Introduction
Estrogens are hormones that are important for sexual
and reproductive development, mainly in women. Estrogen is
instrumental in bone formation, and together with vitamin D,
calcium and other hormones, it effectively breaks down and rebuilds
bones according to the body's natural processes. As estrogen levels
begin to decline in middled-aged women, the process of and the
ability to rebuild bone dimishes. Consequently, in postmenopausal
women, the amount of bone being broken down excedes that of bone
being rebuilt (1).
Estrogen deficiency, which is vital in the
pathogenesis of postmenopausal osteoporosis (2), has received increasing attention in
studies examining periodontal diseases. There have been reports of
periodontitis-associated bone loss in the mandibular body in
ovariectomized (OVX) animals (3–6);
clinical observations in postmenopausal women have confirmed an
increased prevalence of periodontal disease with low estrogen
levels (7–11). Symptoms, such as thinning of the
mandibular inferior cortex (12,13) and residual ridge reduction of the
edentulous jaw (14,15) have been reported in patients with
postmenopausal osteoporosis, which suggest a link between
postmenopausal osteoporosis and the loss of periodontal tissue.
Conventional periodontal tissue regenerative
therapies, such as guided tissue regeneration, depending on the
individual anatomy of the defects or the amount of resident vital
periodontal ligament, can partially regenerate periodontal tissues
(16–21). Until now, the most ideal strategy
of periodontal tissue regeneration therapies is to control
inflammation and stimulate stem progenitors to regenerate new
periodontal tissues. The periodontal ligaments are very important
in maintaining the integrity of the periodontal tissue, which is
the connective tissue located between the alveolar bone and the
root surface of the tooth. The periodontal ligament stem cells
(PDLSCs) of this ligament exhibit osteoblast-like characteristics
and are capable of differentiating into cementoblasts or
osteoblasts, and can regenerate new functional periodontal support
tissues, including the cementum, the alveolar bone and the
periodontal ligament fiber (22,23), and are the most ideal seed cells
of periodontal tissue regeneration.
Nowadays, bone grafting procedure limitations have
shifted the focus of preclinical research to tissue engineering
strategies, which are strictly dependent on the proliferation and
differentiation ability of mesenchymal stem cells (MSCs); however,
they may be affected by genetics, aging, hormone levels, drug
consumption, or chronic systemic disease (24). MSCs have the ability to
self-renew, proliferate and differentiate toward different lineages
(osteoblasts, adipocytes, chondroblasts and myoblasts); however,
MSCs derived from bone marrow (BM-MSCs), which are the most
commonly employed cells in the clinical setting for different
orthopedic diseases (25), are
reported to be negatively influenced by various factors, most of
which are also responsible for osteoporosis (26), such as systemic diseases,
lifestyle, drug consumption and aging.
Several studies have also investigated the
association between changes in levels of the female sex hormone,
estrogen, and changes in PDLSCs (27,28), which modulates the activity of
PDLSCs by binding to the intracellular estrogen receptor (ER)
(29–31). Zhang et al (32) also reported that estrogen
deficiency leads to the impaired osteogenic differentiation of
PDLSCs in osteoporotic rats. Nonetheless, the mechanisms
responsible for the effects of estrogen on PDLSCs are poorly
understood. It is important to clarify the biological function of
the female sex hormone, estrogen, in PDLSCs, as this hormone may
affect periodontal health. Moreover, the effects of estrogen on the
bone regeneration potential of PDLSCs isolated from osteoporotic
rats in vivo have not yet been fully elucidated. Therefore,
it remains undetermined as to whether functional periodontal tissue
can be regenerated following the transplantation of autologous
PDLSCs isolated from osteoporotic animals.
The nano-hydroxyapatite/collagen/poly(L-lactide)
(nHAC/PLA), a three-dimensional (3D) nanostructured scaffold, is an
attractive bone substitute, as the novel biomimetic strategy used
to generate this scaffold endows it with properties similar to
those of natural bone. Cell culture and animal model experiments
have demonstrated that the composite material is highly
osteoconductive, biocompatible and bioresorbable (33,34).
In the present study, we created an estrogen
deficiency micro-environment by ovariectomy and examined the
effects of estrogen deficiency and estrogen supplementation on the
bone-forming capacity of PDLSCs derived from osteoporotic rats and
seeded on nHAC/PLA, both in in vitro and in vivo.
This may provide an ideal approach for functional periodontal
tissue regeneration in postmenopausal women with periodontal
disease.
Materials and methods
Establishment of the animal model of
osteoporosis
All surgical procedures and care administered to the
animals were approved by the University Animal Care Committee and
performed according to institutional guidelines. A total of 48
healthy 3-month old Sprague-Dawley (SD) female rats (Experimental
Animal Center of the Academy of Military Medical Sciences) were
randomly divided into 2 groups as follows: 24 animals were in the
bilaterally ovariectomized (OVX) group, and the other 24 animals
were subjected to sham surgery (sham-operated group, hereinafter
referred to as the sham group). In the sham procedure, the ovaries
were exteriorized and replaced intact to create a stress similar to
that obtained with bilateral ovariectomy, in accordance with the
method described in the study by Yu et al (35).
Measurement of body weight, bone mineral
density (BMD) and the estrogen level
At 0 (baseline) and 3 months after surgery, the SD
rats from the 2 groups were anesthetized with 10% chloral hydrate
(400 mg/kg) and weighed; blood samples were obtained from the
retro-orbital plexus to evaluate the estrogen level. The BMD of the
lumbar spine was measured using a DEXA scanner (GE Healthcare
Lunar; GE Healthcare, Madison, WI, USA). To measure the lumbar
spine BMD, the rats were placed in the prone position on the DEXA
plate and the X-ray tube was aligned along the longitudinal axis at
the mid-point of the body. The BMD value was determined using small
animal analysis software (GE Lunar Prodigy, Prodigy enCORE software
version 10.50.086; GE Healthcare).
Cell culture and isolation of PDLSCs
The rat PDLCs were isolated in accordance with the
method described in the study by Gay et al (36). Briefly, the sham-operated and
ovariectomized SD rats after 3 months of surgery were sacrificed
for gathering the periodontal ligament. The PDLCs were collected
from the periodontal ligament which were rinsed and cut into
trivial blocks under sterile conditions and digested in 0.3%
collagenase (Invitrogen, Carlsbad, CA, USA) at 37°C for 4 h on a
rotator set at 130 rpm. The cells otained were then seeded in a 75
cm2 culture flask and cultured in growth medium (GM)
containing Dulbecco's modified Eagle's medium (DMEM), 10% fetal
bovine serum (FBS), 100 IU/ml penicillin and 100 IU/ml streptomycin
(all from Invitrogen) in a humidified atmosphere (95% air, 5%
CO2) at 37°C. The cells were fed every 3 days with GM.
After the cells reached confluence, they were trypsinized and
plated in the culture flasks. Subsequently, PDLSCs were obtained as
previously described (37).
Briefly, STRO-1+ stem cells were isolated using
immunomagnetic beads (Dynabeads; Dynal Biotech, Oslo, Norway)
according to the manufacturer's instructions. After washing,
bead-positive cells were segregated using a magnetic particle
separator and subsequently seeded into a 75 cm2 culture
flask and cultured in GM in a humidified atmosphere (95% air, 5%
CO2) at 37°C. PDLSCs at passage 4 were used in each
experiment. At least 3 replicates were included for analysis.
Phenotypic analysis of PDLSCs
The PDLSCs at passage 4 isolated from the rats in
the sham group and OVX group were seeded into chamber slides,
cultured for 7 days in GM, fixed with cold acetone for 10 min and
immunostained for confocal laser scanning microscopy (CLSM). The
cells were blocked and permeabilized 1 h at room temperature in
Tris-buffered saline solution (TBS), pH 7.4, containing 10% FBS, 1%
bovine serum albumin (BSA) and 0.5% Triton X-100 (Sigma Chemical
Co., St. Louis, MO, USA). Antibody labeling was performed overnight
at room temperature [vimentin 1:100 (MAB2105), keratin 1:100
(MAB3165); R&D Systems, Minneapolis, MN, USA]. Secondary
antibodies were applied for 2 h at room temperature [anti-mouse IgG
TRITC 1:50 (sc-3796); Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA]. Following repeated washes with phosphate-buffered saline
(PBS), nuclear staining was performed with
4′,6-diamidino-2-phenylindole (DAPI; Sigma Chemical Co.) at room
temperature for 15 min, and Mowiol 4–88 confocal images were
recorded using a Zeiss LSM 5 Pascal system with a Zeiss Axiovert
microscope (Carl Zeiss, Oberkochen, Germany).
Immunohistochemistry for STRO-1 was performed on the
stem cells isolated from the 2 groups of rats cultured on chamber
slides for 7 days. The cells were fixed with 10% formalin, and were
then blocked and permeabilized 1 h at room temperature in TBS, pH
7.4, containing 10% FBS, 1% BSA and 0.5% Triton X-100. The cells
were then incubated with mouse antibodies against human STRO-1
(1:100, MAB1038) (R&D Systems) overnight at 4°C. The cells were
then washed with PBS and incubated with biotin-conjugated
swine-anti-mouse antibodies (sc-2031; Santa Cruz Biotechnology,
Inc.) for 2 h, washed in PBS and incubated with streptavidin-biotin
complex/horseradish peroxidase (Santa Cruz Biotechnology, Inc.) for
1 h. Staining was visualized using 3,3′-diaminobenzidine (DAB, 0.1
mg/ml, 0.02% H2O2) (Santa Cruz Biotechnology,
Inc.).
Methyl thiazolyl tetrazolium (MTT)
assay
MTT assay was performed as previously described in
the study by E et al (38). For the experiment, the cells were
divided into 3 groups as follows: the sham group, the OVX group and
the OVX + 17β-estradiol (E2) group. Briefly, the cells at passage 4
were plated in 96-well plates at 2×104 cells/ml with 200
µl GM for 24 h to allow attachment. The serum-containing
medium was then removed and replaced with a medium without FBS for
a further 12 h. The cells in the sham group and OVX group were
cultured in GM, and the cells in the OVX + E2 group were cultured
in GM + 10−7 M E2 (Sigma Chemical Co.). Following
culture for days 1, 2, 3, 4, 5, 6, 7 and 8, 20 µl 5 mg/ml
MTT solution/well was added to the cells, followed by culture for 4
h. The cells were washed twice with PBS, and 150 µl dimethyl
sulfoxide (Sigma Chemical Co.) was then added to each well,
followed by shaking for 10 min, and the optical density values for
each well were determined at 490 nm.
