Introduction
The number of obese individuals has rapidly
increased in several countries due to changes in lifestyle and/or
food consumption. Obesity is a serious disease that can shorten the
lifespan of an individual by inducing insulin resistance, type 2
diabetes, dyslipidemia and atherosclerosis (1–3).
Obesity is often defined as the excessive accumulation of visceral
adipose tissue. In general, in the human body, if the energy intake
exceeds the energy consumption, most of this excess energy will be
accumulated as visceral adipocytes (particularly in white adipose
tissue) in the form of triglycerides (TG) and cholesterol (CE)
(4).
It is well known that a number of factors are
involved in obesity (1–3). In adipose, hepatic and muscle
tissues, TG and CE accumulate in the form of lipid droplets. The
shape and fatty acid composition of these lipid droplets has become
a focus of research (5,6). The droplets may be closely related
to obesity or insulin resistance (7–10).
Many lipid droplets that have grown large in size can be seen in
the white adipocytes of both humans and animal models with obesity.
These lipid droplets in adipocytes have a core that is composed of
TG and CE esters and is coated by a phospholipid monolayer
associated with lipid droplet proteins, such as perilipin (PLIN)
(5). Earlier research revealed
that these proteins and the composition of these lipid droplets,
defined as the desaturation index, which is described by the ratio
of monounsaturated fatty acids (MUFAs) to saturated fatty acids
(SFAs), plays a role in obesity which is the result of a high-fat
diet (11,12).
The major MUFAs oleate (C18:1n-9) and palmitoleate
(C16:1n-7) are the main unsaturated components of intracellular
lipid droplets within tissues which are responsible for adipose,
hepatic and muscle lipogenesis. These MUFAs are transformed from
SFAs, i.e., stearate (C18) and palmitate (C16), by stearoyl-CoA
desaturase (SCD). It has become clear that SCD is closely
associated with obesity and insulin resistance through the
regulation of lipogenesis and lipid oxidation (10,13). There are also some enzymes that
regulate lipogenesis. Phosphorylated (p-)AMP kinase (p-AMPK)
inhibits the activity of fatty acid synthase (FAS) to reduce the
synthesis of SFA from malonyl-CoA and acetyl-CoA (14–16). On the other hand, p-AMPK
accelerates the phosphorylation of acetyl-CoA carboxylase (ACC) to
suppress the synthesis of malonyl-CoA from acetyl-CoA (14,16,17).
Lipolysis is also affected by several factors
(18). The activity of carnitine
palmitoyltransferase 1 (CPT1), which plays a role in the transport
of fatty acids into the mitochondria, a site where the oxidation of
fatty acids occurs, is suppressed by malonyl-CoA (19). The expression of uncoupling
protein (UCP) accelerates energy consumption through the uncoupling
of oxidative phosphorylation on mitochondrial membranes (18). The inhibition of the expression of
the PLIN gene promotes lipolysis, which is thought to be due to
increased mitochondrial access to fatty acid substrates through an
increase in the surface area of lipid droplets (5).
In an aim to counteract the negative effects of a
high-fat diet, chickpeas (C. arietinum) have been
investigated for their abitity to alter lipid metabolism.
Currently, there are two types of chickpeas, the Kabuli type and
the Desi type. The Desi type has a darker color and smaller seeds,
and they have been cultivated in Xinjiang (China) for over 2,500
years; they have also been used since ancient times in traditional
Uighur medicine as a natural treatment for type 2 diabetes, obesity
and hepatic steatosis (21).
Previously, using a mouse model, we reported that insulin
resistance, type 2 diabetes, dyslipidemia and obesity-associated
atherosclerosis caused by a high-fat diet can be attenuated
improved by the consumption of chickpeas (20). In our previous study, we used the
Desi type of chickpeas. In the present study, we prepared ethanol
extracts of chickpeas (ECP) using the Desi-type chickpeas for in
vitro experiments. We used the mouse 3T3-L1 cell line, which
can differentiate into cells with an adipose-like phenotype, in
order to determine the effects of the chickpeas on fatty acid
metabolism and to elucidate the underlying mechanisms. For
comparison, we also used extracts of Kabuli-type chickpeas for some
of our experiments.
Materials and methods
Materials
Mouse 3T3-L1 pre-adipocytes were obtained from the
Health Science Research Resources Bank (Osaka, Japan). Dulbecco's
modified Eagle's medium (DMEM) with L-glutamine, sodium pyruvate
and non-fat dry milk were obtained from Nacalai Tesque (Kyoto,
Japan). Gentamicin reagent solution (gentamicin sulfate, 10 mg/ml),
trypsin-EDTA (0.25% trypsin, 1 mM EDTA • 4Na, X1), calf serum, the
TRIzol plus RNA purification kit, the superscript VILO cDNA
synthesis kit and Prolong Gold Antifade Reagent were purchased from
Invitrogen (Carlsbad, CA, USA). Primers and SYBR-Green for reverse
transcription-quantitative PCR (RT-qPCR) and Tris-buffered saline
with Tween-20 (TBS-T) tablets, pH 7.6 (100 tablets) were purchased
from Takara Bio Inc. (Shiga, Japan).
Both types of chickpeas were obtained from Jumpsun
Bio-medicine (Shanghai, China). Phosphate-buffered saline (PBS),
chloroform, methanol, 99.5% ethanol, isopropanol, formaldehyde and
undecanoic acid were purchased from Wako Pure Chemical Industries
(Osaka, Japan). Oil Red O was purchased from Sigma-Aldrich (St.
Louis, MO, USA). M-PER® Mammalian Protein Extraction
Reagent and the protein assay kit (BCA method) were obtained from
Pierce/Thermo Scientific (Rockford, IL, USA). AMPKα (23A3) rabbit
monoclonal antibody (mAb; #2603), p-AMPKα (Thr172; 40H9) rabbit mAb
(#2535), ACC antibody (#3662), p-ACC (Ser79) antibody (#3661), PLIN
(D418) antibody (#3470), SCD1 (C12H5) rabbit mAb (#2794), GAPDH
(14C10) rabbit mAb (#2118), liver kinase B1 (LKB1; D60C5) rabbit
mAb (#3047), p-LKB1 (Ser428; C67A3) rabbit mAb (#3482), fatty acid
synthase (FAS) antibody (#3189), horseradish peroxidase
(HRP)-linked anti-rabbit IgG antibody (#7074), and HRP-linked
anti-mouse IgG antibody (#7076) were obtained from Cell Signaling
Technology (Beverly, MA, USA). Anti-RPS18 antibody (HPA050159) was
purchased from Atlas Antibodies (Stockholm, Sweden). CPT1 (H-40)
antibody (sc-98834) and UCP2 (G-6) antibody (sc-390189) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Anti-elongation of very long chain fatty acids family member 6
(ELOVL6) antibody (ab69857) was purchased from Abcam (Cambridge,
MA, USA).
Preparation of ECP
Desi-type chickpeas (50 g) cultivated in Xinjiang,
China were crushed and extracted 3 times with 500 ml of 99.5%
ethanol for 3 h at 60°C, and the filtrate was concentrated to ≤1%
ethanol under reduced pressure at 60°C. Following condensation, the
moisture content was reduced to <30% of the total weight. The
solid content of the extracts was measured by the loss on a drying
test in the Japanese Society of Pharmacopoeia (22), and adjusted to exactly 70% with
water. These extracts were used as ECPs in the present study and
diluted to 0.2–0.1% in culture medium before use. Kabuli-type
chickpea extracts also prepared by the same method.
Cell culture and the differentiation of
3T3-L1 cells into adipocytes
The 3T3-L1 cells were grown in DMEM supplemented
with 10% calf serum and 0.5 µg/ml gentamicin sulfate. For
differentiation, 3T3-L1 confluent pre-adipocytes were treated for 2
days with DMEM supplemented with 10% fetal bovine serum (FBS) in
the presence of adipogenic inducers (0.5 mM methylisobutylxanthine,
10 µM dexamethasone and 5 µg/ml insulin), as
previously described (23,24).
The cells were then maintained in DMEM supplemented with 10% FBS,
0.5 µg/ml gentamicin sulfate and 5 µg/ml insulin for
6 days. The medium was changed every 2 days. The cells were grown
at 37°C in 5% CO2 with 100% humidity.
Oil Red O staining and protein assay
The Oil Red O staining method was used to measure
the cellular neutral lipid droplet accumulation. Following the
culture of 3T3-L1 cells for 8 days for differentiation in the
presence or absence of 0.02–0.1% ECP, the cells were washed twice
with PBS on ice and fixed with 10% formaldehyde for 10 min. The
cells were then washed twice with PBS on ice and immersed in 60%
isopropanol for 1 min and stained with Oil Red O solution
thereafter. The stock solution of the dye (3 mg/ml in isopropanol)
was diluted to 60% with distilled water prior to use and allowed to
stand for 20 min at room temperature.
After staining, the cells were washed with 60%
isopropanol to remove unbound dye and rinsed twice with PBS on ice.
Images were aquired under a 10×40 fluorescence microscope with
bright-field illumination (All-in one fluorescence microscope
BIOREVO BZ-9000; Keyence, Osaka, Japan). For the measurement of
total dye incorporated into cellular lipid droplets, 2 ml of
isopropanol were added to each culture dish after the complete
removal of PBS, and the culture dish was gyrated for 5 min at room
temperature. Subsequently, 300 µl of isopropanol were placed
into each well of a 96-well plate, and the absorbance at 510 nm was
measured with SpectraMax Paradigm® Multi-Mode Microplate
Reader (Molecular Devices, Sunnyvale, CA, USA). The amounts of
proteins were determined using BCA protein assay kits.
