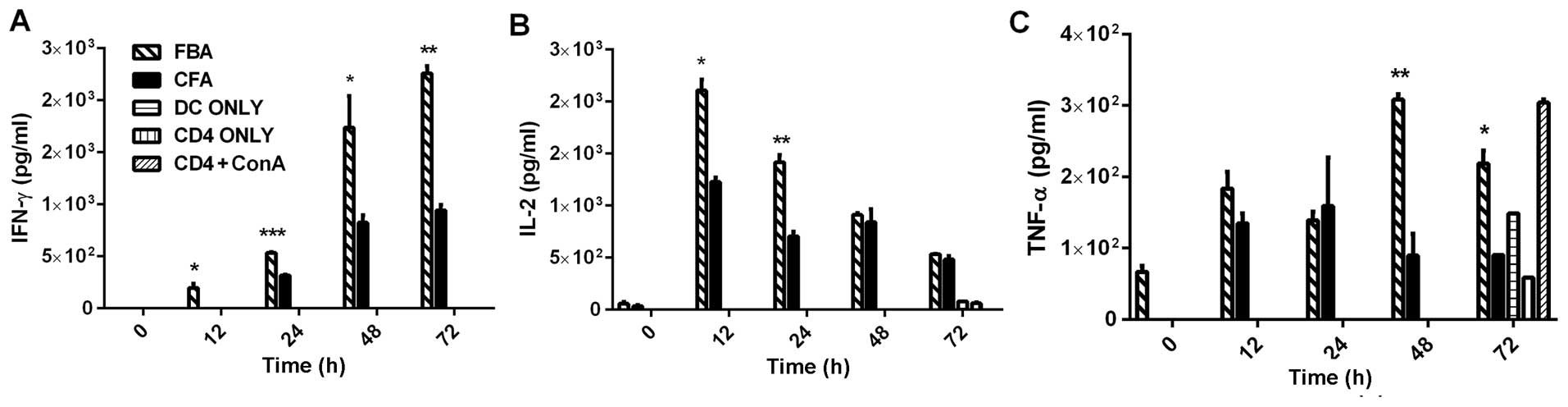
The supernatant of the FBA culture contained levels
of the Th1-related cytokines IFN-γ, IL-2 and tumor necrosis
factor-α (TNF- α) that were significantly higher than those in the
supernatant of any of the other cultures (Fig. 3). More specifically, in the FBA
culture, the levels of IFN-γ were significantly higher than
baseline (Fig. 3A; p<0.01)
between 12 and 72 h following re-stimulation, and they peaked at
48–72 h (Fig. 3A); the levels of
IL-2 peaked at 12 h (Fig. 3B;
p<0.05) and declined thereafter; and the levels of TNF-α peaked
at 48 h (Fig. 3C; p<0.01) and
were then reduced at 72 h (Fig.
3C; p<0.05). Ofnote, the TNF-α level increased at 72 h in
the CD4+ ConA group (as shown in Fig. 3C). No increase was observed in the
secretion of IFN-γ, IL-2 or TNF-α in the supernatants of the CFA,
DC-only, CD4-only, or CD4+ConA cultures.
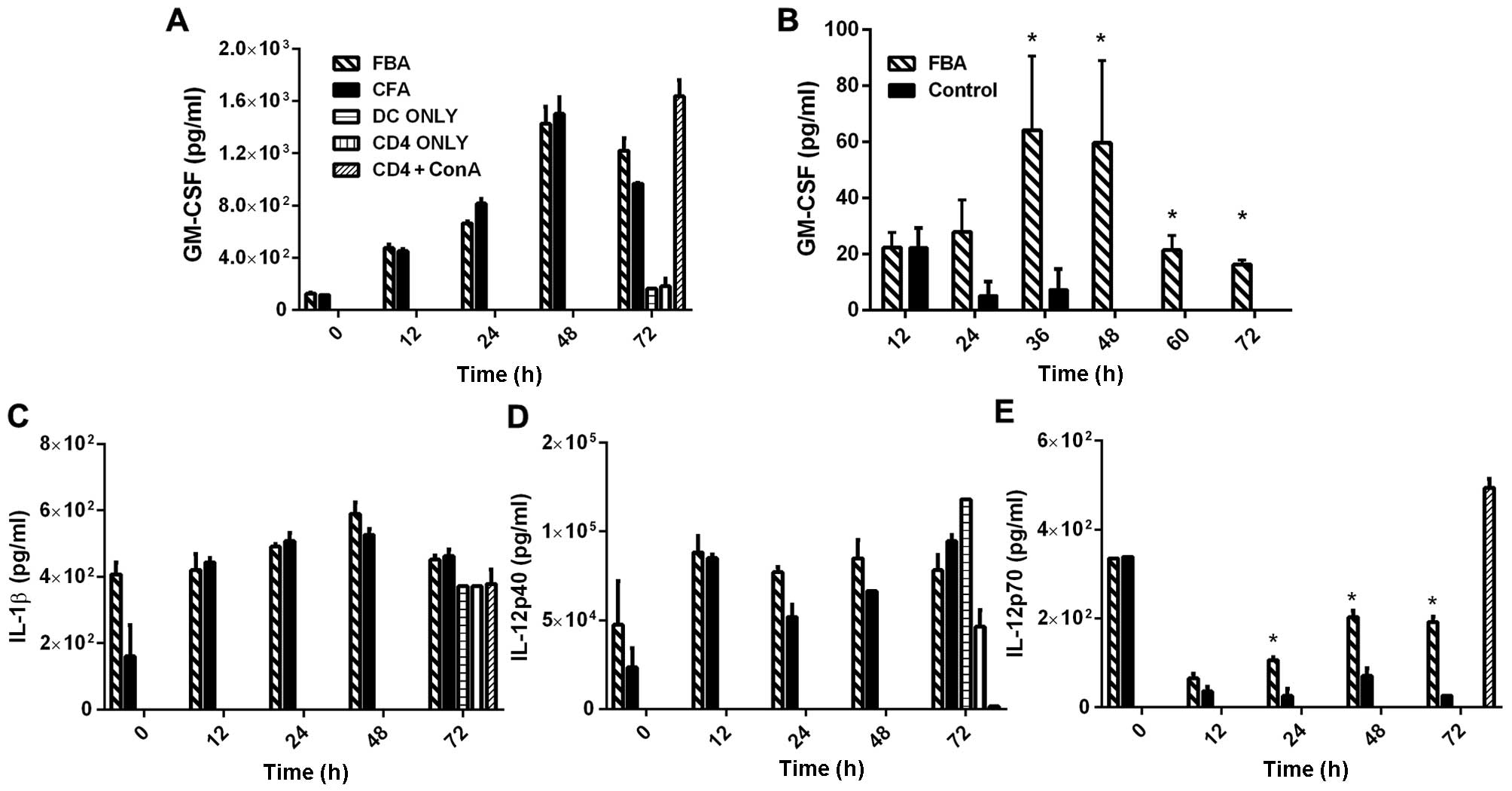
High levels of the Th1-related cytokines,
granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-1β
IL-12p40, co-cultured with the rFBA-stimulated BMDCs, were found in
the supernatants of the FBA and CFA cultures (Fig. 4A, C and D). No such stimulation
was observed in the BMDC-only or in the CD4-only cultures. Notably,
in the supernatant of CD4+ T-cells that were obtained
from the rFBA-immunized mice and co-cultured with naïve
spleen-derived APCs treated with rFBA, the levels of GM-CSF
significantly increased at 36 and 48 h following co-culture
initiation (Fig. 4B; p<0.05)
and declined thereafter (Fig. 4).
