1. Introduction
Fertilization is a sequence of coordinated molecular
events involving the merging of the sperm with the egg, the fusion
of the pronuclei and the intermingling of the maternal and paternal
chromosomes. The first form of human life is the zygote (a diploid
cell) from which the new organism will result. During sexual
intercourse, millions of sperm are deposited into the vagina. A
number of these will die in the acidic environment. However, many
will survive due to the protective elements provided in the fluids
surrounding them. Soon afterwards, the sperm have to swim through
the cervical mucus, towards to the uterus and then on to the
fallopian tubes. As they swim towards these, they decrease in
number, in an attempt to make it through the mucus. Inside the
uterus, the contractions of the uterus assist the journey of the
sperm towards the egg. Fertilization takes place in the ampulla of
the oviduct. If the oocyte is not fertilized here, it slowly passes
to the uterus, where it becomes degenerated and is absorbed.
Achieving fertilization requires the activation of sperm and oocyte
maturation. However, the oocyte/sperm interaction depends on a
number of changes occurring in the egg and the sperm.
The egg complex following ovulation enters the
oviduct consisting of three components: i) the oocyte (egg)
arrested at metaphase of meiosis II; ii) the extracellular matrix
of the egg or zona pellucida consisting of three glycoproteins
(ZP1, ZP2 and ZP3), synthesized and secreted by the oocyte; iii)
the cumulus oophorus, consisting of granulosa cells enriched with
hyaluronic acid (1,2). Cumulus cells support fertilization,
and in vitro fertilization can be achieved more efficiently
with them than without them (3–5).
The journey of the sperm is facilitated by ovarian
hormones that affect the structure, composition and activity of the
secretory epithelia of the cervix, the uterus and the fallopian
tubes and the contractility of these elements. The estrogen
hormones, favor these factors, while progesterone does not.
Oxytocin, which is secreted during intercourse by stimulation of
the posterior pituitary, causes the contraction of the uterus and
the fallopian tubes, as well as by the prostaglandins that affect
the contractility of the uterus and fallopian tubes. During the
ovulation period, the uterus becomes more sensitive to
prostaglandins.
Oocytes acquire the ability to fuse with sperm when
they reach 20 µm in diameter and they are arrested at the
prophase of meiosis II (6).
Spermatozoa undergo a series of events during maturation in order
to acquire motility and fertility, as they move from the proximal
towards the distal end of the epididymis. Only spermatozoa that
have passed through the epididymis are mature enough to be capable
of motility. Moreover, via deposition of new proteins in the
nucleus, the DNA becomes more condensed. The sperm head decreases
in size and becomes more compact. The above is crucial for the
subsequent correct decondensation of the paternal DNA in the
maternal oocyte. The ability for motility is achieved, but at the
same time is inhibited by the milieu. Finally, the structure of the
plasma membrane is altered by the addition of glycoproteins and
other proteins. This affects the motility, the capacitation ability
and the ability for the acrosome reaction (http://www.embryology.ch/anglais/dbefruchtung/bereitstell02.html#genitaltrakt).
Sperm is the male gamete and is derived from the
Greek word 'sperma' (meaning 'seed'). It consists of three parts, a
head (containing DNA and enzymes in order to penetrate the oocyte),
a midpiece (containing cellular elements, centrioles, microtubules
and a number of mitochondria for energy production required to
promote sperm) and a tail (enabling sperm movement) (http://www.newworldencyclopedia.org/entry/Sperm).
The head of normal sperm has an oval shape, a length
of 3–5 µm, a width of 2–3 µm and is 1.5 µm
thick. It consists of the nucleus and the acrosome, which are
covered by the nuclear membrane and the metacrosomic sheath. The
nucleus contains the male genetic material. The DNA of the sperm
has a slightly different structure from the DNA of a diploid
(somatic) cell, meaning that the DNA is concentrated in the
smallest possible volume, so as to save space. The acrosome
contains proteolytic enzymes, such as acrosin, trypsin,
hyaluronidase and proteases, which are released during the acrosome
reaction and hydrolysis in granular cells and the zona pellucida of
the egg in order to assist the penetration of the sperm and the
merger with the egg (7).
The neck or mid-section of the sperm connects the
head to the tail (length, 7.5 µm; and width, 1 mm). It
contains the axial filament surrounded by fibrils with circular
layout and one to two centrosomes. The neck contains mitochondria,
enzymes of glycolysis and oxidation systems and provides the
necessary energy for survival and mobility.
The tail (length, 40–50 mm) consists of a total of
ten pairs of fibrils (a central and nine peripheral) and is
responsible for the typical sperm motility.
The sperm can survive for 3–5 days inside the
vagina, particularly in the vagina or mucous membranes in the upper
part of the female genital tract. Fertilization in this case is
possible if the sperm remain alive. A single sperm is sufficient to
fertilize the egg. Sperm survive outside the female body for up to
several hours. When the sperm meets the oocyte, they swim around
it. Subsequently, one of them adheres to the head of the oocyte and
begins rotational movements about the longitudinal axis. The whole
sperm then penetrates the oocyte, with the acrosome penetrating the
egg cover [http://www.yourarticlelibrary.com/biology/human-reproduction/structure-of-human-sex-gametes-spermatozoan-and-ovum-biology/26913/;
http://www.onmed.gr/ygeia/story/329099/poso-zei-to-sperma-mesa-ston-kolpo#ixzz44lwgUQgb
(in Greek)].
2. Molecules synthesized and secreted from
the oocyte, which orient and stimulate sperm
Resact (respiratory activating peptide) is a peptide
of 14 amino acids, isolated from the egg jelly of the sea urchin,
Arbacia punctulata (8). It
is probably recognized by the guanylate cyclase (guanylate
cyclase), a protein of the cell membrane of the sperm, which causes
an increase in Ca2+ levels. Experimentally, when resact
is added to a solution with spermatozoa, the latter undergoes
helical movements directed towards the point of resact (http://worms.zoology.wisc.edu/dd2/echino/fert/chemo-taxis/chemotaxis.html).
The sperm receptor for resact is a transmembrane
protein. Once it binds resact on the extracellular side, it causes
a conformational change on the cytoplasmic side, and it activates
the receptor's enzymatic activity. This triggers the mitochondrial
ATP-generating apparatus, as well as the dynein ATPase that
stimulates flagellar movement in the sperm (9,10).