Gas chromatographic analysis of lipid
extracts from cells
The sample preparation for the gas chromatography
analysis was performed as previously described (25). Briefly, the cells were washed
twice with PBS on ice, and total cellular lipids were extracted 3
times with chloroform:methanol (2:1, v/v) containing n-undecanoic
acid as an internal standard. The 3 lipid extracts were combined in
a screw-capped glass tube and dried in a heating block under
N2 gas flow at 40°C. Fatty acid methyl esters were
obtained by incubation with 3 M HCl in methanol and extracted with
hexane. Fatty acid methyl esters were identified using a
Hewlett-Packard 5890 gas chromatograph equipped with a 7673 auto
injector and SP-2560 column, 75×0.18 mm ID, 0.14 µm
(Sigma-Aldrich) connected to a flame ionization detector set at
220°C. The column temperature was held at 180°C for 50 min after
the injection. Under these conditions, the 16:0-, 16:1-, 18:0-,
18:1n-9- and 18:1n-7-methyl esters were eluted at 26.6, 29.3, 34.8,
38.4 and 39.0 min, respectively.
Microarrays
A DNA microarray analysis was performed using Gene
SQUARE, a multiple assay DNA microarray metabolic syndrome for mice
(Kurabo Industries, Osaka, Japan) according to the manufacturer's
instructions. Briefly, Alexa Fluor 555-labeled cDNA was prepared
from 10 µg of total RNA from each mouse tissue. Labeled cDNA
was purified and then hybridized at 65°C for 14 h. Following
hybridization, the microarray slide was washed in several types of
washing solution and dried by centrifugation at 450 x g and 20–25°C
for 1 min. The image of the microarray was acquired by a Genepix
4200A microarray scanner (Molecular Devices). The fluorescence
intensities of the scanned images were quantified, corrected for
background noise, and normalized by the intensity of Mus
musculus ribosomal protein S18 (RPS18).
Western blot analysis
The cells grown in 100-mm dishes were washed twice
with 10 ml of PBS and lysed in 1 ml M-PER. The resulting debris was
removed by centrifugation at 15,000 x g for 10 min at 4°C. Equal
amounts of protein (20 µg/well) were applied to Any kD™
Mini-PROTEAN TGX Gels (Bio-Rad Laboratories, Hercules, CA, USA) and
separated by polyacrylamide gel electrophoresis (PAGE). Proteins on
the gel were then transferred to PVDF membranes with an iBlot gel
transfer system (Invitrogen) or a Trans-Blot Turbo Transfer system
(Bio-Rad Laboratories).
After blocking with 5% non-fat dry milk in TBS-T for
1 h at room temperature, the membranes were washed with TBS-T and
incubated with the primary antibody against each protein listed
above (see Materials section) and the secondary goat antibody
against rabbit or mouse IgG conjugated to HRP. Visualization of the
protein bands was performed by the reaction of HRP with SuperSignal
West Dura Extended Duration Substrate (Thermo Fisher Scientific)
and an LAS-3000 luminescent image analyzer (Fujifilm, Tokyo,
Japan). A densitometric analysis of each band was performed with a
Multi Gauge Version 2.1 (Fujifilm).
RR-qPCR
Total RNA was extracted from the 3T3-L1 cells using
a TRIzol Plus RNA purification kit and then reverse transcribed
into cDNA with the use of a SuperScript VILO cDNA synthesis kit.
The abundance of transcripts was assessed by qPCR on a StepOne Plus
(Applied Biosystems by Life Technologies, Foster City, CA, USA)
with a SYBR-Green detection system. For each run, samples were
analyzed in triplicate for both the gene of interest and the Mus
musculus Rps18 gene as an internal control. The expression data
for Scd1 and Rps18 were normalized for the efficiency
of amplification, as determined by a standard curve included on
each run and analyzed using the ΔΔCT method (31). The forward and reverse primers
used were as follows: Scd1, TCTTGTCCCTATAGCCCAATCCAG
(forward) and AGCTCAGAGC GCGTGTTCAA(reverse); and Rps18,
TTCTGGCCAACGGTCTAGACAAC (forward) and CCAGTGGTCTTGGTGTG CTGA
(reverse).
Statistical analysis
The data were subjected to statistical analyses
using SAS Release 9.1 (SAS Institute, Cary, NC, USA). The results
from 3 experiments are presented as the means ± standard deviation
(SD). The statistical analyses were performed using the unpaired
Student's t-test, One-Way ANOVA (Tukey) and the Jonckheere-Terpstra
test. Values of p<0.05 were considered to indicate statistically
significant differences.
Results
Effects of ECP on the growth of the lipid
droplets, and the lipid and PLIN contents
The 3T3-L1 cells cultured with or without ECP at
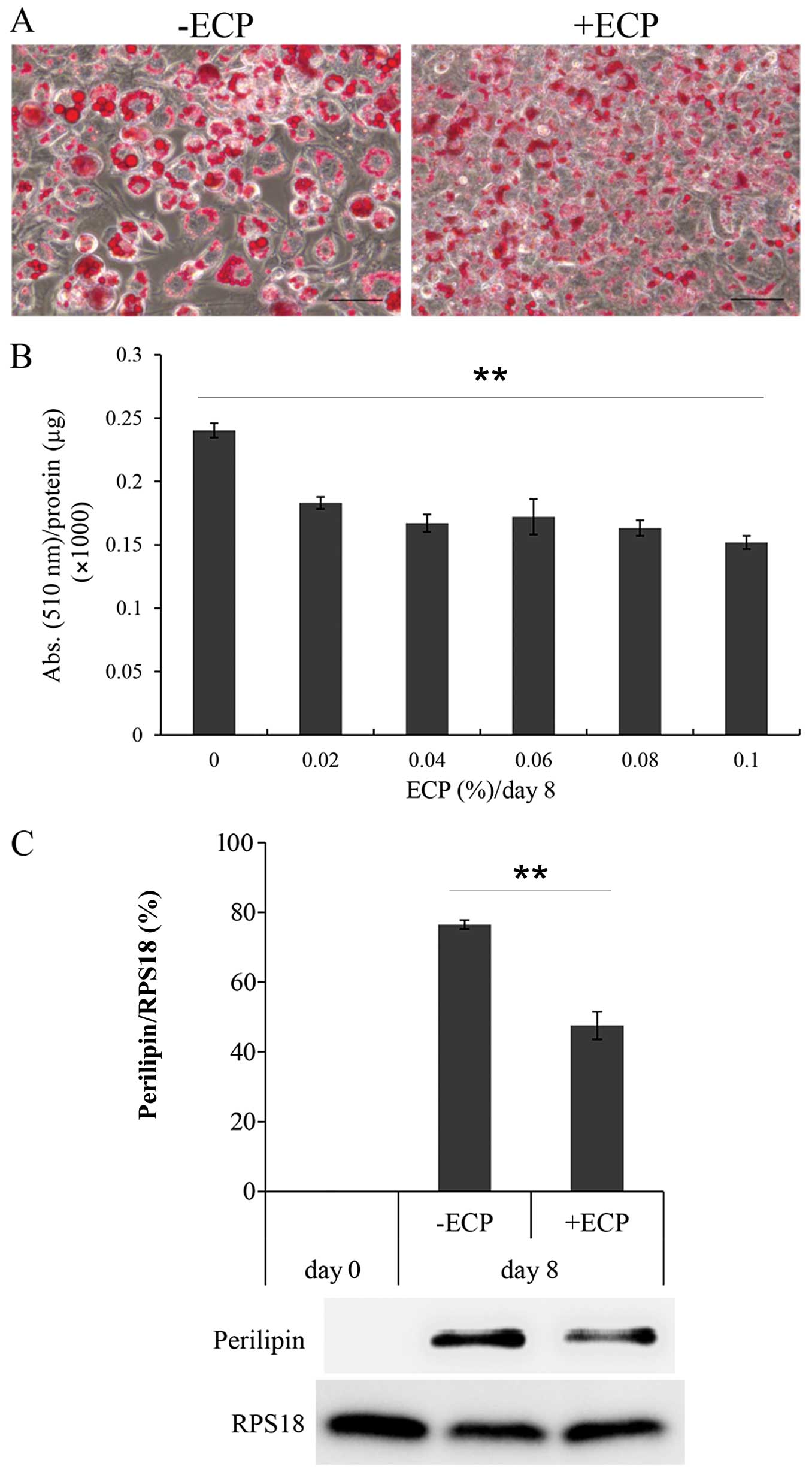
0.1% for 8 days were stained with Oil Red O (Fig. 1A). Large lipid droplets developed
on day 8 when differentiation was completed without ECP. By
contrast, only small droplets were observed in the presence of ECP
at 0.1%. The amount of dye incorporated into the cellular droplets
was then measured as the absorbance at 510 nm/mg protein (Fig. 1B). In the presence of ECP at
0.02–0.1%, the lipid content per mg protein was reduced by 63–76%.
The amount of PLIN was reduced by 63–76% in the presence of ECP at
0.1% (Fig. 1C).
