Introduction
Osteoporosis is a bone disease which affects bone
strength. It is known to increase the risk of bone fractures and
other complications, and has become a major clinical problem that
affects over 200 million individuals worldwide (1). Current strategies for the treatment
of osteoporosis, include the administration of teriparatide,
strontium and bisphosphonates, mainly aimed at reducing bone
resorption (which is regulated by osteoclasts) and increasing bone
formation (which is regulated by osteoblasts). However, these
synthetic medicines have several side-effects that reduce their
efficacy, such as jaw bone osteonecrosis induced by large doses of
bisphosphonates (which act as inhibitors of bone resorption) or an
increased risk of osteosarcoma induced by prolonged treament with
teriparatide (2,3). Thus, healthier and safer agents that
can be used in the prevention or treatment of bone-related
disorders are highly desired.
Previous studies have indicated that the decreased
bone volume in osteoporosis is associated with increased adipose
tissue in bone marrow (4–6). Osteoblasts and marrow adipocytes
differentiate from a common precursor, namely bone marrow-derived
mesenchymal stem cells (BMSCs) (7). It has been previously demonstrated
that compared with the osteogenic capability of mesenchymal stem
cells (MSCs) derived from healthy women, those derived from
post-menopausal women with osteoporosis have a much lower
osteogenic potential (8,9).
Previous studies have demonstrated a large degree of
plasticity of osteoblasts and adipocytes. Specifically,
fully-differentiated osteoblasts derived from huam MSCs (hMSCs) are
capable of dedifferentiation and transdifferentiation into
adipocytes and vice versa (10,11). There is a reciprocal association
and balance between the differentiation of adipocytes and
osteoblasts (12).
The majority of previous data suggest that bone
marrow adipose tissue simply plays the role of filling the marrow
cavity and lacks hematopoietic function (13,14). However, Elbaz et al
reported a lipotoxic effect from marrow adipocytes on osteoblast
differentiation and function (15). Additionally, free-fatty acids
released by adipocytes can inhibit osteoblast proliferation and
induce osteoblast apoptosis (16).
Existing data demonstrate that an excessive amount
of bone marrow adipocytes may be a significant negative risk factor
for skeletal health (17).
Accordingly, inhibiting adipocyte differentiation in bone marrow,
while simultaneously accelerating osteogenesis may be a therapeutic
approach for age-related osteoporosis.
The fruit of Psoralea corylifolia L. (P.
corylifolia) is a widely used Chinese herbal medicine and is
specifically used in the treatment of fractures, and bone and joint
diseases. It has also used in the treatment of other disorders,
such as skin diseases, cardiovascular diseases, tumors and asthma
(18,19). It has been found that psoralen and
isopsoralen (ISO) have growth inhibitory effects on transplanted
tumors in nude rats with osteosarcoma; however, following the
administration of high doses of psoralen and ISO, toxic reactions
such as writhing, lassitude and hypo-activity were observed
(19). Thus, the efficacy and
safety of any agent in the treatment of osteoprosis should also be
consided.
ISO is the main active ingredient extracted from the
seeds of P. corylifolia (psoralen is its isomer). Previous
studies have proposed a positive role for psoralen in promoting
osteoblast differentiation (20),
as well as its stimulatory effects on bone formation (21,22). However, only a few studies have
evaluated the beneficial effects of ISO on bone development
(23,24), and to the best of our knowledge,
none have examined its underlying mechanisms of action.
In addition, to date, at least to the best of our
knowledge, there are no studies available on the role and
mechanisms of action of ISO in bone marrow adipogenesis. Thus, in
the present study, by employing a mouse model of osteoporosis
induced by ovariectomy along with ISO treatment, we aimed to
examine the effects of ISO on bone marrow adipogenesis in
vitro. Furthermore, we evaluated the effects of ISO on the
differentiation of osteoblasts and adipocytes derived from BMSCs
isolated from C57BL/6 mice in an effort to clarify the probable
underlying cellular and molecular mechanisms that occur during this
period. Our results may contribute to the development of novel
therapeutic approaches for the treatment of bone-related
diseases.
Materials and methods
Ethics approval
Ethics approval was provided by the Medical Ethics
Committee of Southern Medical University, Guangzhou, China.
Materials and reagents
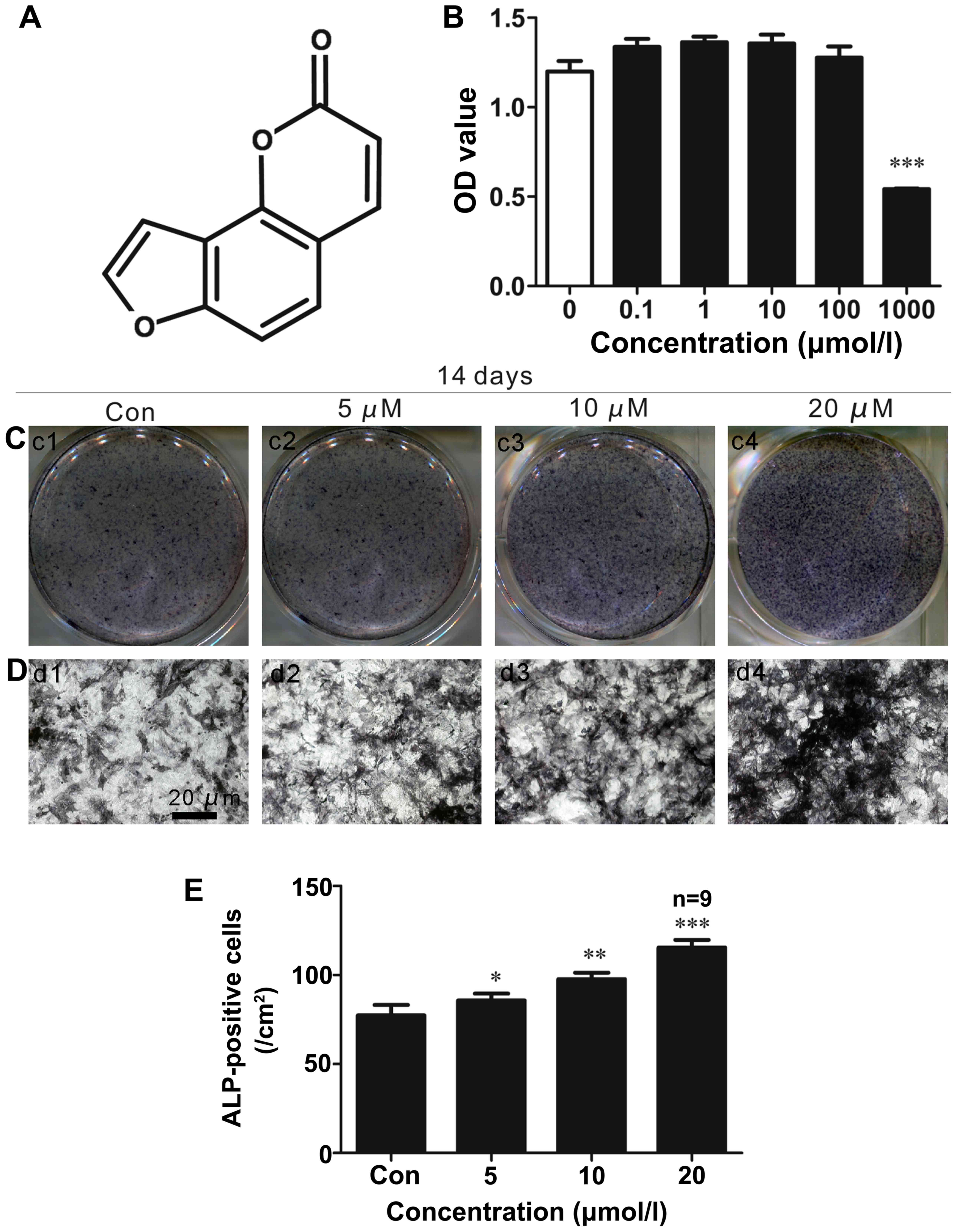
ISO (chemical structure shown in Fig. 1A) was obtained from Sigma (St.
Louis, MO, USA; purity, 99%; molecular weight, 186.1635). Stock
solutions of ISO were prepared in dimethyl sulfoxide (DMSO; Sigma)
and stored at −20°C.