Speract (sperm activating peptide) consists of ten
amino acids and appears to promote the increase in pH and promotes
sperm movement. The speract receptor is localized to the sperm tail
(11), which contains the
axoneme, the organelle of motility. The signaling in the sperm tail
is central to motility regulation. There is evidence that speract
can modulate sperm motility (12).
3. The three stages of fertilization
Sperm preparation: Capacitation and
acrosome reaction
The phase of sperm maturation is known as activation
(sperm capacitation). It occurs in the genital tract of the female,
and acts as a preparatory step for the acrosome reaction.
Activation does not include morphological changes and is
accompanied by hyperactivation of the sperm which is a strong,
non-linear motion (2). An
important role is played by proteolytic enzymes (13).
In mammals, the ejaculated sperm is motile. However,
their ability to fertilize an oocyte is reduced. The latter may
occur after removing inhibitory factors, such as surface-attached
glycoproteins, seminal plasma proteins and the depletion of
membrane cholesterol. This final state of activated sperm is known
as hyperactivation, and is a high energy phase of vigorous
flagellar movement and swimming capacity (14).
Capacitation involves a number of processes, such as
functional coupling of the signal transduction pathways that
regulate the initiation of acrosome reactions by ZP3; alterations
in flagellar motility that may be required to penetrate the zona
pellucida; and the development of the capacity to fuse with eggs
(15). The above-mentioned events
are followed by alterations in metabolism, membrane biophysical
characteristics, changes in the protein phosphorylation state,
elevations in intracellular pH and calcium levels, and
hyperpolarization of membrane potential. In vivo, many
factors are likely to mediate the activation of sperm. Sterol
binding proteins, such as high density lipoprotein, have been
identified in the fallopian tubes and can accelerate the efflux of
cholesterol from the sperm (16).
Furthermore, progesterone can regulate some aspects of sperm
activation. It is present in the environment of the fallopian
tubes, derived both from follicular fluid and cells of the cumulus
oophorus (17). Glucose is known
to be essential for successful capacitation. It functions not only
as an energy molecule allowing spermatozoa to swim, but it also
enables the spermatozoa to fertilize eggs (18).
The agents of the oocyte mucous that cause the
acrosome reaction are specific for each species. In mammals, sperm
recognizes and binds to ZP3 glycoprotein of the zona pellucida. The
O-polysaccharide connections to the core protein of ZP3 seem to be
necessary in this step. It is believed that the terminal galactose
of O-linked oligosaccharides reacts with galactosyltransferase (Mr
56 kDa), a cell membrane protein in the anterior end of the sperm
head. The acrosome reaction of the sperm is caused by the core
protein of ZP3. Once the core protein of ZP3 is damaged, this
reaction is inhibited, but not the binding of the sperm to the zona
pellucida. The sperm reaching the transparent zone is connected via
SED1 protein to ZP3. As a result of irreversible binding of the
sperm to the egg, the zona pellucida triggers the acrosome
reaction.
The outer plasma membrane of the acrosome fuses at
multiple sites with the plasma membrane and the contents of the
acrosome are released. After the acrosome reaction, it is believed
that the protein pre-acrosin of the acrosomal vesicle binds to ZP2,
and becomes activated, forming the enzyme acrosin which digests the
transparent area at this point. Digestion of the zona pellucida is
followed by the fusion of the membrane of the sperm to the egg
membrane, which seems to be caused by the PH-30 sperm protein
(19).
PH-30 is a transmembrane glycoprotein consisting of
an α and β subunit. The α subunit (289 amino acids) contains a
sequence between 90 and 111 which has many similarities to the
'fusion peptide' E2 glycoprotein of the rubella virus. The β
subunit (353 amino acids) contains the tripeptide RGD
(arginine-glycine-aspartic acid) in the last 90 amino acids of the
N-terminus. The tripeptide RGD is the active site which can be
recognized by integrins of the cell membrane of the egg. This
recognition is followed by the fusion of the membranes, which is
caused by the α subunit of PH-30. Both subunits derive from
precursors that undergo proteolytic modification during the
differentiation of sperm in the testes and their maturation in the
epididymis. PH-30 is located in the cell membrane in the anterior
portion of the side part of the mature sperm head (19,20). The acrosome reaction involves the
fusion of the acrosome membrane with the overlying plasma membrane
of the sperm and vesicle formation. The result is the release of
peptidases from the acrosomal vesicle that digest the mucous coat.
The fusion of the membranes appears to be caused by the entry of
Ca+2 ions and the exit of K+ ions. The sperm
plasma membrane that does not participate in the acrosome reaction
and overlying the acrosome is known as the equatorial region. This
seems to be the site of sperm and egg fusion (2). An acrosomal filament, is formed by
the polymerization of G (Globular) actin to F (Fibrilar) actin.
Desmin (30.5 kDa) is exposed to the acrosomal filament. The
polymerization of actin is induced by the increase in pH (~7.4),
which results from the input of Na+ and H+
output. The activation of dynein ATPase and the use of ATP results
in an increased movement of the sperm. cAMP levels increase
(approximately 400-fold) in the cytoplasm of sperm. This increase
marks the beginning of the process of chromatin decondensation of
the sperm, before the formation of the zygote nucleus.
P4 (progesterone) is a well-studied molecule
involved in the modulation of sperm function. Its role in sperm
capacitation has been revealed; however, the exact mechanisms
involved are not yet fully understood. Progesterone is involved not
only in capacitation, but also in other events, such as
hyperactivated motility, flagellar bending, in chemotaxis, in
acrosome reaction and in sperm-ZP binding and sperm-oocyte fusion.
Its effects on sperm function have been studied in humans, as well
as in other mammals (mouse, boar) (21–24).
Evans and Florman, in Nature Cell Biology and Nature
Medicine (15), propose a model
according to which in the sperm head, the receptor TPC2-ZP3 is
triggered by its association with the zona pellucida. This results
in calcium entry through T-type channels, leading to a transient
increase in calcium levels in the cytoplasm and the activation of
phospholipase C (PLC) via protein G. PLC is divided into IP3 and
DAG. This causes Ca+ entrance and leads to the acrosome
reaction.