Effects of ECP on the fatty acid contents
and desaturation index of the lipid droplets
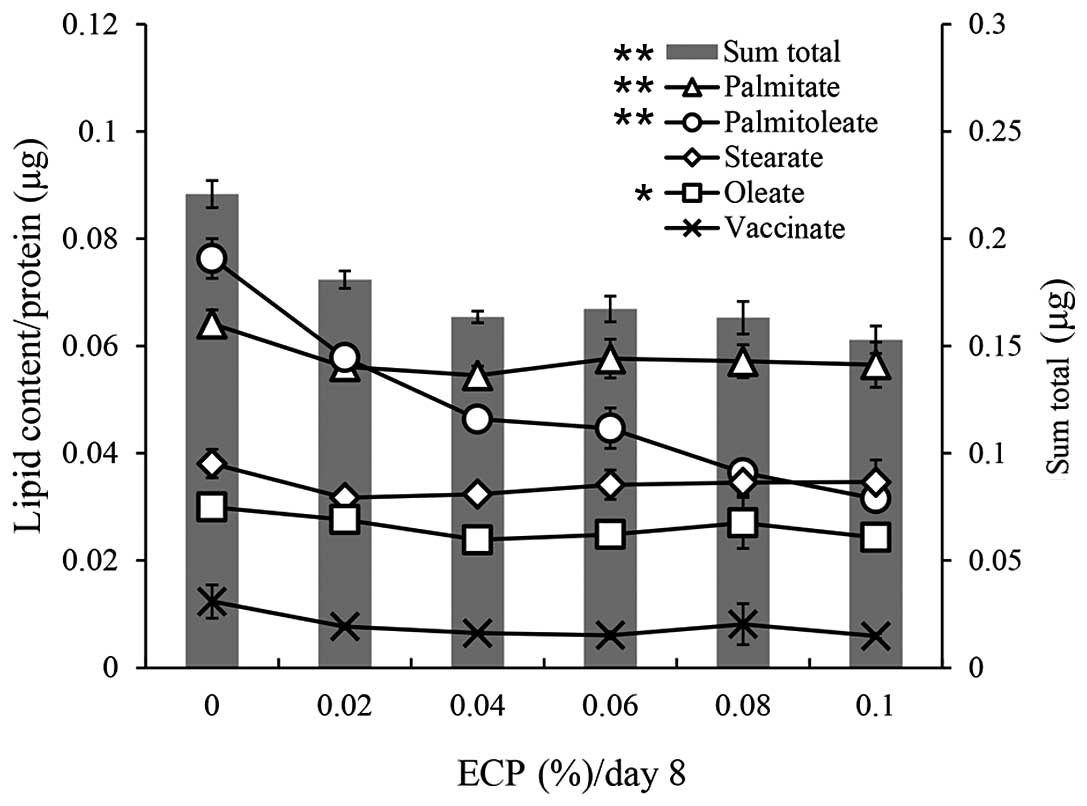
A gas chromatography analysis was performed for
cellular lipid droplets. The contents of each fatty acid and the
sum total of each fatty acid are shown in Fig. 2. The reduction by ECP at 0.02–0.1%
of the sum total (70–81% of sum total without ECP) was similar to
that of the lipid contents measured by the Oil Red O staining
method (Fig. 1A). The contents of
palmitate (C16:0), stearate (C18:0) and oleate (C18:1n-9) were
slightly reduced by ECP at 0.02–0.1%, whereas the contents of
palmitoleate (C16:1) and vaccinate (C18:1n-7) were significantly
reduced by the same ECP content.
The desaturation index values (the MUFA/SFA ratio
for palmitoleate/palmitate, oleate/stearate and vaccenate/stearate)
are shown in Table I. The ratio
of palmitoleate/palmitate was markedly suppressed by the presence
of ECP in a dose-dependent manner, which reflects the significant
reduction in palmitoleate by ECP, as shown in Fig. 2. The ratio of oleate/stearate was
only slightly reduced by treatment with ECP.
 | Table IThe desaturation index values: the
MUFA/SFA ratio for palmitoleate/palmitate, oleate/stearate and
vaccinate/stearate. |
Table I
The desaturation index values: the
MUFA/SFA ratio for palmitoleate/palmitate, oleate/stearate and
vaccinate/stearate.
| ECP (%) |
Palmitoleate/palmitate
C16:1/C16:0 |
Oleate/stearate
C18:1n-9/C18:0 |
Vaccinate/stearate
C18:1n-7/C18:0 |
|---|
| 0 | 1.19±0.08 | 0.79±0.07 | 0.16±0.04 |
| 0.02 | 1.03±0.05 | 0.87±0.09 | 0.13±0.01 |
| 0.04 | 0.85±0.03 | 0.74±0.04 | 0.14±0.01 |
| 0.06 | 0.77±0.08 | 0.73±0.07 | 0.14±0.02 |
| 0.08 | 0.64±0.05 | 0.78±0.15 | 0.22±0.11 |
| 0.10 | 0.56±0.06 | 0.70±0.09 | 0.19±0.02 |
Effects of ECP on the differentiation of
3T3-L1 cells
As the development of lipid droplets is associated
with adipocyte differentiation (26), we examined the effects of 0.1% ECP
on the expression of differentiation marker genes in 3T3-L1 cells
in order to determine whether ECP affects differentiation itself.
We used the Kurabo Gene SQUARE multiple-assay DNA microarray and
focused on the expression levels of the early differentiation
marker genes, CCAAT-enhancer-binding protein (Cebp)β and
Cebpδ, and the expression levels of the late differentiation
marker genes, Cebpα and peroxisome proliferator-activated
receptor γ (PPARγ). We compared the expression levels of
each gene in the presence of ECP at 0.1% with those in the absence
of ECP. For this purpose, we calculated the ratios of the gene
expression levels with or without ECP on days 1–8/those on day 0.
The ratios are shown in Table
II. The expression level of each gene slightly or markedly
increased during the 8-day culture for differentiation in the
absence of ECP. The presence of ECP caused only slight differences
in the ratio (Table II).
| Table IIThe effects of ECP on the expression
