Introduction
Sepsis is a life-threatening organ dysfunction
caused by a dysregulated host response to infection (1). In response to invading
microorganisms and their products [e.g., lipopolysaccharide (LPS)],
immune cells produce a broad array of inflammatory mediators,
including cytokines (2).
Cytokines mediate a variety of responses; for example, interleukin
(IL)-1β induces the expression of adhesion molecules on endothelial
cells to promote the recruitment of inflammatory cells at the site
of inflammation (3). Notably,
inflammatory cytokines are overwhelmingly produced during sepsis,
and the dysregulated production of cytokines can lead to
hypotension, multiple organ failure and ultimately death (2,4).
In recent years, it has been reported that cell death contributes
to the production of cytokines in sepsis (5,6),
and that inhibition of cell death (e.g., pyroptosis) reduces the
production of cytokines and improves the survival of septic models
(5–8). Thus, much attention has been focused
on the relationship between the production of cytokines and host
cell death during sepsis.
Neutrophils, the most abundant leukocytes in humans,
defend against invading microorganisms and are considered as an
essential part of the innate immune system (9,10).
Neutrophils exhibit antimicrobial potential by the ability to
phagocytose microorganisms and release antimicrobial agents (e.g.,
antimicrobial peptides and lytic enzymes) (9). Moreover, upon exposure to bacteria,
neutrophils undergo NETosis (a type of programmed cell death) and
release neutrophil extracellular traps (NETs) (11). Notably, NETs are described as an
antimicrobial mechanism, based on the fact that NETs can trap
microorganisms and exert bactericidal activity by the action of
NET-associated components [e.g., antimicrobial peptides and
neutrophil elastase (NE)] (12,13). In contrast, the major components
of NETs (histone, DNA and granule proteins) are recognized as
damage-associated molecular pattern molecules (DAMPs), which
trigger inflammatory signals to induce cell death, inflammation and
organ failure (14–18); for example, extracellular histones
were found to mediate mortality in mouse fatal liver injury
(17). Importantly, NETs,
recognized as a complex of DAMPs, were found to induce the cell
death of epithelial and endothelial cells (15), and provide a scaffold for platelet
binding and aggregation to form the thrombosis in sepsis (19). Moreover, NETs activate macrophages
to induce cytokine production in atherosclerosis (20).
However, it has not been clarified how NETs affect
and induce the production of cytokines by macrophages in sepsis.
Therefore, in the present study, we utilized LPS (the component of
Gram-negative bacteria) as a stimulus to mimic the septic
environment, and treated mouse macrophage-like J774 cells with NETs
in combination with LPS. The results indicated that NETs
significantly induced the production of IL-1β by J774 cells in the
presence of LPS. Notably, the NET/LPS-induced IL-1β production was
inhibited by both caspase-1 and caspase-8 inhibitors. Moreover,
nucleases and serine protease inhibitors significantly inhibited
the NET/LPS-induced IL-1β production. These observations suggest
that NETs induce the production of IL-1β from LPS-stimulated
macrophages via the caspase-1 and caspase-8 pathways, and
NET-associated DNA and serine proteases are likely involved in the
NET/LPS-induced IL-1β production as essential components.
Materials and methods
Reagents
Ac-YVAD (Tyr-Val-Ala-Asp)-CHO was purchased from
Peptide Institute (Osaka, Japan). Ac-IETD (Ile-Glu-Thr-Asp)-CHO was
purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). Z-VAD (Val-Ala-Asp)-FMK was purchased from InvivoGen (San
Diego, CA, USA). Deoxyribonuclease I (DNase I) and micrococcal
nuclease (MNase) were purchased from Takara Bio, Inc. (Shiga,
Japan). LPS (Escherichia coli B55:05),
4-(2-aminoethyl)benzenesulfonyl fluoride hydrochloride (ABESF),
α1-anti-trypsin, calf thymus (CT)-histone, CT-DNA,
N-acetyl-L-cysteine (NAC) [a reactive oxygen species (ROS)
scavenger] and neutrophil elastase (elastase from human leukocytes)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). Mouse
histone H2AX monoclonal antibody (MAB3406) and mouse histone H4
polyclonal antibody (AF5215) were purchased from R&D Systems,
Inc. (Minneapolis, MN, USA), and used for the neutralization of the
NET-associated histones.