Animal model and animal feeding
Female C57BL/6 mice (aged 2 months; n=18) were
provided by the Experimental Animal Center of Southern Medical
University. The mice were randomly divided into the sham-operated,
ovariectomized (OVX) or OVX plus ISO (n=6/group) groups. The mice
in the OVX plus ISO group were intragastrically administered ISO at
a dose of 20 mg/kg/day for 5 days prior to being subjected to
ovariectomy and this was maintained for 2 months after the mice
were subjected to ovariectomy. A previous study found that a
significant amount of bone loss in vertebrae and femurs was
observed in C57BL/6 mice at only 4 weeks following ovariectomy
(25). Thus, ISO treatment was
maintained for 2 months in this study. The mice were anesthetized
with 1% pentobarbital sodium. The operation area was disinfected by
iodophor. The back skin was then longitudinally cut step by step at
the second lumbar level by one incision (1.5 cm in length) and 2
sides of the ovaries were removed. Ovary peripheral vascular flow
was blocked by ligature using sutures (4-0#). The incision was then
closed and the mice were allowed to recover from the anesthesia at
room temperature. The mice in the sham-operated group only had some
fat tissue around the ovaries removed. The mice were sacrificed by
cervical dislocation. The femurs were then removed using ophthalmic
scissors and fixed in 10% paraformaldehyde solution.
Bone marrow adiposity analyses
For bone marrow adiposity analyses, the distal
portion of the femurs was fixed, decalcified and sectioned into
2-µm-thick sections, and subjected to hematoxylin and eosin
staining according to standard histological protocols. To quantify
proximal metaphyseal adipocyte parameters, we calculated the
adipocyte number (AD#, per mm2). A uniform number of
fields was screened in all sections by 3 authors (Jian Wang,
Sheng-Fa Li and Ting Wang), starting 3 fields from the left end and
3 fields from the top endocortical surface, excluding adipocytes
with disruption in the fields. To avoid any bias in the final
analyses, all sections were interpreted while blinded, without
knowledge of the groups (sham-operated, CON or CON + ISO). Images
were obtained at ×20 magnification using an Olympus BX51RF
stereomicroscope (Olympus).
Microcomputed tomography (μCT)
analyses
Micro-CT (µCT 80; Scanco Medical,
Bassersdorf, Switzerland) analyses were performed on the distal
portion of the femurs. Distal femurs were selected for scanning and
corrected for the CT value, a 70 kV scanning voltage, 30 W power,
429 µA current, and 20 µm scan thickness. Our
analyses included various bone parameters: trabecular thickness
(Tb.Th), bone volume (BV)/total volume (TV) and trabecular number
(Tb.N). The assessment of bone microstructure was carried out
according to the guidelines provided in the study by Bouxsein et
al (26).
Isolation of BMSCs and cell culture
BMSCs were isolated from C57BL/6 mice (n=10; aged 4
weeks; also from the Southern Medical University). The mice were
sacrificed using CO2, and the mouse femurs were
dissected free of surrounding soft tissue. The bone marrow was
flushed with α-MEM (Invitrogen, Carlsbad, CA, USA). The marrow
content from 4 bones was plated in culture flasks containing BMSC
growth medium [α-MEM containing 10% fetal bovine serum (FBS), 100
U/ml penicillin, 100 mg/ml streptomycin sulfate (Gibco, Auckland,
New Zealand). We used a centrifuge (1,000 rpm for 5 min) to isolate
the cells from the extra soft tissue. The cells were re-suspended
and non-adherent cells were removed, and adherent BMSCs were
cultured and expanded for further experiments. The cell culture
medium were replaced every 3 days. The cells were seeded in 96-well
plates and 6-well plates at densities of 1×104 and
1×106 cells/well, respectively, and cultured in a
humidified atmosphere of 5% CO2 and 95% air at 37°C.
After examing the levels of different proteins characteristic of
BMSCs by western blot analysis (see below), we found that the cells
were CD29+ and Sca-1+, and CD45−
and CD11b−; this confirmed these cells were actually
BMSCs.
For osteogenic differentiation, the BMSCs were grown
to 90% confluence in 6-well-plates. The culture medium was replaced
with osteogenic medium (α-MEM supplemented with 15% fetal calf
serum (FCS) plus 1% penicillin/streptomycin, 100 nM dexamethasone,
50 µg/ml ascorbate-2-phosphate and 10 mM β-glycerol
phosphate). The medium was changed every 3 days.
For adipocyte-induced culture, the putative BMSCs
were harvested and seeded into 6-well cell culture plates. Upon
attaining 90% confluency, the cells were treated with adipogenic
differentiation medium: BMSC maintenance medium supplemented with 1
µM dexamethasone, 100 µM indomethacin and 500
µM 1-methyl-3-isobutylxanthine (all from Sigma). For the
initial induction, 5 µg/ml bovine insulin (Sigma) were added
together with adipogenic differentiation medium. The medium was
changed every fourth day.
Cell proliferation assays
Primary BMSCs were seeded in 96-well plates at a
density of 1×104 cells/well. Following culture for 2
days, the cells were treated for 48 h with ISO at concentrations of
0, 0.1, 1, 10, 100 and 1,000 µM. Cell proliferation assays
were performed using Caspase-8 Colorimetric Assay kits (KeyGEN
Biotech, Nanjing, Jiangsu, China) according to the manufacturer's
instructions. The absorbance in the wells was measured at 450
nm.
Alkaline phosphatase (ALP) staining
assay
The BMSCs were cultured in the presence of
osteogenic inducers and ISO (0, 5, 10 and 20 µM) for 2
weeks. ALP activity was also evaluated in the cells stained using
an ALP staining kit (Beyotime, Nanjing, Jiangsu, China) according
to the manufacturer's instructions. For ALP staining, the cells
were washed in phosphate-buffered saline (PBS), fixed in 4%
paraformaldehyde for 20 min at room temperature, and rinsed in
distilled water. The ALP staining mixture was added for 30 min at
room temperature in the dark. The cells were rinsed in distilled
water and PBS to reduce non-specific staining.
Oil Red O staining
The BMSCs were cultured in the presence of
adipogenic inducers and ISO (0, 5, 10 and 20 µM) for 7 and
14 days. Fat droplets that formed in differentiated adipocytes from
the BMSCs were observed using Oil Red O staining. The cells were
fixed in 4% formaldehyde for 15 min at room temperature, washed in
PBS, and stained with 0.6% (w/v) Oil Red O solution (60%
isopropanol, 40% water) for 1 h at 37°C. The cells were washed with
PBS to remove unbound dye and isopropyl alcohol (1 ml) was added to
the culture plates.
Immunocytochemical staining
The murine BMSCs (104 cells/well) were
inoculated into 12-well assay plates with collagen-coated
coverslips. Various concentrations of ISO (0, 5, 10 or 20
µM) were added at room temperature. The cells were fixed in
4% paraformaldehyde for 15 min at room temperature, rinsed with
PBS, and incubated for 1 h with blocking buffer (1X PBST, 0.5%
Triton X-100) supplemented with 5% serum from the same species as
the secondary antibody. The blocking solution was removed and
primary mouse monoclonal antibodies against osteocalcin (OCN; Cat.
no. sc-365797; 1:100) and proliferator-activated receptor γ (PPARγ;
sc-7273; 1:200) (both from Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) were added followed by incubation overnight at 4°C.
Subsequently, the secondary antibody, goat anti-mouse IgG
(SAB4600004; 1:200; Sigma) was added, followed by incubation for 1
h at room temperature. The secondary antibody was aspirated and the
wells were washed 3 times with PBST. DAB (Sangon Biotech, Shanghai,
China) was added followed by incubation for 5 min at room
temperature in the dark. The slides were restained with hematoxylin
(Sigma). Images were acquired using an Olympus BX51RF
stereomicroscope (Olympus, Tokyo, Japan). The mean optical density
(MOD) was examed using Image-Pro Plus software (IPP; Media
Cybernetics, Rockville, MD, USA).
Immunofluorescence staining and
microscopic analyses
The BMSCs were grown on collagen-coated coverslips,
washed with PBS, fixed for 15 min at room temperature in 4%
para-formaldehyde, washed, permeabilized with 0.5% Triton X-100 (5
min, room temperature) and blocked for 30 in blocking buffer (5%
FBS in PBST). The cells were sequentially probed with primary
antibodies against runt-related transcription fact or 2 (RUNX2;
Cat. no. 12556; 1:100) and CCAAT/enhancer binding protein β
(C/EBPβ; Cat. no. 3802; 1:200) (both from Cell Signaling
Technology, Danvers, MA, USA), and washed 3 times with PBST. The
FITC-conjugated sheep anti-rabbi t IgG (Cat. no. F5137; 1:500) and
TRITC-conjugated sheep anti-mouse IgG (Cat. no. T5393; 1:500) (both
from Sigma) were added followed by incubation for 1 h at room
temperature, followed by PBST washes. The cells were incubated with
4′,6-diamidino-2-phenylindole (DAPI; 1 µg/ml) for 15 min and
then rinsed 3 times with PBST. Immunofluorescence staining was also
performed on the femur histological sections using a standard
protocol. Following ovariectomy and ISO treatment, the mice were
then sacrifced. The femurs were fixed in 4% paraformaldehyde in 0.1
M phosphate buffer (pH 7.4). The fixed bones were decalcified by
immersion in 10% EDTA (pH 7.0) for 14 days at room temperature and
embedded in paraffin. The paraffin-embedded longitudinal bone
sections (5-µmp-thick) were blocked in 5% goat serum for 1 h
at room temperature, and incubated overnight with the primery
antibodies. The sections were then incubated with secondary
antibodies for 1 h at room temperature. Femur histological sections
were imaged using a FV1000 confocal microscope and positive cells
were evaluated using the IPP software program. The cells and femur
histologiocal sections were imaged using a laser-scanning confocal
microscope (FV1000; Olympus). Finally, MOD was measured using IPP
software.