of differentiation marker genes. |
Table II
The effects of ECP on the expression
of differentiation marker genes.
| Differentiation
marker | Gene symbol | −CP
| +CP
|
|---|
| Day 0 | Day 1 | Day 2 | Day 4 | Day 6 | Day 8 | Day 1 | Day 2 | Day 4 | Day 6 | Day 8 |
|---|
| Early phase | Cebpβ | 1.0 | 2.2 | 2.8 | 2.5 | 3.1 | 3.3 | 3.1 | 3.8 | 2.9 | 3.1 | 3.8 |
| Cebpδ | 1.0 | 1.0 | 1.0 | 1.2 | 2.6 | 1.0 | 0.9 | 0.9 | 1.4 | 1.9 | 1.1 |
| Late phase | Cebpα | 1.0 | 1.3 | 3.0 | 4.2 | 8.7 | 5.9 | 1.3 | 3.2 | 4.9 | 6.5 | 8.8 |
| Pparγ | 1.0 | 1.0 | 2.1 | 12.0 | 11.2 | 9.9 | 1.0 | 4.5 | 9.7 | 10.8 | 13.2 |
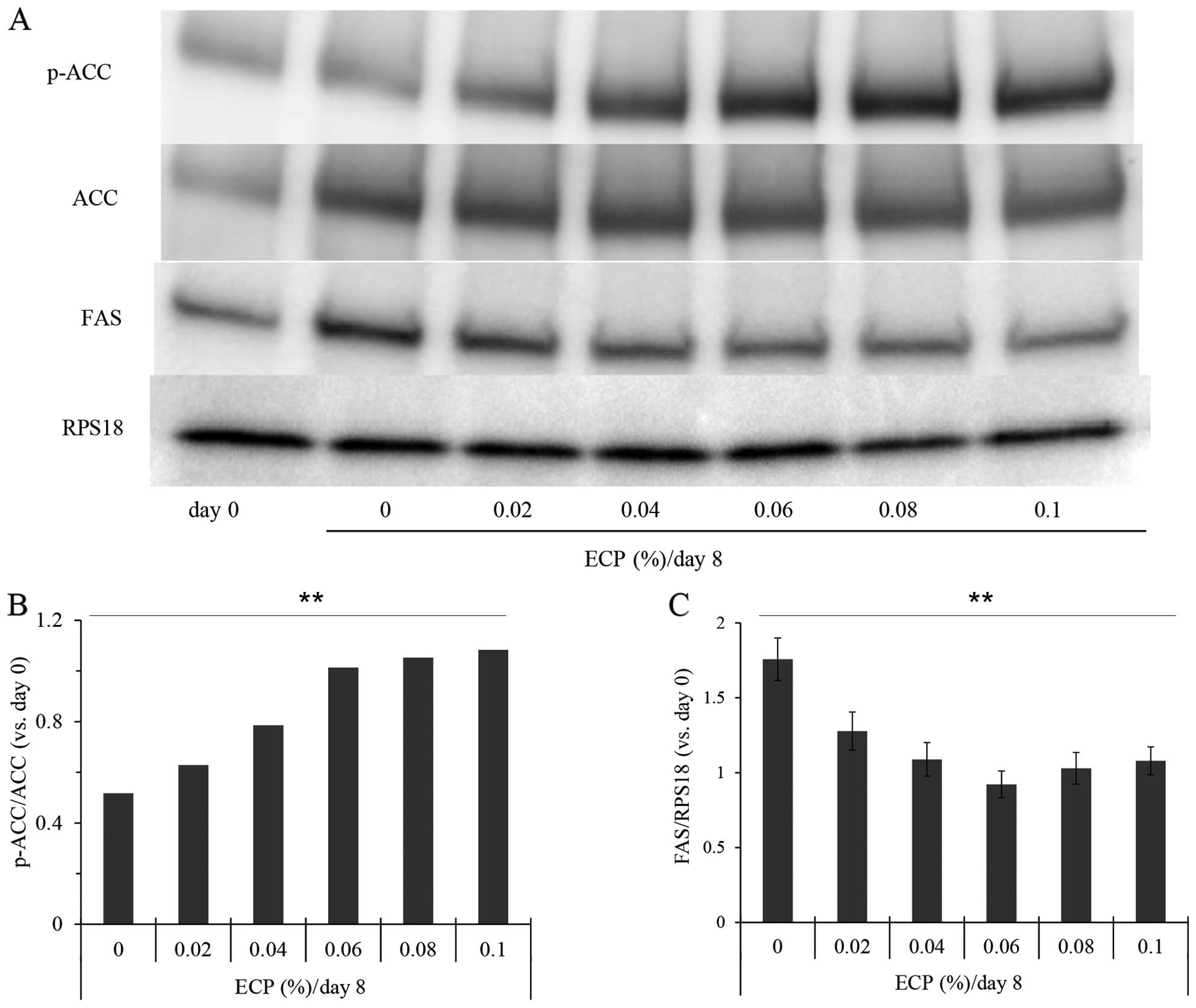
Effects of ECP on SCD1 and ELOVL6 and on
the expression of the Scd1 gene
Since SCD catalyzes the conversion from saturated
fatty acids to unsaturated acids as a key enzyme of lipogenesis,
this enzyme has been suggested to be involved in alterations of the
desaturation index (27,28). We therefore examined the amount of
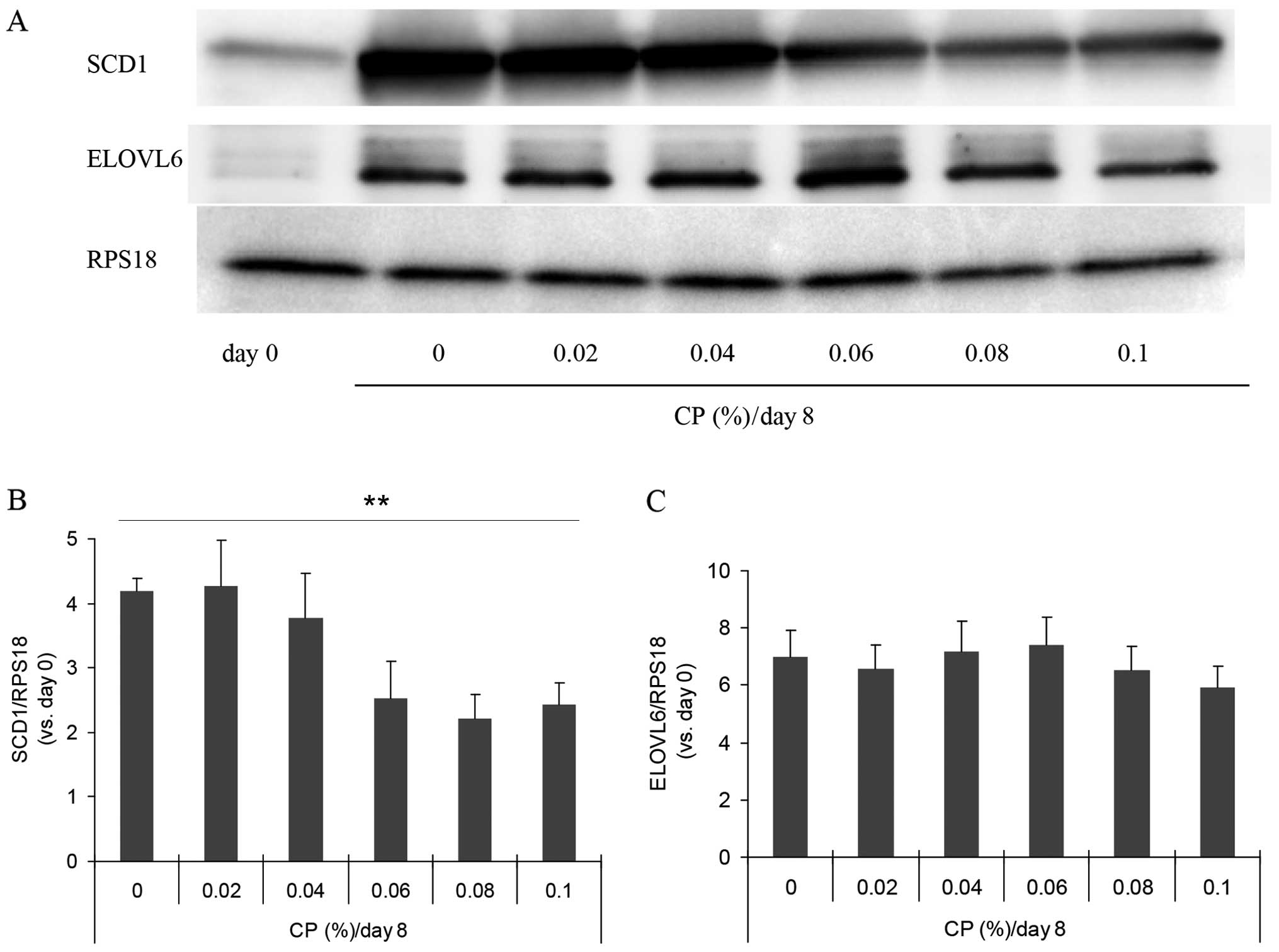
SCD1 protein by western blot analysis (Fig. 3A). The amount was increased by
approximatelt 4-fold during culture for differentiation for 8 days,
and this increase was suppressed in the presence of ECP at
0.06–0.1% (Fig. 3B). In all
western blot analyses, we used RPS18 as the internal standard and
observed no substantial alterations in the amount of this protein
during differentiation.
By contrast, the amounts of GAPDH and β-actin
usually used as internal standards were altered during the
differentiation of 3T3-L1 cells (data not shown). The effects of
ECP on the expression of the Scd1 gene were also examined by
RT-qPCR. The expression was suppressed to 89, 75, 64, 73 and 61% by
ECP at 0.02, 0.04, 0.06, 0.08 and 0.1%, respectively (data not
shown). The amount of ELOVL6, a fatty acid elongase that uses
malonyl-CoA as a two-carbon donor in the step of fatty acid
elongation, was not altered by the presence of 0.02–0.1% ECP
(Fig. 3A and C).
Effects of ECP on other enzymes
participating in lipogenesis
As the suppression of Scd1 gene expression
often causes an increase in the phosphorylation of AMPK (p-AMPK)
and ACC (p-ACC) (19,29), and as the phosphorylated form of
LKB1, the upstream kinase in the AMPK cascade, promotes the
phosphorylation of AMPK (30), we
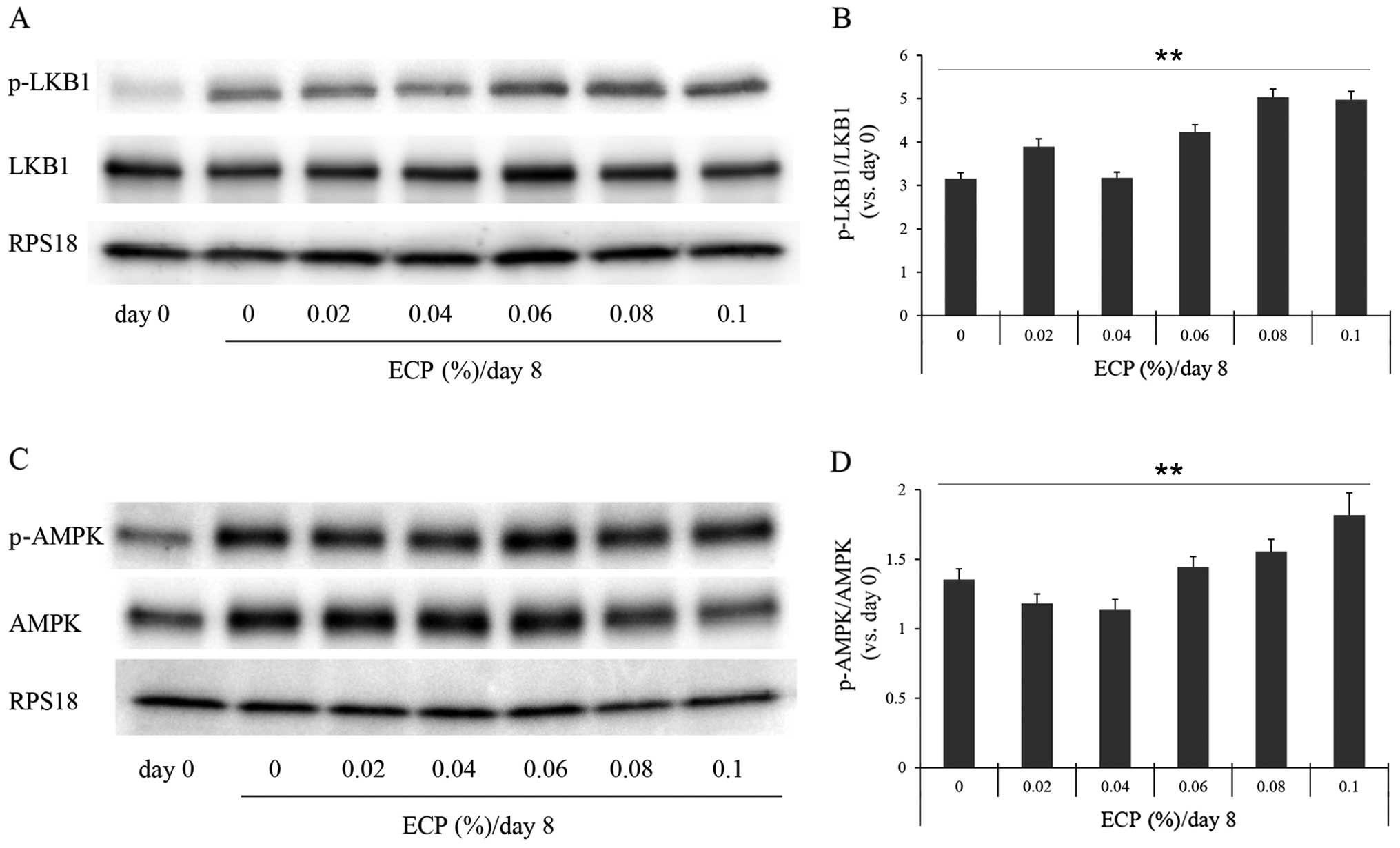
examined the phosphorylation levels of these enzymes by western
blot analysis (Figs. 4 and
5). The phosphorylation levels of
LKB1, AMPK and ACC were significantly increased by the presence of
0.06–0.1% ECP (Figs. 4 and
5A and B). The phosphorylation
levels of LKB1 and AMPK were not significantly altered by ECP at
0.02 or 0.04% (Fig. 4). As it has
been reported that the expression of FAS is suppressed by the
phosphorylated form (31) and
that FAS promotes a reaction, producing SFA from malonyl-CoA, we
also examined the effects of ECP on the level of FAS. The amount of
FAS was reduced by the presence of 0.02–0.1% ECP (Fig. 5A and C).