Mice
Male BALB/c mice (15 weeks old; Sankyo Labo Service,
Tokyo, Japan) were used in the experiments. Mice were bred under
specific pathogen-free (SPF) conditions, and housed in
temperature-controlled, air-conditioned facilities with a 12/12-h
light/dark cycle, and food and water ad libitum. All
experiments were approved by the Ethics Committee for the Use of
Laboratory Animals of Juntendo University, Graduate School of
Medicine (Tokyo, Japan; permit no. 270193).
Isolation of neutrophils
Murine bone marrow-derived neutrophils were
harvested from BALB/c mice. Briefly, the tibia and femur were
isolated and flushed with RPMI-1640 medium (Sigma-Aldrich). The red
blood cells were then removed from bone marrow cells by hypotonic
lysis (0.2% NaCl), and the resultant cell suspension was
centrifuged at 600 × g for 5 min. The cell pellet was resuspended
in PBS, and further centrifuged (1,600 × g, 30 min) over 62%
Percoll (GE Healthcare Life Sciences, Marlborough, MA, USA). Murine
neutrophils at the bottom of a 62% layer were collected and washed
in PBS (21). Neutrophils were
resuspended in RMPI-1640 medium at a cell density of
1.7×106 cells/ml.
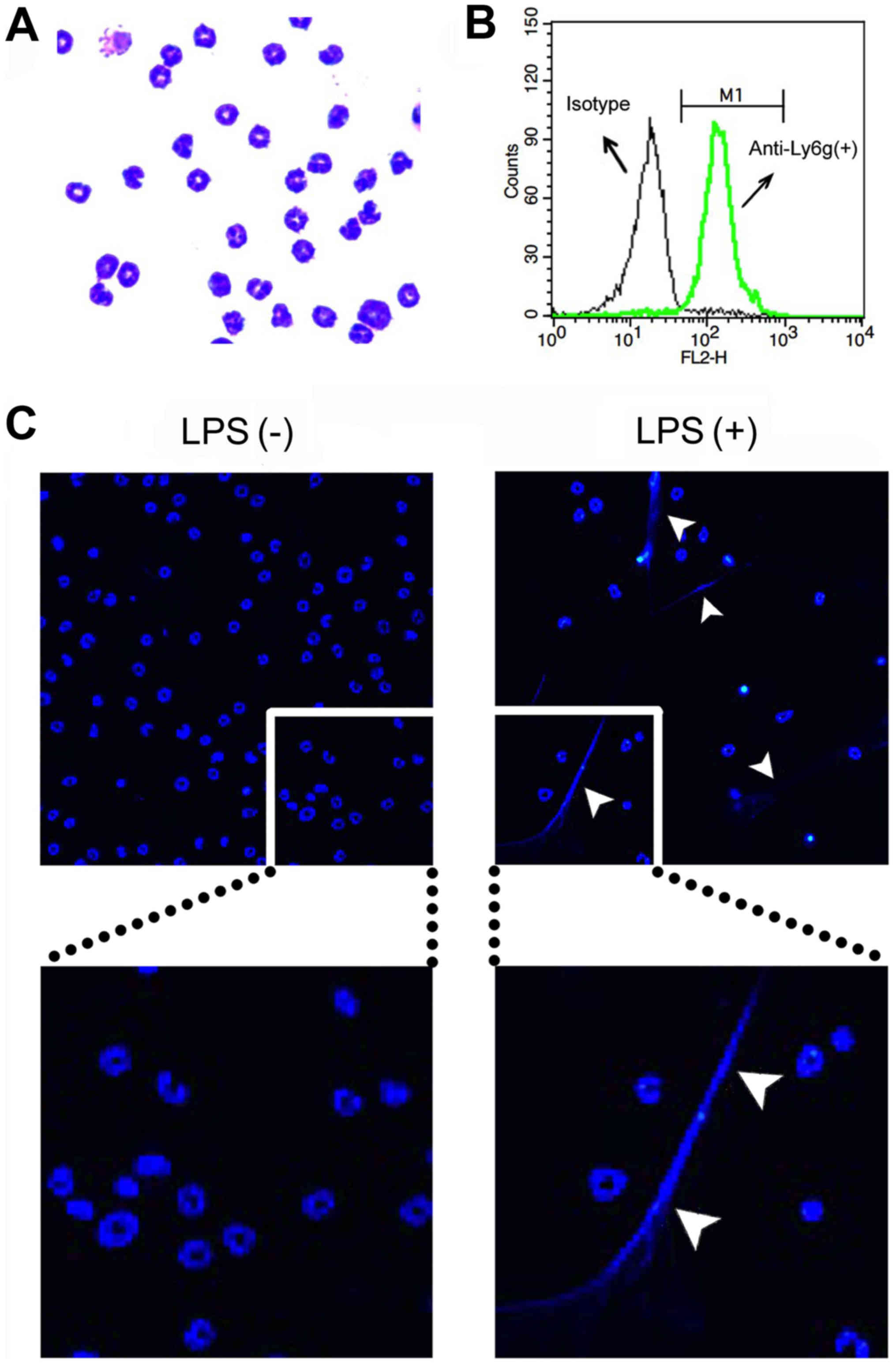
Purities of the neutrophils were determined by both
cytospin counts with Giemsa stain (Wako Pure Chemical Industries,
Ltd., Osaka, Japan) and forward scatter/side scatter gating of
cells stained with PE/Cy5-conjugated anti-Ly6g (a
neutrophil-specific marker) monoclonal antibody (RB6-8C5; Abcam,
Cambridge, MA, USA) [isotype control, a PE/Cy5-conjugated rat IgG2b
monoclonal antibody (eB149/10H5), eBioscience, Inc., San Diego, CA,
USA] using a flow cytometer (FACSCalibur; BD Biosciences, Franklin
Lakes, NJ, USA). The neutrophil fraction contained >90%
neutrophils as determined by Giemsa staining (Fig. 1A) and flow cytometry (Ly6g
staining) (Fig. 1B).
Formation and preparation of NETs
To evaluate NET formation by microscopy, neutrophils
(1.3×106 cells/well) were seeded over a 12-mm glass
coverslip (Thermo Fisher Scientific, Pittsburg, PA, USA) in 24-well