Cell culture protocol for histochemical
analysis of alkaline phosphatase (ALP) activity and
mineralization
For the experiment, the cells were divided into 3
groups: the sham group, the OVX group and the OVX + E2 group.
Briefly, the cells at passage 4 were plated at 1×105
cells/ml in 6-well culture plates with GM for 24 h to allow
attachment. The serum-containing medium was then removed and
replaced with a medium without FBS for a further 12 h. The cells in
the sham group and OVX group were cultured in an osteogenic medium
(ODM) [growth medium containing 10 nM dexamethasone, 50 mg/ml
ascorbic acid and 100 mM β-glycerophosphate (Sigma Chemical Co.)],
and the cells in the OVX + E2 group were cultured in ODM +
10−7 M E2. For the histochemical analysis of ALP
activity and mineralization, the cells were cultured for 21 days.
The medium was replaced every 4 days.
To determine ALP activity, the differentiated cells
cultured for 21 days were fixed with 10% formalin and stained using
the Gomori calcium-cobalt method. Briefly, the cells were washed
with PBS for 5 min. An incubation solution containing 5 ml 2%
barbital sodium, 5 ml 3% β-sodium glycerophosphate (Sigma Chemical
Co.), 10 ml 2% calcium nitrate, 5 ml 2% magnesium sulfate and 25 ml
distilled water was placed on each slide followed by incubation for
4 h at 37°C. The slides were then washed with distilled water and
incubated in 2% calcium nitrate for 2 min at room temperature.
Subsequently, the slides were incubated in 2% cobaltous nitrate for
2 min at room temperature. The slides were then washed with
distilled water and incubated in 1% ammonium sulfide for 1 min at
room temperature. The slides were then washed with running tap
water and left to dry.
To detect extracellular matrix calcification with
Alizarin red staining, on day 21 of differentiation, the osteogenic
medium-cultured cells were fixed with 10% formalin. The cells were
washed with PBS for 5 min. A 2% Alizarin red solution (Sigma
Chemical Co.) was placed on each slide followed by incubation for
10 min at room temperature. The slides were then washed with
running tap water for 5 min and left to dry.
Biometrics preparation of nHAC/PLA
grafts
The nHAC/PLA material (Beijing Allgens Medical
Science and Technology Co., Ltd., China, http://www.allgensmed.com/cn/index.aspx) mimics the
composition and the micro-structure characteristics of the natural
bone, the porosity is 80±10%, the pore size is 300±250 µm
and the calcium phosphate content is 45±5%. The mechanical strength
is ≥0.8 Mpa. The nHAC/PLA materials were cut into 10×4×3 or 5×4×3
mm blocks. The samples were then rinsed with 100% alcohol in order
to remove organic residues and with double distilled water in order
to remove inorganic residues (each solution for 10 min). The
samples were then sterilized by cobalt 60.
Seeding of cells onto nHAC/PLA
grafts
The PDLSCs isolated from the rats in the sham and
OVX groups were respectively seeded onto the nHAC/PLA scaffold. The
constructs were incubated in ODM for 2 h at 37°C, allowing the
cells to adhere to the scaffolds and 1 ml of additional ODM was
then added. The medium was replaced by ODM with or without E2 on
day 4 of incubation and the grafts were then ready for in
vitro experiments and in vivo implantation.
Scanning electron microscopy
The PDLSCs isolated from the rats in the sham and
OVX group were respectively seeded onto the nHAC/PLA scaffolds at
1×106 cells/cm2 per graft, and were then
cultured in ODM. Fixative was prepared from 2% paraformaldehyde and
2.5% glutaraldehyde (Sigma Chemical Co.) in 0.1 mol/l phosphate
buffer. Following fixation (30 min, 37°C), the samples from
nHAC/PLA, sham PDLSCs + nHAC/PLA, OVX PDLSCs + nHAC/PLA and OVX
PDLSCs + nHAC/PLA + E2 cultured in ODM for 7 days were rinsed twice
in PBS for 10 min and then washed 5 times (15 min each) in
different ethanol concentrations (50, 75, 90 and 95% v/v ethanol)
in distilled water and 3 times for 10 min each in analytical
ethanol. After the ethanol washes, the samples were rinsed in a
series of different hexamethyldisilazane (HMDS) concentrations
(33.3, 50 and 66.6% v/v) in analytic ethanol and 3 times in 100%
HMDS (1 min each). The morphological characterization of the cells
and materials was carried out by means of scanning electron
microscopy (SEM) using a Quanta 200 ESEM/SEM, FEI (Phillips,
Madison, WI, USA) with beam energies of 6–25 kV and fitted with an
energy dispersive spectroscopy (EDS) apparatus. The samples were
glued with conducing paste to appropriate mounting stabs, which
were then coated with a several nanometer-thick layer of gold. The
samples were examined under a Hitachi S-520 scanning electron
microscope (Hitachi, Tokyo, Japan).
Estimation of ALP activity and
osteocalcin (OCN) secretion
For the experiment, the cells were divided into 3
groups: sham PDLSCs + nHAC/PLA, OVX PDLSCs + nHAC/PLA and OVX
PDLSCs + nHAC/PLA + E2. The cells at passage 4 were plated in
6-well plates, at 1×105 cells/ml, with DMEM containing
10% FBS for 12 h to allow attachment. The serum-containing medium
was removed and replaced with a medium without serum for 12 h, and
the cells were then cultured in ODM. After 1–7, 7–14 and 14–21
days, the medium was collected from the wells.
ALP activity in the medium was assayed using a
biochemistry automatic analyzer (Hitachi 7600; Hitachi).
OCN in the medium was assayed using a mouse-specific
IRMA (Immutopics, Inc., San Clemente, CA, USA). Briefly, the sample
containing mouse OCN was incubated stimultaneously with an
antibody-coated bead and the 125I-labeled antibody. OCN contained
in the sample is immunologically bound by the immobilized antibody
and the radiolabeled antibody to form a 'sandwich' complex:
bead/anti-mouse OCN, mouse OCN, 125I-anti-mouse OCN. At the end of
the overnight incubation period, the bead was washed to remove any
unbound labeled antibody and other components. The radioactivity
bound to the bead was measured in a gamma counter. The
radioactivity of the bound antibody complex is directly
proportional to the bound antibody complex in the sample. As the
amount of extracellular matrix proteins interferes with the total
cellular protein determination, the data were determined and
expressed (as ng/ml) for each culture dish.
Mineral formation assay
For this experiment, the cells were divided into 3
groups as follows: the sham PDLSCs s+ nHAC/PLA, OVX PDLSCs +
nHAC/PLA and OVX PDLSCs + nHAC/PLA + E2 groups. The PDLSCs were
respectively seeded on nHAC/PLA at 1×105
cells/cm2 per well/graft for 24 h to allow attachment in
growth medium. The cells were starved using serum-free medium
containing 2% BSA, 100 U/ml penicillan and 100 U/ml streptomycin
for 24 h. The cells were then cultured in ODM. After 21 days of
culture, the samples were rinsed with PBS 3 times, fixed with 10%
formalin, rinsed with PBS, and stained at room temperature for 20
min with Alizarin red solution. After staining, all constructs were
washed with distilled water until the supernatant was clear.
Digital images were recorded. For optical density measurements,
each construct was eluted for 30 min with 1 ml 10% cetylpyridinium
chloride monohydrate. The optical density at 540 nm was determined
using a universal microplate reader (ELx800UV, Bio-Tek Instruments,
Inc., Bad Friedrichshall, Germany). The optical density was
calculated by taking the average optical absorbance of constructs
grown in ODM.
Real-time-polymerase chain reaction
(RT-PCR)
For the experiment, the cells were divided into 3
groups as follows: the sham PDLSCs + nHAC/PLA, OVX PDLSCs +
nHAC/PLA and OVX PDLSCs + nHAC/PLA + E2 groups. The PDLSCs were
respectively seeded on nHAC/PLA at 1×105
cells/cm2 graft for 24 h to allow attachment in ordinary
medium. The cells were starved using serum-free medium containing
2% BSA, 100 U/ml penicillan and 100 U/ml streptomycin for 24 h. The
constructs were then cultured in ODM with or without
10−7 M E2. After 14 days of culture, the cells were
crushed in lysis buffer (Roche, Indianapolis, IN, USA) with an
RNase-free piston (Pellet), vortexed and spun. The clear cell
lysate was transferred to QiaShredder (Qiagen, Inc., Valencia, CA,
USA) columns for RNA purification. RNase-free DNase (Roche) was
used to eliminate DNA contamination of RNA samples. Purified RNA
was dissolved in RNase-free water and its concentration was
assessed by reading at 260 nm. RNA quality was checked on a 2%
agarose gel with 1 mg/ml ethidium bromide. The samples were stored
at −80°C until use. Complementary DNA (cDNA) was synthesized from 2
µg of total RNA using the First Strand cDNA synthesis kit
for RT-PCR (AMV; Roche).
cDNA mixture (8 µl) diluted 1:20 in water was
subjected to real-time PCR using SYBR-Green I dye (LightCycler
FastStart DNA master SYBR-Green I; Roche, Penzberg, Germany).