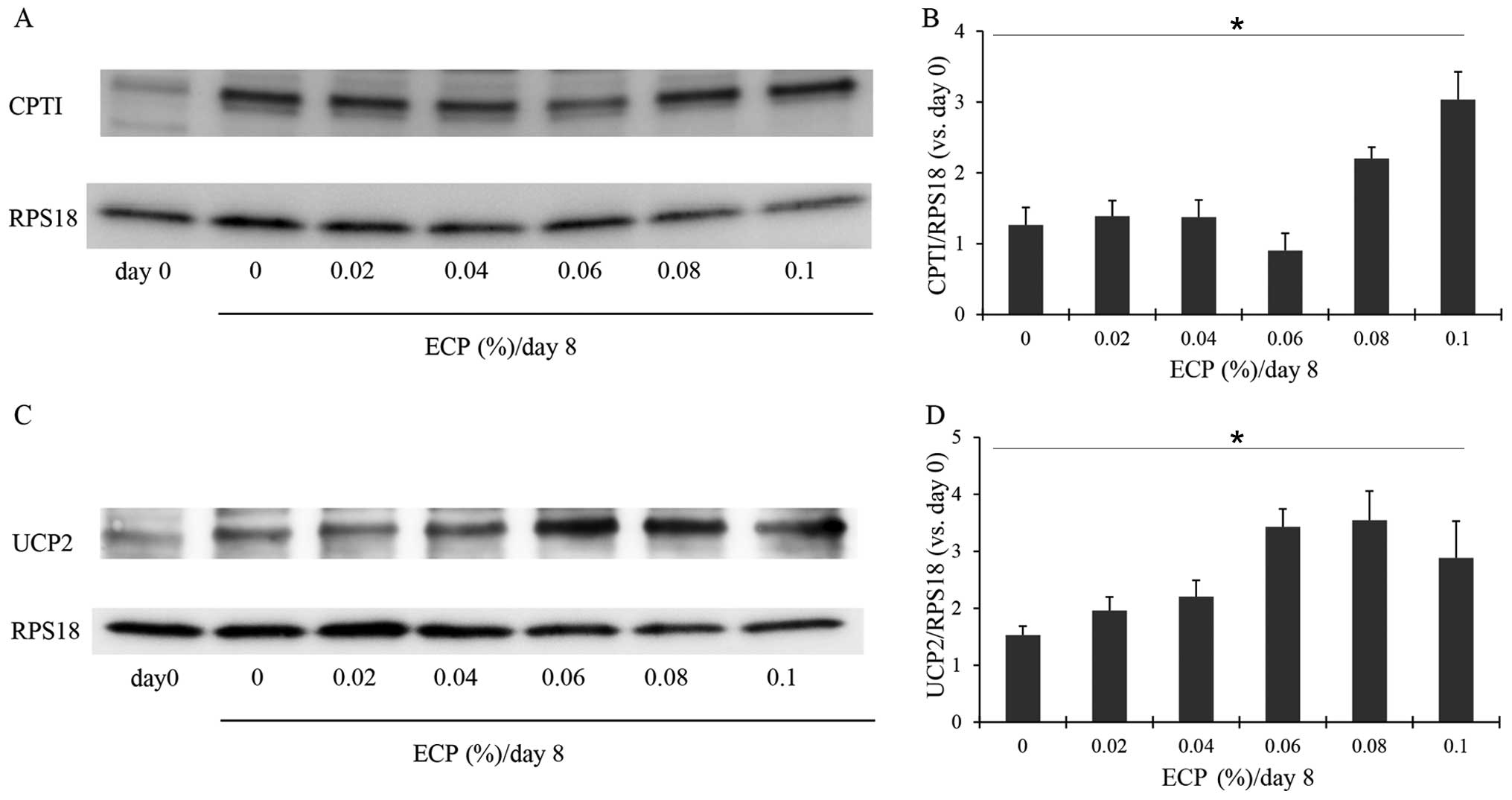
Effects of ECP on enzymes participating
in lipolysis
CPT1 is an enzyme that is important for lipid
metabolism. It promotes the transport of long-chain fatty acids
into the mitochondria (32). We
thus examined the changes in the amount of CPT1 in the 3T3-L1
cells. On day 0, the 3T3-L1 cells had only a small amount of CPT1,
but on day 8 when differentiation was complete, the cells exhibited
an increased amount of CPT1 (Fig. 6A
and B). The presence of 0.08–0.1% ECP caused a significant
increase in the amount of CPT1 (Fig.
6A and B), although ECP at 0.02–0.06% almost had no effect on
the levels of CPT1. Mitochondrial UCP2, located mainly in brown fat
cells, uncouples oxidative phosphorylation and is responsible for
thermogenesis without the production of ATP. This enzyme has been
suggested to promote the oxidation of fatty acids (33). Hence the effects of ECP on UCP2
were examined in this study. ECP at 0.02–0.1% increased the amount
of UCP2 (Fig. 6C and D).
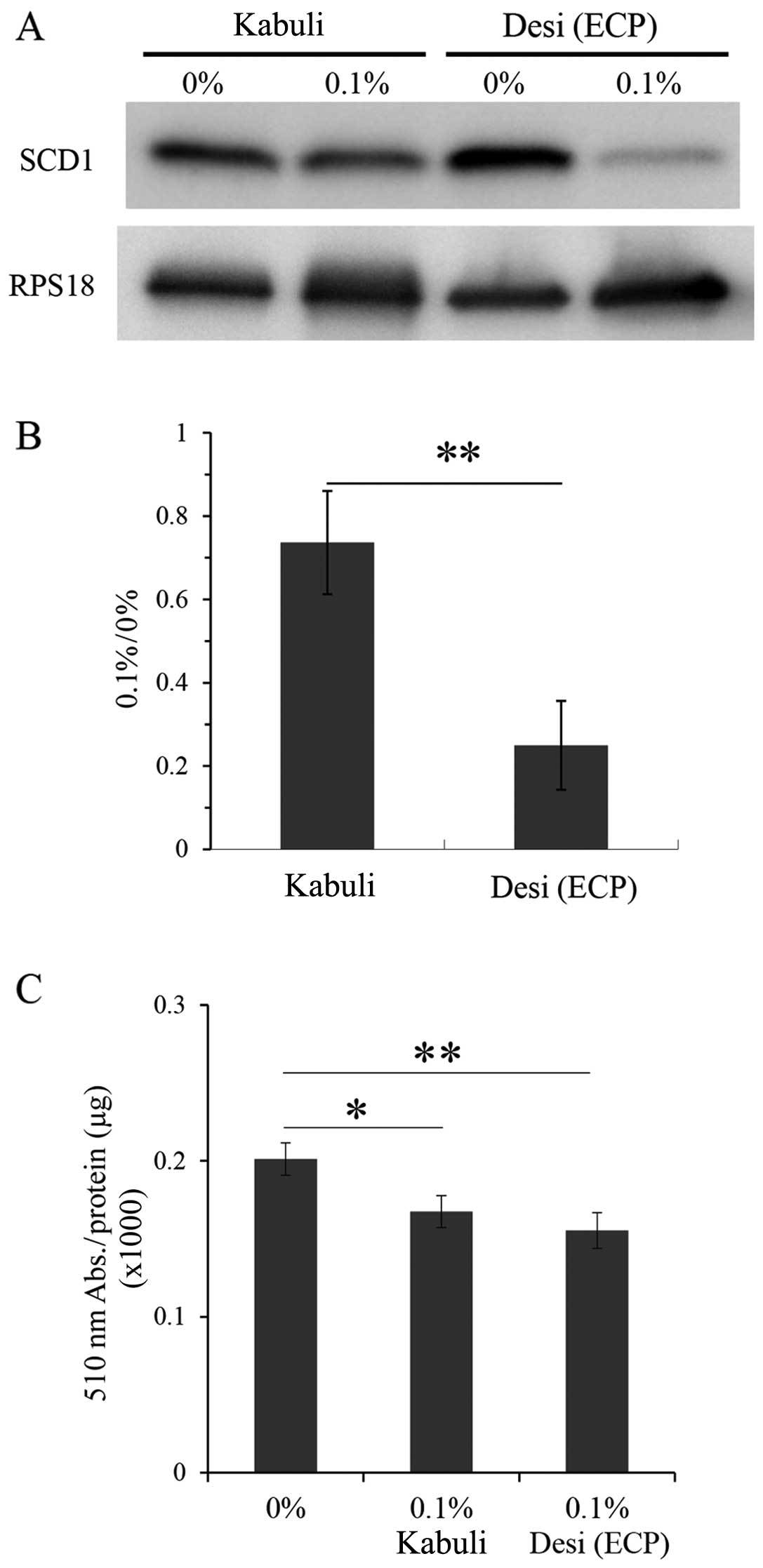
Effects of ethanol extracts of
Kabuli-type chickpeas on SCD1 and lipid contents
We also prepared ethanol extracts of the Kabuli-type
chickpeas using the same method as that used for the Desi-type
chickpeas, as described in the Materials and methods, and we
examined the effects of these extracts on CPT1 and lipid contents
(Fig. 7). The results of western
blot analysis revealed that the ethanol extracts of the Kabuli-type
chicikpeas had less prominent effects on SCD1 expression, as
compared to the Deci-type ECP (Fig.