However, high levels of GM-CSF were observed in the
CD4+ConA cultures (Fig.
4A). IL-1β and IL-12p40 were detected in the supernatant of
DC-only cultures (Fig. 4C and D,
respectively) and in the supernatant of CD4-only cultures (at
<3%), probably due to residual APCs in the supernatant following
the negative selection procedure. Seventy-two hours following the
initiation of the cultures, the levels of IL-1β increased to a
similar extent in all the control cultures, while the levels of
IL-12p40 decreased in the CD4+ConA culture. The IL-12p70
levels were elevated in the FBA and CFA cultures at the time of
initiation of the co-cultures and declined at 12 h after culture
initiation. The levels of IL-12p70 in the FBA culture co-cultured
with BMDCs increased from the reduced levels at 12 h following the
initiation of the co-cultures and reached a plateau at 48 h
(Fig. 4E; p<0.05) ; this
pattern was not observed in any of the control cultures.
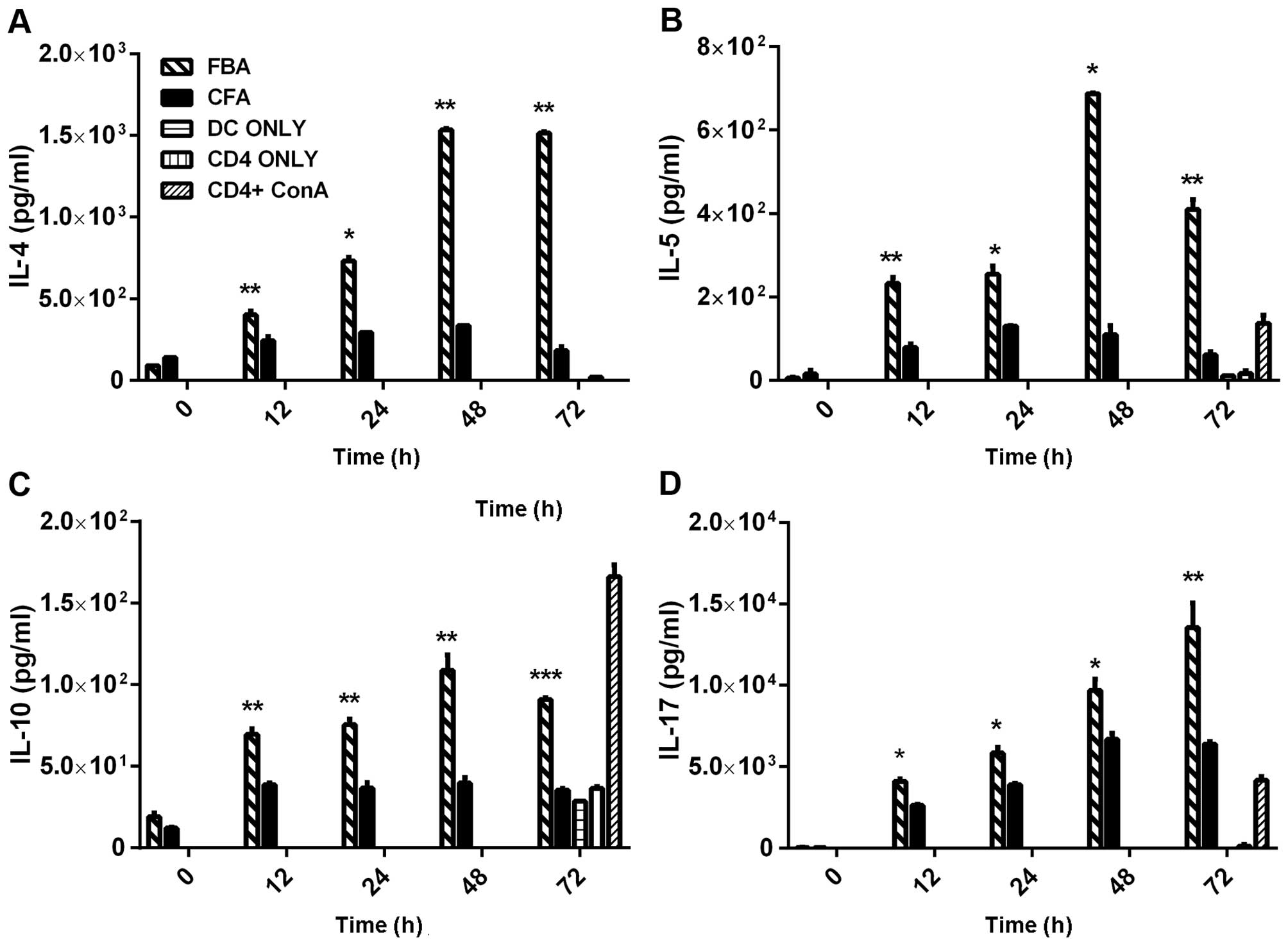
The supernatants of the FBA cultures demonstrated
significantly higher levels of the Th2-related cytokines, IL-4,
IL-5 and IL-10, at 12–48 h after co-incubation with the rFBA-pulsed
BMDCs (Fig. 5A-C, respectively).
As compared with their baseline levels, the levels of each of those
three cytokines peaked at 48 h (p<0.05); then, the levels of
IL-4 remained elevated (Fig. 5A;
p<0.05), whereas the levels of IL-5 (Fig. 5B) and IL-10 (Fig. 5C) declined, remaining higher than
their baseline levels (p<0.05 for each cytokine). The secretion
of IL-4, IL-5 and IL-10 was significantly decreased in all control
cultures, except for an increase in IL-10 in the
CD4+ConA culture (Fig.
