Introduction
Acute myocardial infarction (AMI) is caused by the
sudden blockage of the coronary blood supply and leads to the
hibernation and irreversible death of cardiomyocytes. It is usually
followed by myocardial fibrosis, which results in decreased
ventricular compliance, ventricular dilatation, and eventually,
heart failure (1). Cell therapy
using mesenchymal stem cells (MSCs) to improve the viability of
hibernated cardiomyocytes and reduce fibrosis is an attractive
prospect. Several clinical trials have highlighted the therapeutic
effects of MSC transplantation, by reducing the size of the infarct
area and improving the left ventricular ejection fraction (2,3).
However, as previously demonstrated in a rat model of AMI,
MSC-based therapy only achieved short-term benefits rather than a
long-term impact on cardiac function (4). This may be explained by the findings
of an in vivo animal study which revealed that less than 1%
of engrafted MSCs had survived by day 4 following transplantation
(5). The ischemic
microenvironment, together with risk factors, including anoxia, as
well as serum and glucose deficiency, contribute to the death of
transplanted MSCs, by activating cellular signaling mechanisms,
such as oxidative stress, endoplasmic reticulum (ER) stress and
changes in mitochondrial permeability. ER stress triggered by
ischemia is an important cause of cell death (6). Despite attempts to improve MSC
survival with growth factors, drug pre-treatment, a gene
transfection-activated survival pathway and by reducing
mitochondrial-mediated apoptosis (7–9),
few of these therapies have exerted beneficial effects on ER
stress-induced apoptosis.
Glucagon-like peptide-1 (GLP-1) is a peptide
secreted from L-cells in the small intestine and the proximal
colon. As a cognate receptor for GLP-1, GLP-1 receptor (GLP-1R) is
expressed in various types of tissue, such as the brain and the
pancreas tissue. Thus, GLP-1 exerts pleiotropic effects, including
the enhanced synthesis and release of insulin, enhanced satiety,
delayed gastric emptying and increased cellular survival (10). GLP-1 is rapidly cleaved by
dipeptidyl peptidase IV (DPPIV) and thus, it has a short half-life.
Exendin-4 (ex-4), a 39 amino acid agonist of GLP-1R, has similar
biochemical effects to GLP-1; however, it has a longer half-life
(11). At present, ex-4 is being
used to increase insulin production for the clinical treatment of
type 2 diabetes (12). Apart from
the insulinotropic effects of ex-4, it has been shown to protect
the heart from ischemia-reperfusion injury (13), and it has also been shown to
render cells resistant to ischemic-related injury in an
experimental model of transient cerebral ischemic damage (14). Previous research has demonstrated
that ex-4 attenuates atherosclerotic plaque formation by inhibiting
the inflammatory response in macrophages (15). Moreover, ex-4 has been shown to
improve the survival of several types of cells, such as β-cells,
cardiomyocytes and cholangiocytes (16–18). A growing body of evidence supports
the notion that ex-4 plays an important role in the regulation of
ER stress, thus exerting cytoprotective effects (16,19). However, to the best of our
knowledge, whether ex-4 protects MSCs from ischemia-induced
apoptosis and the involvement of ER stress in this process remains
unknown.
In consideration of the above-mentioned findings, we
hypothesized that ex-4 may confer resistance to apoptosis in MSCs.
In this study, we investigated the potential protective effects of
ex-4 in rat bone marrow-derived mesenchymal stem cells (BM-MSCs)
subjected to oxygen, glucose and serum deprivation (OGD)
conditions, as well as the underlying mechanisms.
Materials and methods
Animals and cell culture
This study was approved by the Institutional Animal
Care and Use Committee of Harbin Medical University (Harbin, China)
and was performed in strict accordance with the recommendations in
the Guide for the Care and Use of Laboratory Animals of the
National Institutes of Health (20). Male Sprague Dawley (SD) rats, 2–4
weeks old and weighing 50–60 g, were used in the present study. The
pancreases of the rats were isolated and RNA and protein were
extracted from the pancreatic tissue. The extraction and culture of
the rat MSCs was performed as previously described (9). Briefly, bone marrow was washed out
from the tibia and femur marrow cavities of the SD rats, and
suspended in sterile phosphate-buffered saline (PBS). Red cells
were then lysed and removed, and 5×105 cells were plated
in a 25 cm2 flask with F12/Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% fetal bovine serum (both from
HyClone, Logan, UT, USA) and 1% penicillin-streptomycin. Following
culture for 48 h, the suspended cells and medium were removed, and
the adherent MSCs continued to grow in medium that had been
refreshed. The MSCs were cultured to 80–90% confluence before
passaging. All experiments were performed using MSCs at passages
3–5 and the MSCs were identified by their immunophenotypic
characteristics. Briefly, the cells were harvested, washed with PBS
and labeled with the following conjugated antibodies: phycoerythrin
(PE)-labeled anti-CD45 (553091) and fluorescein isothiocyanate
(FITC)-labeled-CD29 (555005), CD44 (561859) and CD34 (560238; all
from BD Biosciences, Franklin Lakes, NJ, USA). The labeled cells
were detected by flow cytometry and analyzed using FACSDiva
software (Becton-Dickinson, San Jose, CA, USA).
Cell treatments
To induce the apoptosis of the BM-MSCs by subjecting
them to OGD conditions, the original medium was removed and
replaced with glucose- and serum-free medium. The MSCs were then
placed in an oxygen free incubator (855-AC; Plas-Labs Inc.,
Lansing, MI, USA) at 37°C, cultured for 0, 2, 4 and 8 h and then
harvested for the analysis of apoptosis. To determine the possible
mechanisms responsible for OGD-induced injury, the MSCs were
subjected to OGD conditions for 0, 1, 2, 4 and 8 h before being
harvesting for further experiments. To determine the optimal
concentration of ex-4 (ProSpec-Tany TechnoGene, Ltd., Ness-Ziona,
Israel), 10, 50 and 100 nM ex-4 were added to the complete medium
12 h before the cells were subjected to OGD conditions. Cell
viability was examined to determine the optimal concentration of
ex-4. To examine the anti-apoptotic effects of ex-4, the cells were
incubated with ex-4 for 12 h and then subjected to OGD conditions
for 4 h. To determine the role of GLP-1R, the cells were incubated
with the GLP-1R antagonist, exendin9-39 (ex9-39; 50 nM; Aladdin
Reagents (Shanghai) Co., Ltd., Shanghai, China), and ex-4 for 90
min and then subjected to OGD conditions. To determine the
involvement of protein kinase A (PKA) in the biological effects of
ex-4, the PKA inhibitor, H89 (10 µM; Sigma-Aldrich, St.
Louis, MO, USA), was added to complete medium with ex-4, 90 min
prior to the cells being subjected to OGD conditions. After being
subjected to the experimental treatments, the cells were harvested
for use in subsequent experiments.
Gene knockdown of activating
transcription factor 4 (ATF-4) and C/EBP homologous protein (CHOP)
by small interference RNA (siRNA)
The MSCs were seeded in 6-well plates and cultured
in serum-free medium for 24 h, and then transfected with siRNA for
rat ATF-4/CHOP and non-target siRNA (scramble siRNA) using
X-tremeGENE siRNA transfection reagent (Roche, Mannheim, Germany)
according to the manufacturer's instructions. The siRNA sequences
for ATF-4 and CHOP were 5′-AUCGAAGUCAAACUCUUUCAGGUCC-3′ and
antisense, 5′-GGACCUGAAAGAGUUUGACUUCGAU-3′; and
5′-GGAAGAACUAGGAAACGGA-3′ and antisense, 5′-UCCGUUUCCUAGUUCUUCC-3′,
respectively. The siRNAs were dissolved in diethylpryocarbonate
(DEPC)-treated water and diluted to 0.2 µM with 250
µl Opti-MEM (obtained from Invitrogen, Carlsbad, CA, USA)
for 10 min. A total of 10 µl of X-tremeGENE siRNA
transfection reagent (Roche) was also diluted with 250 µl
Opti-MEM for 20 min. The siRNA and transfection reagent were then
mixed (500 µl) and blended gently for 20 min and added to
the cell plate with 2 ml culture medium. The MSCs transfected with
the siRNAs were cultured for 48 h and then subjected to OGD
conditions as described above. The cells were harvested for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analysis to determine the silencing rate, and for use
in subsequent experiments.