In the capacitation process,
phosphatidyl-inositol-3-kinase (PI3K) is phosphorylated/activated
via a protein kinase A (PKA)-dependent cascade, and is
downregulated by PKCa. At the beginning of capacitation, PKCa is
active and results in PI3K inactivation. PKCa and PP1c2 are
degraded by a PKA-dependent mechanism, and this allows the
activation of PI3K. The activation of PKA, which depends on cAMP
produced by the bicarbonate-dependent soluble adenylyl cyclase,
leads to an increase in actin polymerization, an essential step so
as to reach hyperactivated motility, which is necessary for
successful fertilization (25).
The sperm-specific CatSper ion channels have been
suggested to control the intracellular Ca2+
concentration and, thereby, the swimming behavior of spermatozoa
(26). The CatSper channel seems
to be activated by progesterone in human spermatozoa (29), which is interesting from the
sperm-egg interaction viewpoint, as cumulus cells are known to
produce progesterone (5).
Sperm-egg binding and fusion
The plasma membrane of the oocyte consists of two
sites, a microvilli-free region and a microvilli-rich region. The
fusion of sperm and oocyte takes place in the microvilli-rich
region (28,29). The interaction between sperm and
oocyte is preceded by acrosomal exocytosis, triggered by sperm and
ZP binding. It involves sperm attachment to the oocyte and
cell-cell adhesion, leading to membrane fusion of the two gametes
(30). The inner acrosomal
membrane of the sperm, being exposed following the acrosome
reaction, comes into contact with the oocyte membrane (31). The equatorial segment adheres to
the posterior head of the sperm and fuses with the oocyte membrane
(32,33).
Several molecules have been identified in sperm and
oocytes, with a crucial role in gamete binding. Fertilin α,
fertilin β and cyritestin are also known as ADAM1, ADAM2 and ADAM3,
respectively. Members of this family consist of a signal sequence
domain, a metalloprotease domain, a disintegrin-like domain, a
cysteine-rich domain and an epidermal growth factor-like repeat. A
number of studies have revealed a role of ADAM1, ADAM2 and ADAM3 in
sperm oocyte binding (30).
Fertilin β has been shown not to be essential for plasma membrane
binding and fusion. Studies on fertilin β and cyritestin with
knockout mice have shown a reduced adhesion to the oocyte plasma
membrane; however, some sperm adhere and they fuse with the oocyte
(15,34). Moreover, they show poor adhesion
pattern to the ZP (34–36).
As regards oocytes, integrins found on the egg
surface are thought to be receptors for sperm ADAMs. Studies have
revealed that α6β1 integrin is an egg receptor for fertilin β
(37–39). Other studies have indicated that
α9β1 integrin is a receptor for fertilin β (40,41) CD46 has been found to be expressed
in rodents on the acrosomal membrane of sperm (42). CD46 interacts directly with β1
integrin and indirectly with tetraspanins in human cells (43). However, the key role of CD46
appears to stabilize the acrosomal membrane (44).
As an integrin-associated protein, in the oocyte,
CD9 is essential for sperm-egg interactions. It is a member of a
tetraspanin protein family expressed on the mouse egg surface
(45). The role of CD9 in sperm
oocyte fusion has been demonstrated in a number of studies with
CD9-null oocytes which showed reduced ability for strong sperm
adhesion (46). CD9-deficient
mice show reduced fertility (47–49). Another tetraspanin member, CD81,
is expressed on the oocyte surface and interacts with CD9 (50). Deletion of the CD81 gene results
in a reduction in fertility. However, its role in sperm oocyte
interaction has not yet been fully elucidated (51,52).
CRISP1, a sperm protein expressed by the cumulus
cells surrounding the oocyte, stimulates sperm orientation through
the modulation of sperm hyperactivation and it seems to regulate
CatSper (53). Recent studies
have revealed an oocyte-derived chemotactic activity associated
with a hydrophobic non-peptide molecule in human sperm (54) and an association between follicle
rupture and uterine contractions with the success of human in
vivo insemination, suggesting the existence of possible
chemoattractive substances in the female tract (55). It is not clear if there is a
binding partner on the oocyte membrane for sperm-associated CRISP
protein, along with the exact mechanisms through which gamete
binding is achieved.
The sperm-specific protein, Izumo, is essential for
sperm-egg plasma membrane binding and fusion (56). IZUMO interacts directly with some
molecules on the oolema. On the oocyte side, Juno is a member of
the folate receptor family and recognizes the sperm IZUMO,
facilitating fertilization. It has been revealed that mice lacking
Juno on the surface of their egg cells are infertile as their egg
cells do not fuse with normal sperm. This finding demonstrates the
essential role of Juno in the fertility of female mice (57). Other studies have shown that a
helical dimer of fragments of the N-terminus domain of IZUMO is
required for sperm-oocyte fusion (58). IZUMO forms complexes with other
proteins on the sperm surface via its N-terminal domain which forms
dimers (56). Therefore, IZUMO
plays a key role in organizing and stabilizing a protein-like
complex crucial for membrane fusion. Inoue et al (56) found a protein,
angiotensin-converting enzyme 3 (ACE3) on the sperm acrosomal cap
capable of interacting with IZUMO (59).
Studies have revealed the rapid loss of Juno from
the oocyte membrane soon after fertilization. This implies that
Juno is essential for the process of fertilization, thus, being the
basis for polyspermy block in mammals. One possible explanation for
this process is that Juno is shed in vesicles following
fertilization, generating a zone of 'decoy oocyte' confined within
the perivitelline space that could bind acrosome-reacted sperm and
therefore avoid supernumerary sperm entry (60). Although it has been validated that
Juno protein as the first cell surface receptor conserved in
mammals, the interaction between IZUMO1 and Juno seems to be a
necessary and an essential adhesion step; however, its role in the
gamete fusion mechanism is not yet clear (57).
Two molecules, trypsin-like acrosin and spermosin
proteases have been proposed to be involved in the first physical
contact of the two gametes, suggesting that a proteasome system
enables sperm to penetrate the chorion or participate in the
process as sperm-related 'moveable' binding proteins (52). Sperm hyaluronidases are believed
to play an intriguing role in fertilization in mammals, and
sperm-specific SPAM1 and HYAL5 hyaluronidase have been suggested to
be involved in sperm-ZP binding in mice. Recent studies have shown
that hyaluronidases are not required for fertilization (61,62).