plates (Iwaki Brand; Asahi Techno Glass Corporation, Tokyo, Japan)
for 30 min. After adhesion, the cells were treated with 200 ng/ml
LPS for 4 h to induce NETs. Then, the cells were washed with
RPMI-1640 medium and stained with Hoechst 33342 (0.8 µg/ml).
Thereafter, the coverslips were mounted on a slide glass by using
aqueous medium Vectashield (Vector Laboratories, Inc., Burlingame,
CA, USA), and the cells were photographed using a fluorescence
microscope system Axioplan 2 (Carl Zeiss, Jena, Germany) (Fig. 1C).
To prepare NETs as a stimulant, neutrophils
(1.7×106 cells/well) were seeded in 6-well plates for 30
min in a total volume of 1 ml RPMI-1640 medium, and then were
treated with 200 ng/ml LPS for 4 h to induce NETs. Then, the cells
were washed twice with fresh PRMI-1640 medium, and the NETs were
collected by extensively pipetting with 1 ml RPMI-1640 medium
(22). Thereafter, the NETs were
recovered as a supernatant at a concentration of 1.7×106
cell equivalents/ml by centrifugation at 400 × g and 10,000 × g.
Alternatively, the neutrophils were incubated without LPS for 4 h,
and the supernatants were recovered and used as the control.
Measurement of NET-associated
protein
The concentration of the NET-associated protein
(µg/ml) was quantified using the Bio-Rad protein assay
reagent (Bio-Rad Laboratories, Inc., Hercules, CA, USA) according
to the manufacturer's instructions using samples prepared with (+)
or without (−) LPS stimulation of neutrophils.
Measurement of NET-associated DNA
The concentration of NET-associated DNA was
quantified by adding 5 µg/ml Hoechst 33342 (FLICA™ caspase-1
assay) to samples (100 µl) in a 96-well plate (Nunc™
MicroWell™ 96-well microplates; Thermo Fisher Scientific). The
emission (E) at 460 nm (excitation at 355 nm) was measured using
samples prepared with (+) or without (−) LPS stimulation of
neutrophils, and RPMI-1640 medium as a background. The relative
fold change was calculated as follows: Relative fold change =
(Ewith LPS − ERPMI-1640)/(Ewithout
LPS − ERPMI-1640).