MTT assay
The cells were seeded in 96-well plates at a
starting density of 1×104 cells/well. After being
subjected to the experimental treatments, the cells were washed and
incubated with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) solution (Sigma-Aldrich) at 37°C for 4 h. The
supernatant was removed, and 150 µl dimethyl sulfoxide
(DMSO) were added to each well. The absorbance (OD) of the reaction
solution was examined at 490 nm. Five separate experiments in
triplicate were conducted with different concentrations of
ex-4.
Detection of apoptosis by flow cytometric
analysis
Apoptosis was assessed by Annexin V-FITC/propidium
iodide (PI) (BD Biosciences) double staining and measured by
fluorescence-activated cell sorting (FACS). We obtained an Annexin
V/PI apoptosis analysis kit and performed the experiment according
to the manufacturer's instructions. Briefly, the cells were
collected and sorted into assay tubes at a density of
105–5×105 cells/tube. The cells were then
stained with Annexin V-FITC and PI for 15 min and detected by FACS
with FACSDiva Software (Becton-Dickinson)
Hoechst 33258 staining
The MSCs were seeded in 6-well plates. After being
subjected to the experimental treatments, the cells were washed
twice with PBS and incubated with Hoechst 33258 staining solution
(Beyotime Institute of Biotechnology, Haimen, China) for 30 min at
37°C. Subsequently, the staining solution was removed and the cells
were washed twice with cold PBS and images were captured using a
fluorescence microscope (DMI4000B; Leica, Wetzlar, Germany). The
cells with bright white fluorescent nuclei were considered to be
apoptotic cells and those with homogeneous blue fluorescence in the
nuclei were considered to be viable cells.
Intracellular cAMP ELISA assay
The MSCs were plated at a density of
5×104 cells in 48-well plates, and after being subjected
to the experimental treatments, the cells were collected and lysed
by freeze and thaw cycle 5 times. The intracellular cAMP
concentrations were measured using a rat cAMP ELISA kit (Jiang Lai
Biotechnology, Shanghai, China) according to the manufacturer's
instructions. The absorbance (OD) of the reaction solution was
detected using a microplate reader (BioTek Instruments, San Jose,
CA, USA) at 595 nm.
RT-qPCR
Total RNA was extracted from the MSCs using TRIzol
(Invitrogen). The concentration of the RNA was measured by
ultraviolet spectrophotometry and 1 µg RNA was reverse
transcribed into first-strand cDNA using a Transcriptor First
Strand cDNA Synthesis kit (Roche). Quantitative (real-time) (qPCR)
was performed using FastStart Universal SYBR-Green Master according
to the instructions provided by the manufacturer. The primer
sequences were as follows: GLP-1R forward, ACCTGTCGGAGTGCGAAGAGT
and reverse, ACAGTGCTCGGAGGATGAAGG; CHOP forward,
AGGTCCTGTCCTCAGATGAAAT and reverse, CAGGGTCAAGAGTAGTGAAGGTTT; ATF-4
forward, CCTTCG ACCAGTCGGGTTTG and reverse, CTGTCCCGGAAAAGGCATCC;
GAPDH forward, CATCAAGAAGGTGGTGAAGC and reverse,
ACCACCCTGTTGCTGTAG. The threshold cycle (Ct) value was detected
using an ABI Real Time PCR System (Applied Biosystems, Foster City,
CA, USA. The ΔCt value was calculated by subtracting the Ct number
of the target gene from that of the housekeeping gene, GAPDH. The
fold-change in the transcript level was calculated based on the
ΔΔCt method.
Western blot analysis
The MSCs subjected to different experimental
treatments were washed with PBS and lysed in RIPA lysis buffer
blended with protease and phosphatase inhibitors on ice for 30 min,
and subsequently shattered by ultrasound. The cell extracts were
centrifuged for 15 min at 12,000 × g and the supernatants were
collected. Total protein in the supernatant was quantified using a
Bradford protein assay kit (Beyotime Institute of Biotechnology).
Proteins (20–30 µg) were separated by SDS-PAGE on a 10%
polyacrylamide gel (15% polyacrylamide gel for caspase-3) and then
transferred onto methanol-activated PVDF membranes. The membranes
were blocked for 1 h in 5% skim milk diluted with TBS (50 mM Tris
and 150 mM NaCl) containing 0.1% Tween-20 (TBST) at 37°C and
incubated overnight at 4°C with the following primary antibodies:
phosphorylated (p-) protein kinase RNA-like endoplasmic reticulum
kinase (PERK; 3179), PERK (3192), binding immunoglobulin protein
(BIP; 3183), ATF-4 (11815), CHOP (2895), caspase-3 (9665) and Bcl-2
(2870; all from Cell Signaling Technology, Danvers, MA, USA) and
GLP-1R (ab39072; Abcam, Cambridge, UK). The membranes were washed 3
times with TBST and then incubated for 1 h with a secondary
antibody conjugated with horseradish peroxidase (HRP) (anti-rabbit;
sc-25778; Zhongshan Golden Bridge Biotechnology, Beijing, China).
The membranes were washed with TBST 3 times. The immune complexes
images were developed by ECL in the dark and images were captured
using Bio-Rad ChemiDoc XRS equipment, and the protein band density
was quantified and analyzed by Quantity One software (both from
Bio-Rad, Hercules, CA, USA).
Statistical analysis
Experimental values are expressed as the means ± SD,
and the difference between groups was analyzed by one-way ANOVA
with Tukey's and Newman Keuls post tests. Statistical analysis was
performed by SPSS 19.0 software (IBM Corp., Armonk, NY, USA).
P-values <0.05 were considered to indicate a statistically
significant difference.
Results
Identification and characterization of
BM-MSCs
BM-MSCs from passages 3–5 were collected for
immunophenotypic identification. The mesenchymal origin markers,
CD29 and CD44, were highly expressed, whereas the hematopoietic
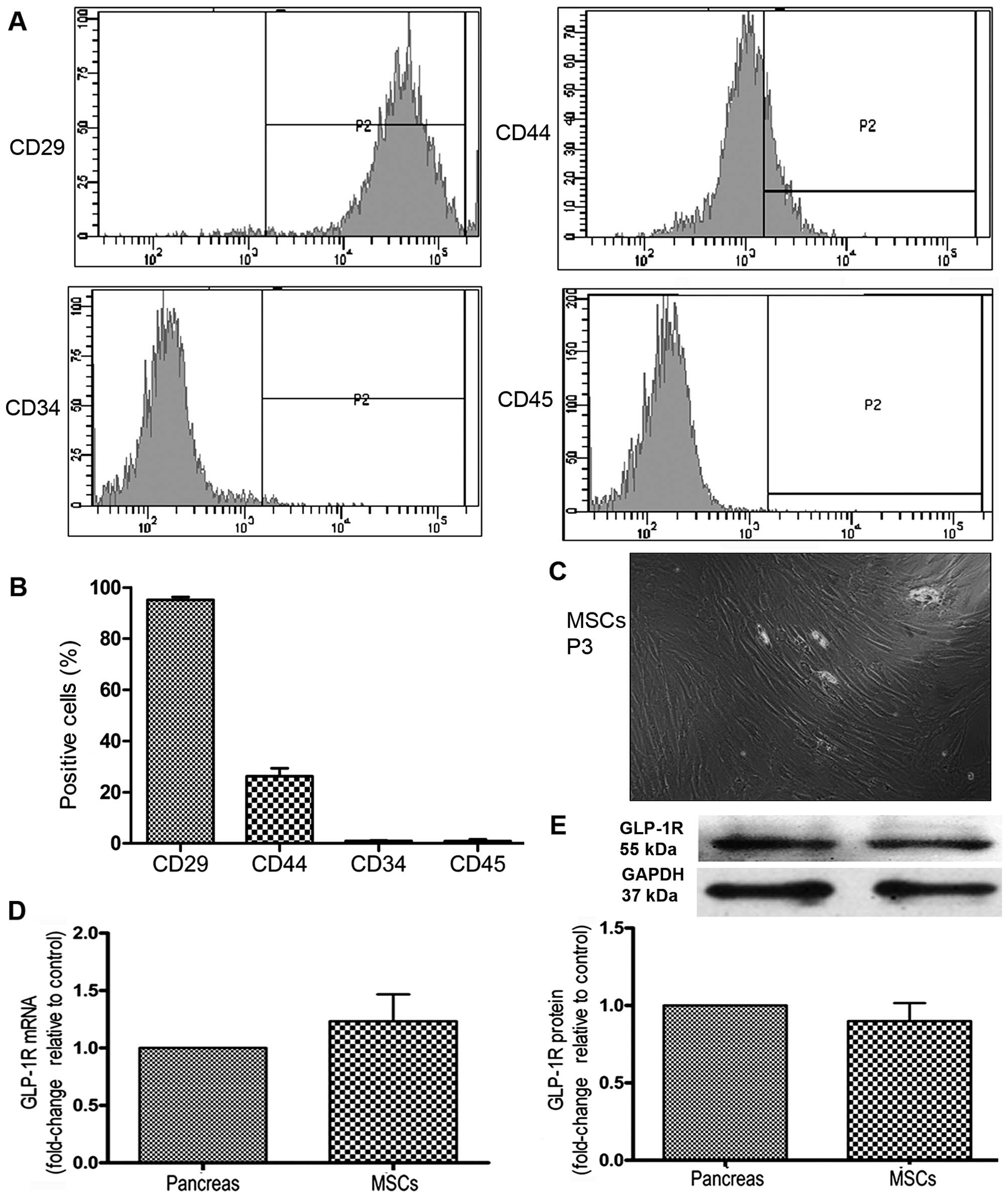
origin markers, CD45 and CD34, were minimally expressed (Fig. 1A and B). The MSCs were
spindle-shaped (Fig. 1C). To
examine the effects of the GLP-1R agonist, ex-4, we detected the
expression of its specific receptor, GLP-1R, on the MSCs using
RT-qPCR and western blot analysis. The results indicated that the
MSCs expressed GLP-1R at the mRNA and protein level (Fig. 1D and E).