5C).
The levels of the Th17-related cytokine, IL-17A,
increased significantly (from 34 pg/ml to 13,500 pg/ml; Fig. 5D; p<0.05) in the FBA culture at
72 h following the co-incubation of the CD4+ T-cells
with the rFBA-pulsed BMDCs (Fig.
5D). Such an increase was not observed in any of the control
cultures.
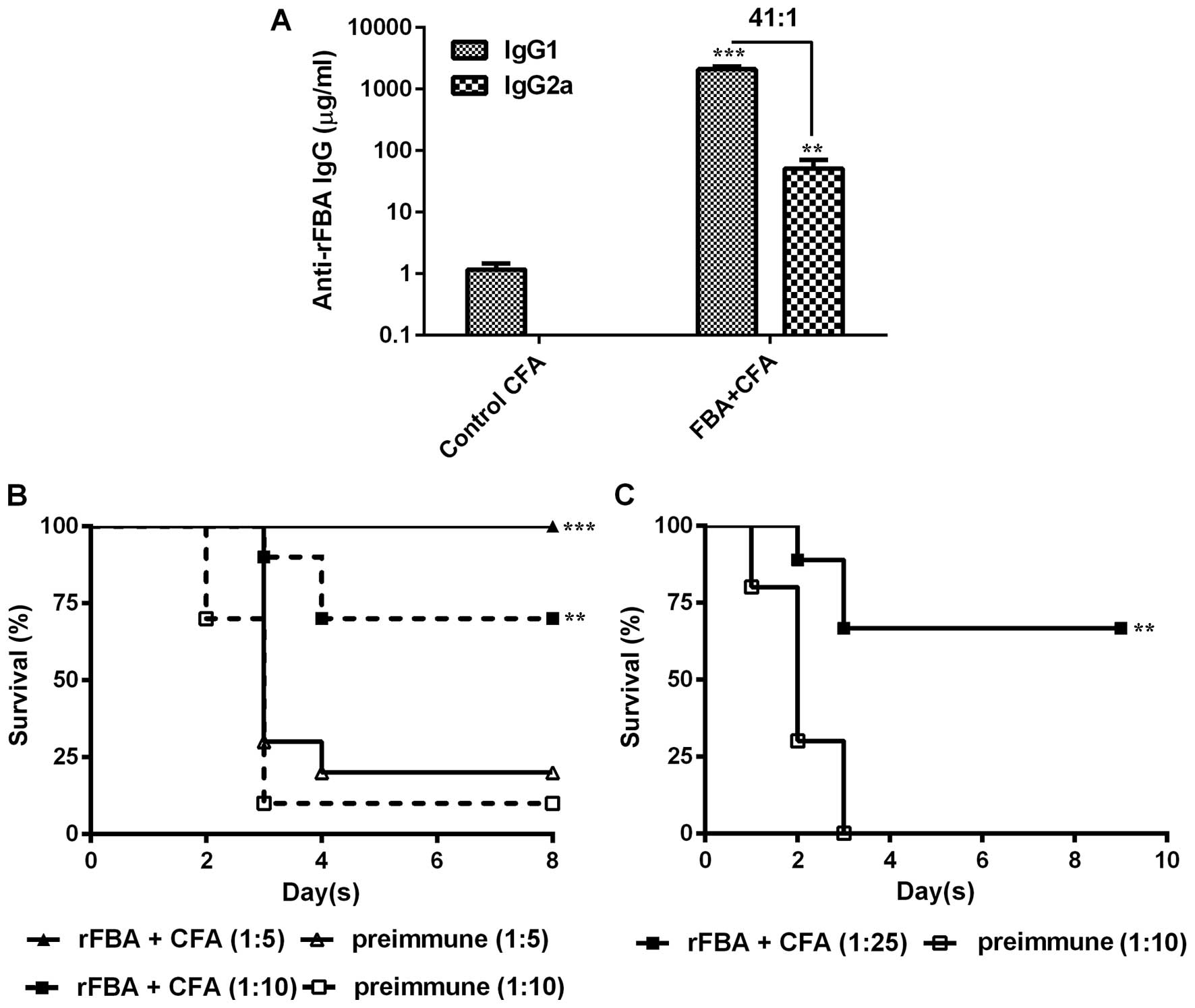
Concentrations of anti-rFBA IgG1 and
IgG2a subtypes and protective ability of anti-rFBA antiserum
In addition to the cytokine profile expressed in
vitro, we determined the concentrations of IgG1 and IgG2a
antibodies in sera obtained from the mice immunized with rFBA +
CFA/IFA/IFA or with CFA/IFA/IFA alone (control). The mice immunized
with rFBA + CFA/IFA/IFA showed anti-rFBA IgG1 (Fig. 6A; p<0.001) and anti-rFBA IgG2a
(Fig. 6A; p<0.05) titers that
were significantly higher than those in the control mice (Fig. 6A). Analysis of the concentration
ratio of IgG1 to IgG2a antibodies showed that immunization with
rFBA + CFA/IFA/IFA resulted in an IgG1/IgG2a ratio of 41.
We then investigated the ability of the serum
obtained from the rFBA-immunized rabbits to neutralize S.
pneumoniae virulence. The BALB/c mice inoculated IP with WU2
bacteria that were pre-incubated ex vivo with rabbit
anti-rFBA antiserum survived significantly longer (Fig. 6B; p<0.01) than the mice
inoculated with bacteria treated with pre-immune rabbit serum
(Fig. 6B). In addition, the
BALB/c mice inoculated intravenously with a lethal dose of WU2
pre-incubated ex vivo with CBA/Nxid mouse
anti-rFBA antiserum survived significantly longer than the mice
inoculated with bacteria pre-incubated in vitro with mouse
pre-immune serum (Fig. 6C;
p<0.01).
Discussion
The surface localization of FBA has been previously
demonstrated by immunoblotting and MALDI-TOF analysis of thge
bacterial cell wall (22) and by
flow cytometric analysis of live bacteria stained with rabbit
anti-rFBA antibodies (24). rFBA
in the presence of alum has been shown to elicit a protective
immune response in a mouse lethal challenge model (22). Thus, similar to other glycolytic
enzymes (26,27), FBA was suspected to have important
functions other than its principal role as a cytoplasmic enzyme in
glycolysis. Indeed, FBA was later found to mediate the adhesion of
S. pneumoniae to host cells, as FBA and anti-FBA antibodies
inhibited S. pneumoniae adhesion to A549 cells (24). In addition, using a peptide
combinatorial library expressed in the filamentous phage, FCR was
identified as the putative receptor of FBA. A peptide derived from
FCR inhibited S. pneumoniae adhesion to A549 cells in
vitro and prevented disease development in vivo in the
mouse model following S. pneumoniae challenge (24). In the present study, we aimed to
elucidate the nature of the protective immune response elicited by
rFBA.