It is important to mention that the oviductal
environment and its secretions also play an essential role in the
transport and interaction of male and female gametes. The
expression of lactoferrin, a human oviductal protein, is able to
inhibit gamete interaction in vitro and it seems to be
involved in the regulation of the reproductive process through a
role in polyspermy prevention (63). It has also been shown that
lactoferrin causes a decrease in sperm α-D-mannose binding sites
and an increase in tyrosine phosphorylation of sperm proteins, thus
implying that this protein is able to modulate the parameters of
sperm function (64).
Cortical reaction: Meiosis resumption of
oocyte and activation of the zygote
Once the sperm fuses with the oocyte, the beating of
the tail stops immediately. The fusion of sperm and the oocyte
membrane appears to cause the polymerization of actin and
microvilli extension. The sperm instead is drawn into the oocyte by
elongation and fusion of the microvilli of the egg. As a result,
the sperm nucleus and other organelles are incorporated into the
oocyte cytoplasm. The actin filaments are essential for the
attraction of the sperm into the oocyte. The cytoplasm swells and
forms colliculus (~7 microns in length and 2 microns wide)
resembling a so-called fertilization cone. In some invertebrates
and amphibians, there are certain areas of the oocyte membrane for
the binding and fusion of the sperm.
Seconds after fertilization, the membrane potential
of the oocyte undergoes a large depolarization via a massive influx
of Na+ ions. The depolarization of the egg takes about a
minute to repolarize via K+ leakage. This is the fast
block to polyspermy: sperm cannot fuse to a membrane that is not
−70 mV (https://structureandfunction.wordpress.com/2013/02/28/the-fast-and-slow-blocks-to-polyspermy-and-egg-activation/).
CG distribution and the cortical reaction following
calcium oscillation are crucial steps in the prevention of
polyspermy. Calcium released from the endoplasmic reticulum is
dependent on the quality and quantity of the mitochondria, which
are markers of oocyte quality, since a low mitochondrial DNA copy
number results in poor oocyte developmental competence (65,66).
PLCζ from spermatozoa is considered to be the
responsible activator during this process (67); however, other factors may also be
involved (5,68).
The cortical reaction is a process through which
cortical granules from the oocyte are released preventing
polyspermy. The fast block of polyspermy immediately prevents
additional sperm getting attached to the oocyte. On the other hand,
the cortical reaction establishes a permanent barrier to sperm
entry and functions as the main part of the slow block of
polyspermy in many animals. The cortical reaction is propagated
over the surface of the egg by a wave of Ca++.
In this process, secretory vesicles located in the
region below the plasma membrane of the oocyte, are fused with the
oocyte plasma membrane. This results in the release of contents of
the cortical granules, modifying the extracellular matrix so as to
be impenetrable to other sperm. The cortical granules contain
hydrolytic enzymes, such as proteases that clip perivitelline
proteins, peroxidases that harden the vitelline envelope and
glycosaminoglycans that attract water into the perivitelline space,
causing it to expand and form the hyaline layer. Calcium ions from
the cortical smooth endoplasmic reticulum are responsible for
cortical granule rupture. It is believed that this is provoked by
the activation of a G-protein in the plasma membrane of the oocyte.
The sperm binds to ZP3 which is consistent with the G-protein and
becomes active. Bindin binds the sperm to the oocyte cell membrane,
but another sperm factor activates a receptor, which in turn
activates the G-protein. G-proteins of the spermatozoon itself
(activated when sperm was in contact with the mucous of the egg
shell with the initiation of the acrosome reaction) activate
enzymes, which results in increased Ca+2 levels in the
egg.
In mammals, the cortical reaction modifies the zona
pellucida, leading to the block of polyspermy. Several enzymes are
released by the cortical granules, leading to the digestion of the
sperm receptor glycoproteins ZP2 and ZP3, so that they can no
longer bind spermatozoon (https://en.wikipedia.org/wiki/Cortical_reaction).
It is interesting to know hows sperm factor causes
the release of Ca+2. Activated G-protein activates the
enzyme, PLC. The latter cleaves the lipid phosphotidylinositol
4,5-biphosphate (PIP2) in diacyglycerol (DAG) and inositol
1,4,5-triphosphate (IP3). PIP2 and DAG are second messengers. IP3
binds to its receptors in the endoplasmic reticulum, at the entry
point of the sperm. This results in the release of Ca+2
from the endoplasmic reticulum at this point. Ca+2 ions
bind to sensitive Ca+2 receptors in the endoplasmic
reticulum around the cortical granules. Ca+2 binding to
these receptors results in the release of more Ca+2,
which spreads like a wave in the region (cortex) of the egg and
causes the rupture of cortical granules. DAG activates PKC which
phosphorylates protein exchange ions Na+ to
H+. The protein that exchanges ions, is activated by
Ca+2 and the result of this activation is the entrance
to Na+ and H+ output and an increase in pH
from 6.8 to 7.3 in the egg. This 'wakes up' the oocyte from the
metabolic inertia (69).
The last phase of oocyte activation is the
resumption and completion of meiosis. This leads to polar body
extrusion, cleavage of the zygote and embryonic cell divisions.
There are widespread changes in molecules, such as proteins and
RNAs that are not necessarily involved in meiosis resumption.
Changes in proteomes and their composition have been revealed,
showing significant degradation of maternal proteins. This is due
to protein degradation, phosphorylation, post-translational
modifications and new translation of maternal RNAs (70).
Cytoplasmic polyadenylation involves the elongation
of the poly(A) tail after the export of mRNAs to the cytoplasm. It
has been observed and described in the oocytes and early embryos of
many animal species, from invertebrates to mammals, and is
universally considered to be a regulatory mechanism for protein
expression from specific mRNAs. The mediators of this process
(cytoplasmic polyadenylation elements and their binding proteins)
have been described in detail and some new findings have been
reviewed recently (71,72).
The sperm nucleus undergoes a series of changes,
including chromatin decondensation and the formation of a new
nuclear envelope, to form the male pronucleus. The latter uses
microtubules to migrate to the center of the cell, where it fuses
with the female pronucleus to form a diploid nucleus. Other sperm
organelles (e.g., mitochondria) persist during the early cleavage
stages of the embryo and they may play a role in development
(19).