Western blot analysis
Western blot analysis was performed using a sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)
electrophoresis system. Protein samples (20 µg) were
resuspended in reduced sample buffer, electrophoresed on a 7.5-10%
Tris gel with Tris running buffer, blotted onto PVDF membranes, and
sequentially probed with primary antibodies against RUNX2 (Cat. no.
12556; 1:1,000; Cell Signaling Technology), OCN (Cat. no.
sc-365797; 1:1,000; Santa Cruz Biotechnology, Inc.), eukaryotic
translation initiation factor 4E-binding protein 1 (4E/BP1;
Thr37/46; Cat. no. 9644; 1:1,000; Cell Signaling Technology),
phospho-S6 ribosomal protein (P-S6; S235/S236; Cat. no. sc-293143;
1:1,000), PPARγ (Cat. no. sc-7273; 1:1,000) (both from Santa Cruz
Biotechnology, Inc.), C/EBPβ (Cat. no. 3802; 1:2,000), Sca-1 (Cat.
no. 9664; 1:4,000), CD29 (Cat. no. 4706; 1:2,000), CD45 (Cat. no.
13917; 1:6,000), CD11b (Cat. no. 14271; 1:3,000) and β-actin (Cat.
no. 3700; 1:2,500) (all from Cell Signaling Technology).
Horseradish peroxidase-conjugated goat anti-rabbit (Cat. no.
SAB4600223; 1:1,000) or anti-mouse (Cat. no. SAB4600004; 1:1,000)
antibodies (both from Sigma) were added, and secondary antibodies
were detected using enhanced chemiluminescence (ECL Plus; General
Electric Healthcare, Milwaukee, WI, USA).
Statistical analyses
We used one-way analyses of variance (ANOVA) to
analyze the data. The homogeneity of the variance tests was used to
evaluate data homogeneity (IBM SPSS Statistics 19.0 software). If
the variance was confirmed to be equal, least-significant
difference tests were used for data analyses. If the variance was
determined to be unequal, Dunnett's T3 tests were used. The results
are presented as the the means ± standard deviation (SD). A value
of P<0.05 was considered to indicate a statistically significant
difference.
Results
Cell proliferation assays
We examined the effects of ISO on the proliferation
of primary mouse BMSCs using CCK8 assays. As shown in Fig. 1B, ISO did not significantly affect
cell growth at concentrations of 1–100 µM after 48 h.
However, BMSC proliferation was significantly inhibited following
treatment with 1,000 µM ISO.
ALP staining in vitro
To determine whether ISO affects BMSC
differentiation into osteoblasts, ALP staining (Fig. 1C–E) was performed as increased ALP
activity is an important indicator of osteoblast differentiation.
We found that ISO enhanced ALP activity in the primary BMSCs in a
dose-dependent manner (Fig.
1C–D). The most significant effects were observed at 2 weeks
after osteogenic induction with a concentration of 20 µM ISO
(Fig. 1E). These results
suggested that ISO signifi-cantly stimulated BMSC differentiation
into osteoblasts.
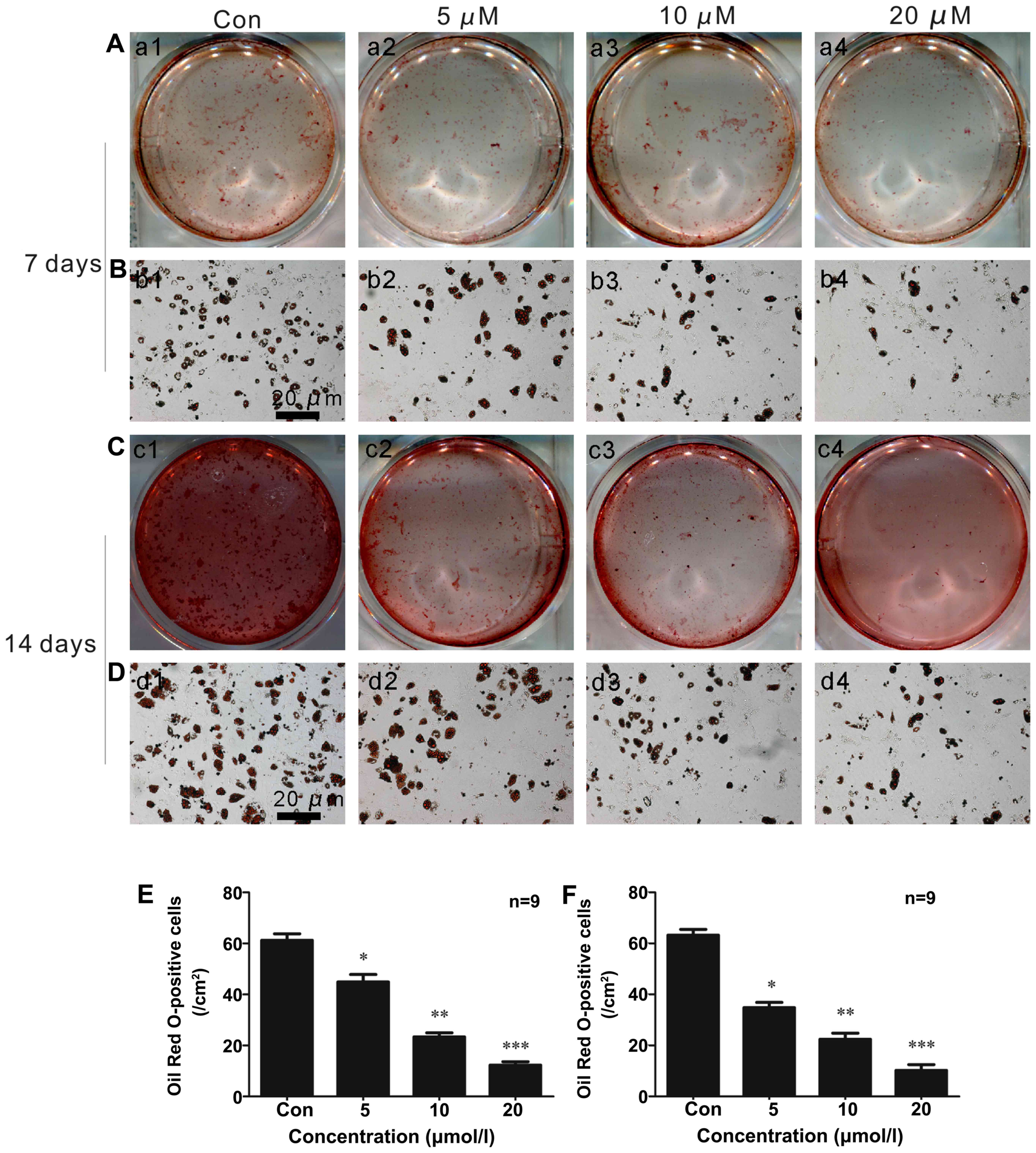
Oil Red O staining in vitro
ISO inhibited the adipocytic differentiation of
BMSCs induced by adipogenic inducers in a concentration-dependent
manner. As shown in Fig. 2A–F,
fewer lipid droplets appeared in the cytoplasm of adipocytes that
were stained with Oil Red O in the cells treated with ISO (Fig. 2E) under adipogenic differentiation
conditions for 7 days (Fig. 2A, B and
E). After 2 weeks of adipocytic differentiation, the adipogenic
rate was at its lowest level in the presence of 20 µM ISO
(Fig. 2C and D and F) compared to
the controls (untreated cells; Fig.
2F). These data suggested that ISO inhibited BMSC
differentiation into adipocytes.