Reactions were performed in 20 µl PCR mixture containing 4
µl 5X Master Mix (dNTP mixture with dUTP instead of dTTP,
MgCl2, SYBR-Green I dye, TaqDNA polymerase and reaction
buffer), 2 µl of 10 µM primers. The primer sequences
(Shanghai Sangon Biological Engineering Technology and Services
Co., Ltd., Shanghai, China) for ALP, OCN, ERα, ERβ and β-actin are
listed in Table I. β-actin
real-time PCR was run as a control to monitor RNA integrity and to
be used for normalization. The specificity of each primer pair was
confirmed by melting curve analysis.
 | Table IPrimers used for real-time PCR with
SYBR-Green. |
Table I
Primers used for real-time PCR with
SYBR-Green.
| Gene | Primer
sequence | Species | Product size
(bp) |
|---|
| ERα | F: 5′-CAT CGA TAA
GAA CCG GAG GA-3′ | Rat | 190 |
| R: 5′-AAG GTT GGC
AGC TCT CAT GT-3′ | | |
| ERβ | F: 5′-AGC AAC TGG
TGC TCA CCC T-3′ | Rat | 94 |
| R: 5′-GTC CGC CAG
CTT AGT GAG G-3′ | | |
| ALP | F: 5′-TCC CAC GTT
TTC ACG TTT-3′ | Rat | 140 |
| R: 5′-GAG ACG TTC
TCC CGT TCA C-3′ | | |
| OCN | F: 5′-TGA GGA CCC
TCT CTC TGC TC-3 | Rat | 150 |
| R: 5′-AGG TAGCGC
CGG AGT CTA TT-3 | | |
| β-actin | F: 5′-CCC ATC TAT
GAG GGT TAC GC-3′ | Rat | 150 |
| R: 5′-TTT AAT GTC
ACG CAC GAT TTC-3′ | | |
Histological and morphometric analysis of
the in vivo experiments
For this experiment, the cells were divided into 4
groups as follows: the nHAC/PLA, sham PDLSCs + nHAC/PLA, OVX PDLSCs
+ nHAC/PLA and OVX PDLSCs + nHAC/PLA + E2 groups. The PDLSCs were
seeded onto nHAC/PLA scaffolds at 1×108
cells/cm2 per graft. Following 7 days of culture in
vitro, the engineered constructs were then implanted into the
backs of 8 severe combined immunodeficient (SCID) mice (age, 6–8
weeks; weight, 20 g) for in vivo bone regeneration. Briefly,
the SCID mice were anesthetized by an injection of pentobarbital
sodium (70 mg/kg) to the abdominal cavity, and the skin on the
backs of the mice was disinfected with iodine. After a small
incision was made, the 4 grafts were transplanted into the backs of
the SCID mice using tweezers, and the incision was then sutured.
After 12 weeks, the SCID mice were sacrificed for removing the
implanted constructs. The harvested constructs were then fixed in
10% formalin neutral buffer solution at pH 7.4 for 2 days and were
then trimmed using waterproof polishing paper without
demineralization and cut into 5-µm-thick sections, and
stained with hematoxylin and eosin (H&E) for light microscopic
observation. All the samples were analyzed microscopically and
compared with the controls. Digital images were recorded.
For morphometric analysis, 5 sequential sections per
implant were selected for evaluation under low magnification,
allowing the coverage of the entire implant. Using a Leica-Qwin 3.2
image analysis system (Leitz DMRD; Leica Microsystems Inc.,
Bannockburn, IL, USA), all slides were analyzed to identify the
type of tissue (mature bone-like and osteoid-like). The extent of
newly formed mature bone and osteoid was indicated by the
percentage of bone formation area within the section, Total scores
per section were calculated and averaged for all sections to obtain
an overall score for each implant.
Statistical analysis
All data are presented as the means ± standard
deviation (SD). The statistical analyses of the results were
performed using computer-based SPSS 16.0 software. A one-way
analysis of variance with the post hoc test LSD for multiple
comparisons was used to determine statistically significant
differences in the cell proliferation, mineral formation, the
expression levels of ALP, OCN, ERα and ERβ mRNAs and the percentage
of bone formation area between the study groups. The repeated
measures were used to determine the statistically significant
differences in ALP activity and OCN secretion. Values of P<0.05
were considered to indicate statistically significant
differences.
Results
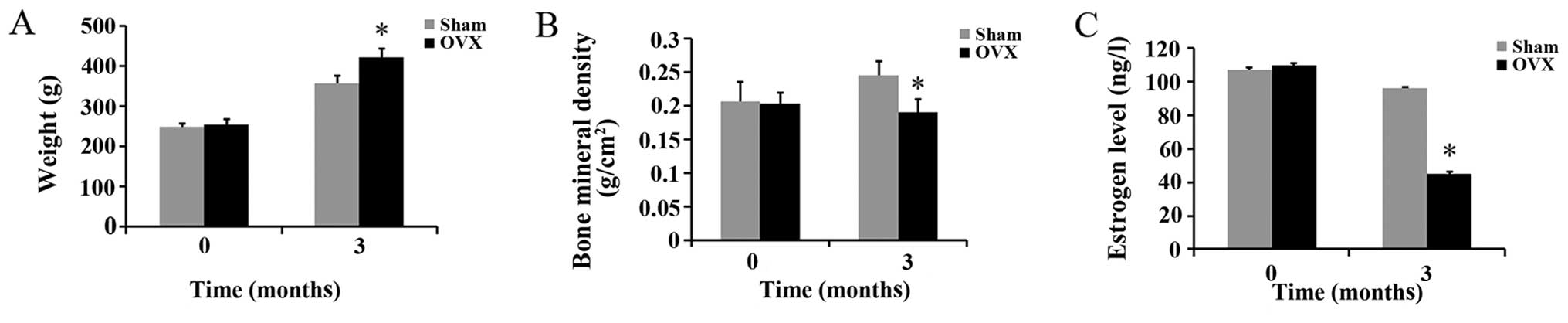
Effects of ovariectomy on body weight and
BMD, and estrogen levels in rats
There were no significant differences in body
weight, BMD and estrogen levels between the rats in the sham and
OVX groups before the experiment. Three months after the
ovariectomy, the body weight of the rats in the OVX group was
significantly higher than that of the rats in the sham group;
however, the BMD and estrogen levels of the rats in the OVX group
were significantly lower than those of the rats in the sham group
(Fig. 1).
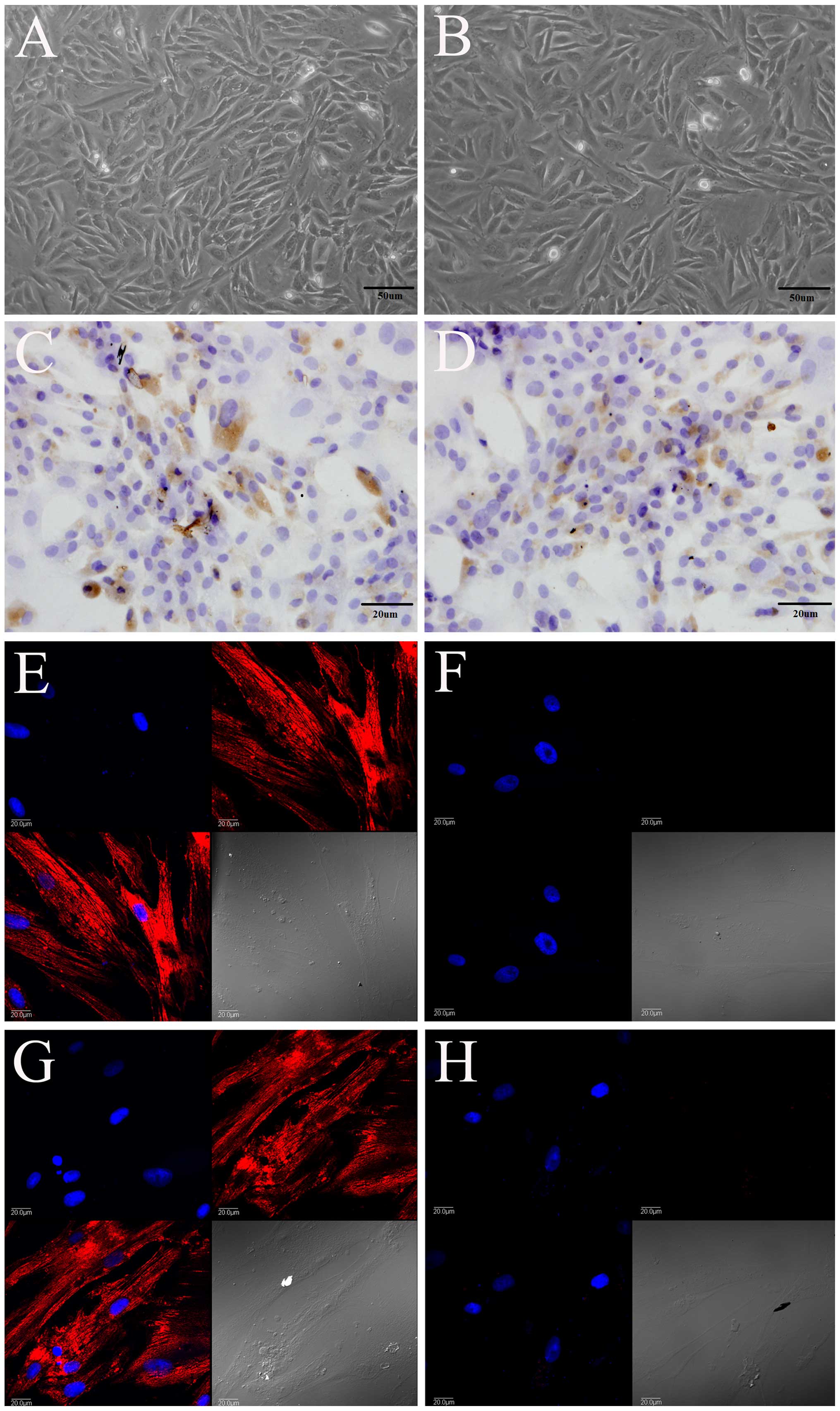
Morphology and identification of rat
PDLSCs
Morphologically, the PDLSCs isolated from the rats
in the sham group (Fig. 2A) and
OVX group (Fig. 2B) had a
triangular, spindle and fusiform shape, and expressed STRO-1, as
shown by immunohistochemical staining (Fig. 2C and D). They also expressed
vimentin (Fig. 2E and G), but did
not express keratin (Fig. 2F and
H), as shown by immunofluorescence staining. Vimentin was
labeled red with TRITC, and the nucleus was labeled blue with
DAPI.