The beginning, condensation of chromatin seems to
happen with two histone phosphorylations of the sperm and their
exchange with histones of the oocyte. Chromatin is condensed
following interaction with histones H3 and H4, and X and Y
sperm-specific proteins. In the cytoplasm of the ovum is the
protein nycleoplasmin connected with histones H2A and H2B. As
nucleoplasmin has greater affinity for the X and Y sperm proteins,
the exchange of histones H2A/H2B with X and Y proteins occurs. This
exchange allows the de-concentration and sharpness of the chromatin
nucleus of the sperm. The process of decompression begins from the
periphery to the center of the nucleus. When this process is
finished, the vesicles of the nuclear membrane of the sperm bind to
vesicles from the endoplasmic reticulum of the egg. The membrane of
the pronucleus of the sperm is produced. While the pro nucleus of
the sperm is formed, the nucleus of the oocyte completes the second
division of meiosis and eliminates the second polar body. After
entering the oocyte, the nucleus and the sperm centrosome rotate
180°, so that the centrosome is positioned between the nucleus of
the sperm and ovum. The centrosome organizes microtubules that
attach and pull the pronucleus of the sperm and the egg. Both
pronuclei migrate towards each other and they are directed towards
the center of the cytoplasm of the ovum. The membranes of the two
pronuclei rupture and mixing of nuclear material is observed,
resulting in the formation of the diploid nucleus of the zygote.
The fusion of the pronuclei occurs in <1 h e.g., in
Echinus (the meiotic division is finished before ovulation).
Approximately 12 h in mammals including most eggs (e.g., human) is
needed to complete the second division of meiosis with the
excretion of the second polar body (73). Fig.
1 illustrates the events taking place in fertilization.
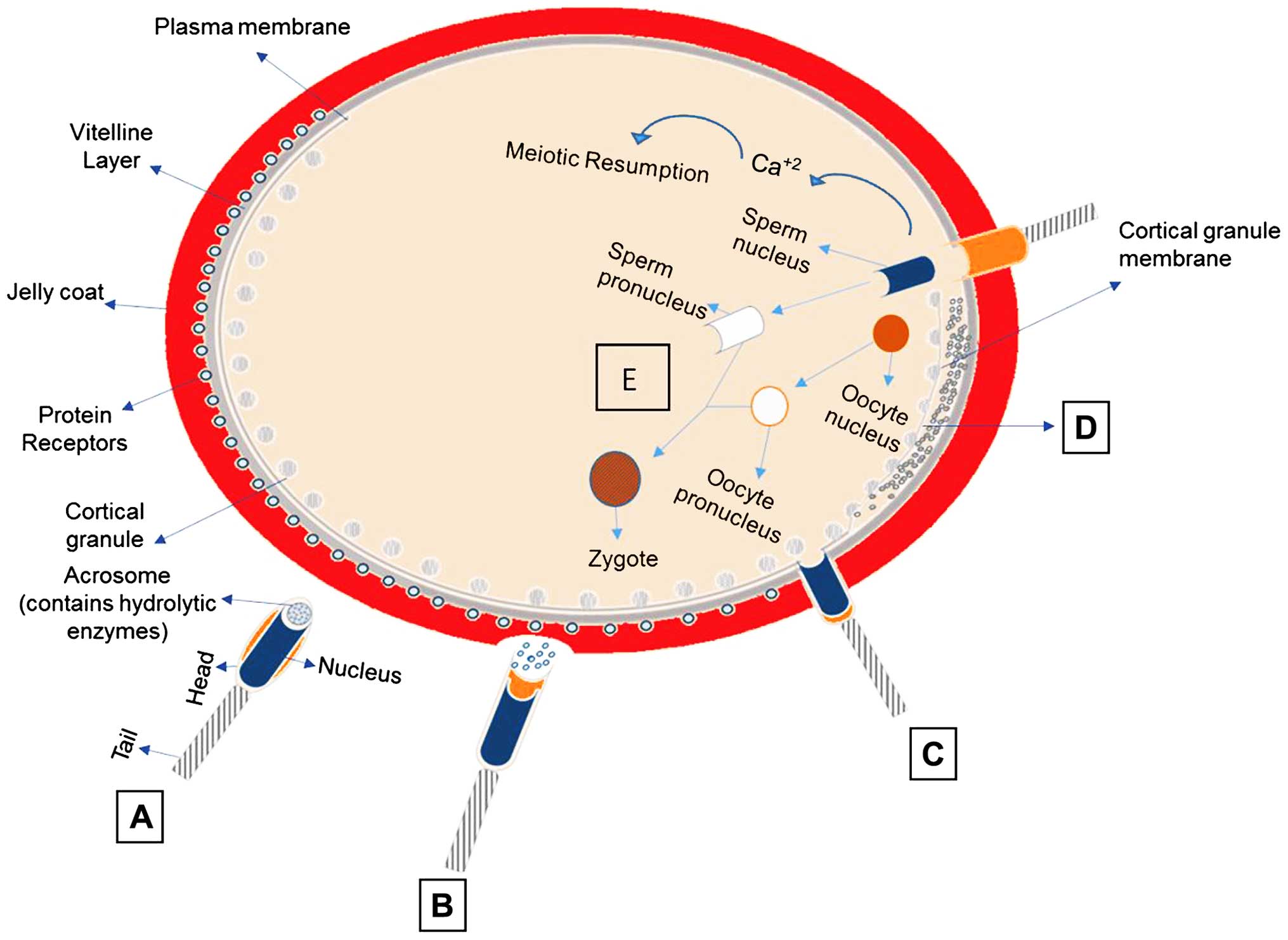 | Figure 1The events taking place in
fertilization. (A) Sperm preparation-capacitation: Molecules
(resact, speract) secreted from the oocyte, orient and stimulate
sperm (guanylate cyclase). (B) Acrosome reaction: release of
hydrolytic enzymes. The sperm via SED1 protein is connected to ZP3.