OGD mimics in vitro ischemic conditions
and induces the apoptosis of MSCs
In the present study, the MSCs were subjected to OGD
conditions for 0 to 8 h in order to induce typical apoptosis. The
apoptotic ratio of the MSCs was detected using Annexin V/PI
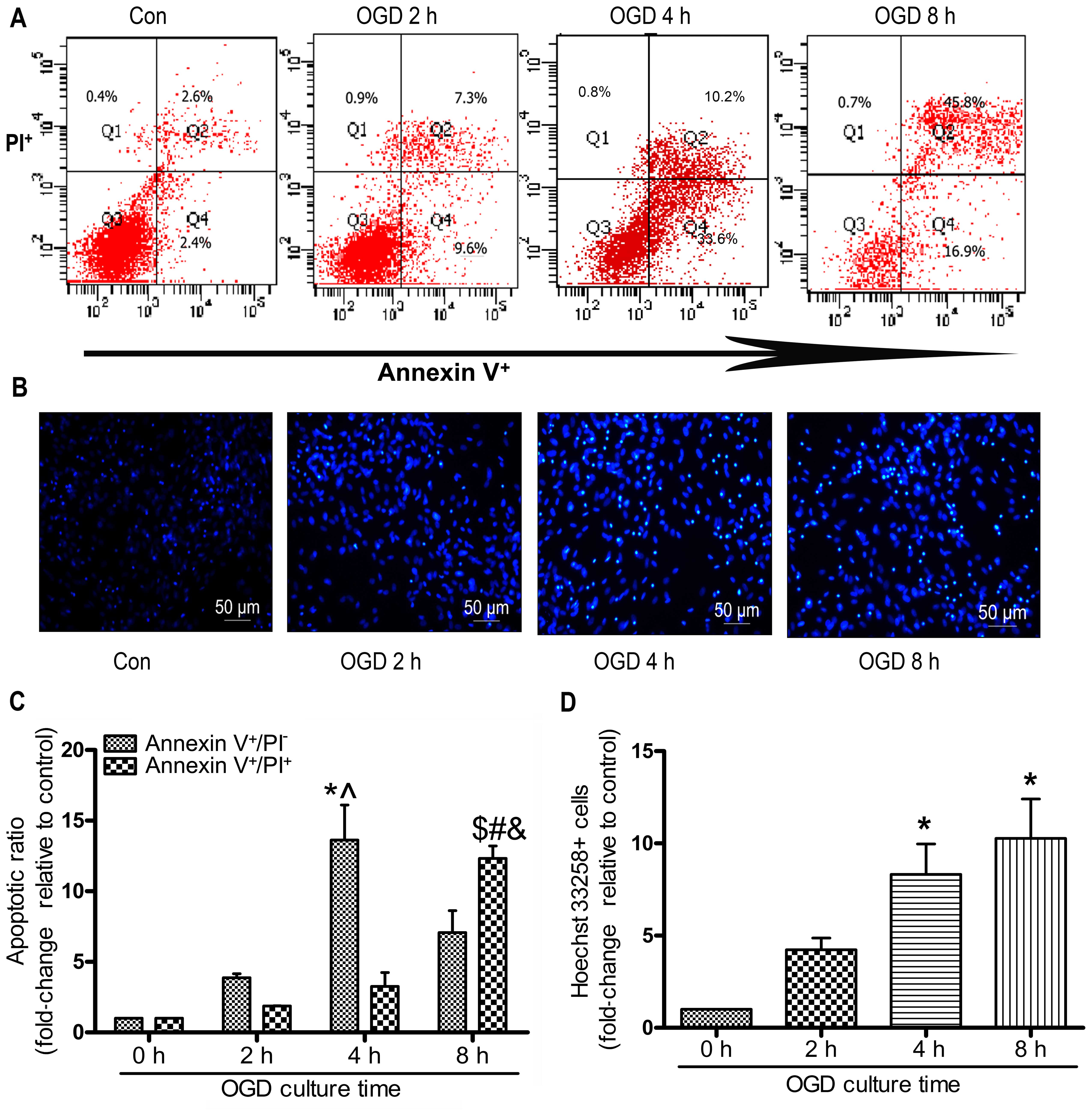
staining and flow cytometry. As shown in Fig. 2A and C, the early-stage apoptotic
ratio within 4 h increased in a time-dependent manner. Subjecting
the MSCs to OGD contitions for 4 h resulted in the most notable
early-stage apoptotic rate compared with 2 and 8 h of being
subjected to OGD contitions (4 h, 13.63±4.28 vs. 2 h 3.86±0.50; 8
h, 7.07±2.69, P<0.05; Fig.
2C). The late-stage apoptotic ratio increased gradually and
there was an evident increase in the cells exposed for 8 h to OGD
compared with those exposed for 4 or 2 h. The numbers of Hoechst
33258-positive stained cells at 4 and 8 h were significantly
increased compared with those at 0 h (4 h, 7.40±0.40; 8 h,
9.20±1.20 vs. 2 h 3.90±0.61; P<0.05; Fig. 2D). However, there was no
significant difference observed in the number of Hoechst
33258-positive cells following 4 and 8 h of incubation (P>0.05;
Fig. 2D).