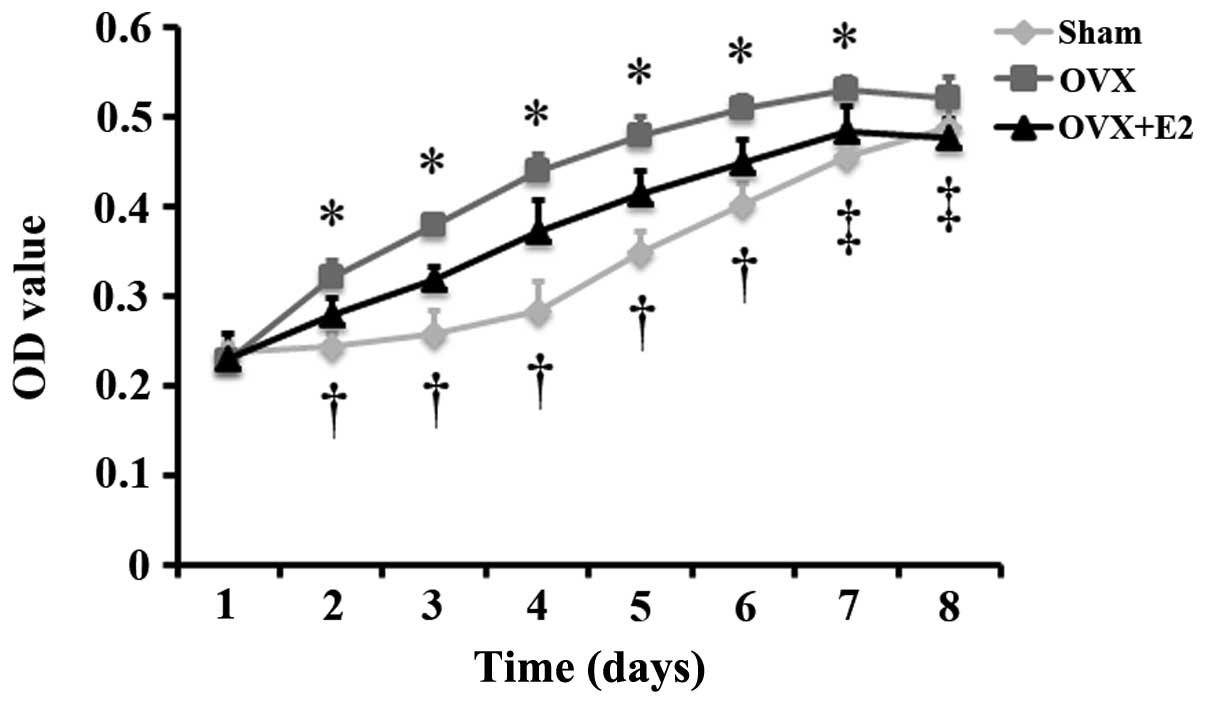
Effect of estrogen on the proliferation
of PDLSCs isolated from OVX rats
To observe the differences in the proliferation of
cells from the sham and OVX group, and to examine the effects of
estrogen on the proliferation of cells from the OVX group, MTT
assay was carried out to monitor the proliferation of the 3 groups
of cells (sham group, OVX group and OVX + group). The results
revealed that the proliferation of the cells in the 3 groups
increased gradually with the extension of the culture time, and
that the proliferation of the cells in the OVX and OVX + E2 groups
reached peak levels on on day 7. Following culture for 2, 3, 4, 5,
6 and 7 days, the proliferation of the cells in the OVX group was
significantly greater than that of the cells in the sham group. The
proliferation of the cells in the OVX + E2 group was significantly
lower than that of the cells in the OVX group following culture for
2, 3, 4, 5, 6, 7 and 8 days, and was significantly greater than
that of the cells in the sham group following culture for 2, 3, 4,
5 and 6 days (Fig. 3).
Effects of estrogen on the osteogenic
differentiation of the PDLSCs isolated from OVX rats
Following culture in ODM for 21 days, the cells from
the sham group exhitibed intense staining for ALP (Fig. 4A), and also exhibited calcium
deposition, with formed and developed mineralization nodules
(Fig. 4D), as revealed by the
Gomori calcium-cobalt method and Alizarin red staining. Weaker
staining was observed in the cells from the OVX group (Fig. 4B and E). When the cells from the
OVX group were cultured in ODM + 10−7 E2 for 21 days,
the cells exhibited intense staining for ALP (Fig. 4C), and also exhibited calcium
deposition, with formed and developed mineralization nodules
(Fig. 4E).
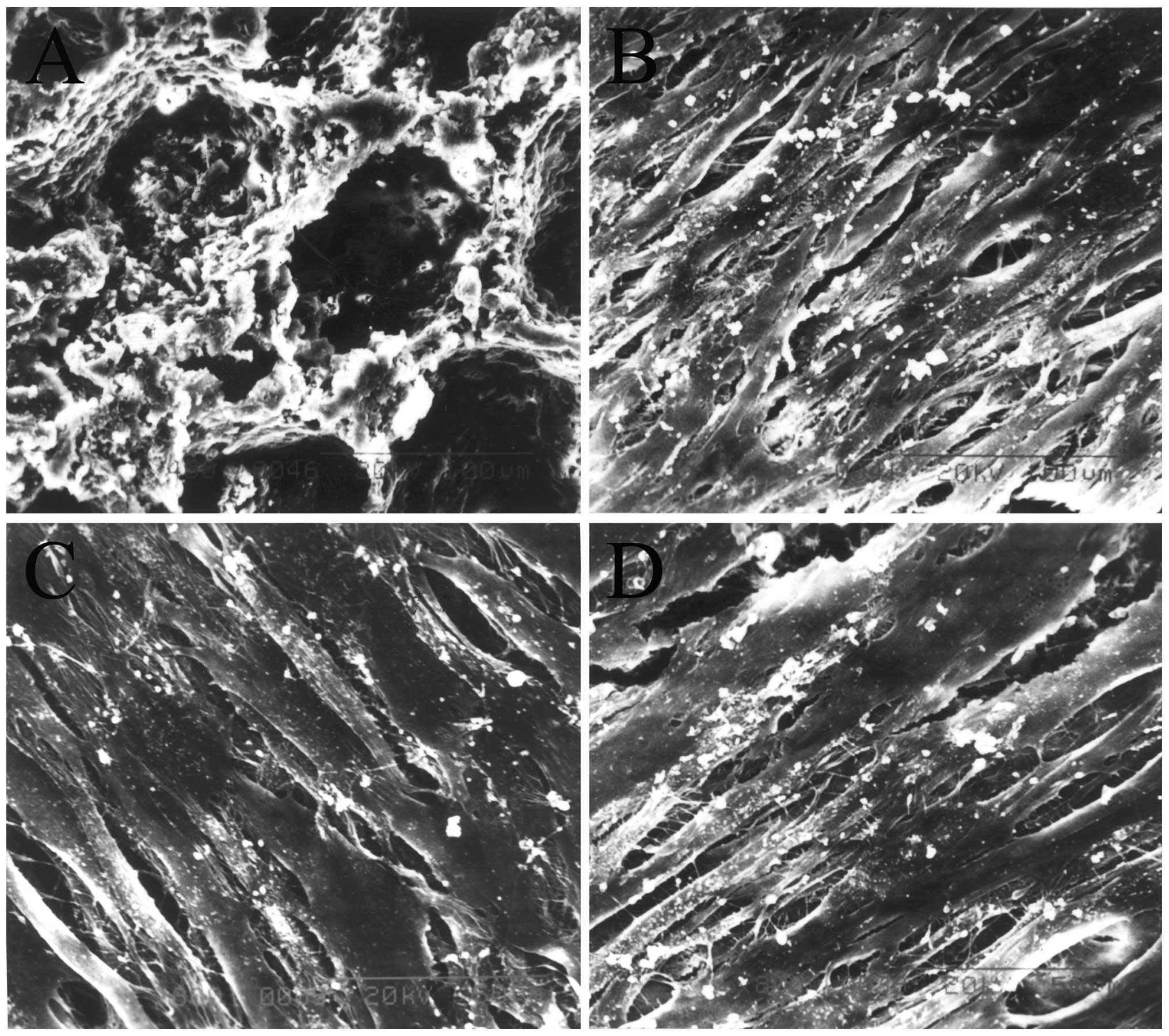
Scanning electron microscopy
The results of SEM revealed that the nHAC/PLA had
the micro-structure characteristics of natural bone (Fig. 5A). When the constructs were
cultured in ODM for 7 days, a large number of cells in the sham
PDLSCs + nHAC/PLA (Fig. 5B), OVX
PDLSCs + nHAC/PLA (Fig. 5C) and
OVX PDLSCs + nHAC/PLA + E2 (Fig.
5D) groups had adhered to, and had expanded and proliferated on
the nHAC/PLA. There were many filamentous extracellular matrices on
the surface of the cells. Some of the cells in the sham PDLSCs +
nHAC/PLA and OVX PDLSCs + nHAC/PLA + E2 groups were covered by more
mineral deposits.