(C) Fusion of sperm with plasma membrane of the oocyte: sperm
pre-acrosin binds to ZP2. Proteins of sperm IZUMO, ADAMs 1, ADAMs
2, ADAMs 3 and CRISP1 bind to receptors on the oocyte (Juno,
integrins, CD9, CD81). Other molecules identified playing role in
gamete fusion are: Trypsin-like acrosin, spermosin, SPAM1, HYAL5,
ACE3. (D) Cortical Reaction: Ca+2 release/wave of
Ca+2 and formation of fertilization cone. Enzymes
released by cortical granules, digest sperm receptors ZP2 and ZP3
(block of polyspermy). (E) Sperm chromatin decondensation to form
male pronucleus: The oocyte nucleus completes the 2nd meiosis and
eliminates the 2nd polar body. |
4. Reorganization and partition of the
zygote
Once the sperm enters the oocyte, the cytoplasmic
material is reorganized. The cytoplasm of the oocyte contains
cytoplasmic determinants that become distributed on different cells
during cleavage. The cytoplasmic determinants (e.g., Vg1 and
bicoid) directly or indirectly activate or inactivate genes and
control events of differentiation in an early embryo. The point
through which sperm has penetrated the oocyte marks the bilateral
symmetry of the future adult organism. Following fertilization,
cortical rotation occurs. Tubulin microtubules are organized as
train tracks between the cortical and the endoplasm in the vegetal
pole. The result is the appearance of a gray area, the gray
crescent, which marks the future dorsal side of the embryo. Between
the midpoint of the gray crescent and the entry point of sperm is
the level of the lateral symmetry normally marked with the first
cleavage.
The gray crescent is a region of zygote in which
early morphogenetic movements will begin to form the blastopore,
which marks the beginning of the process of gastrulation in the
early embryo. Following fertilization, a sequence of cell divisions
results in the formation of the early embryo. The zygote begins to
divide mitotically (cleavage), and the resulting cells are termed
blastomeres. As long as cleavage progresses, the cells proliferate
and become progressively smaller, and the fetal size and general
shape remains the same (73).
The first cleavage begins as 'a ring' (cleavage
furrow) around the zygote and is parallel to the polar axis
(animal-vegetal pole) of the zygote and normal to the longitudinal
axis (axis joining the poles of the spindle) of the mitotic
spindle. The ring consists of actin-myosin found in the cortical
separating the zygote inwardly. The myosin plays the role of ATPase
and controls the actin. The level of the second cleavage is
parallel to the polar axis. At this stage, the embryo consists of
four blastomeres, extending from the animal to the vegetal pole.
Cleavage is a fractionating process, resulting in the formation of
an eight-cell stage and in a 16–32 cell so-called morula (69).
5. Fertilization failure
Some fertility issues may arise from failure in
fertilization events, in the molecular base, described in this
review. Defects in the expression of receptors in the oocyte cell
membrane (e.g., CD9), destruction of the gamete fusion mechanism,
insufficient or abnormal variation of Ca2+ and failure
of exocytosis of cortical granules, inadequate completion of second
meiotic division of the oocyte, defective sperm chromatin
decondensation, defective pronucleus due to abnormal remodeling of
chromatin (multiple pronuclei) or abnormal formation/function of
the mitotic spindle, may account for infertility.
References
|
1
|
Wassarman PM and Florman HM: Cellular
mechanisms during mammalian fertilization. Handbook of Physiology:
Section 14-Cell Physiology. Oxford University Press; New York, NY:
pp. 885–938. 1997
|
|
2
|
Yanagimachi R: The Physiology of
Reproduction. Raven Press; New York, NY: pp. 189–317. 1994
|
|
3
|
Jin M, Fujiwara E, Kakiuchi Y, Okabe M,
Satouh Y, Baba SA, Chiba K and Hirohashi N: Most fertilizing mouse
spermatozoa begin their acrosome reaction before contact with the
zona pellucida during in vitro fertilization. Proc Natl Acad Sci
USA. 108:4892–4896. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tokuhiro K, Ikawa M, Benham AM and Okabe
M: Protein disulfide isomerase homolog PDILT is required for
quality control of sperm membrane protein ADAM3 and male fertility
[corrected]. Proc Natl Acad Sci USA. 109:3850–3855. 2012.
View Article : Google Scholar
|
|
5
|
Okabe M: The cell biology of mammalian
fertilization. Development. 140:4471–4479. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zuccotti M, Piccinelli A, Giorgi Rossi P,
Garagna S and Redi CA: Chromatin organization during mouse oocyte
growth. Mol Reprod Dev. 41:479–485. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nelson L: Enzymes associated with sperm
cell function. Biology of Fertilization. Metz CB and Monroy A:
London Academic Press; Orlando, FL: pp. 215–131. 1985, View Article : Google Scholar
|
|
8
|
Ward GE, Brokaw CJ, Garbers DL and
Vacquier VD: Chemotaxis of Arbacia punctulata spermatozoa to
resact, a peptide from the egg jelly layer. J Cell Biol.
101:2324–2329. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shimomura H, Dangott LJ and Garbers DL:
Covalent coupling of a resact analogue to guanylate cyclase. J Biol
Chem. 261:15778–15782. 1986.PubMed/NCBI
|
|
10
|
Cook SP and Babcock DF: Selective
modulation by cGMP of the K+ channel activated by
speract. J Biol Chem. 268:22402–22407. 1993.PubMed/NCBI
|
|
11
|
Cardullo RA, Herrick SB, Peterson MJ and
Dangott LJ: Speract receptors are localized on sea urchin sperm
flagella using a fluorescent peptide analog. Dev Biol. 162:600–607.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cook SP, Brokaw CJ, Muller CH and Babcock
DF: Sperm chemotaxis: Egg peptides control cytosolic calcium to
regulate flagellar responses. Dev Biol. 165:10–19. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Talbot P and Franklin LE: Surface
modification of guinea pig sperm during in vitro capacitation: An
assessment using lectin-induced agglutination of living sperm. J
Exp Zool. 203:1–14. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lishko PV, Kirichok Y, Ren D, Navarro B,
Chung JJ and Clapham DE: The control of male fertility by
spermatozoan ion channels. Annu Rev Physiol. 74:453–475. 2012.
View Article : Google Scholar
|
|
15
|
Evans JP and Florman HM: The state of the
union: the cell biology of fertilization. Nat Cell Biol. 4:s57–s63.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Visconti PE, Ning X, Fornés MW, Alvarez
JG, Stein P, Connors SA and Kopf GS: Cholesterol efflux-mediated
signal transduction in mammalian sperm: Cholesterol release signals
an increase in protein tyrosine phosphorylation during mouse sperm
capacitation. Dev Biol. 214:429–443. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Contreras HR and Llanos MN: Detection of
progesterone receptors in human spermatozoa and their correlation
with morphological and functional properties. Int J Androl.
24:246–252. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Goodson SG, Qiu Y, Sutton KA, Xie G, Jia W
and O'Brien DA: Metabolic substrates exhibit differential effects
on functional parameters of mouse sperm capacitation. Biol Reprod.