Exposure to OGD in vitro triggers ER
stress in MSCs
PERK is a critical ER stress sensor, and its
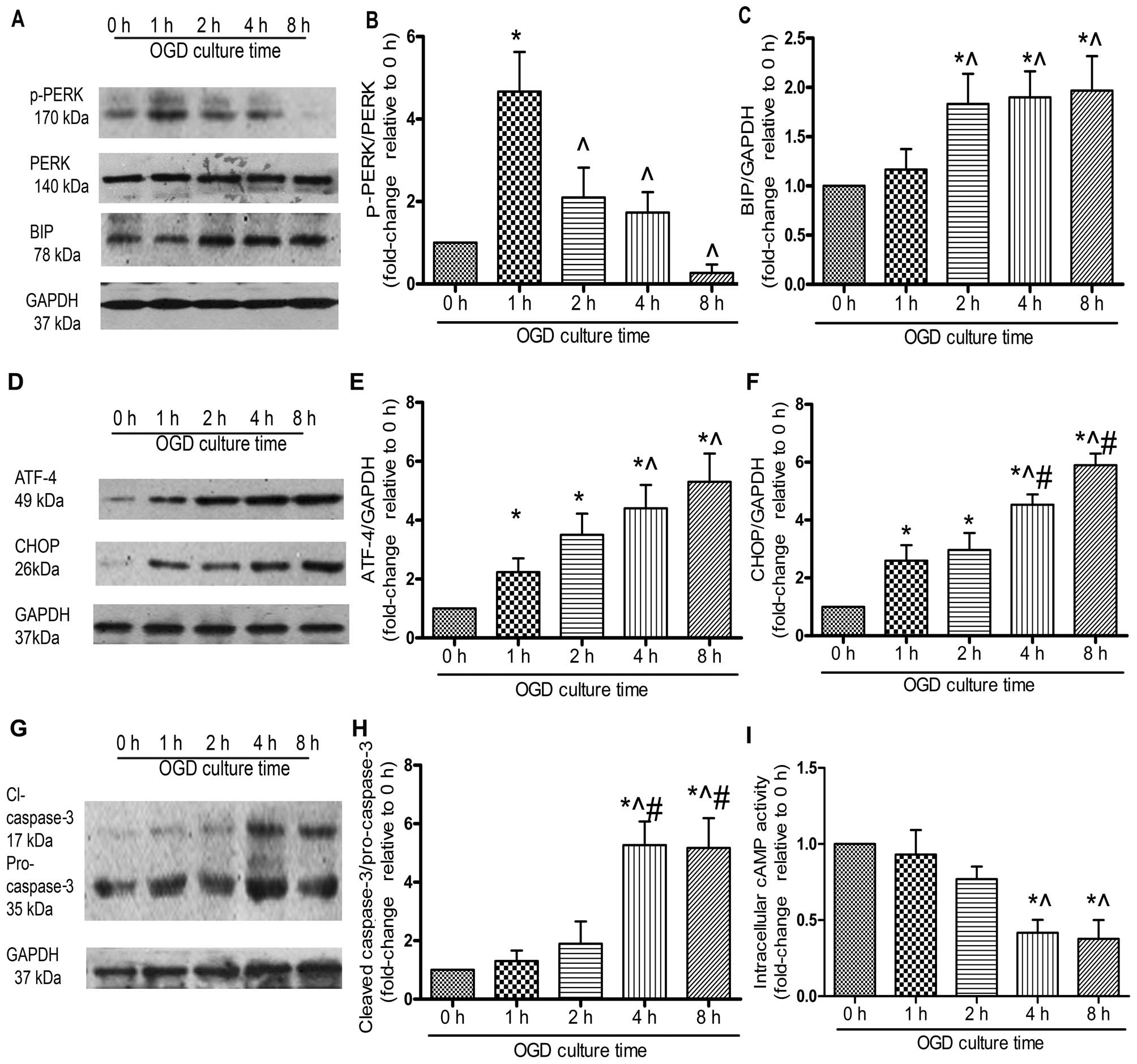
activation suggests the involvement of ER stress (21). In the present study, PERK was
phosphorylated at 1 h of OGD incubation and this lasted for 4 h
(Fig. 3A and B). The protein
level of BIP, an ER chaperone, increased significantly following
exposure to OGD for 2 h (P<0.05; Fig. 3A and C). ATF-4 protein expression
increased gradually. There was a significant elevation at 4 and 8 h
compared with that at 1 h (P<0.05; Fig. 3D and E), whereas no marked
difference was observed between 4 and 8 h (P>0.05). Similarly,
CHOP protein expression at 4 h was significantly increased compared
with that at 1 and 2 h, whereas no significant increase was
observed between 4 and 8 h (Fig. 3D
and F). The cleavage of caspase-3, a typical indicator of cell
apoptosis, was significantly upregulated under OGD conditions in a
time-dependent manner (Fig. 3G and
H). To determine the role of cAMP signaling in the OGD-induced
apoptosis of MSCs, we examined the activity of intracellular cAMP,
which was significantly decreased under OGD conditions. By 4 h, the
activity of intracellular cAMP had decreased by approximately 50%,
which suggested that cAMP signaling was abrogated during the
OGD-induced apoptosis of MSCs (Fig.
3I).
Ex-4 exerts anti-apoptotic effects on
MSCs under OGD conditions
In order to preliminarily evaluate the
cytoprotective effects, as well as the corresponding optimal
concentration of ex-4 in the MSCs under OGD conditions, an MTT
assay was performed on the MSCs pre-treated with 0, 10, 50 and 100
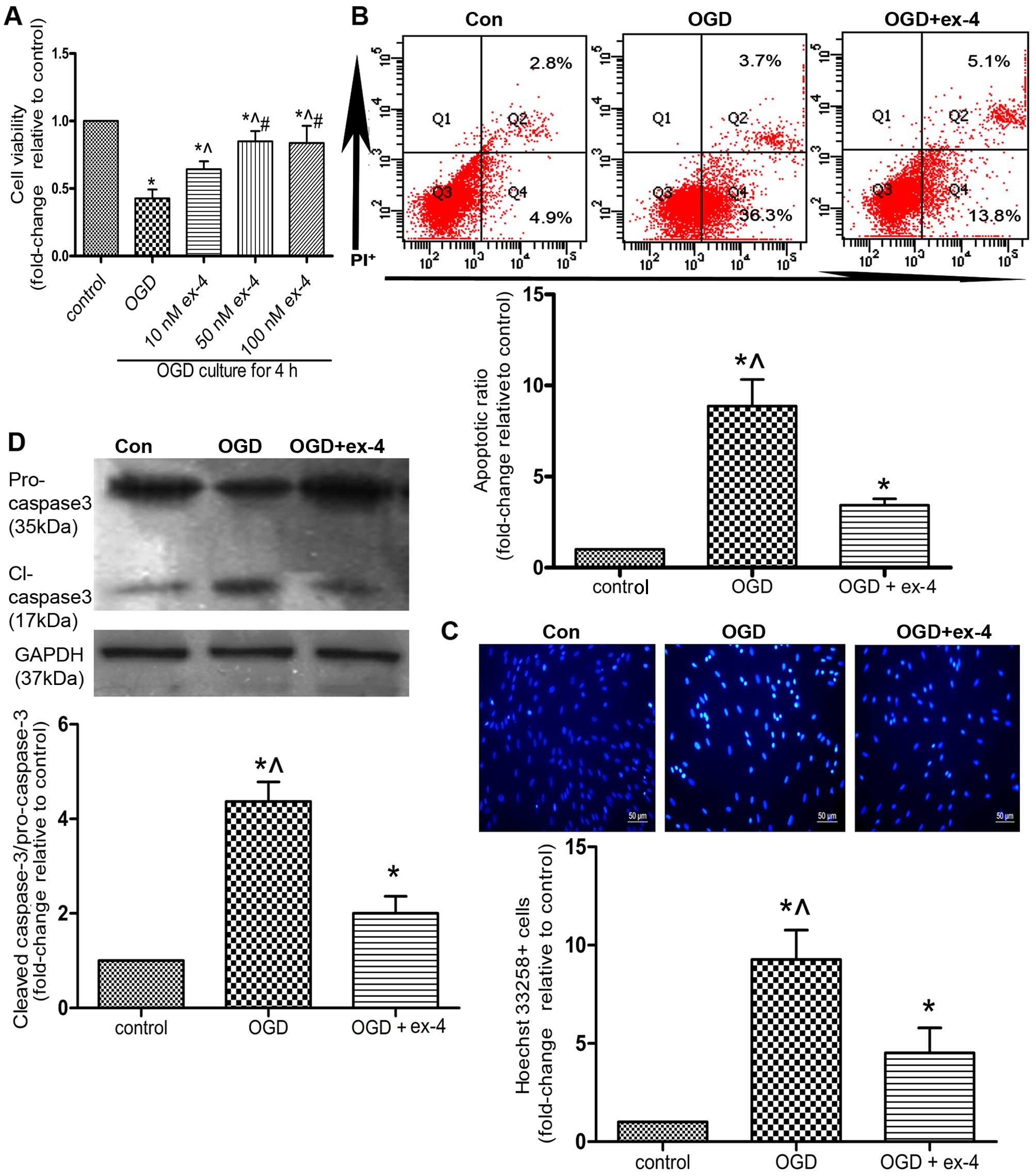
nM of ex-4. As shown in Fig. 4A,
pre-treatment of the MSCs with 50 nM ex-4 for 12 h led to the
restoration of cell viability by almost 50% compared with the OGD
group (cells not treated with ex-4; 0.85±0.04 vs. 0.43±0.04,
P<0.05). However, there was no significant difference observed
between the 50 nM ex-4- and 100 nM ex-4-treated groups (P>0.05;
Fig. 4A). Thus, for subsequent
experiments, the MSCs were pre-treated with 50 nM ex-4 in order to
evaluate its cytoprotective effects.