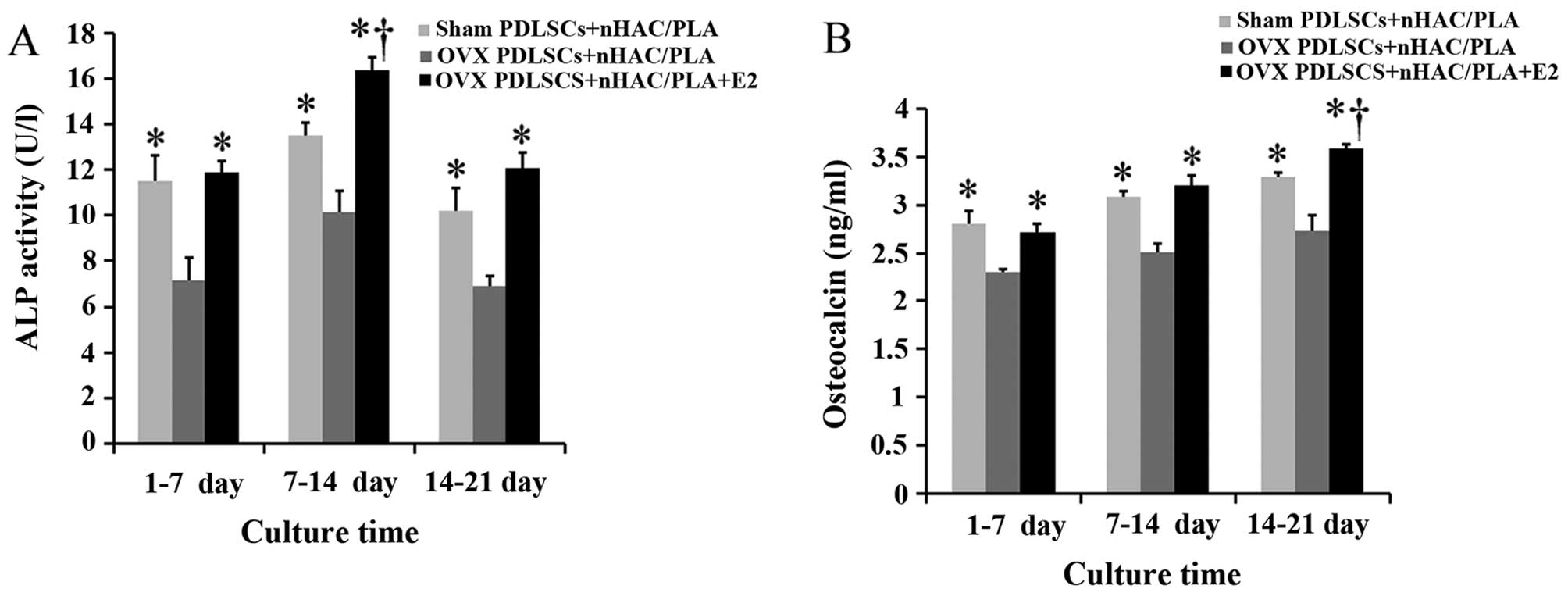
Effects of estrogen on ALP activity and
OCN secretion in the PDLSCs isolated from OVX rats and seeded on
nHAC/PLA
The specific ALP activity of the PDLSCs from the 3
groups reached the highest levels during the culture period of 7–14
days. Compared with the cells isolated from the sham-operated rats,
the ovariectomy significantly decreased ALP activity in the cells
during the culture period of 1–7, 7–14 and 14–21 days.
Nevertheless, treatment with E2 significantly increased ALP
activity in the cells isolated from the OVX rats, and the value of
ALP activity in the OVX PDLSCs + nHAC/PLA + E2 group was
significantly higher than that of the sham PDLSCs + nHAC/PLA group
during the culture period 7–14 days (Fig. 6A).
OCN secretion from the PDLSCs in the 3 groups
gradually increased with the extension of the culture time.
Compared with the cells isolated from the sham-operated rats, the
ovariectomy significantly decreased OCN secretion from the cells
isolated from the OVX rats during the culture period of 1–7, 7–14
and 14–21 days. Nevertheless, E2 significantly increased OCN
secretion from the cells isolated from the OVX rats, and the value
of OCN secretion in the OVX PDLSCs + nHAC/PLA + E2 group was
significantly higher than that of the sham PDLSCs + nHAC/PLA group
during the culture period of 14–21 days (Fig. 6B).
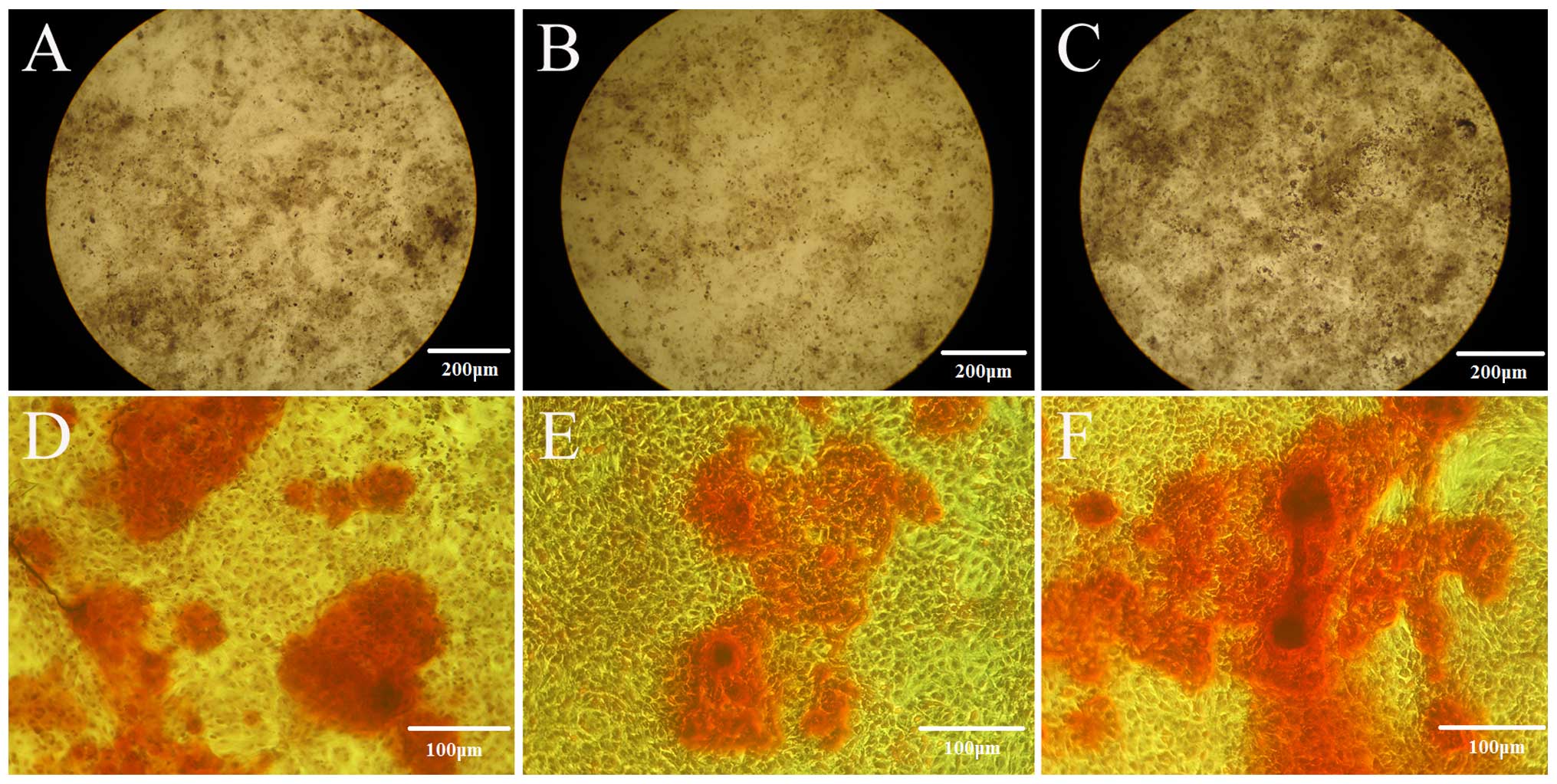
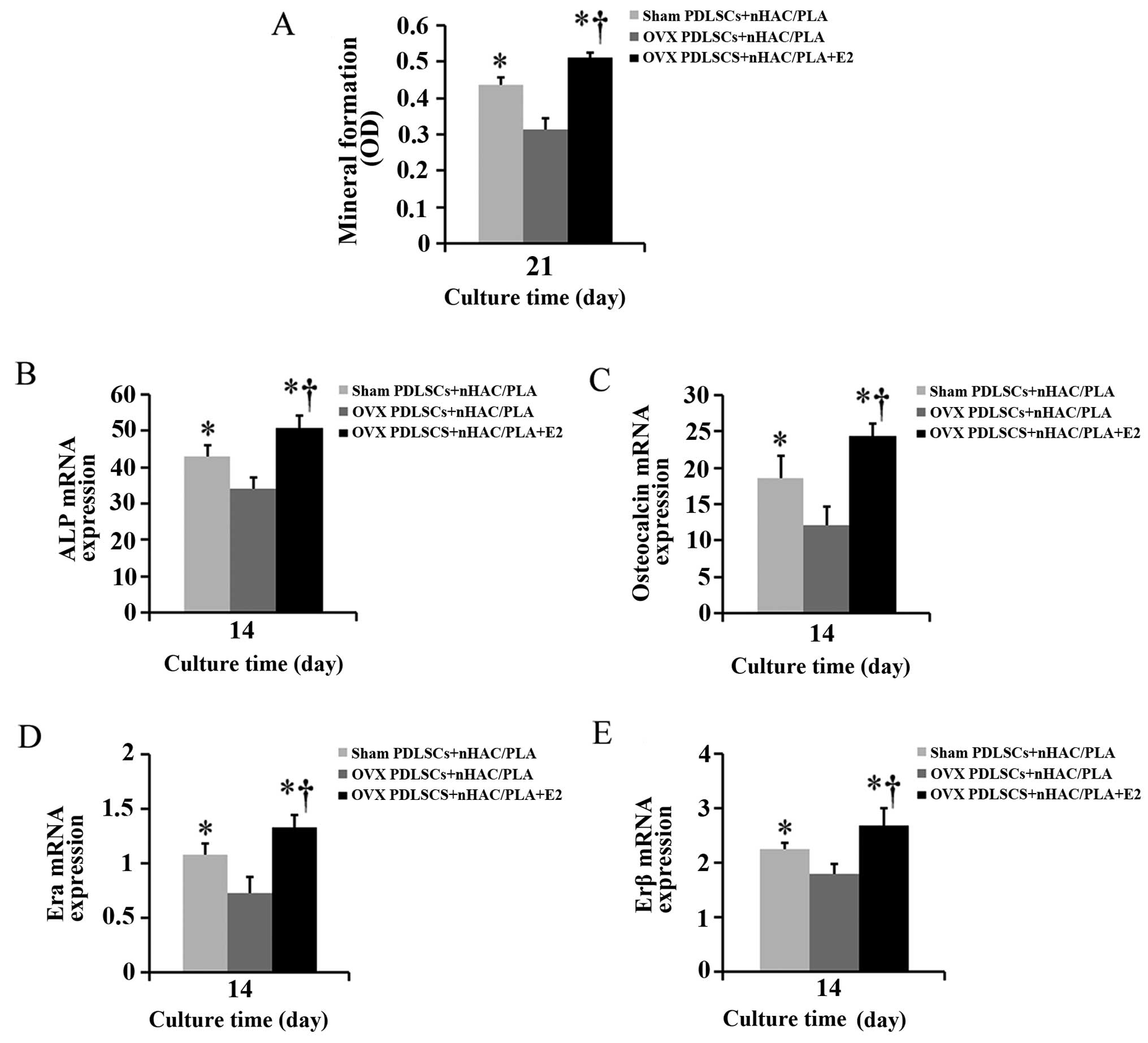
Effect of estrogen on mineral formation
in the PDLSCs isolated from OVX rats and seeded on the nHAC/PLA
scaffold
Alizarin red staining was used to quantify calcium
phosphate mineral formation in the cells on the sham PDLSCs +
nHAC/PLA, OVX PDLSCs + nHAC/PLA and OVX PDLSCs + nHAC/PLA + E2
constructs following culture for 21 days. The mineral formation in
the cells on the OVX PDLSCs + nHAC/PLA construct was significantly
less than that of the cells on the sham PDLSCs + nHAC/PLA and OVX
PDLSCs + nHAC/PLA + E2 (Fig.