87:752012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Blobel CP, Myles DG, Primakoff P and White
JM: Proteolytic processing of a protein involved in sperm-egg
fusion correlates with acquisition of fertilization competence. J
Cell Biol. 111:69–78. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Primakoff P, Hyatt H and Tredick-Kline J:
Identification and purification of a sperm surface protein with a
potential role in sperm-egg membrane fusion. J Cell Biol.
104:141–149. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
López-Torres AS and Chirinos M: Modulation
of human sperm capacitation by progesterone, estradiol, and
luteinizing hormone. Reprod Sci. 12:1–9. 2016.
|
|
22
|
Roldan ER, Murase T and Shi QX: Exocytosis
in spermatozoa in response to progesterone and zona pellucida.
Science. 266:1578–1581. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu JT, Chiang KC and Cheng FP: Expression
of progesterone receptor(s) during capacitation and incidence of
acrosome reaction induced by progesterone and zona proteins in boar
spermatozoa. Anim Reprod Sci. 93:34–45. 2006. View Article : Google Scholar
|
|
24
|
de Lamirande E, Harakat A and Gagnon C:
Human sperm capacitation induced by biological fluids and
progesterone, but not by NADH or NADPH, is associated with the
production of superoxide anion. J Androl. 19:215–225.
1998.PubMed/NCBI
|
|
25
|
Ickowicz D, Finkelstein M and Breitbart H:
Mechanism of sperm capacitation and the acrosome reaction: Role of
protein kinases. Asian J Androl. 14:816–821. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chang H and Suarez SS: Two distinct Ca(2+)
signaling pathways modulate sperm flagellar beating patterns in
mice. Biol Reprod. 85:296–305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brenker C, Goodwin N, Weyand I, Kashikar
ND, Naruse M, Krähling M, Müller A, Kaupp UB and Strünker T: The
CatSper channel: A polymodal chemosensor in human sperm. EMBO J.
31:1654–1665. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ebensperger C and Barros C: Changes at the
hamster oocyte surface from the germinal vesicle stage to
ovulation. Gamete Res. 9:387–397. 1984. View Article : Google Scholar
|
|
29
|
Johnson MH, Eager D, Muggleton-Harris A
and Grave HM: Mosaicism in organisation concanavalin A receptors on
surface membrane of mouse egg. Nature. 257:321–322. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Evans JP: The molecular basis of
sperm-oocyte membrane interactions during mammalian fertilization.
Hum Reprod Update. 8:297–311. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang TTF Jr and Yanagimachi R: Inner
acrosomal membrane of mammalian spermatozoa: Its properties and
possible functions in fertilization. Am J Anat. 174:249–268. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yanagimachi R and Noda YD: Physiological
changes in the postnuclear cap region of mammalian spermatozoa: A
necessary preliminary to the membrane fusion between sperm and egg
cells. J Ultrastruct Res. 31:486–493. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bedford JM, Moore HDM and Franklin LE:
Significance of the equatorial segment of the acrosome of the
spermatozoon in eutherian mammals. Exp Cell Res. 119:119–126. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cho C, Bunch DO, Faure JE, Goulding EH,
Eddy EM, Primakoff P and Myles DG: Fertilization defects in sperm
from mice lacking fertilin β. Science. 281:1857–1859. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shamsadin R, Adham IM, Nayernia K,
Heinlein UAO, Oberwinkler H and Engel W: Male mice deficient for
germ-cell cyritestin are infertile. Biol Reprod. 61:1445–1451.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nishimura H, Cho C, Branciforte DR, Myles
DG and Primakoff P: Analysis of loss of adhesive function in sperm
lacking cyritestin or fertilin beta. Dev Biol. 233:204–213. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Almeida EA, Huovila AP, Sutherland AE,
Stephens LE, Calarco PG, Shaw LM, Mercurio AM, Sonnenberg A,
Primakoff P, Myles DG and White JM: Mouse egg integrin α 6 β 1
functions as a sperm receptor. Cell. 81:1095–1104. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen MS, Tung KS, Coonrod SA, Takahashi Y,
Bigler D, Chang A, Yamashita Y, Kincade PW, Herr JC and White JM:
Role of the integrin-associated protein CD9 in binding between
sperm ADAM 2 and the egg integrin α6β1: Implications for murine
fertilization. Proc Natl Acad Sci USA. 96:11830–11835. 1999.
View Article : Google Scholar
|
|
39
|
Chen H and Sampson NS: Mediation of
sperm-egg fusion: Evidence that mouse egg α6β1 integrin is the
receptor for sperm fertilinbeta. Chem Biol. 6:1–10. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Eto K, Huet C, Tarui T, Kupriyanov S, Liu
HZ, Puzon-McLaughlin W, Zhang XP, Sheppard D, Engvall E and Takada
Y: Functional classification of ADAMs based on a conserved motif
for binding to integrin α 9β 1: Implications for sperm-egg binding
and other cell interactions. J Biol Chem. 277:17804–17810. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu X and Evans JP: Analysis of the roles
of RGD-binding integrins, α(4)/α(9) integrins, α(6) integrins, and
CD9 in the interaction of the fertilin β (ADAM2) disintegrin domain
with the mouse egg membrane. Biol Reprod. 66:1193–1202. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mizuno M, Harris CL, Johnson PM and Morgan
BP: Rat membrane cofactor protein (MCP; CD46) is expressed only in
the acrosome of developing and mature spermatozoa and mediates
binding to immobilized activated C3. Biol Reprod. 71:1374–1383.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lozahic S, Christiansen D, Manié S,
Gerlier D, Billard M, Boucheix C and Rubinstein E: CD46 (membrane
cofactor protein) associates with multiple beta1 integrins and
tetraspans. Eur J Immunol. 30:900–907. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Johnson PM, Clift LE, Andrlíková P,
Jursová M, Flanagan BF, Cummerson JA, Stopka P and
Dvorakova-Hortova K: Rapid sperm acrosome reaction in the absence
of acrosomal CD46 expression in promiscuous field mice (Apodemus).
Reproduction. 134:739–747. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Stein KK, Primakoff P and Myles D:
Sperm-egg fusion: Events at the plasma membrane. J Cell Sci.