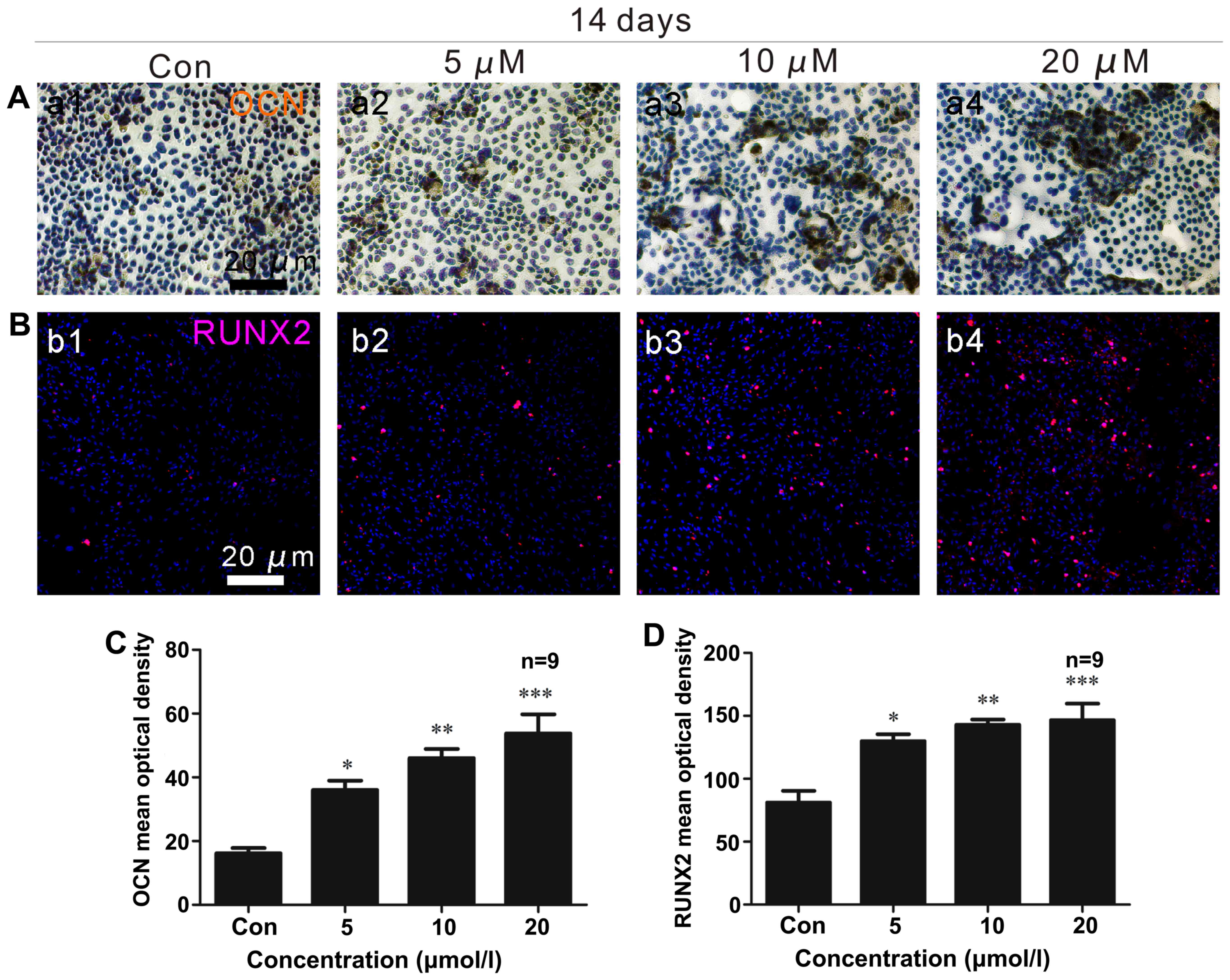
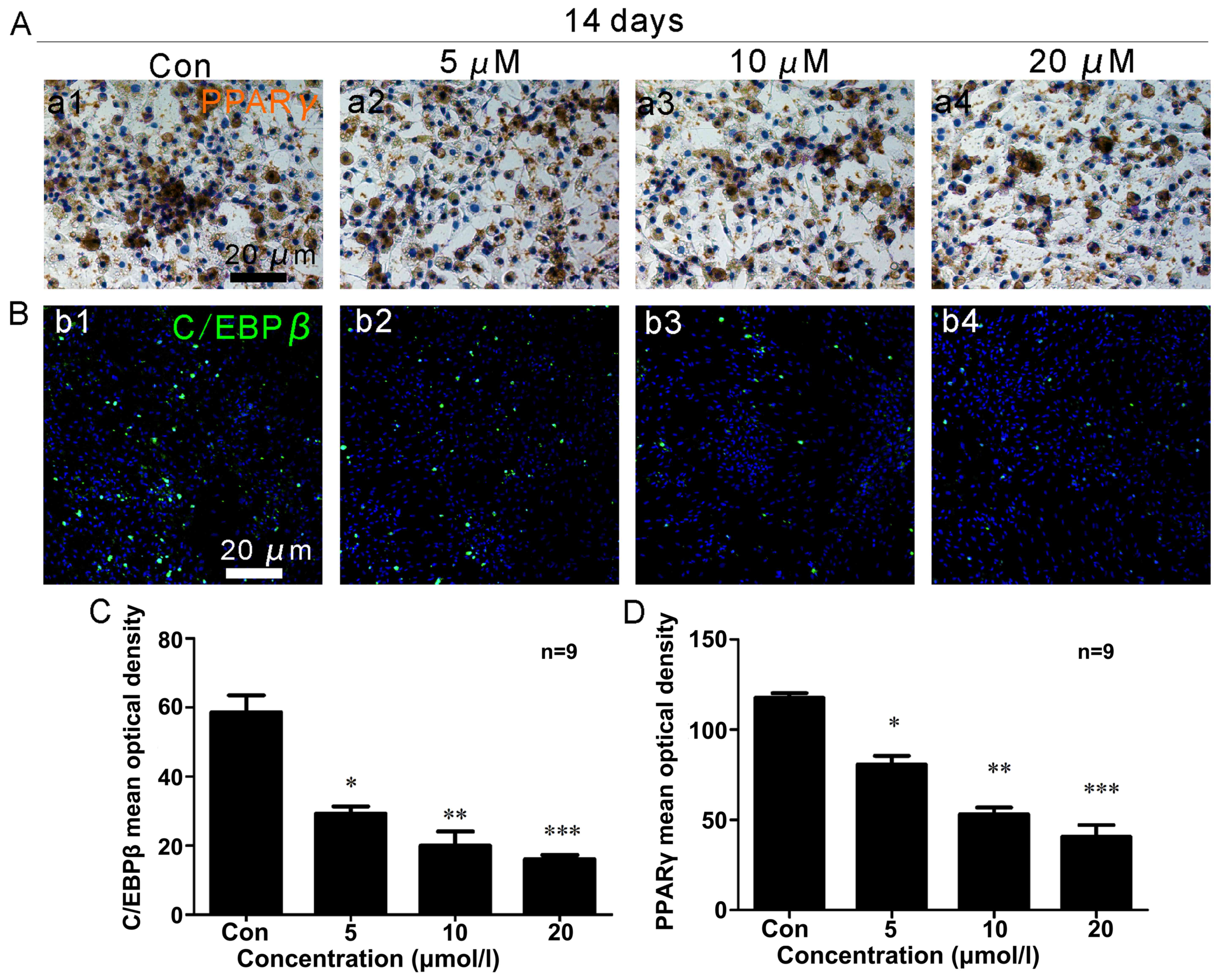
OCN and PPARγ immunocytochemistry
Two weeks following osteoblast differentiation, OCN
expression (brown) increased (Fig. 3A
and C), with a maximum level observed in the cells treated with
a concentration of 20 µM ISO (Fig. 3C). Under adipogenic conditions for
2 weeks, the generation of PPARγ (brown) was significantly reduced
(Fig. 4A and D). The lowest
expression of PPARγ was noted in the presence of 20 µM ISO
(Fig. 4A and D).
RUNX2 and C/EBPβ immunofluorescence
After 2 weeks of osteogenic differentiation, RUNX2
expression (red) in the osteoblasts was upregulated (Fig. 3B and D), and ISO promoted RUNX2
expression in a dose-dependent manner. In the presence of 20
µM ISO, RUNX2 expression was at its maximum level (Fig. 3D). Additionally, after 2 weeks of
adipogenic differentiation, C/EBPβ (green) expression decreased
(Fig. 4B and C). The minimum
density of C/EBPβ staining occurred in the presence of 20 µM
ISO (Fig. 3C).