9A–C) constructs, and mineral formation in the cells on the OVX
PDLSCs + nHAC/PLA + E2 construct was significantly greater than
that of the cells on the sham PDLSCs + nHAC/PLA (Fig. 7A).
Effect of estrogen on the mRNA expression
levels of ALP, OCN, ERα and ERβ in the PDLSCs isolated from OVX
rats and seeded on nHAC/PLA
When the PDLSCs from the sham-operated and OVX rats
were seeded on the nHAC/PLA and were cultured in ODM with or
without E2 for 14 days, real-time PCR analysis revealed that the
mRNA expression levels of ALP (Fig.
7B), OCN (Fig. 7C), ERα
(Fig. 7D) and ERβ (Fig. 7E) in the PDLSCs from the OVX rats
were significantly down-regulated due to the ovariectomy. E2
significantly increased the mRNA expression levels of ALP, OCN, ERα
and ERβ in the PDLSCs from the OVX rats, and the expression levels
in the OVX PDLSCs + nHAC/PLA + E2 group were significantly higher
than those in the sham PDLSCs + nHAC/PLA group.
Histological and morphometric analysis of
in vivo experiments
The PDLSCs isolated from the sham-operated and OVX
rats were seeded onto the nHAC/PLA porous scaffolds and cultured in
ODM or ODM + E2 for 7 days, and the cell/scaffold constructs were
then implanted subcutaneously into SCID mice. After 12 weeks, the
subcutaneous implants were removed from the mice and analyzed for
new bone formation. H&E staining revealed that nHAC/PLA alone
exhibited no new bone formation, while the marked ingrowth of soft
connective tissue into the grafts was observed (Fig. 8A). Newly formed bones were
observed in the sham PDLSCs + nHAC/PLA (Fig. 8B), OVX PDLSCs + nHAC/PLA (Fig. 8C) and OVX PDLSCs + nHAC/PLA + E2
(Fig. 8D) constructs. Newly
formed blood vessels were observed in the micropore of the
nHAC/PLA. Osteoblastic cells lined the surface of the newly formed
bone.
 | Figure 8In vivo osteogenesis. (A)
nHAC/PLA, (B) sham PDLSCs + nHAC/PLA, (C) OVX PDLSCs + nHAC/PLA,
(D) OVX PDLSCs + nHAC/PLA + E2. Magnification, ×200, NB, newly
formed bone. Black arrowheads indicate blood vessels, white
arrowheads indicate osteoblastic cells. OVX, ovariectomized PDLSCs,
periodontal ligament stem cells; nHAC/PLA,
nano-hydroxyapatite/collagen/poly(L-lactide). |
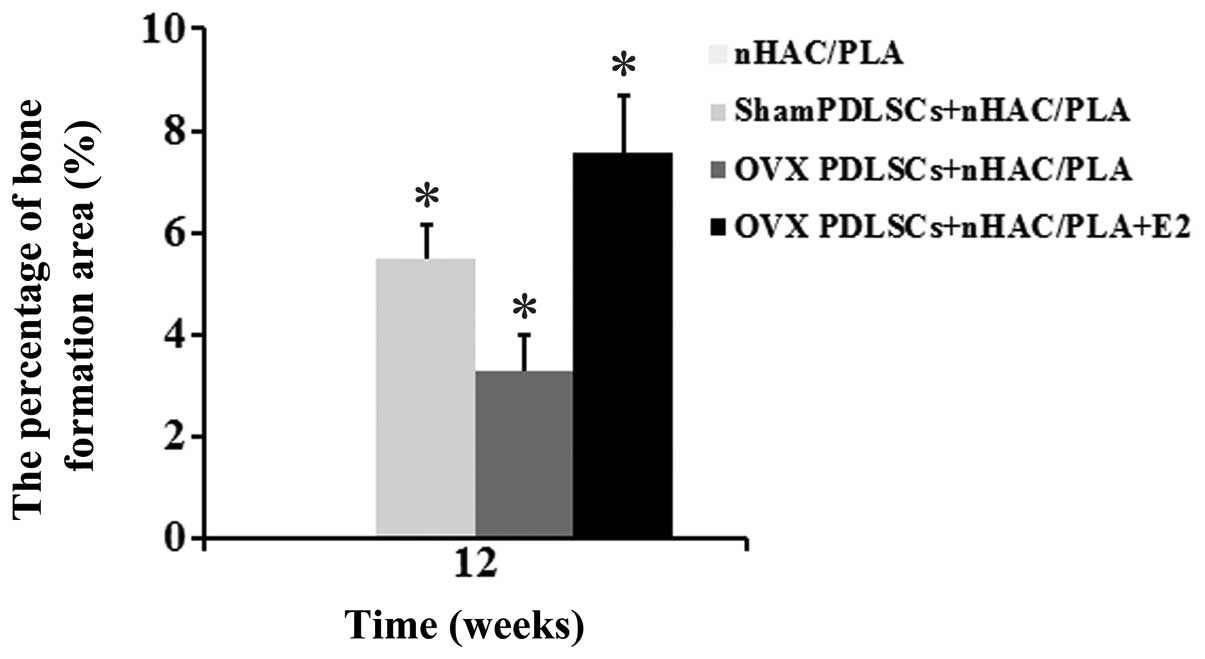
The results of histomorphometric analysis are
summarized in Fig. 9. After 12
weeks of implantation, the percentages of bone formation area
between the groups differed significantly. The percentage of bone
formation area in the sham PDLSCs + nHAC/PLA group (5.498±0.65%)
and was significantly higher than that of the OVX PDLSCs + nHAC/PLA
group (3.269±0.72%), but was significantly lower than that of the
OVX PDLSCs + nHAC/PLA + E2 group (7.573±1.1%). The nHAC/PLA alone
exhibited no bone formation.
Discussion
Autologous adult stem cell-based tissue engineering
and regenerative medicine has been considered a promising
substitute for current clinical treatments that restore tissue and
organ deficiencies (39–41). Successful tissue regeneration
requires a sufficient cell population with high differentiation
potential.
PDLSCs can regenerate new functional periodontal
support tissue, including cementum, alveolar bone and periodontal
ligament fibers. Therefore, PDLSC-based tissue engineering has now
emerged as a promising and ideal alternative approach for the
clinical treatment of periodontal tissue defects (22,23). However, a number of factors
(including advanced age, degenerative diseases of donors and the
microenvironment of systemic disease, such as hyperglycemia and
estrogen deficiency) affect the PDLSC population, the proliferation
rate and the differentiation potential (32,37,42), indicating that the changes in the
microenvironment are responsible for PDLSCs exhibiting different
characteristics.
In light of the above findings, the aim of the
present study was the detailed characterization of the properties
of PDLSCs derived from OVX rats. An ovariectomy was performed in
order to create an estrogen-deficient microenvironment. A number of
studies have suggested that an ovariectomy can create an
estrogen-deficient microenvironment (32,43–46). In this study, the weight, BMD of
the lumbar spine and estrogen levels in the 2 groups of rats
(sham-operated and OVX rats) were examined, an these values
differed significantly between the sham and OVX group at the 3rd
month after surgery, but not at day 0 after surgery. The OVX rats
had a higher weight, and lower BMD and a lower estrogen level.
These results demonstrated that we successfully created an
estrogen-deficient microenvironment by ovariectomy.
Our first goal was to define the proliferative
ability of the PDLSCs in regards estrogen depletion. In our primary
PDLSC cultures, we observed that morphologically, the 2 groups of
cells had a triangular, spindle and fusiform shape, with no
significant differences between the 2 groups. The PDLSCs from the
sham group attached more easily, and expanded earlier than those
from the OVX group. After the primary cells were passaged, the
PDLSCs from the OVX group proliferated more rapidly. This indicated
that the estrogen deficiency enhanced the proliferative ability of
the PDLSCs. However, ALP staining and Alizarin red staining
demonstrated that estrogen deficiency inhibited the osteogenic
differentiation of the PDLSCs in monolayer culture conditions.
These results were consistent with those reported in the study by
Zhang et al (32). A
possible explanation for the increased proliferation is that
estrogen deficiency makes an organism produce more PDLSCs in a
compensative manner. However, the increased proliferative ability
of the PDLSCs failed to enhance the osteogenic differentiation
ability, which is why postmenopausal women often suffer from
osteoporosis. However, estrogen supplements can reverse the effects
of estrogen deficiency. This may be why postmenopausal women adopt
estrogen replacement therapy to reduce immediate and long-term
excessive bone mass loss caused by low levels of estrogen, and to
prevent or delay the onset of osteoporosis.