A major challenge in the development of subunit
vaccines is the development of an efficient adjuvant or of a lymph
node delivery system that would replace the highly potent CFA
adjuvant (28–31). Aluminum salts elicit purely
Th2-type responses (28), DNA
vaccines elicit mainly Th1-type immune responses (by interacting
with TLR9) (29), and CFA/IFA
adjuvants elicit both a Th1-type (30) and a Th2-type (31) immune responses. A Th17-type
response was previously observed in experimental autoimmune
encephalomyelitis induced by immunization with myelin-derived
peptides in CFA (32), and,
therefore, we suspected that a Th17-type response will be induced
in the presence of CFA as the adjuvant. Th17 has recently been
shown to be associated with protective immunity against S.
pneumoniae (34). Despite
extensive efforts to develop new adjuvants, none has thus far
surpassed the adjuvancy capacity of CFA (5,33).
CFA/IFA adjuvants are widely used in preclinical studies
investigating candidates for a S. pneumoniae protein-based
vaccine (34–36).
In the present experiments, the protection observed
against a lethal S. pneumoniae challenge following
immunization with rFBA in the presence of CFA/IFA surpassed the
protection observed following immunization with pVA
Cfba or of rFBA in the presence of alum salt.
Intranasal inoculation of mice with a whole cell vaccine (WCV) in
the presence of cholera toxin or labile toxin, or of their
non-toxic mutated derivatives, has been shown to elicit protection
against intranasal inoculation with S. pneumoniae (37), which was Th17-dependent and
antibody-independent (38).
However, parenteral immunization with a WCV in the presence of alum
as the adjuvant, either subcutaneously or intramuscularly, elicited
a mucosal and systemic protection that was antibody dependent and
T-cell-independent (39). WCVs
contain pathogen-associated molecular patterns (PAMPs), such as
lipoprotein and lipoteichoic acid that interact with TLR2,
pneumolysin that interacts with TLR4, or peptidoglycan that
interacts with nucleotide-binding oligomerization domain containing
2 (NOD2), which may contribute to the protective immune response
elicited by the host (40).
Lipidated membrane proteins have also been shown to elicit an
increase in Th17 response and improve protection in comparison with
non-lipidated proteins as a result of TLR2 activation (41). We suspected that the better
protection following immunization with a single protein and with
CFA/IFA as the adjuvant will elicit protection of the three arms of
the immune response due to the mycobacterial components of CFA
(40).
In this study, initial experiments performed on
CD4+ T-cells co-incubated with rFBA-stimulated APCs
revealed a significantly increased proliferation of memory
CD4+ T-cells in comparison with the following three
control groups: CD4− T-cells obtained from the
rFBA-immunized mice, CD4+ T-cells obtained from the mice
immunized with the adjuvant only, and CD4− T-cells
obtained from the mice immunized with the adjuvant only. The extent
of CD4+ T-cell proliferation was similar to that
described for other bacterial protein immunogens (42,43).
The nature of the adaptive immune response to rFBA
was examined by analyzing the cytokine secretion profile in the
co-cultures of rFBA-pulsed naïve BMDCs and CD4+ T-cells
obtained from rFBA-immunized mice. In the supernatant of those
cultures, the levels of IL-2, IFN-γ, TNF-α, IL-12p70, IL-4, IL-5,
IL-10 and IL-17 were significantly higher than those in the
supernatants of the co-cultures of rFBA-treated BMDCs with
CD4+ T-cells obtained from the mice immunized with the
adjuvant only. The supernatants of the CD4+ T-cells
obtained from the rFBA-immunized mice in the absence of BMDCs did
not show an increased cytokine secretion. Similarly, the
CD4− T-cells obtained from either the rFBA + CFA/IFA/IFA
or the adjuvant only-immunized mice did not show increased cytokine
secretion. This finding confirms the importance of the cross-talk
between CD4+ T-cells and rFBA-treated dendritic cells
for the secretion of those cytokines, which are known to be
secreted from Th1-, Th2- and Th17-type memory CD4+
T-cells (44,45).
The secretion of IL-2 peaked as early as 12 h
following the initiation of the co-culture, and subsequently
decreased. This temporal pattern is in accordance with the known
secretion pattern of IL-2, which is rapid and transient, and the
decrease in IL-2 production was due to the negative feedback loop
required to limit IL-2 production during helper T-cell
differentiation (46). The
significantly increased secretion of IFN-γ that was detected in
this study is consistent with that of previous studies on children,
which demonstrated the importance of IFN-γ for the naturally
acquired immune response to S. pneumoniae (47) and for the acquired immune response
to S. pneumoniae proteins in mice (48). In this study, the secretion of
IL-2 and IFN-γ cytokines suggests the existence of a Th1-type
response to immunization with rFBA. The significantly increased
secretion of the proinflammatory cytokine TNF-α is in accordance
with previous research that demonstrated the importance of TNF-α in
the development of the adaptive humoral immune response to rPspA
following immunization with unencapsulated R36A (49).
The levels of GM-CSF, IL-12p40 and IL-1β observed
in the cultures of CD4+ T-cells obtained from the
rFBA-immunized mice were insignificant compared to those in
CD4+ T-cells obtained from the adjuvant-immunized mice.
Dendritic cell maturation was performed by exposing the cells to
LPS prior to stimulation with rFBA. Thus, the increased levels of
the GM-CSF, IL-12p40 and IL-1β cytokines were probably due to the
effect of LPS on the dendritic cells, as was described previously
(50). Indeed, a significant
increase was observed in the levels of GM-CSF in the supernatant of
CD4+ T-cells that were obtained from the rFBA-immunized
mice and co-cultured with APCs pulsed with rFBA but not treated
with LPS, as compared with those observed in the supernatant of
CD4+ T-cells that were obtained from the
adjuvant-immunized mice and co-cultured with rFBA-pulsed APCs
(Fig. 4B).
The levels of IL-12p70 detected in the culture
supernatant of CD4+ T-cells obtained from the
rFBA-immunized mice and co-cultured with the rFBA-treated BMDCs
were significantly higher than those detected in the culture
supernatant of CD4+ T-cells obtained from the
adjuvant-immunized mice in the presence of rFBA-treated BMDCs. This
phenomenon was not observed in the rFBA-treated BMDCs co-cultured
with CD4+ T-cells obtained from the adjuvant-immunized
mice, suggesting that a cross-talk exists between CD4+
T-cells and the rFBA-treated BMDCs. Indeed, such a cross-talk is
crucial in the development of Th1-type responses, as described
previously (51).