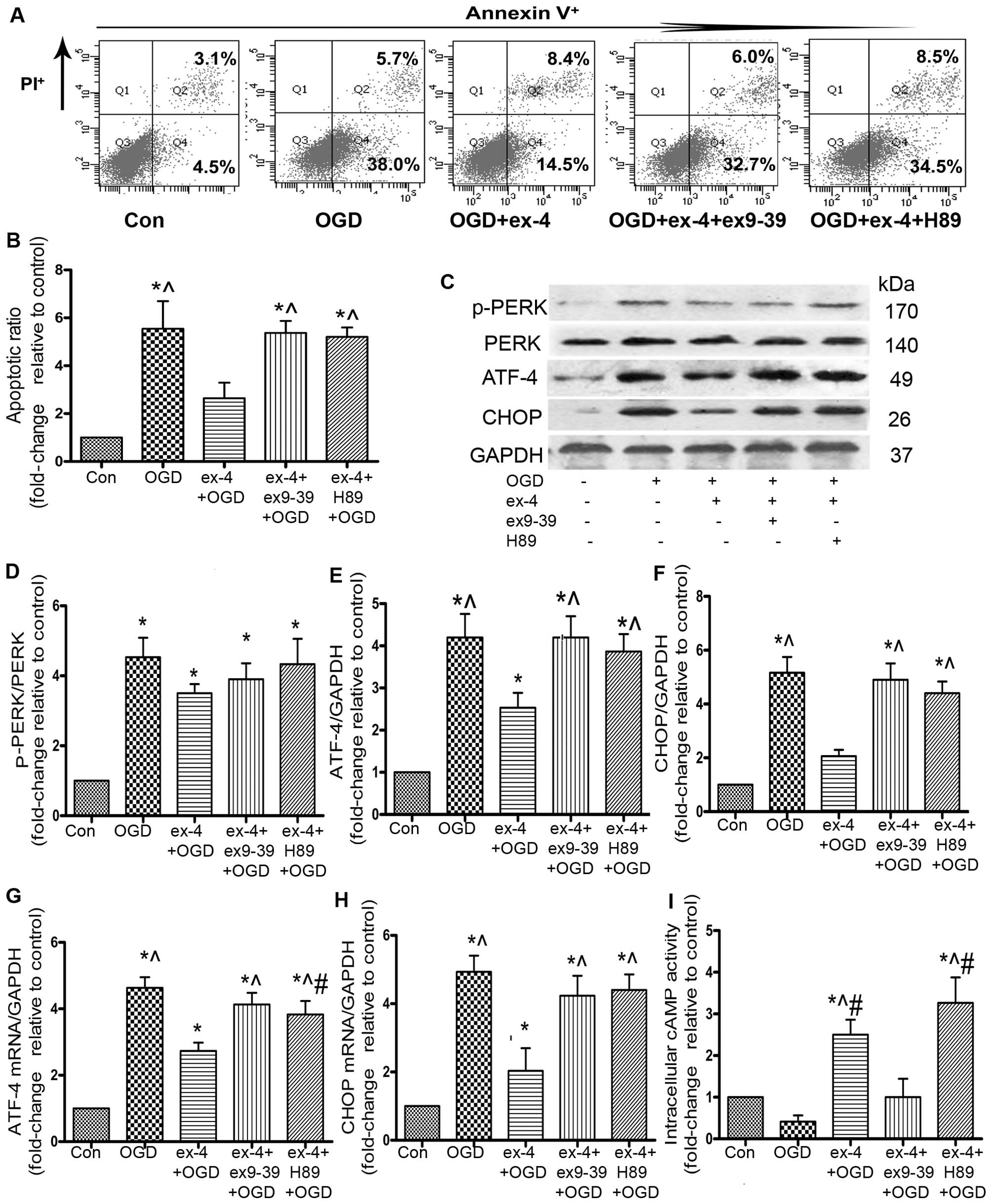
The apoptotic ratio of the MSCs was detected by
Annexin V/PI staining and flow cytometry. As indicated in Fig 4B, pre-treatment with ex-4 reduced
the apoptotic ratio by 61.3% compared with that in the OGD group
(3.43±0.35 vs. 8.87±1.46, P<0.05). In a parallel experiment, the
apoptosis of the MSCs was assessed by Hoechst 33258 staining. The
number of Hoechst 33258-positive cells was also reduced by 51.6% in
the OGD + ex-4 group compared with that in the OGD group (4.52±1.28
vs. 9.27±1.50, P<0.05; Fig.
4C). Moreover, it was demonstrated that the levels of cleaved
caspase-3 were markedly reduced in the OGD + ex-4 group compared
with those in the OGD group (2.00±0.36 vs. 4.37±0.42, P<0.05;
Fig. 4D). Taken together, these
findings indicated that ex-4 exerted anti-apoptotic effects on the
MSCs under OGD conditions.
Ex-4 protects MSCs from OGD-induced
apoptosis by decreasing the ATF-4 and CHOP levels and by activating
the GLP-1R/cAMP/PKA pathway
It has been demonstrated that ATF-4 and CHOP are
critical factors that contribute to cell apoptosis induced by ER
stress (22). In the present
study, the ATF-4 mRNA levels decreased by 41.3% (2.73±0.25 vs.
4.63±0.32, P<0.05; Fig. 5G)
and the CHOP mRNA levels decreased by 59.2% in the ex-4 + OGD group
compared with the OGD group (2.03±0.67 vs. 4.93±0.47, P<0.05;
Fig. 5H). To further confirm the
inhibition of the ATF-4/CHOP pathway, the ATF-4 and CHOP protein
levels were examined by western blot analysis; ex-4 significantly
reduced the protein expression of ATF-4 by 39.8% compared with the
OGD group (2.53±0.61 vs. 4.20±0.96, P<0.05; Fig. 5E), and CHOP protein expression
decreased by 60% compared with the OGD group (2.07±0.40 vs.
5.17±1.00, P<0.05; Fig. 5F).
There was no significant difference observed in the p-PERK protein
levels between the ex-4 + OGD and the OGD groups (P>0.05;
Fig. 5D).
To examine the role of GLP-1R in the MSCs
pre-treated with ex-4, the MSCs were incubated with the GLP-1R
antagonist, ex9-39, as well as ex-4, prior to incubation under OGD
conditions. The apoptotic rate was much higher in the ex-4 + ex9-39
+ OGD group than that in the ex-4 + OGD group (5.37±0.50 vs.
2.64±0.65, P<0.05; Fig. 5A and
B). Following incubation with ex9-39, the abrogation of ATF-4
(2.53±0.61 vs. 4.20±0.87, P<0.05; Fig. 5E) and CHOP protein expression was
inhibited (2.07±0.40 vs. 4.90±1.06, P<0.05; Fig. 5F). The mRNA expression of ATF-4
and CHOP was also significantly higher in the ex-4 + ex9-39 + OGD
group than that in the ex-4 + OGD group (P<0.05; Fig. 5G and H).
The cAMP-dependent PKA pathway has been found to
play a role in cell survival (23). Thus, to examine the involvement of
the cAMP/PKA pathway in the anti-apoptotic effects of ex-4, we
conducted an intracellular cAMP activity ELISA, which showed that
the cAMP activity in the ex-4 + OGD group was markedly increased
compared with that in the OGD and ex9-39 + ex-4 + OGD group (ex-4
2.50±0.36 vs. OGD 0.42±0.18, ex9-39 1.00±0.44, P<0.05; Fig. 5I), which suggested that ex-4
enhanced the activity of cAMP. In order to examine the role of PKA
in the cytoprotective effects of ex-4, the MSCs were pre-incubated
with H89. FACS analysis revealed that the apoptotic ratio in the
ex-4 + H89 + OGD group was significantly increased compared with
that in the ex-4 + OGD group (ex-4, 2.64±0.65 vs. H89, 5.20±0.39,
P<0.05; Fig. 5B), which
suggests that PKA plays a major role in the anti-apoptotic effects
of ex-4. As regards the role of PKA in the inhibitory effects of
ex-4 on ATF-4/CHOP, the results revealed that there was no marked
difference between the OGD and the ex-4 + H89 + OGD groups in the
ATF-4/CHOP protein levels, as well as in the CHOP mRNA level
(P>0.05; Fig. 5E, F and H),
whereas there were prominent differences in the mRNA and protein
levels of ATF-4 and CHOP between the ex-4 + OGD group and the ex-4
+ H89 + OGD group (P<0.05; Fig.