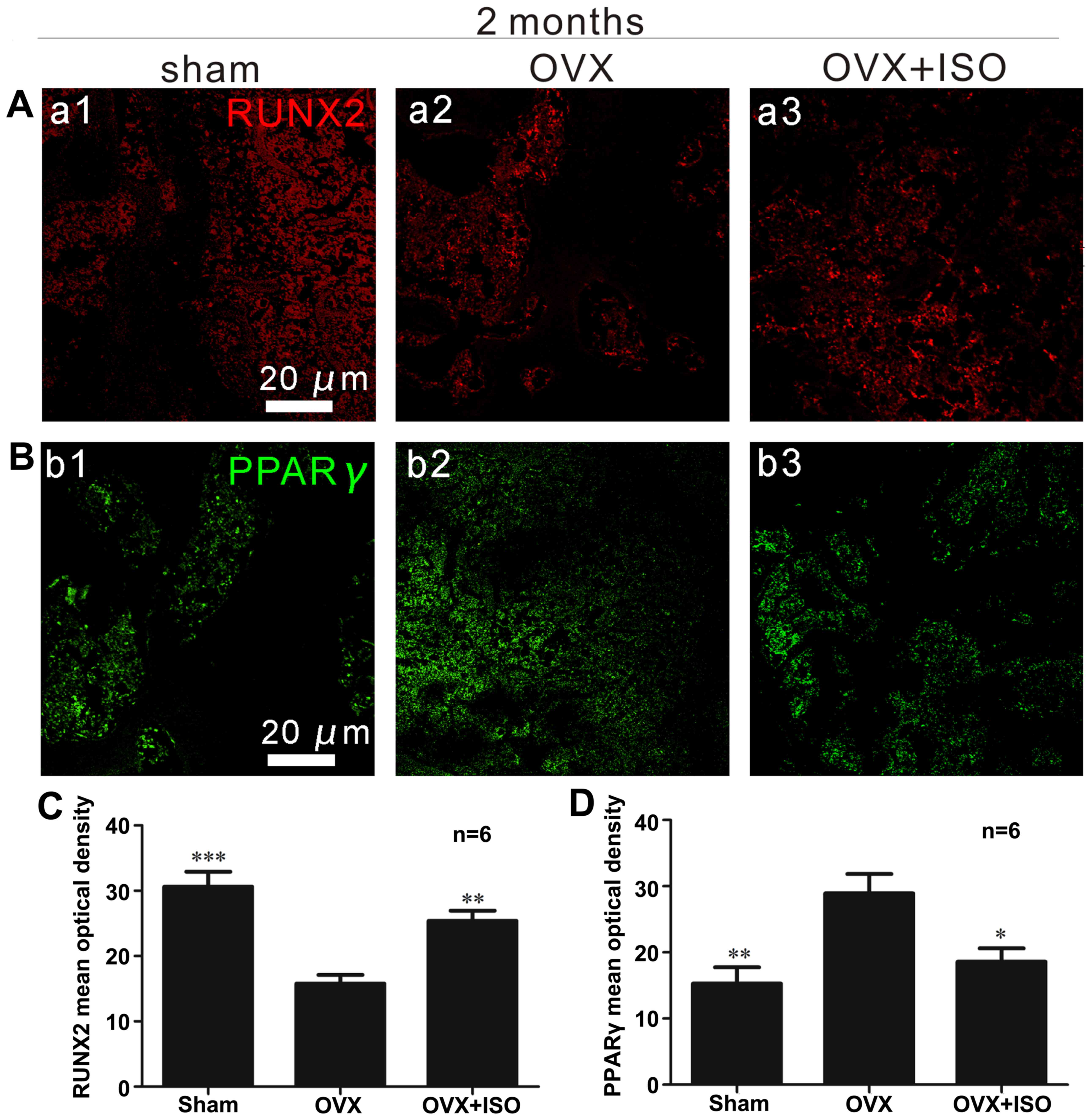
RUNX2 immunofluorescence in vivo
RUNX2 is very important for bone remodeling, and
functions by regulating the differentiation of osteoblasts
(27,37). The intensity of positive RUNX2
staining (red) was reduced in the OVX group (Fig. 5A), whereas it was significantly
elevated in the OVX + ISO group (Fig.
5A and C).
PPARγ immunofluorescence in vivo
Increased PPARγ (green) expression was observed in
the OVX group compared with the OVX + ISO group (Fig. 5B and D). PPARγ expression was
markedly increased in mice in the OVX group (Fig. B, panel b2 and D) compared to the
mice in the OVX + ISO group (Fig. 5B,
panel b3 and D). In addition, there were also statistically
significant differences between the sham-operaged group (Fig. 5B, panel b1 and D) and the OVX
group. PPARγ is considered indispensable for adipocyte
differentiation.
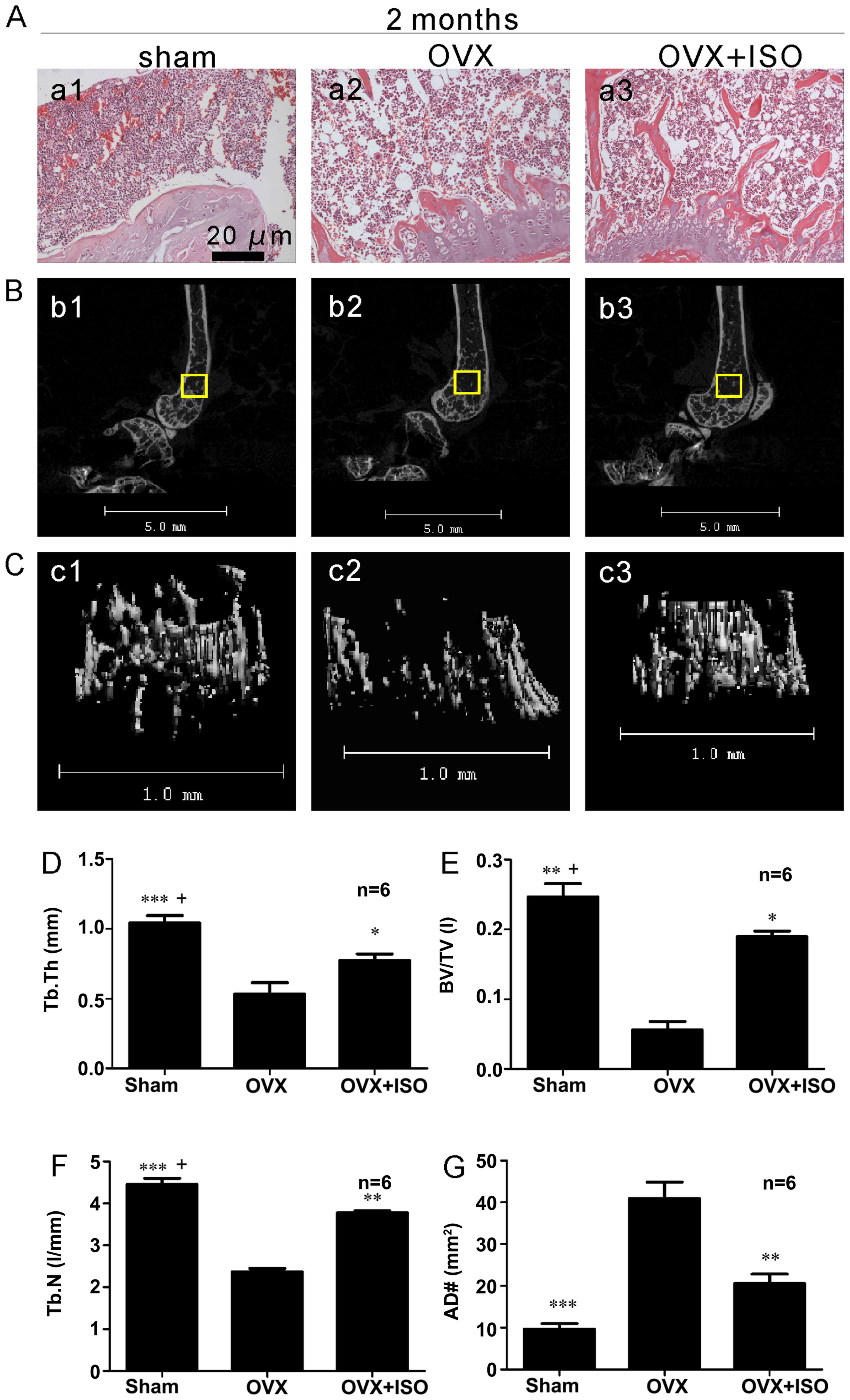
Bone histomorphometries
Representative hematoxylin and eosin images of bone
from the OVX and OVX + ISO groups (Fig. 6A) indicated that ISO reduced the
number of adipocytes (AD#) in the bone marrow (Fig. 6A and G).
Micro-CT analyses
We performed µCT scans on the mouse distal
femurs (Fig. 6B–F). Two months
after the surgery, bone turnover was significantly decreased in the
OVX mice compared with the sham-operated group (Fig. 6D–F). Treatment with ISO treatment
significantly rescued this bone loss. The values for Tb.Th
(Fig. 6D), BV/TV (Fig. 6E) and Tb.N (Fig. 6F) in the OVX group were markedly
decreased compared with the sham-operated group. Following
treatment with ISO for 2 months, these indicators were
significantly improved (Fig. 6D and
E). However, compared to the sham-operated group, the levels of
Tb.N, Tb.Th and BV/TV were still decreased in the OVX + ISO group
(Fig. 6D–F). This result reminded
us of the fact that ISO also cannot be a perfect substitute for
estrogen.