Measurement of NET-associated NE
The NET-associated NE was detected by western
blotting. Briefly, samples prepared with (+) or without (−) LPS
stimulation of neutrophils were subjected to sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
polyvinylidene fluoride membranes (Immobilon-P; Millipore Corp.,
Billerica, MA, USA). The membranes were blocked with BlockOne
(Nakara, Tokyo, Japan), probed with rat anti-mouse NE mAb (R&D
Systems, Inc.), followed by horseradish peroxidase-conjugated
anti-rat IgG (Santa Cruz Biotechnology, Inc.). Signals were
detected with SuperSignal West Dura chemiluminescent substrate
(Pierce Biotechnology, Inc., Rockford, IL, USA), and quantified as
grey-scale using LAS-3000 luminescent image analyzer (Fujifilm,
Tokyo, Japan) and MultiGauge software (Fujifilm). Relative fold
change was calculated as follows; Relative fold change =
grey-scalewith LPS/grey-scalewithout LPS.
Measurement of NET-associated
myeloperoxidase (MPO)-DNA complex
The NET-associated MPO-DNA complex was detected by
MPO-DNA ELISA. Briefly, a 96-well plate (Thermo Fisher Scientific)
was coated with 5 µg/ml anti-mouse MPO monoclonal antibody
(AF3667; R&D Systems, Inc.) overnight at 4°C. After washing 3
times, 20 µl of the samples prepared with (+) or without (−)
LPS stimulation of neutrophils and RPMI-1640 medium (as a
background) were added to the wells with 80 µl of incubation
buffer containing a peroxidase-labeled anti-DNA monoclonal antibody
(MCA-33; Cell Death ELISAPlus; Roche Diagnostics,
Indianapolis, IN, USA). The plate was incubated for 2 h with
shaking at 300 rpm at room temperature. After 3 washes, 100
µl peroxidase substrate (ABTS; Cell Death
ELISAPlus) was added, and absorbance (A) at 405 nm was
measured after a 20-min incubation at room temperature in the dark.
Relative fold change was calculated as follows: Relative fold
change = (Awith LPS −
ARPMI-1640)/(Awithout LPS −
ARPMI-1640).
Measurement of NET-associated histone-DNA
complex
The NET-associated histone-DNA complex was detected
using Cell Death ELISAPlus (Roche Diagnostics) according
to the manufacturer's instructions. Briefly, 20 µl of the
samples prepared with (+) or without (−) LPS stimulation of
neutrophils and RPMI-1640 medium (as a background) were added to a
streptavidin-coated plate with 80 µl of incubation buffer
containing a peroxidase-labeled anti-DNA mAb. The plate was
incubated for 2 h with shaking at 300 rpm at room temperature.
After 3 washes, 100 µl peroxidase substrate (ABTS) was
added, and absorbance at 405 nm was measured after a 20-min
incubation at room temperature in the dark. Relative fold change
was calculated as follows: Relative fold change = (Awith
LPS − ARPMI-1640)/(Awithout LPS −
ARPMI-1640).
Cell culture
A murine macrophage-like cell line J774 was
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and maintained in RPMI-1640 medium supplemented
with 10% fetal bovine serum (FBS; Sanko Junyaku, Tokyo, Japan) and
100 U/ml penicillin/100 µg/ml streptomycin (Nacalai Tesque,
Kyoto, Japan) at 37°C in 5% CO2. J774 cells
(2×104 cells/well) were seeded in 96-well culture plates
(Iwaki Brand) overnight, and then treated with NETs
(1.7×105 cell equivalents) for 24 h in the absence or
presence of 10 ng/ml LPS at 37°C in 5% CO2. Thereafter,
the supernatants were recovered for the IL-1β and lactate
dehydrogenase (LDH) assays.