5E–H). Taken together, these findings suggest that the
activation of PKA may participate in the abrogation of ATF-4/CHOP
levels by ex-4.
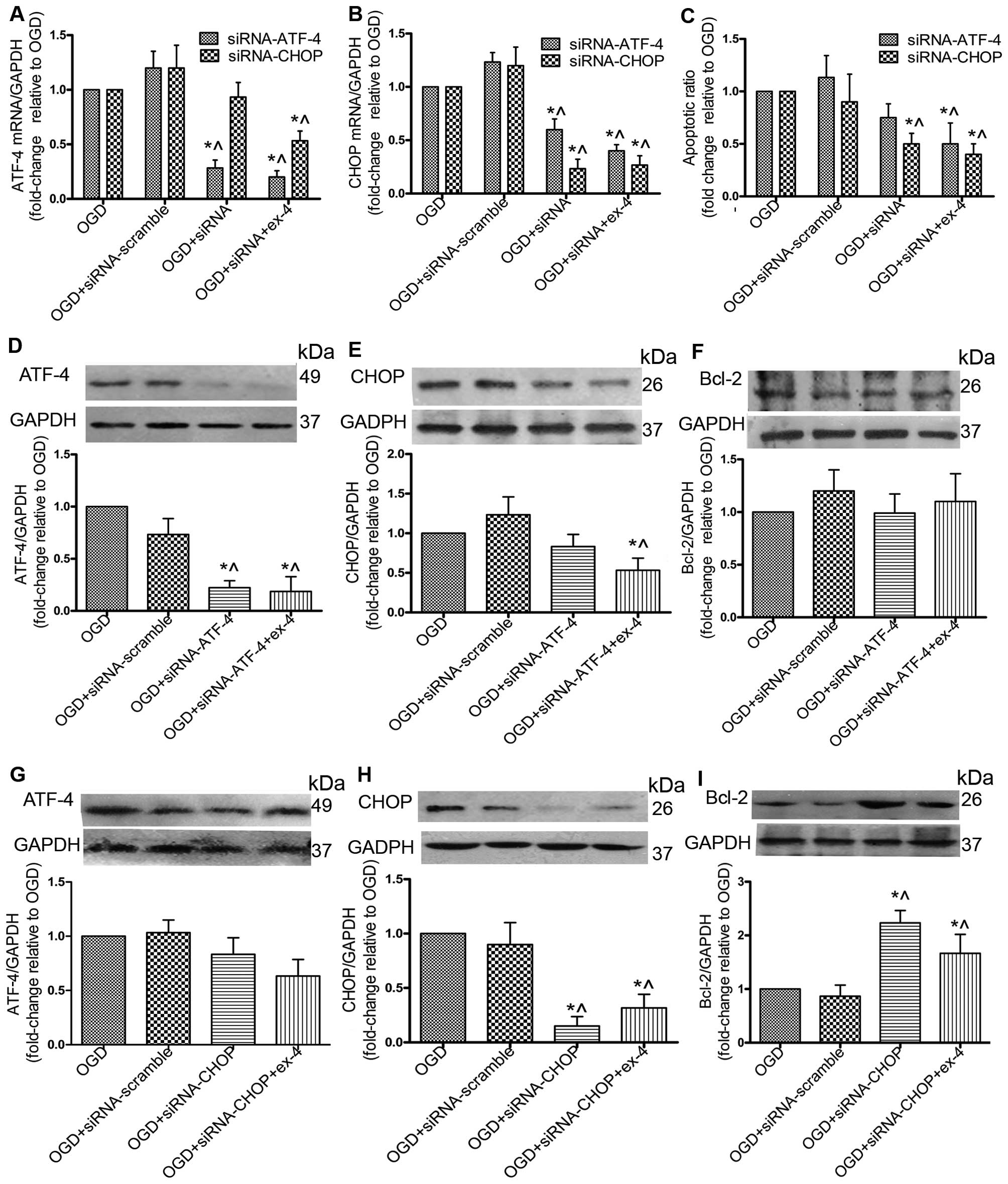
siRNA against CHOP (siRNA-CHOP) protects
MSCs by upregulating Bcl-2 and does not impair the cytoprotective
effects of ex-4
To further explore the role of the ATF-4/CHOP
pathway in the anti-apoptotic effects mediated by ex-4, the siRNA
knockdown technique was adopted to reduce the expression of ATF-4
and CHOP. By silencing ATF-4, the mRNA expression of ATF-4 was
reduced by 72% (P<0.05; Fig.
6A), and the protein expression was reduced by 73% (P<0.05;
Fig. 6D); however, no significant
difference in the apoptotic ratio and Bcl-2 protein expression was
observed between the OGD + siRNA-ATF-4 group and the OGD group
(P>0.05; Fig. 6C and F).
Treatment with Ex-4 combined with siRNA-ATF-4 significantly
decreased the apoptotic ratio (Fig.
6C) and the protein expression of CHOP (Fig. 6E). By silencing CHOP, its protein
expression was reduced by 87% (Fig.
6H) and the apoptotic ratio was decreased by 50% (Fig. 6C). There was no significant change
in the ATF-4 protein levels (Fig.
6G); however, Bcl-2 protein expression was increased 1.23-fold
(Fig. 6I). With ex-4 + siRNA-CHOP
treatment, the apoptotic ratio was decreased (P<0.05; Fig. 6C), and in addition, the Bcl-2
protein levels were significantly upregulated compared with the OGD
group (P<0.05; Fig. 6I). These
results suggested that transfection of the cells with siRNA-CHOP
reduced the apoptotic rate in the MSCs without impairing the
anti-apoptotic effects of ex-4.
Discussion
BM-MSCs originate from the mesodermal germ layer and
perform a supportive function in the stroma, as well as giving rise
to cells of multiple cell lineages, including adipocytes and
osteocytes. Evidence suggests that MSCs possess the ability to
differentiate into cardiomyocytes and to secrete a wide array of
cytokines and growth factors to suppress the inflammatory response,
inhibit fibrosis and enhance angiogenesis in the infarcted
myocardium (24). These findings
provide a solid theoretical basis for the application of MSC
engraftment as a therapy in AMI; however, MSCs do not in fact
survive for long following transplantation, which may explain the
negligible effect on cardiac function one year after MSC
engraftment (25). Based on
findings from a bioluminescence imaging study by van der Bogt et
al, all engrafted MSCs had died by week 6 (26). In addition, Mangi et al
observed robust cell death early following transplantation after
transfecting MSCs with the pro-survival gene, Akt-1 (8). The ischemic microenvironment in the
infarcted heart proves hostile to engrafted MSCs, resulting in
decreased cell survival. To mimic the ischemic milieu in
vitro, hypoxia and serum deprivation (H/SD) is a commonly used
model, and our group previously found that the H/SD model induced
the apoptosis of approximately 25% of MSCs for up to 24 h (7). As glucose is an indispensible energy
nutrient for cells and is found in short supply in the ischemic
context, the OGD model was adopted in the present study and
exposure to OGD induced the apoptosis of approximately 40% of the
MSCs within 4 h. In comparison, the OGD model represents a more
time-saving and aggressive approach to mediate MSC death in
vitro.
To date, studies aiming to find ways to protect MSCs
from apoptosis have focused on maintaining the integrity of the
mitochondria or activating survival signaling pathways (7). Few studies have highlighted the
importance of ER stress induced by ischemia (27). As a matter of fact, ER stress
plays a pivotal role in the pathophysiological mechanisms of AMI
and ischemia-induced apoptosis (6). The ER is an organelle involved in
protein folding, calcium homeostasis and lipid biosynthesis.