7A and B). The lipid contents were reduced by the ethanol
extracts of the Kabuli-type chickpeas at 0.1% (Fig. 7C). However, this reduction was
less prominent than that induced by the Deci-type ECP (Fig. 7C).
Discussion
Kabuli-type chickpeas are presently grown in many
temperate-zone countries, and the smaller, darker Desi type is
grown only in Xinjiang, China and a few areas in India, in the
semi-arid tropics. In a previous study, we used the Desi-type
chickpeas obtained from Xinjiang Uighur and also used a mouse model
of diabetes, and our findings indicated that several dysfunctions
associated with obesity were improved by the Desi-type chickpeas
(20). The results of the present
study demonstrated that ethanol extracts of Desi-type chickpeas
affected fatty acid metabolism in 3T3-L1 cells, indicating that the
effects of Desi-type chickpeas observed in our previous mouse model
may be attributed at least in part to fatty acid metabolism at the
cellular level.
We also used the Kabuli type, which had significant,
but yet limited effects on cellular lipid contents and the amount
of SCD1 (Fig. 7). It is thus
possible that a long period of geographic isolation may have caused
differences in the quantity and/or quality of the effects of the
two types of chickpeas on obesity and fatty acid metabolism. The
differences in the effects of the two types of chickpeas could also
explain the reason why Desi-type chickpeas have been used as
traditional Uighur medicine for patients with type 2 diabetes,
obesity and hepatic steatosis (21). There are studies demonstrating
that a diet containing chickpeas leads to small improvements in
glucose tolerance, the serum lipids profile and/or
hypercholesterolemia, although the studies do not describe the type
of chickpeas used (32–36).
In the present study, in the presence of adipogenic
inducers for 8 days, the 3T3-L1 cells differentiated into
adipocytes. During this period, it was established that there is an
induction of the expression of differentiation marker genes
(37–39). As shown in Table II, the expression levels of early
differentiation marker genes (Cebpβ and Cebpδ) and
late differentiation marker genes (Cebpα and PPARγ)
were increased during culture. ECP did not significantly alter the
expression levels of these genes (Table II), indicating that ECP did not
influence 3T3-L1 cell differentiation itself.
Our microscopic analysis revealed that there was a
large number of ECP-treated cells with small-sized intracellular
lipid droplets, whereas large lipid droplets were found inside
normally differentiated adipocytes (Fig. 1A). As reported previously, small
multilocular lipid droplets produced in the endoplasmic reticulum
grow into unilocular lipid droplets in the cytoplasm by fusing
(8,9,40).
During this enlargement process, it seems to be essential for lipid
droplet coat proteins such as PLIN and FSP27 to coat the surface of
the lipid droplets (9). In the
present study, we observed that the amount of PLIN was
significantly decreased by ECP (Fig.
1C and D). It is thus possible that a decrease in the amount of
PLIN interferes with the growth of large unilocular droplets. Of
note, smaller forms of droplets have been reported to allow for a
larger total surface, which increases the mitochondrial access to
fatty acid substrates, thereby promoting lipid oxidation (9). However, we have no data on lipid
oxidation at present.
It is well known that there is a key enzyme that
controls lipid metabolism (41).
As illustrated in Fig. 8, SCD is
an enzyme that converts SFAs, i.e., palmitic acid (C16:0) and
stearic acid (C18:0), to produce MUFAs, i.e., palmitoleic acid
(C16:1) and oleic acid (C18:1n-9), respectively. Under conditions
in which the expression of Scd1 is suppressed, the
desaturation index (MUFAs/FSA) is decreased (42). The genetic expression of SCD1, one
of four isoforms of this enzyme, is regulated by insulin and leptin
(43–45). An enhanced expression of SCD1
leads to the accumulation of fat in adipose, hepatic and muscle
tissue (43,44,46). The deletion of Scd1 in mice
causes resistance to high-fat diet-induced obesity and glucose
intolerance (42).
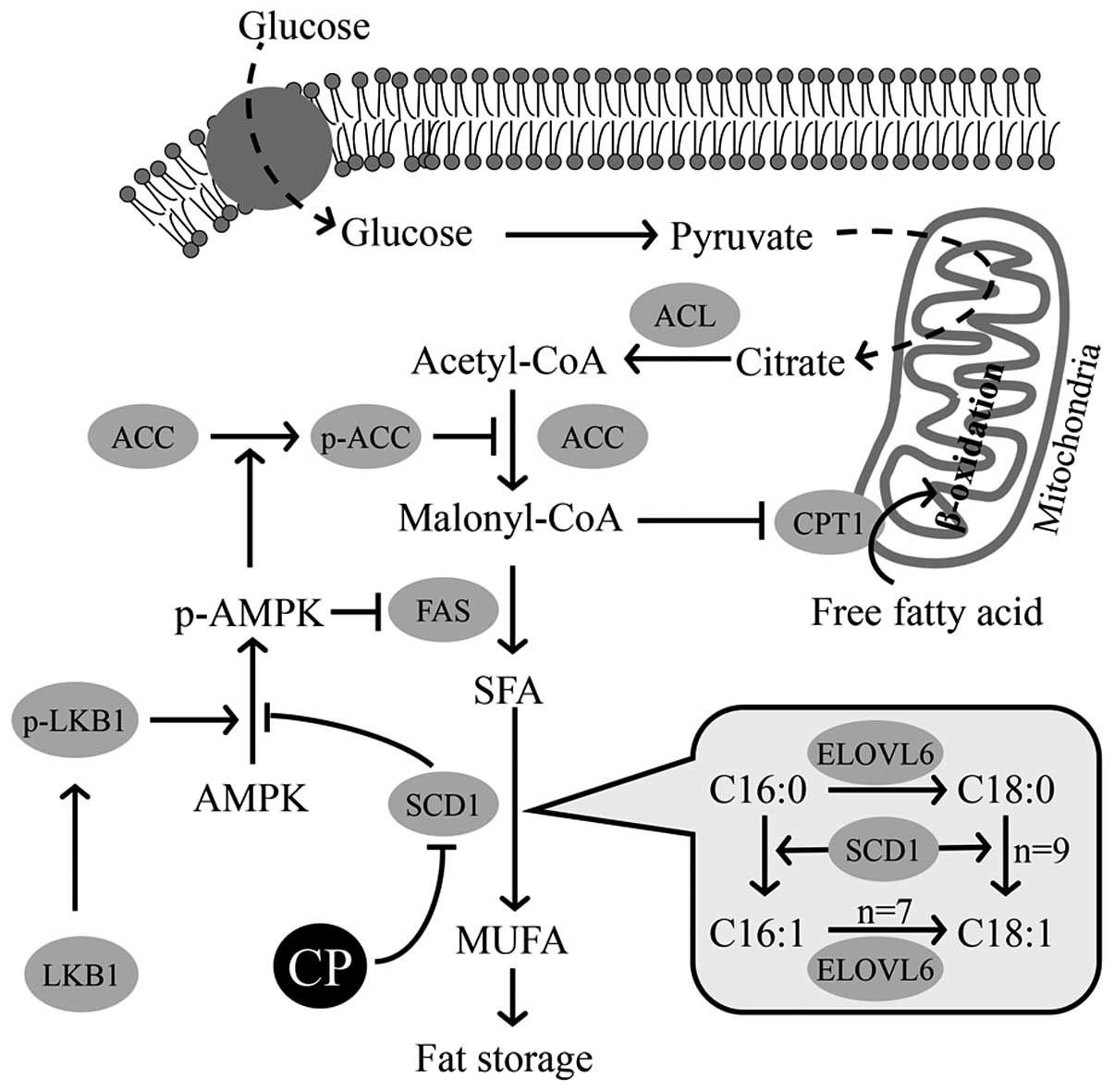 | Figure 8Hypothetical regulation of
lipogenesis and lipolysis by stearoyl-CoA desaturase 1 (SCD1) and
other enzymes. The hypothetical regulation of lipogenesis and
lipolysis by extracts of chickpeas (ECP), based on our findings in
this study. Three enzymes, acetyl-CoA carboxylase (ACC), fatty acid
synthase (FAS) and SCD1, participating in lipogenesis in each step
are described. SCD1 promotes the desaturation of saturated fatty
acid (SFA) of liver kinase B1 (LKB1), causing an increase in the
phosphorylation of AMP activated protein kinase (AMPK). The
phosphorylated form of AMPK then promotes the phosphorylation of
ACC to inactivate ACC, thereby suppressing the reaction from
acetyl-CoA to malonyl-CoA. The depletion of malonyl-CoA simulates
the transportation of fatty acids into the mitochondria by
carnitine palmitoyltransferase 1 (CPT1). SCD1 itself catalyzes the
reaction from SFA (16:0, 18:0) to MUFA (16:1, 18:1), as shown in
the balloon. p-AMPK, phospho-AMPK; p-ACC, phospho-ACC; ACL, ATP
citrate lyase; ELOVL6, elongation of very long chain fatty acids
family member 6; MUFA, monounsaturated fatty acid. |
As shown in Fig.