Assays for caspase-1 and caspase-8
activation
J774 cells (2×104 cells/well) were seeded
in CC2 Chamber Slide system 8-wells (Thermo Fisher Scientific)
overnight, and then treated with NETs (1.7×105 cell
equivalents) for 24 h in the absence or presence of 10 ng/ml LPS at
37°C in 5% CO2. Thereafter, the activation of caspase-1
and caspase-8 was assayed using the FLICA™ caspase-1 assay kit
(ImmunoChemistry Technologies, LLC, Bloomington, MN, USA) and
FLICA™ caspase-8 assay kit (ImmunoChemistry Technologies, LLC),
respectively. Briefly, after treatment with LPS and NETs, the cells
were washed with fresh RPMI-1640 medium, and then the cells were
labeled with FAM-YVAD-FMK or FAM-LETD-FMK (fluorescent-labeled
caspase-1 or caspase-8 inhibitor that binds with activated
caspase-1 or caspase-8, respectively) by incubation for 1 h at
37°C. After washing, the cells were labeled with Hoechst 33342 (0.8
µg/ml) for 10 min. Finally, the slide was mounted with a
cover glass (Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen,
Germany) using aqueous medium Vectashield, and the cells were
photo-graphed using a fluorescence microscope system Axioplan
2.
Quantification of IL-1β and LDH
IL-1β 1evels in the supernatants were determined
using a commercially available mouse IL-1β ELISA kit (eBioscience,
Inc.), according to the manufacturer's instructions. LDH activity
in the supernatants was determined for the evaluation of cell
death; LDH activity in the supernatants and 1% Triton X-100-lysed
cells (as a total activity of 100%) was determined using a