Various factors that interfere with ER function lead to the
accumulation of misfolded or unfolded proteins, including oxidative
stress, ischemia and disturbances in calcium homeostasis. Okada
et al demonstrated that ER stress-initiated apoptosis
occurred in cardiomocytes in a mouse model of aortic constriction,
and found that suppressing the ER stress pathway may diminish the
death of cardiomocytes (28).
Additionally, it was suggested to be the main mechanism responsible
for the apoptosis of various types of cells in models of ischemia
in vivo or in vitro (29,30). In line with these results, the
present study demonstrated that the levels of p-PERK/BIP/ATF-4/CHOP
markedly increased and those of the apoptotic indicator, cleaved
caspase-3, were upregulated, which strongly suggested that
excessive and prolonged ER stress is an important mechanism
responsible for the apoptosis of MSCs in the setting of ischemia.
ER stress serves as a predominant mechanism and a therapeutic
target with which to improve the survival of both cardiomocytes and
MSCs in AMI.
Ex-4, a long-acting agonist of GLP-1, is known to
regulate the perturbation of ER stress. As a GLP-1 agonist, ex-4
depends on GLP-1R to play its physiological role, and this study
demonstrated that GLP-1R was expressed on rat MSCs and played a key
role in mediating the anti-apoptotic effects of ex-4. Tsunekawa
et al (31) observed that
ex-4 distinctly decreased the levels of the ER stress-related
molecules, BIP and CHOP, in β-cell-specific
calmodulin-overexpressing mice and reduced the apoptosis of
β-cells. In agreement with these findings, the present study found
that ex-4 markedly decreased the apoptosis of MSCs by inhibiting
the activation of the ATF-4/CHOP pathway. Furthermore, in a rat
model of myocardial infarction, pre-conditioning with ex-4 was
shown to improve the adhesion and therapeutic efficacy of
transplanted adipose-derived stem cell transplantation therapy
(32). In a previous study, the
post-MI delivery of encapsulated GLP-1 MSCs that had been
genetically modified increased angiogenesis and attenuated fibrosis
than treatment with MSCs alone (33). Moreover, ex-4 aids in the survival
of cardiomyocytes and endothelial cells, and the pro-inflammatory
factors in the infacted myocardium were significantly decreased by
ex-4 infiltration (15,16,34). Taken together, the findings from
previous studies and the present study indicate that ex-4 is a
promising candidate for optimizing MSC therapy for AMI.
The ATF-4/CHOP pathway is an important branch of ER
stress, and the present study demonstrated that ex-4 reduced the
OGD-induced apoptosis of MSCs by suppressing the ATF-4/CHOP
pathway. CHOP serves as a pivotal stimulus for cell death, and it
is mainly induced by ATF-4 (35).
The present study found that CHOP-siRNA significantly decreased the
apoptosis of MSCs under OGD conditions. In line with these results,
a previous study by Zinszner et al (36) demonstrated that embryonic
fibroblasts derived from CHOP−/− mice exhibited
significantly less apoptosis. CHOP mediated the transcription of
various types of apoptotic-related molecules, including Bcl-2,
GADD34 and ER oxidoreductin 1 (ERO1) (37). Bcl-2 promotes cell survival by
restoring the mitochondrial potential, and the present study
indicated that the Bcl-2 protein levels were significantly
increased following transfection with siRNA-CHOP and ex-4
preconditioning. This implied that Bcl-2 participated in the
anti-apoptotic effects of ex-4. Moreover, CHOP activated GADD34 to
initiate or enhance apoptosis by increasing protein synthesis,
which resulted in the aggravation of ER stress. By activating ERO1,
CHOP promoted the oxidizing state in the ER and therefore increased
the aggregation of hostile elements leading to cell death (37). Thus, the downregulation of CHOP in
MSCs may be the principal mechanism responsible for the
cytoprotective effects of ex-4.
The cAMP-dependent PKA pathway plays a role in the
promotion of cell proliferation and in the inhibition of apoptosis
(23). The signal transduction to
ER by ex-4 has been reported to be associated with the cAMP/PKA
pathway in β-cells (38). In the
present study, the activity of intracellular cAMP was impaired by
OGD, and restored by ex-4. The protective effects of ex-4 were
notably reversed by H89, whereas the attenuation of ATF-4 and CHOP
levels by ex-4 was partly restored. Thus, this study strongly
suggested that the cAMP-dependent PKA pathway may also participate
in the anti-apoptotic effects of ex-4 under OGD conditions.
In conclusion, the present study demonstrates that
ex-4 confers resistance to OGD-mediated apoptosis in BM-MSCs, and
the possible mechanisms responsible for these effects involve the
activation of the GLP-1R/cAMP/PKA pathway and the attenuation of ER
stress. These findings highlight ER stress as a pivotal target for
protecting MSCs from ischemia and provide evidence of the
cytoprotective effects of ex-4. Together with the reported
protective effects against ischemia or ischemia-reperfusion injury
in cardiomyocytes, ex-4 may represent a therapeutic agent with the
potential to optimize MSC therapy in AMI.
Acknowledgments
We would like to thank Dr Hulun Li for his guidance
with cell culture and treatment, and Dr Wei Liu for her expert
assistance with PCR and western blot analysis. Dr Hulun Li and Dr
Wei Liu are members of the Key Laboratory of Myocardial Ischemia
Mechanism and Treatment (Harbin Medical University), Ministry of
Education. The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81330033
and 81300201) and a grant from the Key Laboratory of Myocardial
Ischemia, Harbin Medical University, Ministry of Education (no.
KF201408).
References
|
1
|
Boersma E, Mercado N, Poldermans D,
Gardien M, Vos J and Simoons ML: Acute myocardial infarction.
361:847–858. 2003.
|
|
2
|
Uccelli A, Moretta L and Pistoia V:
Mesenchymal stem cells in health and disease. Nat Rev Immunol.
8:726–736. 2008. View
Article : Google Scholar
|
|
3
|
Gnecchi M, He H, Liang OD, Melo LG,
Morello F, Mu H, Noiseux N, Zhang L, Pratt RE, Ingwall JS and Dzau
VJ: Paracrine action accounts for marked protection of ischemic
heart by Akt-modified mesenchymal stem cells. Nat Med. 11:367–368.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dai W, Hale SL, Martin BJ, Kuang JQ, Dow
JS, Wold LE and Kloner RA: Allogeneic mesenchymal stem cell
transplantation in postinfarcted rat myocardium: short- and
long-term effects. Circulation. 112:214–223. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Toma C, Pittenger MF, Cahill KS, Byrne BJ
and Kessler PD: Human mesenchymal stem cells differentiate to a
cardiomyocyte phenotype in the adult murine heart. Circulation.
105:93–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Minamino T and Kitakaze M: ER stress in
cardiovascular disease. J Mol Cell Cardiol. 48:1105–1110. 2010.
View Article : Google Scholar
|
|
7
|
Hou M, Liu J, Liu F, Liu K and Yu B: C1q
tumor necrosis factor-related protein-3 protects mesenchymal stem
cells against hypoxia- and serum deprivation-induced apoptosis
through the phosphoinositide 3-kinase/Akt pathway. Int J Mol Med.
33:97–104. 2014.
|
|
8
|
Mangi AA, Noiseux N, Kong D, He H, Rezvani
M, Ingwall JS and Dzau VJ: Mesenchymal stem cells modified with Akt
prevent remodeling and restore performance of infarcted hearts. Nat
Med. 9:1195–1201. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hou M, Cui J, Liu J, Liu F, Jiang R, Liu
K, Wang Y, Yin L, Liu W and Yu B: Angiopoietin-like 4 confers
resistance to hypoxia/serum deprivation-induced apoptosis through
PI3K/Akt and ERK1/2 signaling pathways in mesenchymal stem cells.
PLoS One. 9:e858082014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Willms B, Werner J, Holst JJ, Orskov C,
Creutzfeldt W and Nauck MA: Gastric emptying, glucose responses,
and insulin secretion after a liquid test meal: effects of
exogenous glucagon-like peptide-1 (GLP-1)-(7-36) amide in type 2
(noninsulin-dependent) diabetic patients. J Clin Endocrinol Metab.