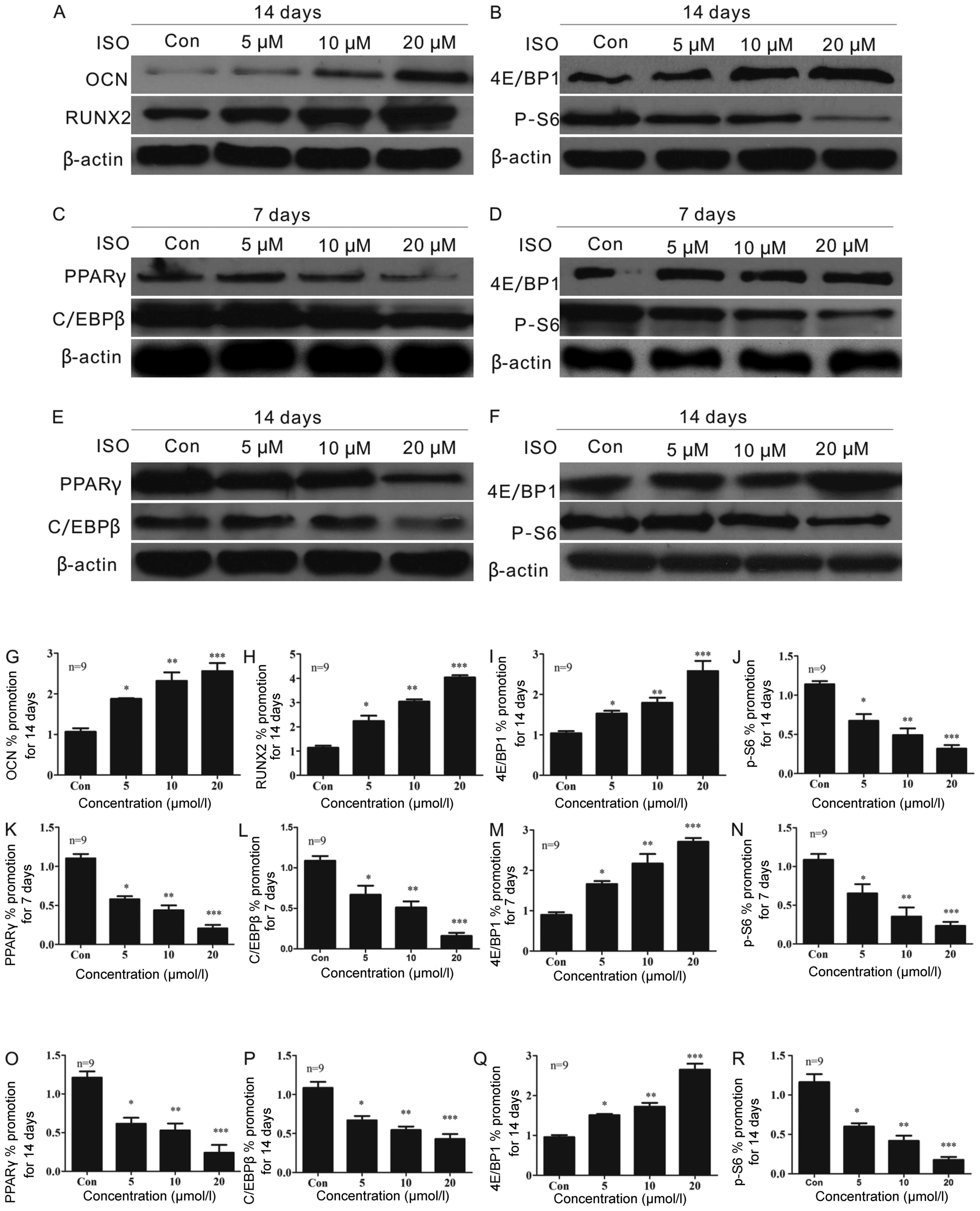
Western blot analysis
To determine whether ISO affects BMSC
differentiation, we performed western blot analysis (Fig. 7). ISO promoted BMSC osteoblast
differentiation in a dose-dependent manner, as demonstrated by the
upregulation of the osteoblast-specific markers, OCN (Fig. 7A and G) and RUNX2 (Fig. 7A and H). OCN activity was
increased in the cells treated with ISO for 2 weeks (Fig. 7A and G). ISO had the most
significant effect on OCN expression at a concentration of 20
µM (Fig. 7G). In addition,
the maximum expression of RUNX2 (Fig.
7A and H) was observed at this concentration of ISO, although
ISO regulated RUNX2 expression in a dose-dependent manner (Fig. 7H).
PPARγ and C/EBPβ are adipocyte-specific markers. One
week after adipogenic induction, PPARγ (Fig. 7C and K) and C/EBPβ (Fig. 7C and L) activity was decreased in
the cells treated with ISO in a dose-dependent manner (Fig. 7K and L). We also found that the
expression of PPARγ (Fig. 7E and
O) and C/EBPβ (Fig. 7E and P)
was markedly decreased in the presence of ISO at day 14 in a
dose-dependent manner (Fig. 7O and
P).
To explore the signaling pathways involved in the
regulatory effects of ISO on BMSC adipogenesis, we assessed
mammalian target of rapamycin complex 1 (mTORC1) signaling. It is
notable that mTORC1 signaling plays an important role in
PPARγ-mediated adipogenesis (28–30). We demonstrated that the
phosphorylation of S6 (S235/236) was inhibited and the
phosphorylation of 4E/BP1 (Thr37/46) was promoted by ISO. These
proteins are direct downstream effectors of mTORC1, suggesting that
ISO may prevent BMSC adipogenesis via the inhibition of mTORC1
signaling. To determine whether the ISO-induced BMSC
differentiation was dependent on the inhibition of mTOR signaling
(Fig. 7B, D and F), we measured
the in vitro expression of 4E/BP1 (Thr37/46) (Fig. 7I, M and Q) and P-S6 (S235/236)
(Fig. 7J, N and R). We observed
an increased 4E/BP1 (Thr37/46) expression (Fig. 7B and I) and a decreased P-S6
(S235/236) expression (Fig. 7B and
J) 2 weeks after osteogenic induction. Importantly, similar
results were observed 7 days after the induction of adipocyte
differentiation (Fig. 7D, M and
N). In adipogenic differentiation conditions and in the
presence of ISO for 2 weeks, we observed that the expression of
4E/BP1 (Thr37/46) (Fig. 7F and Q)
was significantly upregulated and that of P-S6 (S235/236) remained
suppressed (Fig. 7F and R). These
results were observed in a dose-dependent manner (Fig. 7Q and R).
Discussion
The active components extracted from the seeds of
P. corylifolia have been widely investigated, including
psoralen, which has been shown to exert beneficial effects on
skeletal health (20,31,32). Psoralen and ISO are the two main
active ingredients of P.corylifolia fruit extracts. However,
studies evaluating the protective effects of ISO against
osteoporosis are limited, particularly studies that investigate its
cellular and molecular mechanisms (19,23,33).
In this study, we demonstrated that 2 months of ISO
treatment (20 mg/kg/day) increased the bone mass of the distal
femoral metaphysis in OVX mice (Tb.Th, BV/TV and Tb.N). The above
bone metabolism parameters (BV/TV, Tb.Th and Tb.N) are usually
important indicators reflect the level of osteoporosis in
vivo. Based on the close association between bone marrow
adipogenesis and bone loss during the pathogenesis of osteoporosis,
the effects of ISO on bone marrow were evaluated using histological
and immunofluorescence analyses. In ISO-treated OVX mice, we
observed increased trabecular bone in parallel with reduced adipose
tissue in bone marrow. Moreover, an enhanced RUNX2 secretion and
decreased PPARγ secretion were observed.
PPARγ and C/EBPβ are essential transcription factors
for adipogenesis (34). Its
expression and/or activity determines the commitment of BMSCs into
the osteoblasts or adipocyte lineage (35). ALP, RUNX2 and OCN are key
regulators of osteoblast differentiation and play an important role
in bone formation (36). We used
the above parameters as measurements of osteogenesis in the present
study.
To the best of our knowledge, this is the first
study to demonstrate a role for ISO in the promotion of
osteogenesis and the attenuation of adipogenesis in vivo.
This mechanism of action, which focuses on BMSCs, differs from the
mechanisms of currently available agents for osteoporosis that
target mature osteoblasts and osteoclasts (37).