commercially available LDH assay kit (Takara Bio, Inc.), according
to the manufacturer's instructions.
Statistical analysis
Data are shown as the means ± standard deviation. A
statistical test was performed using one-way analysis of variance
(ANOVA), followed by Bonferroni's multiple comparison test
(GraphPad Prism; GraphPad Software, Inc., San Diego, CA, USA).
Otherwise, a statistical test was performed using the Student's
t-test for experiments with two groups (GraphPad Prism). P<0.05
was considered to indicate a statistically significant
difference.
Results
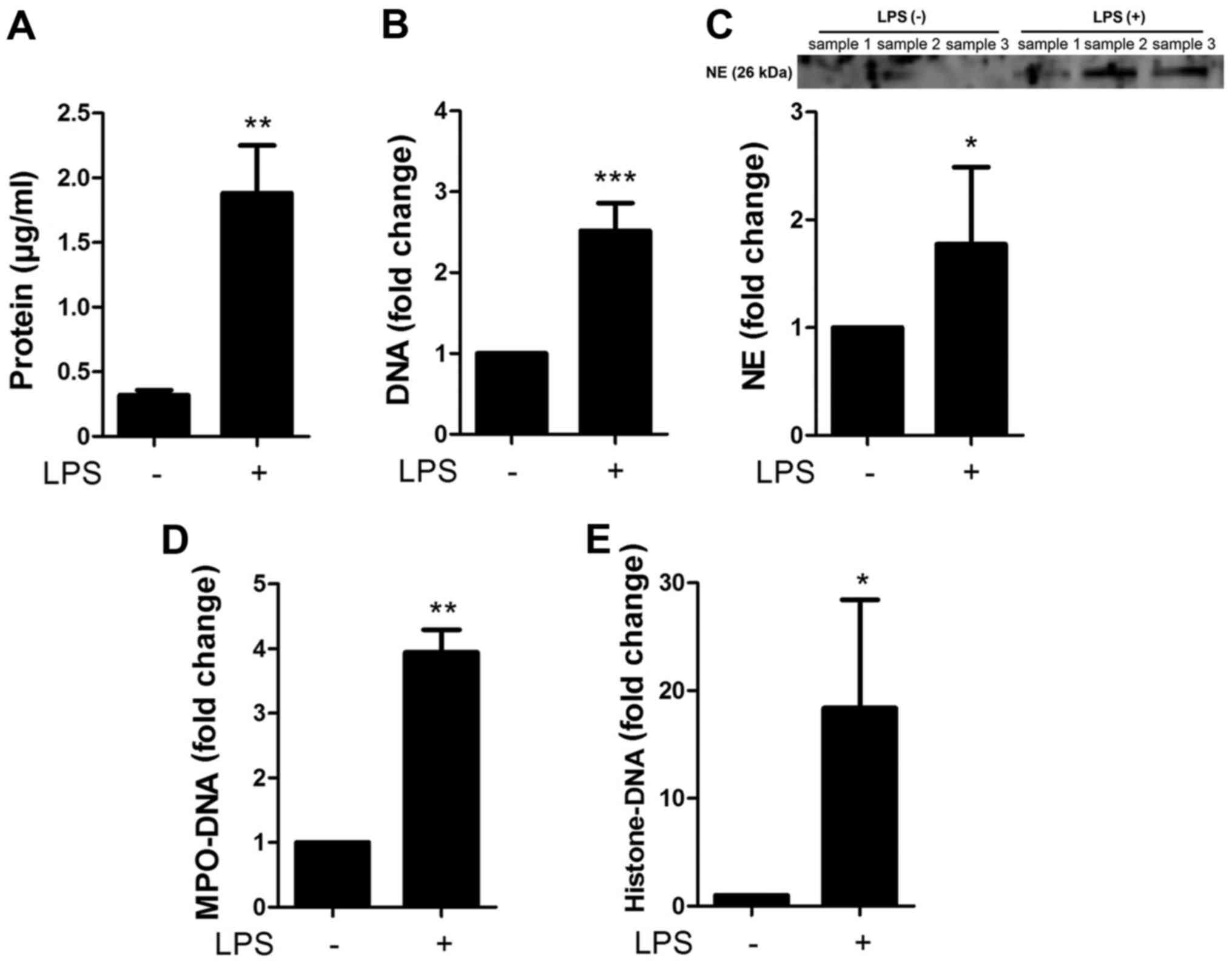
Characterization of the NETs
The NETs were characterized by measuring the
concentrations of proteins, DNA, NE, MPO-DNA complex and
histone-DNA complex contained in the preparations of NETs (with LPS
stimulation) and the controls (without LPS stimulation) (Fig. 2A). The preparation of NETs
contained 1.88±0.37 µg/ml protein, whereas the controls
contained 0.32±0.04 µg/ml protein. Moreover, the NETs
contained 2.52±0.34-fold higher DNA, 1.77±0.72-fold higher NE,
3.94±0.35-fold higher MPO-DNA complex and 18.4±10.0-fold higher
histone-DNA complex than the controls (Fig. 2B–E), confirming that NETs contain
the complexes of DNA, histone and granule proteins.
Treatment with NETS and LPS induces IL-1β
production by macrophages via the caspase-1 and caspase-8
pathways
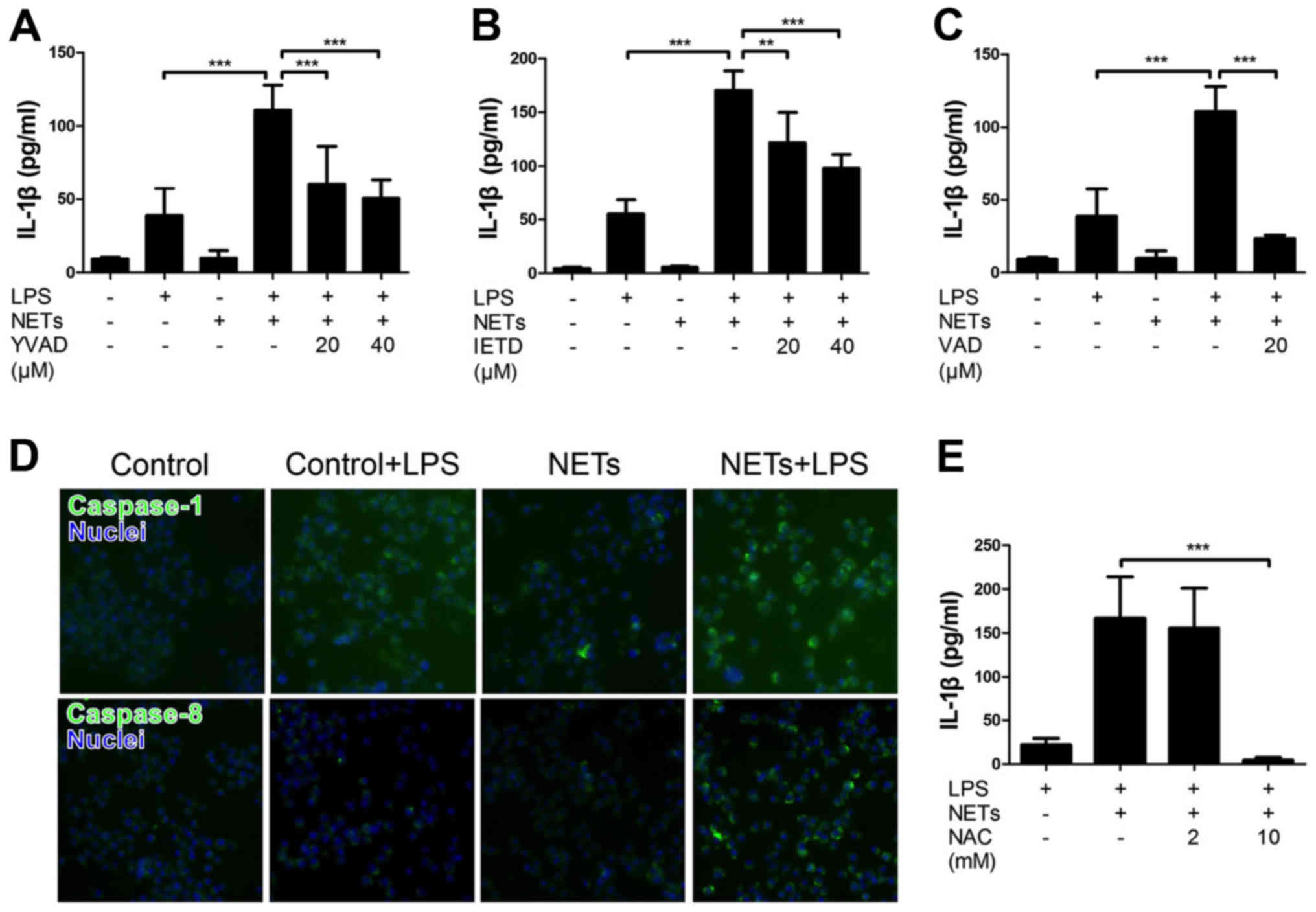
We first examined the effect of NETs and LPS
treatment on the cytokine production by macrophages by measuring