Our second goal was to analyze the osteogenic
differentiation ability of PDLSCs derived from OVX rats cultured in
3D nHAC/PLA nanostructured scaffolds. A number of studies have
demonstrated that the population, proliferation rate and
differentiation potential of adult stem cells are affected by
systemic diseases, lifestyle, drug consumption and aging (24,26,32,37,42). The results from the study by
Bressan et al (47)
demonstrated that dental pulp stem cells derived from an aged group
exhibited a low proliferative and osteogenic differentiation
ability in monolayer culture conditions; when cultured on
hydroxyapatite (HA) nanostructured granules and used in vivo
to repair critical size defects, they exhibited the same ability as
those of the younger group in terms of time to repair and the
quality of extracellular matrix. Their study indicates that 3D HA
nanostructured granules can reverse the effects of advanced age on
the proliferative ability and the osteogenic differentiation
ability of dental pulp stem cells. However, in the present study,
the 3D nHAC/PLA nanostructured scaffolds did not alter the effects
of estrogen deficiency on the osteogenic differentiation ability of
the PDLSCs. When the PDLSCs were cultured on nHAC/PLA in ODM, the 3
groups of cells (sham PDLSCs + nHAC/PLA, OVX PDLSCs + nHAC/PLA and
OVX PDLSCs + nHAC/PLA + E2) all adhered, expanded and proliferated,
and produced a large amount of mineral matrix, as shown by SEM;
however, the ALP activity and OCN secretion in the cells derived
from the OVX group decreased due to estrogen deficiency. The
results from real-time PCR further confirmed this by assessing the
intrinsic expression of osteogenic markers. In order to further
examine the effects of estrogen deficiency on the osteogenic
differentiation of PDLSCs derived from OVX rats, we also examined
the mineral formation of the cells cultured in monolayer conditions
and on the nHAC/PLA. The results revealed that, in monolayer
conditions, more positive strong Alizarin red staining existed in
the cells derived from the sham group. In 3D nHAC/PLA conditions,
quantitative analysis of Alizarin red staining suggested that
estrogen deficiency significantly decreased mineral formation in
the cells. Previous studies have demonstrated that estrogen plays
an important role in the osteogenic differentiation of PDLSCs by
binding specific intracellular ER (29–32). Thus, we also examined the mRNA
expression of ER in the cells derived from OVX rats. Estrogen
deficiency downregulated the mRNA expression of ERα and ERβ. There
were no differences between the cells cultured on 3D nHAC/PLA
nanostructured scaffolds in our study and the cells were cultured
in monolayer conditions in a previous study (32). When the PDLSCs derived from OVX
rats were treated with estrogen, the results revealed that the
estrogen-treated PDLSCs exhibited increased ALP activity, OCN
secretion and mineral formation, as well as an increased mRNA
expression of ALP and OCN, and ERα and ERβ. These results indicated
that the estrogen-deficient microenvironment impaired the
osteogenic differentiation of PDLSCs derived from OVX rats.
Estrogen enhanced the osteogenic differentiation of PDLSCs derived
from OVX rats and seeded on nHAC/PLA in vitro. Both ERα and
Erβ were involved in the osteogenic differentiation of the
PDLSCs.
In in vitro experiments, we demonstrated that
treatment with estrogen restored the osteogenic differentiation
capacity of the PDLSCs seeded on nHAC/PLA, which had been impaired
due to estrogen deficiency. As a final goal, we examined whether
the PDLSCs derived from OVX rats and seeded on 3D nano-structured
scaffolds were able to be undergo osteogenesis in vivo. The
cells seeded on the nHAC/PLA were cultured for 7 days in
vitro, and the constructs were then implanted subcutaneously
into the backs of SCID mice for 12 weeks, and we then observed that
the impaired PDLSCs (OVX PDLSCs + nHAC/PLA) exhibited new bone
formation abilities, but their regenerative ability was
significantly lower than that of the other 2 groups (sham PDLSCs +
nHAC/PLA and OVX PDLSCs + nHAC/PLA + E2). This ability was
comparable between the estrogen-treated group and sham group.
In conclusion, to date, a number of studies have
demonstrated that postnatal stem cells can be isolated from various
adult tissues, such as bone marrow (25), skeletal muscle (48), brain (49), liver (50), pancreas (51), lungs (52), heart (53) and kidneys (54), and that these cells exhibit
self-renewal capacity and multilineage differentiation potential.
However, there are some disadvantages associated with harvesting
stem cells, such as additional damage to the body. The harvesting
of stem cells from various tissues or organs can damage those
organs, such as in stem cells derived from bone marrow (25), skeletal muscle (48), brain (49), liver (50), pancreas (51), lungs (52), heart (53) and the kidneys (54). Therefore, we focused on utilizing
tissues that can be obtained without additional injury, as well as
on new stem cell sources, such as periodontal ligament stem cells
from teeth extracted for orthodontic purposes, prophylactically
extracted nondecayed third molar teeth. Periodontal ligament stem
cells have many advantages, and the results to date suggest that
teeth are a viable source of adult MSCs for a wide range of
clinical applications (22,23,37). Thus far, no investigation on the
stemness of PDLSCs cultured on 3D nHAC/PLA nanostructured scaffolds
and its correlation with estrogen deficiency and estrogen
supplements has been performed, at least to the best of our
knowledge.
Our data provide three important insights. The first
is that this study clearly demonstrates that PDLSCs exhibit a
response to estrogen deficiency at the 3rd month following an
ovariectomy. The PDLSCs derived from OVX rats exhibit an increased
proliferative ability and a weaker osteogenic differential ability.
The second important conclusion is that if cells from OVX rats are
cultured on 3D nHAC/PLA nanostructured scaffolds, they regain the
same biological properties as those of cells cultured in monolayer
conditions. Both ERα and Erβ are involved in the osteogenic
differentiation of PDLSCs. The third conclusion is that estrogen
can enhance the bone regeneration potential of periodontal ligament
stem cells derived from OVX rats and seeded on nHAC/PLA in
vivo. The impaired PDLSCs from OVX rats can form new bone, but
their regenerative ability is limited. Further studies are required
to investigate the in situ bone regenerative capacity of
estrogen-treated PDLSCs derived from OVX rats and seeded on 3D
nHAC/PLA nanostructured scaffolds.
Acknowledgments
The authors would like to thank the staff and
faculty members of the Institute of Stomatology of the General
Hospital of People's Liberation Army of China. This study was
funded by the National High-tech R&D Program (863 Program),
grant no. 2013AA032201, the PLA General Hospital Clinical Research
Support Fund, grant no. 2015FC-CXYY-1003.
References
|
1
|
Bertonazzi A, Nelson B, Salvador J and
Umland E: The smallest available estradiol transdermal patch: a new
treatment option for the prevention of postmenopausal osteoporosis.
Womens Health (Lond Engl). 11:815–824. 2015. View Article : Google Scholar
|
|
2
|
Daniell HW: Postmenopausal tooth loss.
Contributions to edentulism by osteoporosis and cigarette smoking.
Arch Intern Med. 143:1678–1682. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tominari T, Hirata M, Matsumoto C, Inada M
and Miyaura C: Polymethoxy flavonoids, nobiletin and tangeretin,
prevent lipopolysaccharide-induced inflammatory bone loss in an
experimental model for periodontitis. J Pharmacol Sci. 119:390–394.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kobayashi M, Matsumoto C, Hirata M,
Tominari T, Inada M and Miyaura C: The correlation between
postmenopausal osteoporosis and inflammatory periodontitis
regarding bone loss in experimental models. Exp Anim. 61:183–187.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marques MR, da Silva MA, Manzi FR,
Cesar-Neto JB, Nociti FH Jr and Barros SP: Effect of intermittent
PTH administration in the periodontitis-associated bone loss in
ovariectomized rats. Arch Oral Biol. 50:421–429. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Duarte PM, de Assis DR, Casati MZ, Sallum
AW, Sallum EA and Nociti FH Jr: Alendronate may protect against
increased periodontitis-related bone loss in estrogen-deficient
rats. J Periodontol. 75:1196–1202. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reinhardt RA, Payne JB, Maze CA, Patil KD,
Gallagher SJ and Mattson JS: Influence of estrogen and
osteopenia/osteoporosis on clinical periodontitis in postmenopausal
women. J Periodontol. 70:823–828. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Palomo L, Chitguppi R, Buencamino MC,
Santos D and Thacker H: A need to educate postmenopausal women of
their periodontal health. J Indian Soc Periodontol. 17:225–227.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pepelassi E, Nicopoulou-Karayianni K,
Archontopoulou AD, Mitsea A, Kavadella A, Tsiklakis K, Vrotsos I,
Devlin H and Horner K: The relationship between osteoporosis and
periodontitis in women aged 45–70 years. Oral Dis. 18:353–359.
2012. View Article : Google Scholar
|
|
10
|
Haas AN, Rösing CK, Oppermann RV, Albandar
JM and Susin C: Association among menopause, hormone replacement
therapy, and periodontal attachment loss in southern Brazilian
women. J Periodontol. 80:1380–1387. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lerner UH: Inflammation-induced bone
remodeling in periodontal disease and the influence of
post-menopausal osteoporosis. J Dent Res. 85:596–607. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Taguchi A, Tanimoto K, Suei Y, Otani K and
Wada T: Oral signs as indicators of possible osteoporosis in
elderly women. Oral Surg Oral Med Oral Pathol Oral Radiol Endod.