We then found that the ISO-enhanced ALP activity is
a dose-dependent osteogenic inducer of BMSCs that does not affect
cell growth in bone marrow. This result is similar to that of
previous studies on psoralen (20,38). As previously demonstrated, in
patients with age-related osteoporosis, BMSCs in bone marrow
preferentially differentiate into adipocytes rather than
osteoblasts (8,39). Accordingly, in this study, BMSCs
were cultured under adipogenic conditions, which mimics
osteoporosis in humans. Our data indicated that ISO inhibited BMSC
adipogenesis in a dose-dependent manner. The pro-osteogenic and
anti-adipogenic effects of ISO on BMSCs were further confirmed by
evaluating the expression levels of key osteogenic and adipogenic
transcription factors. As we had hypothesized, our results
indicated that treatment with ISO increased RUNX2 expression. By
contrast, PPARγ expression was reduced by treatment with ISO. These
findings were consistent with the data from our in vitro
experiments.
Based on the observation that decreased numbers of
osteoblastic cells occur concomitantly with increased fat content
in bone marrow during aging and osteoporosis, studies have
suggested a reciprocal association between the adipocyte
differentiation and osteoblast differentiation of BMSCs (40–42).
As osteoblasts and adipocytes are both derived from
BMSCs (43), we hypothesized that
ISO may exert its anti-osteoporotic effects under low estrogen
conditions by modulating the RUNX2/PPARγ balance, thereby
inhibiting the differentiation of BMSCs towards adipocytes
(7).
There are several limitations to the current study.
First, we did not measure the serum lipid levels of mice in our
in vivo experiments. However, previous studies have
demonstrated that compared with other fat types, alterations in
serum lipid levels do not significantly affect the role and
function of fat in bone marrow (44,45). Additionally, although our results
have indicated a protective effect of ISO on osteoporotic bone and
bone marrow adipose tissue, we did not include a positive control
group, such as strontium and estrogen, which prevent bone loss due
to the inhibition of bone marrow adipogenesis (46,47).
In conclusion, the present study demonstrated that
ISO attenuated bone marrow adipogenesis, which was indicated by
increased RUNX2 levels and decreased PPARγ levels. This shifts BMSC
lineage differentiation toward osteoblasts rather than adipocytes.
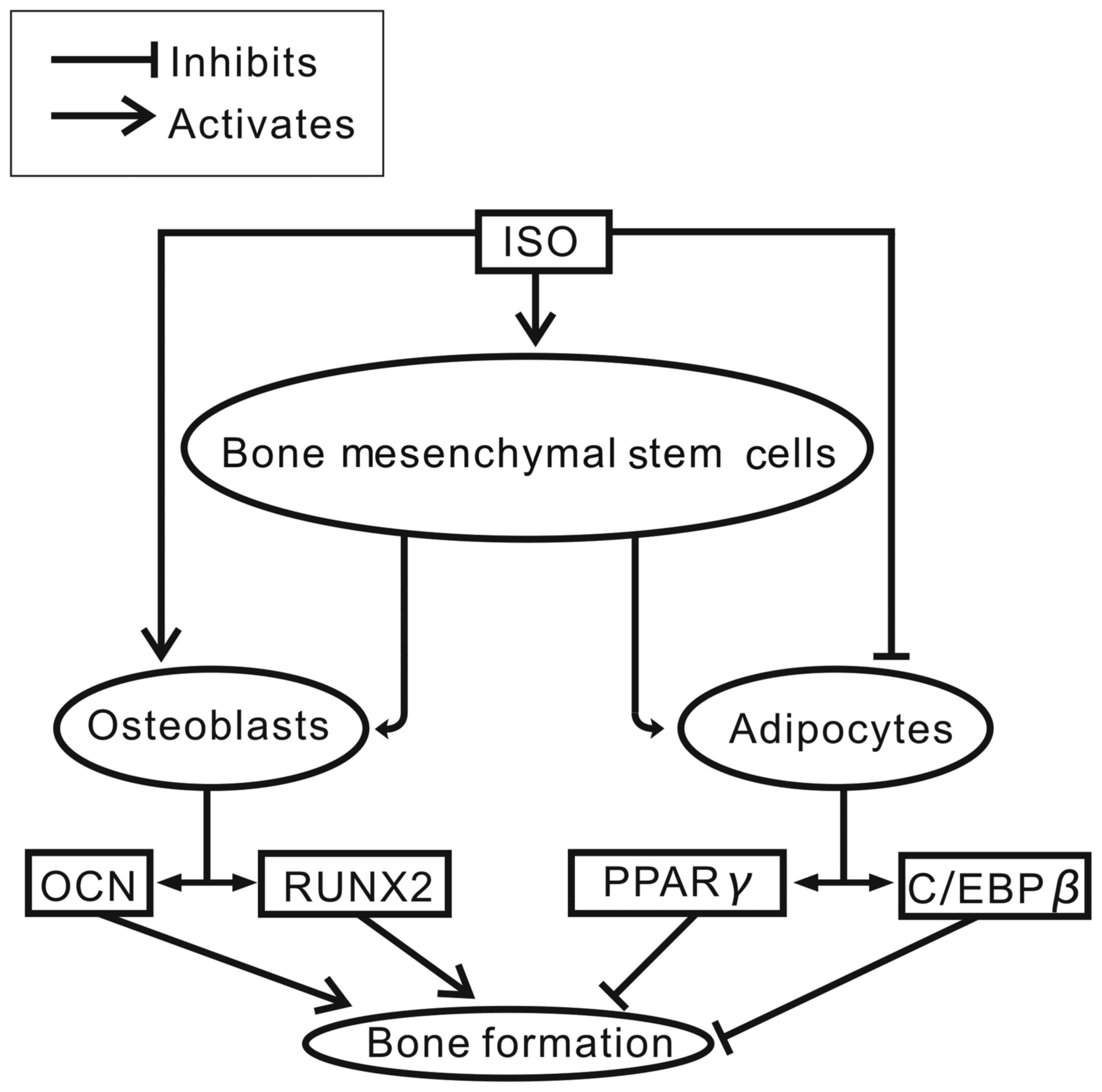
Moreover, we suggest that mTORC1 signaling may be the underlying
signaling pathway involved in this process (Fig. 8). Our data suggest that the
naturally occurring agent, ISO, is safer and more cost-effective
for use in the treatment of post-menopausal bone-related
diseases.
Acknowledgments
We would like to thank Dr Xin Wang (Guangzhou Huayin
Medical Technology Co., Guangdong, China) for providing excellent
technical support with the micro-CT analysis. This study was
supported by the Science and Technology Department of Hunan
Province (grant no. 2014FJ4233) and the Education Department of
Hunan Province (grant no. 14C0914).
References
|
1
|
Schurman L, Bagur A, Claus-Hermberg H,
Messina OD, Negri AL, Sánchez A, González C, Diehl M, Rey P, Gamba
J, et al: Guías 2012 para el diagnóstico, la prevención y el
tratamiento de la osteoporosis. Medicina (B Aires). 73:55–74.
2013.In Spanish.
|
|
2
|
Wang ZQ, Li JL, Sun YL, Yao M, Gao J, Yang
Z, Shi Q, Cui XJ and Wang YJ: Chinese herbal medicine for
osteoporosis: A systematic review of randomized controlled trails.
Evid Based Complement Alternat Med. 2013:3562602013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tastekin N and Zateri C: Probable
osteosarcoma risk after prolonged teriparatide treatment: comment
on the article by Saag et al. Arthritis Rheum. 62:1837–1838. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nuttall ME and Gimble JM: Is there a
therapeutic opportunity to either prevent or treat osteopenic
disorders by inhibiting marrow adipogenesis. Bone. 27:177–184.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Burkhardt R, Kettner G, Böhm W,
Schmidmeier M, Schlag R, Frisch B, Mallmann B, Eisenmenger W and
Gilg T: Changes in trabecular bone, hematopoiesis and bone marrow
vessels in aplastic anemia, primary osteoporosis, and old age: A
comparative histomorphometric study. Bone. 8:157–164. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Griffith JF, Yeung DK, Antonio GE, Wong
SY, Kwok TC, Woo J and Leung PC: Vertebral marrow fat content and
diffusion and perfusion indexes in women with varying bone density:
MR evaluation. Radiology. 241:831–838. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Majors AK, Boehm CA, Nitto H, Midura RJ
and Muschler GF: Characterization of human bone marrow stromal
cells with respect to osteoblastic differentiation. J Orthop Res.
15:546–557. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rodríguez JP, Montecinos L, Ríos S, Reyes
P and Martínez J: Mesenchymal stem cells from osteoporotic patients
produce a type I collagen-deficient extracellular matrix favoring
adipogenic differentiation. J Cell Biochem. 79:557–565. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sekiya I, Larson BL, Vuoristo JT, Cui JG
and Prockop DJ: Adipogenic differentiation of human adult stem
cells from bone marrow stroma (MSCs). J Bone Miner Res. 19:256–264.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Song L and Tuan RS: Transdifferentiation
potential of human mesenchymal stem cells derived from bone marrow.