A significant increase in the secretion of IL-4,
IL-5 and IL-10 was observed in the supernatants of CD4+
T-cells obtained from the rFBA-immunized mice co-cultured with the
rFBA-treated BMDCs. The higher levels of IL-4 resemble those found
after immunizing mice with rGroEL (52), PspA (53) or PsaA (54). The increased secretion of IL-4,
IL-5 and IL-10 suggests that rFBA immunization elicits a Th2-type
immune response, which may be important for protection. Recently,
IL-10 has been shown to be important to avoid excessive tissue
inflammation and to improve host survival, despite the fact that
bacterial dissemination is less efficient in the presence of this
cytokine (55).
The secretion of the IL-17 cytokine by
CD4+ T-cells co-cultured with the rFBA-treated BMDCs
increased continuously throughout the 72 h of the experiment. This
finding is in agreement with the findings of recent studies
demonstrating the importance of IL-17 in protection against S.
pneumoniae. The seminal studies of Malley and Anderson
[reviewed in (8)] have shown
that, in mice, the acquired protective immune response to
pneumococcal colonization following intranasal immunization with
heat-killed unencapsulated bacteria depends on CD4+
T-cells, and particularly on the Th17 arm of the immune response;
however, it is antibody-independent (39).
In the present study, cytokines from the Th1, Th2
and Th17 arms of the immune response were induced following
immunization with a single protein, rFBA. These results confirm
previous findings obtained in vivo in the bronchoalveolar
lavage and the serum of mice immunized with Lactobacillus
lactis expressing the PppA protein from S. pneumoniae
(56). Other studies performed
with human CD4+ T-cells have demonstrated the dominance
of both the Th17-type and Th1-type immune responses to S.
pneumoniae (9,10) or to S. pneumoniae proteins
(4,50). Parenteral immunization with
attenuated WCV in the presence of alum elicited Th1-, Th2- and
Th17-type cytokine expression that was an order of magnitude lower
than that found in the present study (57). An alum-adjuvanted
Staphylococcus aureus antigen [(clumping factor A (ClfA)]
induced IL-17-dependent protection against Staphylococcal challenge
(58). Therefore, the ability of
alum to promote Th17-type responses appears to depend on the nature
of the vaccine antigen; in particular, antigens with inherent
immunomodulatory activity, such as killed bacteria, may synergize
with the adjuvant to promote Th17-type responses.
ConA was used in the present study as a polyclonal
non-specific stimulator for cytokine secretion. ConA is a member of
the legume lectin family and binds specifically to internal and
terminal α-D-mannosyl and α-D-glucosyl groups found in various
sugars, glycoproteins and glycolipids (59). ConA binds the CD3 component of
human T-cell receptors (60) and
most of its stimulatory properties are similar to those of anti-CD3
and anti-TCR antibodies (61).
The effect of ConA on T-cells is generally attributed to its
binding to and cross-linking of the CD3 component of the TCR,
thereby mimicking the physiological T-cell receptor ligand
MHC-peptide complex (62).
However, lectins are more potent stimulators of cell proliferation
than anti-CD3 or anti-CD4 antibodies, possibly as a result of
simultaneous activation of one or more co-stimulatory receptors. In
addition, differences in substrate phosphorylation downstream from
TCR following the binding of an MHC-peptide complex, in comparison
to ConA activation of CD4+ T-cells, were reported
(63). The activation of T-cells
with ConA resulted in reduced secretion of IL-2 and IFN-γ in
comparison with stimulation with an antigen (64), suggesting that antigen-MHC or ConA
activation of CD3 stimulates different levels of cytokine
expression.
The type of antibody produced against rFBA and the
importance of antibodies to protection were analyzed in the sera
obtained from the rFBA-immunized mice. Immunizing mice with rFBA
resulted in a median IgG1/IgG2a ratio of 41, suggesting a primary
Th2 response resulting in IgG1 antibody production, with a
substantial Th1 response resulting in IgG2a antibody production. In
a previous study, the immunization of BALB/c mice with rPspA + alum
induced an IgG1/IgG2a ratio of 1024, suggesting a strong Th2-biased
response. However, immunization with a pspA DNA vaccine
induced an IgG1/IgG2a ratio of 2, suggesting a strong Th1 bias
(65). Th1- and Th2-type immune
responses are mutually exclusive in the presence of all the
components of the immune system in vivo. Thus, the Th1 Th2
exclusion is never absolute but, rather, relative. The induction of
both Th1- and Th2-type responses in vivo is in accordance
with our in vitro results, in which Th1- and Th2-related
cytokines were found upon exposure of the CD4+ T-cells
to rFBA-pulsed BMDCs. The expression of Th1- and Th2-type cytokines
appeared simultaneously; suggesting that memory T-cells from these
arms of the immune response are present in the rFBA-immunized mice.
Activation in vitro with the rFBA-pulsed BMDCs elicited
cytokine production from the Th1, Th2 and Th17 arms of the immune
response. In vivo, re-exposure to an immunogen/pathogen may
culminate in higher expression of one of the arms of the immune
response relative to the others due to activation of control
mechanisms that are lacking in the in vitro system.
Furthermore, rabbit and mouse anti-rFBA antisera pre-incubated
ex vivo with S. pneumoniae significantly neutralized
bacterial virulence and thereby protected the mice against a S.
pneumoniae lethal challenge, as compared with bacteria
pre-incubated with pre-immune serum. This neutralization may result
from interference in bacterial adhesion to host cells (66) and suggests that the antibodies
also enhance phagocytosis of bacteria that shed their capsule
during adhesion.
We conducted our experiments with both the rabbit
and the mouse sera since different species have different
immunoglobulin protein profiles. Humans and mice have five antibody
isotypes (IgA, IgD, IgE, IgG and IgM), whereas, in rabbits, four
isotypes (IgA, IgE, IgG and IgM) have been identified to date, to
the best of our knowledge. In addition, rabbits have only one IgG
subclass, whereas mice have five subclasses (IgG1, IgG2a, IgG2b,
IgG2c and IgG3) and humans have four (IgG1, IgG2, IgG3 and IgG4).
We have previously demonstrated that anti-rFBA antisera inhibit the
adhesion of S. pneumoniae to cultured lung-derived
epithelial cells (24).