117:6269–6274. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jégou A, Ziyyat A, Barraud-Lange V, Perez
E, Wolf JP, Pincet F and Gourier C: CD9 tetraspanin generates
fusion competent sites on the egg membrane for mammalian
fertilization. Proc Natl Acad Sci USA. 108:10946–10951. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kaji K, Oda S, Shikano T, Ohnuki T,
Uematsu Y, Sakagami J, Tada N, Miyazaki S and Kudo A: The gamete
fusion process is defective in eggs of Cd9-deficient mice. Nat
Genet. 24:279–282. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
48
|
Le Naour F, Rubinstein E, Jasmin C,
Prenant M and Boucheix C: Severely reduced female fertility in
CD9-deficient mice. Science. 287:319–321. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Miyado K, Yamada G, Yamada S, Hasuwa H,
Nakamura Y, Ryu F, Suzuki K, Kosai K, Inoue K, Ogura A, et al:
Requirement of CD9 on the egg plasma membrane for fertilization.
Science. 287:321–324. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Horváth G, Serru V, Clay D, Billard M,
Boucheix C and Rubinstein E: CD19 is linked to the
integrin-associated tetraspans CD9, CD81, and CD82. J Biol Chem.
273:30537–30543. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Takahashi Y, Bigler D, Ito Y and White JM:
Sequence-specific interaction between the disintegrin domain of
mouse ADAM 3 and murine eggs: Role of beta1 integrin-associated
proteins CD9, CD81, and CD98. Mol Biol Cell. 12:809–820. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rubinstein E, Ziyyat A, Prenant M, Wrobel
E, Wolf JP, Levy S, Le Naour F and Boucheix C: Reduced fertility of
female mice lacking CD81. Dev Biol. 290:351–358. 2006. View Article : Google Scholar
|
|
53
|
Ernesto JI, Weigel Muñoz M, Battistone MA,
Vasen G, Ma r tínez-L ópez P, Or ta G, Figueiras-Fierro D, De la
Vega-Beltran JL, Moreno IA, Guidobaldi HA, et al: CRISP1 as a novel
CatSper regulator that modulates sperm motility and orientation
during fertilization. J Cell Biol. 210:1213–1224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Armon L, Ben-Ami I, Ron-El R and Eisenbach
M: Human oocyte-derived sperm chemoattractant is a hydrophobic
molecule associated with a carrier protein. Fertil Steril.
102:885–890. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Blasco V, Prados N, Carranza F,
González-Ravina C, Pellicer A and Fernández-Sánchez M: Influence of
follicle rupture and uterine contractions on intrauterine
insemination outcome: A new predictive model. Fertil Steril.
102:1034–1040. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Inoue N, Ikawa M, Isotani A and Okabe M:
The immunoglobulin superfamily protein Izumo is required for sperm
to fuse with eggs. Nature. 434:234–238. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bianchi E, Doe B, Goulding D and Wright
GJ: Juno is the egg Izumo receptor and is essential for mammalian
fertilization. Nature. 508:483–487. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Inoue N, Hamada D, Kamikubo H, Hirata K,
Kataoka M, Yamamoto M, Ikawa M, Okabe M and Hagihara Y: Molecular
dissection of IZUMO1, a sperm protein essential for sperm-egg
fusion. Development. 140:3221–3229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Inoue N, Kasahara T, Ikawa M and Okabe M:
Identification and disruption of sperm-specific angiotensin
converting enzyme-3 (ACE3) in mouse. PLoS One. 5:e103012010.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bianchi E and Wright GJ: Izumo meets Juno:
Preventing polyspermy in fertilization. Cell Cycle. 13:2019–2020.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sawada H, Mino M and Akasaka M: Sperm
proteases and extracellular ubiquitin-proteasome system involved in
fertilization of ascidians and sea urchins. Adv Exp Med Biol.
759:1–11. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhou C, Kang W and Baba T: Functional
characterization of double-knockout mouse sperm lacking SPAM1 and
ACR or SPAM1 and PRSS21 in fertilization. J Reprod Dev. 58:330–337.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yoon S, Chang KT, Cho H, Moon J, Kim JS,
Min SH, Koo DB, Lee SR, Kim SH, Park KE, et al: Characterization of
pig sperm hyaluronidase and improvement of the digestibility of
cumulus cell mass by recombinant pSPAM1 hyaluronidase in an in
vitro fertilization assay. Anim Reprod Sci. 150:107–114. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zumoffen CM, Gil R, Caille AM, Morente C,
Munuce MJ and Ghersevich SA: A protein isolated from human
oviductal tissue in vitro secretion, identified as human
lactoferrin, interacts with spermatozoa and oocytes and modulates
gamete interaction. Hum Reprod. 28:1297–1308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zumoffen CM, Massa E, Caille AM, Munuce MJ
and Ghersevich SA: Effects of lactoferrin, a protein present in the
female reproductive tract, on parameters of human sperm
capacitation and gamete interaction. Andrology. 3:1068–1075. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Tosti E and Ménézo Y: Gamete activation:
Basic knowledge and clinical applications. Hum Reprod Update.
22:420–439. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Nomikos M, Swann K and Lai FA: Starting a
new life: sperm PLC-zeta mobilizes the Ca2+ signal that induces egg
activation and embryo development: an essential phospholipase C
with implications for male infertility. BioEssays. 34:126–134.
2012. View Article : Google Scholar
|
|
68
|
Harada Y, Kawazoe M, Eto Y, Ueno S and
Iwao Y: The Ca2+ increase by the sperm factor in physiologically
polyspermic newt fertilization: Its signaling mechanism in egg
cytoplasm and the species-specificity. Dev Biol. 351:266–276. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Abbott AL and Ducibella T: Calcium and the
control of mammalian cortical granule exocytosis. Front Biosci.
6:D792–D806. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
70
|
Krauchunas AR and Wolfner MF: Molecular
changes during egg activation. Curr Top Dev Biol. 102:267–292.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Charlesworth A, Meijer HA and de Moor CH:
Specificity factors in cytoplasmic polyadenylation. Wiley
Interdiscip Rev RNA. 4:437–461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Sousa Martins JP, Liu X, Oke A, Arora R,
Franciosi F, Viville S, Laird DJ, Fung JC and Conti M: DAZL and
CPEB1 regulate mRNA translation synergistically during oocyte
maturation. J Cell Sci. 129:1271–1282. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gilbert SF: Developmental Biology. 10th
edition. Sinauer Associates; Sunderlan: 2013
|