the release of IL-1β and LDH. Mouse macrophage-like J774 cells were
treated with NETs and LPS for 24 h. The results indicated that LPS
or NET treatment alone did not essentially induce the production of
IL-1β (Fig. 3A) and LDH (cell
death, data not shown). Notably, the treatment of J774 cells with
both NETs and LPS significantly induced the release of IL-1β
(Fig. 3A), but not LDH (data not
shown). Notably, Ac-YVAD-CHO (a caspase-1-specific inhibitor),
Ac-IETD-CHO (a caspase-8-specific inhibitor) and Z-VAD-FMK (a pan
caspase inhibitor) inhibited the NET/LPS-induced production of
IL-1β (Fig. 3A–C), suggesting
that the NET/LPS-induced IL-1β production was dependent on the
activation of caspase-1 and caspase-8. In fact, we confirmed the
activation of caspase-1 and caspase-8 by staining the activated
caspase with FAM-YVAD-FMK (a fluorescence-labeled caspase-1
inhibitor) and FAM-LETD-FMK (a fluorescence-labeled caspase-8
inhibitor), respectively. As shown in Fig. 3D, LPS or NET treatment alone did
not significantly induce either caspase-1 or the caspase-8
activation; however, the NET/LPS treatment considerably increased
the activation of both caspase-1 and caspase-8. These observations
indicate that treatment with NETs and LPS induced the production of
IL-1β by macrophages via caspase-1- and caspase-8-dependent
pathways. Furthermore, NAC (a ROS scavenger) inhibited the
NET/LPS-induced IL-1β production by J774 cells (Fig. 3E), suggesting the involvement of
ROS in the NET/LPS-induced IL-1β production by macrophages.