FASEB J. 18:980–982. 2004.PubMed/NCBI
|
|
11
|
Park SR, Oreffo RO and Triffitt JT:
Interconversion potential of cloned human marrow adipocytes in
vitro. Bone. 24:549–554. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Beresford JN, Bennett JH, Devlin C, Leboy
PS and Owen ME: Evidence for an inverse relationship between the
differentiation of adipocytic and osteogenic cells in rat marrow
stromal cell cultures. J Cell Sci. 102:341–351. 1992.PubMed/NCBI
|
|
13
|
Gimble JM: The function of adipocytes in
the bone marrow stroma. New Biol. 2:304–312. 1990.PubMed/NCBI
|
|
14
|
Tavassoli M: Marrow adipose cells and
hemopoiesis: An interpretative review. Exp Hematol. 12:139–146.
1984.PubMed/NCBI
|
|
15
|
Elbaz A, Wu X, Rivas D, Gimble JM and
Duque G: Inhibition of fatty acid biosynthesis prevents adipocyte
lipotoxicity on human osteoblasts in vitro. J Cell Mol Med.
14:982–991. 2010. View Article : Google Scholar :
|
|
16
|
Dong X, Bi L, He S, Meng G, Wei B, Jia S
and Liu J: FFAs-ROS-ERK/P38 pathway plays a key role in adipocyte
lipotoxicity on osteoblasts in co-culture. Biochimie. 101:123–131.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Greco EA, Lenzi A and Migliaccio S: The
obesity of bone. Ther Adv Endocrinol Metab. 6:273–286. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pae HO, Cho H, Oh GS, Kim NY, Song EK, Kim
YC, Yun YG, Kang CL, Kim JD, Kim JM and Chung HT: Bakuchiol from
Psoralea corylifolia inhibits the expression of inducible nitric
oxide synthase gene via the inactivation of nuclear transcription
factor-kappaB in RAW 264.7 macrophages. Int Immunopharmacol.
1:1849–1855. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu H, Zhang L, Liu D, Tang P and Song F:
Isolation and purification of psoralen and isopsoralen and their
efficacy and safety in the treatment of osteosarcoma in nude rats.
Afr Health Sci. 14:641–647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang DZ, Yang F, Yang Z, Huang J, Shi Q,
Chen D and Wang YJ: Psoralen stimulates osteoblast differentiation
through activation of BMP signaling. Biochem Biophys Res Commun.
405:256–261. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wong RW and Rabie AB: Effect of psoralen
on bone formation. J Orthop Res. 29:158–164. 2011. View Article : Google Scholar
|
|
22
|
Tsai MH, Huang GS, Hung YC, Bin L, Liao LT
and Lin LW: Psoralea corylifolia extract ameliorates experimental
osteoporosis in ovariectomized rats. Am J Chin Med. 35:669–680.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ming L, Ge B, Chen K, Ma H and Zhai Y:
Effects of isopsoralen on bone marrow stromal stem cells
differentiate and proliferate in vitro. Zhongguo Zhong Yao Za Zhi.
36:2124–2128. 2011.In Chinese. PubMed/NCBI
|
|
24
|
Ming LG, Cheng KM, Ge BF, Ma HP and Zai
YK: Effect of isopsoralen on the proliferation and differentiate of
osteoblasts in vitro. Zhong Yao Cai. 34:404–408. 2011.In Chinese.
PubMed/NCBI
|
|
25
|
Lee SK, Kalinowski JF, Jacquin C, Adams
DJ, Gronowicz G and Lorenzo JA: Interleukin-7 influences osteoclast
function in vivo but is not a critical factor in
ovariectomy-induced bone loss. J Bone Miner Res. 21:695–702. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bouxsein ML, Boyd SK, Christiansen BA,
Guldberg RE, Jepsen KJ and Muller R: Guidelines for assessment of
bone microstructure in rodents using micro-computed tomography. J
Bone Miner Res. 25:1468–1486. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rossini M, Gatti D and Adami S:
Involvement of WNT/β-catenin signaling in the treatment of
osteoporosis. Calcif Tissue Int. 93:121–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cybulski N, Polak P, Auwerx J, Rüegg MA
and Hall MN: mTOR complex 2 in adipose tissue negatively controls
whole-body growth. Proc Natl Acad Sci USA. 106:9902–9907. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiang X, Yuan F, Zhao J, Li Z, Wang X,
Guan Y, Tang C, Sun G, Li Y and Zhang W: Deficiency in pulmonary
surfactant proteins in mice with fatty acid binding protein
4-Cre-mediated knockout of the tuberous sclerosis complex 1 gene.
Exp Physiol. 98:830–841. 2013. View Article : Google Scholar :
|
|
30
|
Polak P, Cybulski N, Feige JN, Auwerx J,
Rüegg MA and Hall MN: Adipose-specific knockout of raptor results
in lean mice with enhanced mitochondrial respiration. Cell Metab.
8:399–410. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lim SH, Ha TY, Kim SR, Ahn J, Park HJ and
Kim S: Ethanol extract of Psoralea corylifolia L. and its main
constituent, bakuchiol, reduce bone loss in ovariectomised
Sprague-Dawley rats. Br J Nutr. 101:1031–1039. 2009. View Article : Google Scholar
|
|
32
|
Wang D, Li F and Jiang Z: Osteoblastic
proliferation stimulating activity of Psoralea corylifolia extracts
and two of its flavonoids. Planta Med. 67:748–749. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yuan X, Bi Y, Yan Z, Pu W, Li Y and Zhou
K: Psoralen and isopsoralen ameliorate sex hormone
deficiency-induced osteoporosis in female and male mice. Biomed Res
Int. 2016:68694522016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wood RJ: Vitamin D and adipogenesis: New
molecular insights. Nutr Rev. 66:40–46. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Moerman EJ1, Teng K, Lipschitz DA and
Lecka-Czernik B: Aging activates adipogenic and suppresses
osteogenic programs in mesenchymal marrow stroma/stem cells: the
role of PPAR-gamma2 transcription factor and TGF-beta/BMP signaling
pathways. Aging Cell. 3:379–389. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yoshida CA, Furuichi T, Fujita T, Fukuyama
R, Kanatani N, Kobayashi S, Satake M, Takada K and Komori T:
Core-binding factor beta interacts with Runx2 and is required for
skeletal development. Nat Genet. 32:633–638. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Peng S, Zhang G, He Y, Wang X, Leung P,
Leung K and Qin L: Epimedium-derived flavonoids promote
osteoblastogenesis and suppress adipogenesis in bone marrow stromal
cells while exerting an anabolic effect on osteoporotic bone. Bone.
45:534–544. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang Z, Huang JH, Liu SF, Zhao YJ, Shen
ZY, Wang YJ and Bian Q: The osteoprotective effect of psoralen in
ovariectomy-induced osteoporotic rats via stimulating the
osteoblastic differentiation from bone mesenchymal stem cells.
Menopause. 19:1156–1164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Duque G: Bone and fat connection in aging
bone. Curr Opin Rheumatol. 20:429–434. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chan GK and Duque G: Age-related bone
loss: Old bone, new facts. Gerontology. 48:62–71. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Verma S, Rajaratnam JH, Denton J, Hoyland
JA and Byers RJ: Adipocytic proportion of bone marrow is inversely
related to bone formation in osteoporosis. J Clin Pathol.
55:693–698. 2005. View Article : Google Scholar
|
|
42
|
Rodríguez JP, Garat S, Gajardo H, Pino AM
and Seitz G: Abnormal osteogenesis in osteoporotic patients is
reflected by altered mesenchymal stem cells dynamics. J Cell
Biochem. 75:414–423. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Owen M: Marrow stromal stem cells. J Cell
Sci Suppl. 10:63–76. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gasparrini M, Rivas D, Elbaz A and Duque
G: Differential expression of cytokines in subcutaneous and marrow
fat of aging C57BL/6J mice. Exp Gerontol. 44:613–618. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ecklund K, Vajapeyam S, Feldman HA, Buzney
CD, Mulkern RV, Kleinman PK, Rosen CJ and Gordon CM: Bone marrow
changes in adolescent girls with anorexia nervosa. J Bone Miner
Res. 25:298–304. 2010. View Article : Google Scholar
|
|
46
|
Fournier C, Perrier A, Thomas M, Laroche
N, Dumas V, Rattner A, Vico L and Guignandon A: Reduction by
strontium of the bone marrow adiposity in mice and repression of
the adipogenic commitment of multipotent C3H10T1/2 cells. Bone.
50:499–509. 2012. View Article : Google Scholar
|
|
47
|
Syed FA, Oursler MJ, Hefferanm TE,
Peterson JM, Riggs BL and Khosla S: Effects of estrogen therapy on
bone marrow adipocytes in postmenopausal osteoporotic women.
Osteoporos Int. 9:1323–1330. 2008. View Article : Google Scholar
|