Similarly, antibodies to other S. pneumoniae adhesins, among
which are PcpA and PsrP, inhibited the adhesion of S.
pneumoniae to human respiratory epithelial cells (15,67), and antibodies against PcpA were
found to protect mice against a pneumococcal challenge (15). The exposure of bacterial cell-wall
proteins occurs during bacterial adhesion to the host as a result
of capsular shedding (66), which
enables access and binding of the antibodies to their respective
proteins and, possibly, facilitates their neutralization.
Therefore, the observed protection of mice against a S.
pneumoniae challenge by using anti-FBA antibodies may result
from inhibition of bacterial adhesion to the host.
Acknowledgments
This study was supported by grants 4476, 5540 and
3000003867 from the Israel Ministry of Health to Y.M.N.; grant
80904101 from BG Negev Technologies, Ben-Gurion University of the
Negev, to Y.M.N.; grant 2506 from the Center of Emerging Diseases
to Y.M.N.; and grant 613/04 from The Israel Academy of Science to
Y.M.N., the Israeli Ministry of Commerce and Industry to Y.M.N.,
the Israeli Ministry of Commerce and Industry to NasVax and Y.M.N.,
CAREPNEUMO from the European Commission to NasVax and Y.M.N. The
study and some of the authors were partially funded by commercial
companies (Nasvax Ltd. and Protea Vaccine Technologies Ltd.)
through the BG Negev Technologies that is the commercialization arm
of Ben-Gurion University of the Negev. The rFBA protein that is
discussed in the manuscript is protected by several pending
patents: US WO 03/082183; EP 1490104; PCT WO2010/029546 A2. The
protein vaccine is at an early stage of preclinical studies. There
are no marketed products to declare.
References
|
1
|
Dagan R, Poolman J and Siegrist CA:
Glycoconjugate vaccines and immune interference: a review. Vaccine.
28:5513–5523. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
van der Poll T and Opal SM: Pathogenesis,
treatment, and prevention of pneumococcal pneumonia. Lancet.
374:1543–1556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Denoël P, Philipp MT, Doyle L, Martin D,
Carletti G and Poolman JT: A protein-based pneumococcal vaccine
protects rhesus macaques from pneumonia after experimental
infection with Streptococcus pneumoniae. Vaccine. 29:5495–5501.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schmid P, Selak S, Keller M, Luhan B,
Magyarics Z, Seidel S, Schlick P, Reinisch C, Lingnau K, Nagy E and
Grubeck-Loebenstein B: Th17/Th1 biased immunity to the pneumococcal
proteins PcsB, StkP and PsaA in adults of different age. Vaccine.
29:3982–3989. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koff WC, Gust ID and Plotkin SA: Toward a
human vaccines project. Nat Immunol. 15:589–592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
van Rossum AM, Lysenko ES and Weiser JN:
Host and bacterial factors contributing to the clearance of
colonization by Streptococcus pneumoniae in a murine model. Infect
Immun. 73:7718–7726. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Basset A, Thompson CM, Hollingshead SK,
Briles DE, Ades EW, Lipsitch M and Malley R: Antibody-independent,
CD4+ T-cell-dependent protection against pneumococcal
colonization elicited by intranasal immunization with purified
pneumococcal proteins. Infect Immun. 75:5460–5464. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Malley R and Anderson PW:
Serotype-independent pneumococcal experimental vaccines that induce
cellular as well as humoral immunity. Proc Natl Acad Sci USA.
109:3623–3627. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mureithi MW, Finn A, Ota MO, Zhang Q,
Davenport V, Mitchell TJ, Williams NA, Adegbola RA and Heyderman
RS: T cell memory response to pneumococcal protein antigens in an
area of high pneumococcal carriage and disease. J Infect Dis.
200:783–793. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pido-Lopez J, Kwok WW, Mitchell TJ,
Heyderman RS and Williams NA: Acquisition of pneumococci specific
effector and regulatory CD4+ T cells localising within
human upper respiratory-tract mucosal lymphoid tissue. PLoS Pathog.
7:e10023962011. View Article : Google Scholar
|
|
11
|
Rosenow C, Ryan P, Weiser JN, Johnson S,
Fontan P, Ortqvist A and Masure HR: Contribution of novel
choline-binding proteins to adherence, colonization and
immunogenicity of Streptococcus pneumoniae. Mol Microbiol.
25:819–829. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mann B, Thornton J, Heath R, Wade KR,
Tweten RK, Gao G, El Kasmi K, Jordan JB, Mitrea DM, Kriwacki R, et
al: Broadly protective protein-based pneumococcal vaccine composed
of pneumolysin toxoid-CbpA peptide recombinant fusion protein. J
Infect Dis. 209:1116–1125. 2014. View Article : Google Scholar :
|
|
13
|
Glover DT, Hollingshead SK and Briles DE:
Streptococcus pneumoniae surface protein PcpA elicits protection
against lung infection and fatal sepsis. Infect Immun.
76:2767–2776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Talkington DF, Brown BG, Tharpe JA, Koenig
A and Russell H: Protection of mice against fatal pneumococcal
challenge by immunization with pneumococcal surface adhesin A
(PsaA). Microb Pathog. 21:17–22. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rose L, Shivshankar P, Hinojosa E,
Rodriguez A, Sanchez CJ and Orihuela CJ: Antibodies against PsrP, a
novel Streptococcus pneumoniae adhesin, block adhesion and protect
mice against pneumococcal challenge. J Infect Dis. 198:375–383.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Daniels CC, Kim KH, Burton RL, Mirza S,
Walker M, King J, Hale Y, Coan P, Rhee DK, Nahm MH and Briles DE:
Modified opsonization, phagocytosis, and killing assays to measure
potentially protective antibodies against pneumococcal surface
protein A. Clin Vaccine Immunol. 20:1549–1558. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Giefing C, Meinke AL, Hanner M, Henics T,
Bui MD, Gelbmann D, Lundberg U, Senn BM, Schunn M, Habel A, et al:
Discovery of a novel class of highly conserved vaccine antigens
using genomic scale antigenic fingerprinting of pneumococcus with
human antibodies. J Exp Med. 205:117–131. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pancholi V and Chhatwal GS: Housekeeping
enzymes as virulence factors for pathogens. Int J Med Microbiol.
293:391–401. 2003. View Article : Google Scholar
|
|
19
|
Tunio SA, Oldfield NJ, Berry A, Ala'Aldeen
DA, Wooldridge KG and Turner DP: The moonlighting protein
fructose-1,6-bisphosphate aldolase of Neisseria meningitidis:
surface localization and role in host cell adhesion. Mol Microbiol.