2, the contents of SFAs (C16:0, C18:0) were not markedly
affected by ECP, whereas those of MUFAs (C16:1, C18:1n-7, C18:1n-9)
were decreased by ECP. These results are well reflected by the
gradual decrease in the desaturation index with the increasing ECP
concentration (Table I).
Simultaneously, ECP suppressed the gene expression of Scd1
and the amount of SCD1 (Fig. 3A and
B), indicating that the suppression may lead to a decrease in
the desaturation index. On the other hand, there no effect of ECP
on ELOVL6 was observed, which catalyzes the conversion of C16:0 or
C16:1 to C18:0 or C18:1 (Fig. 3A and
C). These results may explain the decreases in the lipid
contents and desaturation index in the ECP-treated cells. Of note,
the desaturation index has been reported to have a proportional
association with human plasma triglycerides (47) and Scd1 mRNA (42).
As schematically depicted in Fig. 8, we suggest that the
phosphorylation of AMPK is induced by a phosphorylated form of
LKB1, which leads to the suppression in the production of SFA from
malonyl-CoA by FAS and to the suppression of the production of
malonyl-CoA from Acetyl-CoA by ACC (19,29,41). ECP promoted the phosphorylation of
LKB1, AMPK and ACC and reduced the amount of FAS (Figs. 4 and 5). The administration of ECP thus
resulted in a series of these changes, as well as in decreased
levels of SCD1 to reduce lipogenesis, although it is not yet clear
whether each enzyme is directly or indirectly affected by ECP.
It has been reported that the expression of CPT1, an
enzyme facilitating the transportation of fatty acids into the
mitochondria, is regulated by the amount of malonyl-CoA and that
the expression has a positive correlation with β-oxidation in the
mitochondria (48,49). As suggested above, malonyl-CoA can
be depleted to a certain extent by the inhibitory effects of ECP on
lipogenesis. It is therefore possible that the increased amount of
CPT1 shown in Fig. 6A and B is
due to decreased levels of malonyl-CoA and that β-oxidation is
promoted. On the other hand, as UCP2 promotes the consumption of
fatty acids without the formation of ATP (50,51), an increase caused by ECP in the
amount of UCP2 may result in the consumption. These two results
suggest that lipolysis may be promoted by ECP.
In the present study, ethanol extracts of chickpeas,
particularly those of the Desi type (which has been used as food
for the control of obesity in a restricted region) were shown to
have effects on several steps in lipid metabolism in an in
vitro system. These effects were suppressive effects on the
storage of fat. The suppression of each step in lipogenesis may
result in a reduction in fat storage (29). The accumulation of droplets with
small sizes and increased amounts of CPT1 and UCP2 may facilitate
the consumption of fat. However, the mechanisms underlying these
broad effects of ECP on lipid metabolism are not clear at
present.
As previously reported (41,52), the regulation of SCD1 has become
an interesting focus of attention as a potential treatment for the
prevention of metabolic syndrome due to diet-induced obesity,
hepatic steatosis and insulin resistance. SCD1 may be a key factor
in lipid metabolism, and its reduction may result in the promoted
phosphorylation of LKB1 (Fig. 4A and
B). As previously reported (29), the phosphorylation of ACC may
reduce the amount of malonyl-CoA and may lead to an increase in the
amount of CPT1 (Fig. 5B). All of
these findings suggest that at least a part of the effectiveness of
ECP against visceral adiposity, dyslipidemia and insulin resistance
that has been shown in mice (20)
can be explained by the effects of ECP on lipid metabolism shown in
the present experiments.
We have almost no information about the molecular
basis of the effects of the ethanol extracts. However, we observed
slight effects with the water soluble fraction and some effects
with the chloroform soluble fraction (data not shown), after the
ECP was shaken with chloroform. Several types of fatty acids and
unsaturated fatty acids, including linoleic acid are found in the
chloroform soluble fraction on a thin layer chromatograph. Of note,
conjugated linoleic acid has been reported to have suppressive
effects on SCD1 (53,54). We have not performed an experiment
to determine whether linoleic acid isolated from the ethanol
extracts exerts effects on SCD1 levels. However, it is possible
that unsaturated fatty acids such as linoleic acid participate in
the effects of ECP.
Thus, it can be concluded that some contents of
Desi-type of chickpeas may alter lipid metabolism, reducing the
lipid content, and these chickpease may prove to be effective in
the prevention of diabetes.
Acknowledgments
We would like to thank E. Ueno and J. Kako at Otsuka
Pharmaceutical Co., Ltd. for providing advice and technical
solutions for this study. We would also like to thank A. Shinohara
and R. Moyer for their assistance with the translation of this
study into English.
References
|
1
|
Flegal KM, Carroll MD, Ogden CL and
Johnson CL: Prevalence and trends in obesity among US adults,
1999–2000. JAMA. 288:1723–1727. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kopelman PG: Obesity as a medical problem.
Nature. 404:635–643. 2000.PubMed/NCBI
|
|
3
|
Kahn BB and Flier JS: Obesity and insulin
resistance. J Clin Invest. 106:473–481. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wajchenberg BL: Subcutaneous and visceral
adipose tissue: their relation to the metabolic syndrome. Endocr
Rev. 21:697–738. 2000. View Article : Google Scholar
|
|
5
|
Brasaemle DL: Thematic review series:
adipocyte biology. The perilipin family of structural lipid droplet
proteins: stabilization of lipid droplets and control of lipolysis.
J Lipid Res. 48:2547–2559. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Londos C, Brasaemle DL, Gruia-Gray J,
Servetnick DA, Schultz CJ, Levin DM and Kimmel AR: Perilipin:
Unique proteins associated with intracellular neutral lipid
droplets in adipocytes and steroidogenic cells. Biochem Soc Trans.
23:611–615. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ducharme NA and Bickel PE: Lipid droplets
in lipogenesis and lipolysis. Endocrinology. 149:942–949. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Puri V and Czech MP: Lipid droplets: FSP27
knockout enhances their sizzle. J Clin Invest. 118:2693–2696.
2008.PubMed/NCBI
|
|
9
|
Nishino N, Tamori Y, Tateya S, Kawaguchi
T, Shibakusa T, Mizunoya W, Inoue K, Kitazawa R, Kitazawa S,
Matsuki Y, et al: FSP27 contributes to efficient energy storage in
murine white adipocytes by promoting the formation of unilocular
lipid droplets. J Clin Invest. 118:2808–2821. 2008.PubMed/NCBI
|
|
10
|
Flowers MT, Miyazaki M, Liu X and Ntambi
JM: Probing the role of stearoyl-CoA desaturase-1 in hepatic
insulin resistance. J Clin Invest. 116:1478–1481. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Flowers MT: The delta9 fatty acid
desaturation index as a predictor of metabolic disease. Clin Chem.
55:2071–2073. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jeyakumar SM, Lopamudra P, Padmini S,
Balakrishna N, Giridharan NV and Vajreswari A: Fatty acid
desaturation index correlates with body mass and adiposity indices
of obesity in Wistar NIN obese mutant rat strains WNIN/Ob and
WNIN/GR-Ob. Nutr Metab (Lond). 6:272009. View Article : Google Scholar
|
|
13
|
Dobrzyn A and Ntambi JM: Stearoyl-CoA
desaturase as a new drug target for obesity treatment. Obes Rev.
6:169–174. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Motoshima H, Goldstein BJ, Igata M and
Araki E: AMPK and cell proliferation - AMPK as a therapeutic target
for atherosclerosis and cancer. J Physiol. 574:63–71. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Foretz M, Carling D, Guichard C, Ferré P
and Foufelle F: AMP-activated protein kinase inhibits the
glucose-activated expression of fatty acid synthase gene in rat
hepatocytes. J Biol Chem. 273:14767–14771. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Woods A, Azzout-Marniche D, Foretz M,
Stein SC, Lemarchand P, Ferré P, Foufelle F and Carling D:
Characterization of the role of AMP-activated protein kinase in the
regulation of glucose-activated gene expression using
constitutively active and dominant negative forms of the kinase.
Mol Cell Biol. 20:6704–6711. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xiang X, Saha AK, Wen R, Ruderman NB and
Luo Z: AMP-activated protein kinase activators can inhibit the
growth of prostate cancer cells by multiple mechanisms. Biochem
Biophys Res Commun. 321:161–167. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ahmadian M, Duncan RE and Sul HS: The
skinny on fat: lipolysis and fatty acid utilization in adipocytes.