81:327–332. 1996.PubMed/NCBI
|
|
11
|
Xu G, Stoffers DA, Habener JF and
Bonner-Weir S: Exendin-4 stimulates both beta-cell replication and
neogenesis, resulting in increased beta-cell mass and improved
glucose tolerance in diabetic rats. Diabetes. 48:2270–2276. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
DeFronzo RA, Ratner RE, Han J, Kim DD,
Fineman MS and Baron AD: Effects of exenatide (exendin-4) on
glycemic control and weight over 30 weeks in metformin-treated
patients withtype 2 diabetes. Diabetes Care. 28:1092–1100. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sonne DP, Engstrøm T and Treiman M:
Protective effects of GLP-1 analogues exendin-4 and GLP-1(9-36)
amide against ischemia-reperfusion injury in rat heart. Regul Pept.
146:243–249. 2008. View Article : Google Scholar
|
|
14
|
Lee CH, Yan B, Yoo KY, Choi JH, Kwon SH,
Her S, Sohn Y, Hwang IK, Cho JH, Kim YM and Won MH:
Ischemia-induced changes in glucagon-like peptide-1 receptor and
neuroprotective effect of its agonist, exendin-4, in experimental
transient cerebral ischemia. J Neurosci Res. 89:1103–1113. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arakawa M, Mita T, Azuma K, Ebato C, Goto
H, Nomiyama T, Fujitani Y, Hirose T, Kawamori R and Watada H:
Inhibition of monocyte adhesion to endothelial cells and
attenuation of atherosclerotic lesion by a glucagon-like peptide-1
receptor agonist, exendin-4. Diabetes. 59:1030–1037. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Younce CW, Burmeister MA and Ayala JE:
Exendin-4 attenuates high glucose-induced cardiomyocyte apoptosis
via inhibition of endoplasmic reticulum stress and activation of
SERCA2a. Am J Physiol Cell Physiol. 304:C508–C518. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marzioni M, Alpini G, Saccomanno S,
Candelaresi C, Venter J, Rychlicki C, Fava G, Francis H, Trozzi L
and Benedetti A: Exendin-4, a glucagon-like peptide 1 receptor
agonist, protects cholangiocytes from apoptosis. Gut. 58:990–997.
2009. View Article : Google Scholar :
|
|
18
|
Ferdaoussi M, Abdelli S, Yang J-Y, Cornu
M, Niederhauser G, Favre D, Widmann C, Regazzi R, Thorens B, Waeber
G and Abderrahmani A: Exendin-4 protects β-cells from interleukin-1
β-induced apoptosis by interfering with the c-Jun NH2-terminal
kinase pathway. Diabetes. 57:1205–1215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee J, Hong SW, Park SE, Rhee EJ, Park CY,
Oh KW, Park SW and Lee WY: Exendin-4 attenuates endoplasmic
reticulum stress through a SIRT1-dependent mechanism. Cell Stress
Chaperones. 19:649–656. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Clark JD, Gebhart GF, Gonder JC, Keeling
ME and Kohn DF: The 1996 guide for the care and use of laboratory
animals. ILAR J. 38:41–48. 1997. View Article : Google Scholar
|
|
21
|
Walter P and Ron D: The unfolded protein
response: from stress pathway to homeostatic regulation. Science.
334:1081–1086. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao J, Dai D-L, Yao L, Yu H-H, Ning B,
Zhang Q, Chen J, Cheng WH, Shen W and Yang ZX: Saturated fatty acid
induction of endoplasmic reticulum stress and apoptosis in human
liver cells via the PERK/ATF4/CHOP signaling pathway. Mol Cell
Biochem. 364:115–129. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Drucker DJ: Glucagon-like peptides:
regulators of cell proliferation, differentiation, and apoptosis.
Mol Endocrinol. 17:161–171. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Parekkadan B and Milwid JM: Mesenchymal
stem cells as therapeutics. Annu Rev Biomed Eng. 12:87–117. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lipinski MJ, Biondi-Zoccai GG, Abbate A,
Khianey R, Sheiban I, Bartunek J, Vanderheyden M, Kim HS, Kang HJ,
Strauer BE and Vetrovec GW: Impact of intracoronary cell therapy on
left ventricular function in the setting of acute myocardial
infarction: a collaborative systematic review and meta-analysis of
controlled clinical trials. J Am Coll Cardiol. 50:1761–1767. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
van der Bogt KE, Schrepfer S, Yu J, Sheikh
AY, Hoyt G, Govaert JA, Velotta JB, Contag CH, Robbins RC and Wu
JC: Comparison of transplantation of adipose tissue- and bone
marrow-derived mesenchymal stem cells in the infarcted heart.
Transplantation. 87:642–652. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Szegezdi E, Duffy A, O'Mahoney ME, Logue
SE, Mylotte LA, O'brien T and Samali A: ER stress contributes to
ischemia-induced cardiomyocyte apoptosis. Biochem Biophys Res
Commun. 349:1406–1411. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Okada K, Minamino T, Tsukamoto Y, Liao Y,
Tsukamoto O, Takashima S, Hirata A, Fujita M, Nagamachi Y, Nakatani
T, et al: Prolonged endoplasmic reticulum stress in hypertrophic
and failing heart after aortic constriction: possible contribution
of endoplasmic reticulum stress to cardiac myocyte apoptosis.
Circulation. 110:705–712. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hillion JA, Takahashi K, Maric D, Ruetzler
C, Barker JL and Hallenbeck JM: Development of an ischemic
tolerance model in a PC12 cell line. J Cereb Blood Flow Metab.
25:154–162. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Badiola N, Penas C, Miñano-Molina A,
Barneda-Zahonero B, Fadó R, Sánchez-Opazo G, Comella JX, Sabriá J,
Zhu C, Blomgren K, et al: Induction of ER stress in response to
oxygen-glucose deprivation of cortical cultures involves the
activation of the PERK and IRE-1 pathways and of caspase-12. Cell
Death Dis. 2:e1492011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsunekawa S, Yamamoto N, Tsukamoto K, Itoh
Y, Kaneko Y, Kimura T, Ariyoshi Y, Miura Y, Oiso Y and Niki I:
Protection of pancreatic beta-cells by exendin-4 may involve the
reduction of endoplasmic reticulum stress; in vivo and in vitro
studies. J Endocrinol. 193:65–74. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu J, Wang H, Wang Y, Yin Y, Wang L, Liu
Z, Yang J, Chen Y and Wang C: Exendin-4 pretreated adipose derived
stem cells are resistant to oxidative stress and improve cardiac
performance via enhanced adhesion in the infarcted heart. PLoS One.
9:e997562014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wright EJ, Farrell KA, Malik N, Kassem M,
Lewis AL, Wallrapp C and Holt CM: Encapsulated glucagon-like
peptide-1-producing mesenchymal stem cells have a beneficial effect
on failing pig hearts. Stem Cells Transl Med. 1:759–769. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Favaro E, Granata R, Miceli I, Baragli A,
Settanni F, Cavallo Perin P, Ghigo E, Camussi G and Zanone MM: The
ghrelin gene products and exendin-4 promote survival of human
pancreatic islet endothelial cells inhyperglycaemic conditions,
through phosphoinositide 3-kinase/Akt, extracellular signal-related
kinase (ERK)1/2 and cAMP/protein kinase A (PKA) signalling
pathways. Diabetologia. 55:1058–1070. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar
|
|
36
|
Zinszner H, Kuroda M, Wang X, Batchvarova
N, Lightfoot RT, Remotti H, Stevens JL and Ron D: CHOP is
implicated in programmed cell death in response to impaired
function of the endoplasmic reticulum. Genes Dev. 12:982–995. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Marciniak SJ, Yun CY, Oyadomari S, Novoa
I, Zhang Y, Jungreis R, Nagata K, Harding HP and Ron D: CHOP
induces death by promoting protein synthesis and oxidation in the
stressed endoplasmic reticulum. Genes Dev. 18:3066–3077. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cunha DA, Ladrière L, Ortis F,
Igoillo-Esteve M, Gurzov EN, Lupi R, Marchetti P, Eizirik DL and
Cnop M: Glucagon-like peptide-1 agonists protect pancreatic β-cells
from lipotoxic endoplasmic reticulum stress through upregulation of
BiP and JunB. Diabetes. 58:2851–2862. 2009. View Article : Google Scholar : PubMed/NCBI
|