76:605–615. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu Z, Zhang W and Lu C: Immunoproteomic
assay of surface proteins of Streptococcus suis serotype 9. FEMS
Immunol Med Microbiol. 53:52–59. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McCarthy JS, Wieseman M, Tropea J, Kaslow
D, Abraham D, Lustigman S, Tuan R, Guderian RH and Nutman TB:
Onchocerca volvulus glycolytic enzyme fructose-1,6-bisphosphate
aldolase as a target for a protective immune response in humans.
Infect Immun. 70:851–858. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ling E, Feldman G, Portnoi M, Dagan R,
Overweg K, Mulholland F, Chalifa-Caspi V, Wells J and
Mizrachi-Nebenzahl Y: Glycolytic enzymes associated with the cell
surface of Streptococcus pneumoniae are antigenic in humans and
elicit protective immune responses in the mouse. Clin Exp Immunol.
138:290–298. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Portnoi M, Ling E, Feldman G, Dagan R and
Mizrachi-Nebenzahl Y: The vaccine potential of Streptococcus
pneumoniae surface lectin- and non-lectin proteins. Vaccine.
24:1868–1873. 2006. View Article : Google Scholar
|
|
24
|
Blau K, Portnoi M, Shagan M, Kaganovich A,
Rom S, Kafka D, Chalifa Caspi V, Porgador A, Givon-Lavi N, Gershoni
JM, et al: Flamingo cadherin: a putative host receptor for
Streptococcus pneumoniae. J Infect Dis. 195:1828–1837. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mori A, Oleszycka E, Sharp FA, Coleman M,
Ozasa Y, Singh M, O'Hagan DT, Tajber L, Corrigan OI, McNeela EA and
Lavelle EC: The vaccine adjuvant alum inhibits IL-12 by promoting
PI3 kinase signaling while chitosan does not inhibit IL-12 and
enhances Th1 and Th17 responses. Eur J Immunol. 42:2709–2719. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chhatwal GS: Anchorless adhesins and
invasins of Gram-positive bacteria: a new class of virulence
factors. Trends Microbiol. 10:205–208. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Henderson B and Martin A: Bacterial
virulence in the moonlight: multitasking bacterial moonlighting
proteins are virulence determinants in infectious disease. Infect
Immun. 79:3476–3491. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Garçon N and Van Mechelen M: Recent
clinical experience with vaccines using MPL- and QS-21-containing
adjuvant systems. Expert Rev Vaccines. 10:471–486. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moon JJ, Huang B and Irvine DJ:
Engineering nano- and microparticles to tune immunity. Adv Mater.
24:3724–3746. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Keler T, He L, Ramakrishna V and Champion
B: Antibody-targeted vaccines. Oncogene. 26:3758–3767. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu H, Moynihan KD, Zheng Y, Szeto GL, Li
AV, Huang B, Van Egeren DS, Park C and Irvine DJ: Structure-based
programming of lymph-node targeting in molecular vaccines. Nature.
507:519–522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Libbey JE and Fujinami RS: Experimental
autoimmune encephalomyelitis as a testing paradigm for adjuvants
and vaccines. Vaccine. 29:3356–3362. 2011. View Article : Google Scholar :
|
|
33
|
Delany I, Rappuoli R and De Gregorio E:
Vaccines for the 21st century. EMBO Mol Med. 6:708–720.
2014.PubMed/NCBI
|
|
34
|
Seiberling M, Bologa M, Brookes R, Ochs M,
Go K, Neveu D, Kamtchoua T, Lashley P, Yuan T and Gurunathan S:
Safety and immunogenicity of a pneumococcal histidine triad protein
D vaccine candidate in adults. Vaccine. 30:7455–7460. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Adamou JE, Heinrichs JH, Erwin AL, Walsh
W, Gayle T, Dormitzer M, Dagan R, Brewah YA, Barren P, Lathigra R,
et al: Identification and characterization of a novel family of
pneumococcal proteins that are protective against sepsis. Infect
Immun. 69:949–958. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wizemann TM, Heinrichs JH, Adamou JE,
Erwin AL, Kunsch C, Choi GH, Barash SC, Rosen CA, Masure HR,
Tuomanen E, et al: Use of a whole genome approach to identify
vaccine molecules affording protection against Streptococcus
pneumoniae infection. Infect Immun. 69:1593–1598. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lu YJ, Yadav P, Clements JD, Forte S,
Srivastava A, Thompson CM, Seid R, Look J, Alderson M, Tate A, et
al: Options for inactivation, adjuvant, and route of topical
administration of a killed, unencapsulated pneumococcal whole-cell
vaccine. Clin Vaccine Immunol. 17:1005–1012. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lu YJ, Leite L, Gonçalves VM, Dias WO,
Liberman C, Fratelli F, Alderson M, Tate A, Maisonneuve JF,
Robertson G, et al: GMP-grade pneumococcal whole-cell vaccine
injected subcutaneously protects mice from nasopharyngeal
colonization and fatal aspiration-sepsis. Vaccine. 28:7468–7475.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Malley R, Srivastava A, Lipsitch M,
Thompson CM, Watkins C, Tzianabos A and Anderson PW:
Antibody-independent, interleukin-17A-mediated, cross-serotype
immunity to pneumococci in mice immunized intranasally with the
cell wall polysaccharide. Infect Immun. 74:2187–2195. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kumar S, Ingle H, Prasad DV and Kumar H:
Recognition of bacterial infection by innate immune sensors. Crit
Rev Microbiol. 39:229–246. 2013. View Article : Google Scholar
|
|
41
|
Moffitt K, Skoberne M, Howard A,
Gavrilescu LC, Gierahn T, Munzer S, Dixit B, Giannasca P, Flechtner
JB and Malley R: Toll-like receptor 2-dependent protection against
pneumococcal carriage by immunization with lipidated pneumococcal
proteins. Infect Immun. 82:2079–2086. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu X, Wetzler LM and Massari P: The PorB
porin from commensal Neisseria lactamica induces Th1 and Th2 immune
responses to ovalbumin in mice and is a potential immune adjuvant.
Vaccine. 26:786–796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang Q, Bagrade L, Bernatoniene J, Clarke
E, Paton JC, Mitchell TJ, Nunez DA and Finn A: Low CD4 T cell
immunity to pneumolysin is associated with nasopharyngeal carriage
of pneumococci in children. J Infect Dis. 195:1194–1202. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mosmann TR and Coffman RL: TH1 and TH2
cells: Different patterns of lymphokine secretion lead to different
functional properties. Annu Rev Immunol. 7:145–173. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Harrington LE, Mangan PR and Weaver CT:
Expanding the effector CD4 T-cell repertoire: the Th17 lineage.