Trends Endocrinol Metab. 20:424–428. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dobrzyn P, Dobrzyn A, Miyazaki M, Cohen P,
Asilmaz E, Hardie DG, Friedman JM and Ntambi JM: Stearoyl-CoA
desaturase 1 deficiency increases fatty acid oxidation by
activating AMP-activated protein kinase in liver. Proc Natl Acad
Sci USA. 101:6409–6414. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang Y, Zhou L, Gu Y, Zhang Y, Tang J, Li
F, Shang W, Jiang B, Yue X and Chen M: Dietary chickpeas reverse
visceral adiposity, dyslipidaemia and insulin resistance in rats
induced by a chronic high-fat diet. Br J Nutr. 98:720–726. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Committee of Chinese Pharmacopoeia:
Chinese pharmacopoeia commission of sanitary ministry of People's
Republic of China. Chinese Pharmacopoeia. Uigur Pharmacopoeia
Fascicule. Xinjiang Science and Technology Sanitation Publisher;
Xinjiang: pp. 1141998
|
|
22
|
The Society of Japanese Pharmacopoeia: The
Japanese Pharmacopoeia. 15th edition. Yakuji Nippo; Tokyo: pp.
442006
|
|
23
|
Hummasti S, Laffitte BA, Watson MA,
Galardi C, Chao LC, Ramamurthy L, Moore JT and Tontonoz P: Liver X
receptors are regulators of adipocyte gene expression but not
differentiation: Identification of apoD as a direct target. J Lipid
Res. 45:616–625. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang F and Tong Q: Transcription factor
PU.1 is expressed in white adipose and inhibits adipocyte
differentiation. Am J Physiol Cell Physiol. 295:C213–C220. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim YC, Gomez FE, Fox BG and Ntambi JM:
Differential regulation of the stearoyl-CoA desaturase genes by
thiazolidinediones in 3T3-L1 adipocytes. J Lipid Res. 41:1310–1316.
2000.PubMed/NCBI
|
|
26
|
Mackall JC, Student AK, Polakis SE and
Lane MD: Induction of lipogenesis during differentiation in a
'preadipocyte' cell line. J Biol Chem. 25:6462–6464. 1976.
|
|
27
|
Enoch HG, Catalá A and Strittmatter P:
Mechanism of rat liver microsomal stearyl-CoA desaturase. Studies
of the substrate specificity, enzyme-substrate interactions, and
the function of lipid. J Biol Chem. 251:5095–5103. 1976.PubMed/NCBI
|
|
28
|
Miyazaki M, Bruggink SM and Ntambi JM:
Identification of mouse palmitoyl-coenzyme A Δ9-desaturase. J.
Lipid Res. 47:700–704. 2006. View Article : Google Scholar
|
|
29
|
Scaglia N, Chisholm JW and Igal RA:
Inhibition of stearoylCoA desaturase-1 inactivates acetyl-CoA
carboxylase and impairs proliferation in cancer cells: Role of
AMPK. PLoS One. 4:e68122009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Woods A, Johnstone SR, Dickerson K, Leiper
FC, Fryer LG, Neumann D, Schlattner U, Wallimann T, Carlson M and
Carling D: LKB1 is the upstream kinase in the AMP-activated protein
kinase cascade. Curr Biol. 13:2004–2008. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang BB, Zhou G and Li C: AMPK: An
emerging drug target for diabetes and the metabolic syndrome. Cell
Metab. 9:407–416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lane MD1, Wolfgang M, Cha SH and Dai Y:
Regulation of food intake and energy expenditure by hypothalamic
malonyl-CoA. Int J Obes (Lond). 32(Suppl 4): S49–S54. 2008.
View Article : Google Scholar
|
|
33
|
Schrauwen P and Hesselink M: UCP2 and UCP3
in muscle controlling body metabolism. J Exp Biol. 205:2275–2285.
2002.PubMed/NCBI
|
|
34
|
Pittaway JK, Ahuja KD, Robertson IK and
Ball MJ: Effects of a controlled diet supplemented with chickpeas
on serum lipids, glucose tolerance, satiety and bowel function. J
Am Coll Nutr. 26:334–340. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pittaway JK, Robertson IK and Ball MJ:
Chickpeas may influence fatty acid and fiber intake in an ad
libitum diet, leading to small improvements in serum lipid profile
and glycemic control. J Am Diet Assoc. 108:1009–1013. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zulet MA and Martinez JA: Corrective role
of chickpea intake on a dietary-induced model of
hypercholesterolemia. Plant Foods Hum Nutr. 48:269–277. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tanaka T, Yoshida N, Kishimoto T and Akira
S: Defective adipocyte differentiation in mice lacking the
C/EBPbeta and/or C/EBPdelta gene. EMBO J. 16:7432–7443. 1997.
View Article : Google Scholar
|
|
38
|
Rosen ED, Sarraf P, Troy AE, Bradwin G,
Moore K, Milstone DS, Spiegelman BM and Mortensen RM: PPAR gamma is
required for the differentiation of adipose tissue in vivo and in
vitro. Mol Cell. 4:611–617. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Linhart HG, Ishimura-Oka K, DeMayo F, Kibe
T, Repka D, Poindexter B, Bick RJ and Darlington GJ: C/EBPalpha is
required for differentiation of white, but not brown, adipose
tissue. Proc Natl Acad Sci USA. 98:12532–12537. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Puri V, Konda S, Ranjit S, Aouadi M,
Chawla A, Chouinard M, Chakladar A and Czech MP: Fat-specific
protein 27, a novel lipid droplet protein that enhances
triglyceride storage. J Biol Chem. 282:34213–34218. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ntambi JM and Miyazaki M: Regulation of
stearoyl-CoA desaturases and role in metabolism. Prog Lipid Res.
43:91–104. 2004. View Article : Google Scholar
|
|
42
|
Peter A, Cegan A, Wagner S, Lehmann R,
Stefan N, Königsrainer A, Königsrainer I, Häring HU and Schleicher
E: Hepatic lipid composition and stearoyl-coenzyme A desaturase 1
mRNA expression can be estimated from plasma VLDL fatty acid
ratios. Clin Chem. 55:2113–2120. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ntambi JM, Miyazaki M, Stoehr JP, Lan H,
Kendziorski CM, Yandell BS, Song Y, Cohen P, Friedman JM and Attie
AD: Loss of stearoyl-CoA desaturase-1 function protects mice
against adiposity. Proc Natl Acad Sci USA. 99:11482–11486. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cohen P, Miyazaki M, Socci ND,
Hagge-Greenberg A, Liedtke W, Soukas AA, Sharma R, Hudgins LC,
Ntambi JM and Friedman JM: Role for stearoyl-CoA desaturase-1 in
leptin-mediated weight loss. Science. 297:240–243. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Waters KM and Ntambi JM: Insulin and
dietary fructose induce stearoyl-CoA desaturase 1 gene expression
of diabetic mice. J Biol Chem. 269:27773–27777. 1994.PubMed/NCBI
|
|
46
|
Rahman SM, Dobrzyn A, Dobrzyn P, Lee SH,
Miyazaki M and Ntambi JM: Stearoyl-CoA desaturase 1 deficiency
elevates insulin-signaling components and down-regulates
protein-tyrosine phosphatase 1B in muscle. Proc Natl Acad Sci USA.
100:11110–11115. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Attie AD, Krauss RM, Gray-Keller MP,
Brownlie A, Miyazaki M, Kastelein JJ, Lusis AJ, Stalenhoef AF,
Stoehr JP, Hayden MR, et al: Relationship between stearoyl-CoA
desaturase activity and plasma triglycerides in human and mouse
hypertriglyceridemia. J Lipid Res. 43:1899–1907. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kerner J and Hoppel C: Fatty acid import
into mitochondria. Biochim Biophys Acta. 1486:1–17. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chien D, Dean D, Saha AK, Flatt JP and
Ruderman NB: Malonyl-CoA content and fatty acid oxidation in rat
muscle and liver in vivo. Am J Physiol Endocrinol Metab.
279:E259–E265. 2000.PubMed/NCBI
|
|
50
|
Brand MD and Esteves TC: Physiological
functions of the mitochondrial uncoupling proteins UCP2 and UCP3.
Cell Metab. 2:85–93. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Nedergaard J and Cannon B: The 'novel'
'uncoupling' proteins UCP2 and UCP3: What do they really do? Pros
and cons for suggested functions. Exp Physiol. 88:65–84. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Voss MD, Beha A, Tennagels N, Tschank G,
Herling AW, Quint M, Gerl M, Metz-Weidmann C, Haun G and Korn M:
Gene expression profiling in skeletal muscle of Zucker diabetic
fatty rats: Implications for a role of stearoyl-CoA desaturase 1 in
insulin resistance. Diabetologia. 48:2622–2630. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ntambi JM: Regulation of stearoyl-CoA
desaturase by polyunsaturated fatty acids and cholesterol. J Lipid
Res. 40:1549–1558. 1999.PubMed/NCBI
|
|
54
|
Choi Y, Kim YC, Han YB, Park Y, Pariza MW
and Ntambi JM: The trans-10,cis-12 isomer of conjugated linoleic
acid down-regulates stearoyl-CoA desaturase 1 gene expression in
3T3-L1 adipocytes. J Nutr. 130:1920–1924. 2000.PubMed/NCBI
|