80:612–616. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Klemetti E: A review of residual ridge
resorption and bone density. J Prosthet Dent. 75:512–514. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hirai T, Ishijima T, Hashikawa Y and
Yajima T: Osteoporosis and reduction of residual ridge in
edentulous patients. J Prosthet Dent. 69:49–56. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
von Wowern N and Kollerup G: Symptomatic
osteoporosis: A risk factor for residual ridge reduction of the
jaws. J Prosthet Dent. 67:656–660. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gonçalves PF, Gurgel BC, Pimentel SP,
Sallum EA, Sallum AW, Casati MZ and Nociti FH Jr: Effect of two
different approaches for root decontamination on new cementum
formation following guided tissue regeneration: A histomorphometric
study in dogs. J Periodontal Res. 41:535–540. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hoffmann T, Richter S, Meyle J, Gonzales
JR, Heinz B, Arjomand M, Sculean A, Reich E, Jepsen K, Jepsen S and
Boedeker RH: A randomized clinical multicentre trial comparing
enamel matrix derivative and membrane treatment of buccal class II
furcation involvement in mandibular molars. Part III: Patient
factors and treatment outcome. J Clin Periodontol. 33:575–583.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Needleman IG, Worthington HV,
Giedrys-Leeper E and Tucker RJ: Guided tissue regeneration for
periodontal infra-bony defects. Cochrane Database Syst Rev. Apr
19–2006.Epub ahead of print. CD001724PubMed/NCBI
|
|
19
|
Venezia E, Goldstein M, Boyan BD and
Schwartz Z: The use of enamel matrix derivative in the treatment of
periodontal defects: A literature review and meta-analysis. Crit
Rev Oral Biol Med. 15:382–402. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kaigler D, Cirelli JA and Giannobile WV:
Growth factor delivery for oral and periodontal tissue engineering.
Expert Opin Drug Deliv. 3:647–662. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Blumenthal NM: A clinical comparison of
collagen membranes with e-PTFE membranes in the treatment of human
mandibular buccal class II furcation defects. J Periodontol.
64:925–933. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Y, Zheng Y, Ding G, Fang D, Zhang C,
Bartold PM, Gronthos S, Shi S and Wang S: Periodontal ligament stem
cell-mediated treatment for periodontitis in miniature swine. Stem
Cells. 26:1065–1073. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Seo BM, Miura M, Gronthos S, Bartold PM,
Batouli S, Brahim J, Young M, Robey PG, Wang CY and Shi S:
Investigation of multi-potent postnatal stem cells from human
periodontal ligament. Lancet. 364:149–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu M, Kohan E, Bradley J, Hedrick M,
Benhaim P and Zuk P: The effect of age on osteogenic, adipogenic
and proliferative potential of female adipose-derived stem cells. J
Tissue Eng Regen Med. 3:290–301. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aldahmash A, Zaher W, Al-Nbaheen M and
Kassem M: Human stromal (mesenchymal) stem cells: Basic biology and
current clinical use for tissue regeneration. Ann Saudi Med.
32:68–77. 2012.
|
|
26
|
Chen FP, Hu CH and Wang KC: Estrogen
modulates osteogenic activity and estrogen receptor mRNA in
mesenchymal stem cells of women. Climacteric. 16:154–160. 2013.
View Article : Google Scholar
|
|
27
|
Morishita M, Yamamura T, Bachchu MA,
Shimazu A and Iwamoto Y: The effects of oestrogen on osteocalcin
production by human periodontal ligament cells. Arch Oral Biol.
43:329–333. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Morishita M, Yamamura T, Shimazu A,
Bachchu AH and Iwamoto Y: Estradiol enhances the production of
mineralized nodules by human periodontal ligament cells. J Clin
Periodontol. 26:748–751. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mamalis A, Markopoulou C, Lagou A and
Vrotsos I: Oestrogen regulates proliferation, osteoblastic
differentiation, collagen synthesis and periostin gene expression
in human periodontal ligament cells through oestrogen receptor
beta. Arch Oral Biol. 56:446–455. 2011. View Article : Google Scholar
|
|
30
|
Liang L, Yu JF, Wang Y, Wang G and Ding Y:
Effect of estrogen receptor beta on the osteoblastic
differentiation function of human periodontal ligament cells. Arch
Oral Biol. 53:553–557. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cao M, Shu L, Li J, Su J, Zhang W, Wang Q,
Guo T and Ding Y: The expression of estrogen receptors and the
effects of estrogen on human periodontal ligament cells. Methods
Find Exp Clin Pharmacol. 29:329–335. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang B, Li Y, Zhou Q and Ding Y: Estrogen
deficiency leads to impaired osteogenic differentiation of
periodontal ligament stem cells in rats. Tohoku J Exp Med.
223:177–186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liao S, Wang W, Uo M, Ohkawa S, Akasaka T,
Tamura K, Cui F and Watari F: A three-layered nano-carbonated
hydroxyapatite/collagen/PLGA composite membrane for guided tissue
regeneration. Biomaterials. 26:7564–7571. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu HC, e LL, Wang DS, Su F, Wu X, Shi ZP,
Lv Y and Wang JZ: Reconstruction of alveolar bone defects using
bone morphogenetic protein 2 mediated rabbit dental pulp stem cells
seeded on nano-hydroxyapatite/collagen/poly(L-lactide). Tissue Eng
Part A. 17:2417–2433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu SJ, Liu HC, Ling-Ling E, Wang DS and
Zhu GX: Proliferation and differentiation of osteoblasts from the
mandible of osteoporotic rats. Exp Biol Med (Maywood). 237:395–406.
2012. View Article : Google Scholar
|
|
36
|
Gay IC, Chen S and MacDougall M: Isolation
and characterization of multipotent human periodontal ligament stem
cells. Orthod Craniofac Res. 10:149–160. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zheng W, Wang S, Ma D, Tang L, Duan Y and
Jin Y: Loss of proliferation and differentiation capacity of aged
human periodontal ligament stem cells and rejuvenation by exposure
to the young extrinsic environment. Tissue Eng Part A.
15:2363–2371. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
E LL, Xu LL, Wu X, Wang DS, Lv Y, Wang JZ
and Liu HC: The interactions between rat-adipose-derived stromal
cells, recombinant human bone morphogenetic protein-2, and
beta-tricalcium phosphate play an important role in bone tissue
engineering. Tissue Eng Part A. 16:2927–2940. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Barrilleaux B, Phinney DG, Prockop DJ and
O'Connor KC: Review: Ex vivo engineering of living tissues with
adult stem cells. Tissue Eng. 12:3007–3019. 2006. View Article : Google Scholar
|
|
40
|
Caplan AI: Adult mesenchymal stem cells
for tissue engineering versus regenerative medicine. J Cell
Physiol. 213:341–347. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Eberli D and Atala A: Tissue engineering
using adult stem cells. Methods Enzymol. 420:287–302. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chang PC, Chien LY, Chong LY, Kuo YP and
Hsiao JK: Glycated matrix up-regulates inflammatory signaling
similarly to Porphyromonas gingivalis lipopolysaccharide. J
Periodontal Res. 48:184–193. 2013. View Article : Google Scholar
|
|
43
|
Fan JZ, Wang Y, Meng Y, Li GW, Chang SX,
Nian H and Liang YJ: Panax notoginseng saponins mitigate
ovariectomy-induced bone loss and inhibit marrow adiposity in rats.
Menopause. 22:1343–1350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee JH, Baek HR, Lee KM, Zheng GB, Shin SJ
and Shim HJ: Effects of ovariectomy and corticosteroid induced
osteoporosis on the osteoinductivity of rhBMP-2 in a segmental
long-bone defect model. Tissue Eng Part A. 21:2262–2271. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fang J, Yang L, Zhang R, Zhu X and Wang P:
Are there differences between Sprague-Dawley and Wistar rats in
long-term effects of ovariectomy as a model for postmenopausal
osteoporosis? Int J Clin Exp Pathol. 8:1491–1502. 2015.PubMed/NCBI
|
|
46
|
Liu Y, Wang L, Liu S, Liu D, Chen C, Xu X,
Chen X and Shi S: Transplantation of SHED prevents bone loss in the
early phase of ovariectomy-induced osteoporosis. J Dent Res.
93:1124–1132. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bressan E, Ferroni L, Gardin C, Pinton P,
Stellini E, Botticelli D, Sivolella S and Zavan B: Donor
age-related biological properties of human dental pulp stem cells
change in nanostructured scaffolds. PLoS One. 7:e491462012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sato C, Iso Y, Mizukami T, Otabe K, Sasai
M, Kurata M, Sanbe T, Sekiya I, Miyazaki A and Suzuki H: Fibroblast
growth factor-23 induces cellular senescence in human mesenchymal
stem cells from skeletal muscle. Biochem Biophys Res Commun.
470:657–662. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hong CJ, Park H and Yu SW: Autophagy for
the quality control of adult hippocampal neural stem cells. Brain
Res. March 9–2016.Epub ahead of print. View Article : Google Scholar
|
|
50
|
Takashima Y, Terada M, Udono M, Miura S,
Yamamoto J and Suzuki A: Suppression of let-7b and miR-125a/b
maturation by Lin28b enables maintenance of stem cell properties in
hepatoblasts. Hepatology. March 17–2016.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Larijani B, Arjmand B, Ahmadbeigi N,
Falahzadeh K, Soleimani M, Sayahpour FA and Aghayan HR: A simple
and cost-effective method for isolation and expansion of human
fetal pancreas derived mesenchymal stem cells. Arch Iran Med.
18:770–775. 2015.PubMed/NCBI
|
|
52
|
Tong L, Zhou J, Rong L, Seeley EJ, Pan J,
Zhu X, Liu J, Wang Q, Tang X, Qu J, et al: Fibroblast growth
factor-10 (FGF-10) mobilizes lung-resident mesenchymal stem cells
and protects against acute lung injury. Sci Rep. 6:216422016.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Drowley L, Koonce C, Peel S, Jonebring A,
Plowright AT, Kattman SJ, Andersson H, Anson B, Swanson BJ, Wang QD
and Brolen G: Human induced pluripotent stem cell-derived cardiac
progenitor cells in phenotypic screening: A transforming growth
factor-β type 1 receptor kinase inhibitor induces efficient cardiac
differentiation. Stem Cells Transl Med. 5:164–174. 2016. View Article : Google Scholar
|
|
54
|
Tuganbekova S, Gaipov A, Turebekov Z,
Saparbayev S, Shaimardanova G, Popova N, Taubaldiyeva Z,
Serebrennikova D and Trimova R: Fetal renal stem cell transplant in
nephrotic and nonnephrotic glomerulonephritis with stage 2–4
chronic kidney disease: Potential effect on proteinuria and
glomerular filtration. rate. 13(Suppl 3): 156–159. 2015.
|