Curr Opin Immunol. 18:349–356. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Villarino AV, Tato CM, Stumhofer JS, Yao
Z, Cui YK, Hennighausen L, O'Shea JJ and Hunter CA: Helper T cell
IL-2 production is limited by negative feedback and STAT-dependent
cytokine signals. J Exp Med. 204:65–71. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
van den Biggelaar AH, Pomat WS,
Phuanukoonnon S, Michael A, Aho C, Nadal-Sims MA, Devitt CJ, Jacoby
PA, Hales BJ, Smith WA, et al: Effect of early carriage of
Streptococcus pneumoniae on the development of pneumococcal
protein-specific cellular immune responses in infancy. Pediatr
Infect Dis J. 31:243–248. 2012. View Article : Google Scholar
|
|
48
|
Ferreira DM, Darrieux M, Silva DA, Leite
LC, Ferreira JM Jr, Ho PL, Miyaji EN and Oliveira ML:
Characterization of protective mucosal and systemic immune
responses elicited by pneumococcal surface protein PspA and PspC
nasal vaccines against a respiratory pneumococcal challenge in
mice. Clin Vaccine Immunol. 16:636–645. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Khan AQ, Shen Y, Wu ZQ, Wynn TA and
Snapper CM: Endogenous pro- and anti-inflammatory cytokines
differentially regulate an in vivo humoral response to
Streptococcus pneumoniae. Infect Immun. 70:749–761. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wei WC, Su YH, Chen SS, Sheu JH and Yang
NS: GM-CSF plays a key role in zymosan-stimulated human dendritic
cells for activation of Th1 and Th17 cells. Cytokine. 55:79–89.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tourret M, Guégan S, Chemin K, Dogniaux S,
Miro F, Bohineust A and Hivroz C: T cell polarity at the
immunological synapse is required for CD154-dependent IL-12
secretion by dendritic cells. J Immunol. 185:6809–6818. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cao J, Zhang X, Gong Y, Zhang Y, Cui Y,
Lai X, Yin Y, Li M, Li D and Zhang L: Protection against
pneumococcal infection elicited by immunization with multiple
pneumococcal heat shock proteins. Vaccine. 31:3564–3571. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li Y, Wang S, Scarpellini G, Gunn B, Xin
W, Wanda SY, Roland KL and Curtiss R III: Evaluation of new
generation Salmonella enterica serovar Typhimurium vaccines with
regulated delayed attenuation to induce immune responses against
PspA. Proc Natl Acad Sci USA. 106:593–598. 2009. View Article : Google Scholar :
|
|
54
|
Miyaji EN, Dias WO, Gamberini M, Gebara
VC, Schenkman RP, Wild J, Riedl P, Reimann J, Schirmbeck R and
Leite LC: PsaA (pneumococcal surface adhesin A) and PspA
(pneumococcal surface protein A) DNA vaccines induce humoral and
cellular immune responses against Streptococcus pneumoniae.
Vaccine. 20:805–812. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Peñaloza HF, Nieto PA, Muñoz-Durango N,
Salazar-Echegarai FJ, Torres J, Parga MJ, Alvarez-Lobos M, Riedel
CA, Kalergis AM and Bueno SM: Interleukin-10 plays a key role in
the modulation of neutrophils recruitment and lung inflammation
during infection by Streptococcus pneumoniae. Immunology.
146:100–112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Vintiñi E, Villena J, Alvarez S and Medina
M: Administration of a probiotic associated with nasal vaccination
with inactivated Lactococcus lactis-PppA induces effective
protection against pneumoccocal infection in young mice. Clin Exp
Immunol. 159:351–362. 2010. View Article : Google Scholar :
|
|
57
|
Wu K, Yao R, Wang H, Pang D, Liu Y, Xu H,
Zhang S, Zhang X and Yin Y: Mucosal and systemic immunization with
a novel attenuated pneumococcal vaccine candidate confer serotype
independent protection against Streptococcus pneumoniae in mice.
Vaccine. 32:4179–4188. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Narita K, Hu DL, Mori F, Wakabayashi K,
Iwakura Y and Nakane A: Role of interleukin-17A in cell-mediated
protection against Staphylococcus aureus infection in mice
immunized with the fibrinogen-binding domain of clumping factor A.
Infect Immun. 78:4234–4242. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Poretz RD and Goldstein IJ:
Protein-carbohydrate interaction. On the mode of bonding of
aromatic moieties to concanavalin A, the phytohemagglutinin of the
jack bean. Biochem Pharmacol. 20:2727–2739. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chilson OP and Kelly-Chilson AE: Mitogenic
lectins bind to the antigen receptor on human lymphocytes. Eur J
Immunol. 19:389–396. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fleischer B: Activation of human T
lymphocytes II. Involvement of the T3 antigen in polyclonal T cell
activation by mitogenic lectins and oxidation. Eur J Immunol.
14:748–752. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Weiss A, Imboden J, Hardy K, Manger B,
Terhorst C and Stobo J: The role of the T3/antigen receptor complex
in T-cell activation. Annu Rev Immunol. 4:593–619. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Pani G, Colavitti R, Borrello S and
Galeotti T: Endogenous oxygen radicals modulate protein tyrosine
phosphorylation and JNK-1 activation in lectin-stimulated
thymocytes. Biochem J. 347:173–181. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Callahan TA and Moynihan JA: Contrasting
pattern of cytokines in antigen-versus mitogen-stimulated
splenocyte cultures from chemically denervated mice. Brain Behav
Immun. 16:764–773. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Vadesilho CF, Ferreira DM, Moreno AT,
Chavez-Olortegui C, Machado de Avila RA, Oliveira ML, Ho PL and
Miyaji EN: Characterization of the antibody response elicited by
immunization with pneumococcal surface protein A (PspA) as
recombinant protein or DNA vaccine and analysis of protection
against an intranasal lethal challenge with Streptococcus
pneumoniae. Microb Pathog. 53:243–249. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hammerschmidt S, Wolff S, Hocke A, Rosseau
S, Müller E and Rohde M: Illustration of pneumococcal
polysaccharide capsule during adherence and invasion of epithelial
cells. Infect Immun. 73:4653–4667. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Khan MN, Sharma SK, Filkins LM and
Pichichero ME: PcpA of Streptococcus pneumoniae mediates adherence
to nasopharyngeal and lung epithelial cells and elicits functional
antibodies in humans. Microbes Infect. 14:1102–1110. 2012.
View Article : Google Scholar : PubMed/NCBI
|