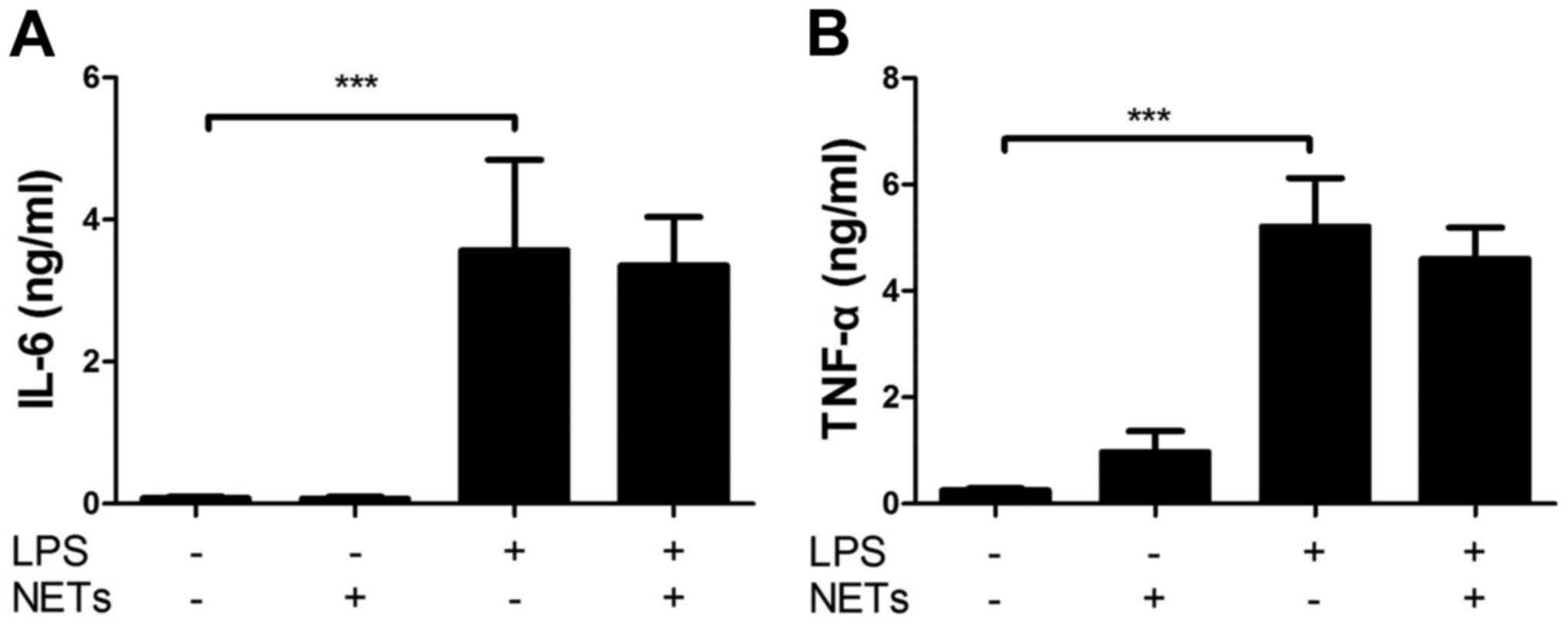
Separately, we confirmed that LPS alone significantly increased the
levels of IL-6 and tumor necrosis factor (TNF)-α compared with the
NETs alone (Fig. 4A and B).
Importantly, NET/LPS treatment did not further increase the levels
of IL-6 and TNF-α production compared with LPS alone. Moreover,
NETs alone did not essentially induce the production of IL-6 and
TNF-α compared with LPS, suggesting that NETs, used in this study,
if any, contained little amount of LPS for activating J774
cells.
NET-associated DNA is an important
component of the NET/LPS-induced IL-1β production
Furthermore, we investigated the involvement of
NET-associated DNA in the NET/LPS-induced IL-1β production by using
nucleases. Thus, J774 cells were treated with NETs/LPS in the
presence of nucleases (DNase I or MNase) for 24 h. Preliminarily,
we confirmed that DNase I and MNase totally degraded DNA (as
evidence by agarose gel electrophoresis) and did not show
cytotoxicity (as evidenced by LDH assay) (data not shown). Notably,
both DNase I and MNase significantly inhibited the NET/LPS-induced
IL-1β production by J774 cells (Fig.
5A and B). Next, we examined the effect of DNA on IL-1β
production by J774 cells using CT-DNA to mimic the NET-associated
DNA. CT-DNA alone did not induce the production of IL-1β; however,
CT-DNA and LPS significantly induced IL-1β production (Fig. 5C). These observations indicate
that NET-associated DNA is an important component of
NET/LPS-induced IL-1β production by macrophages.
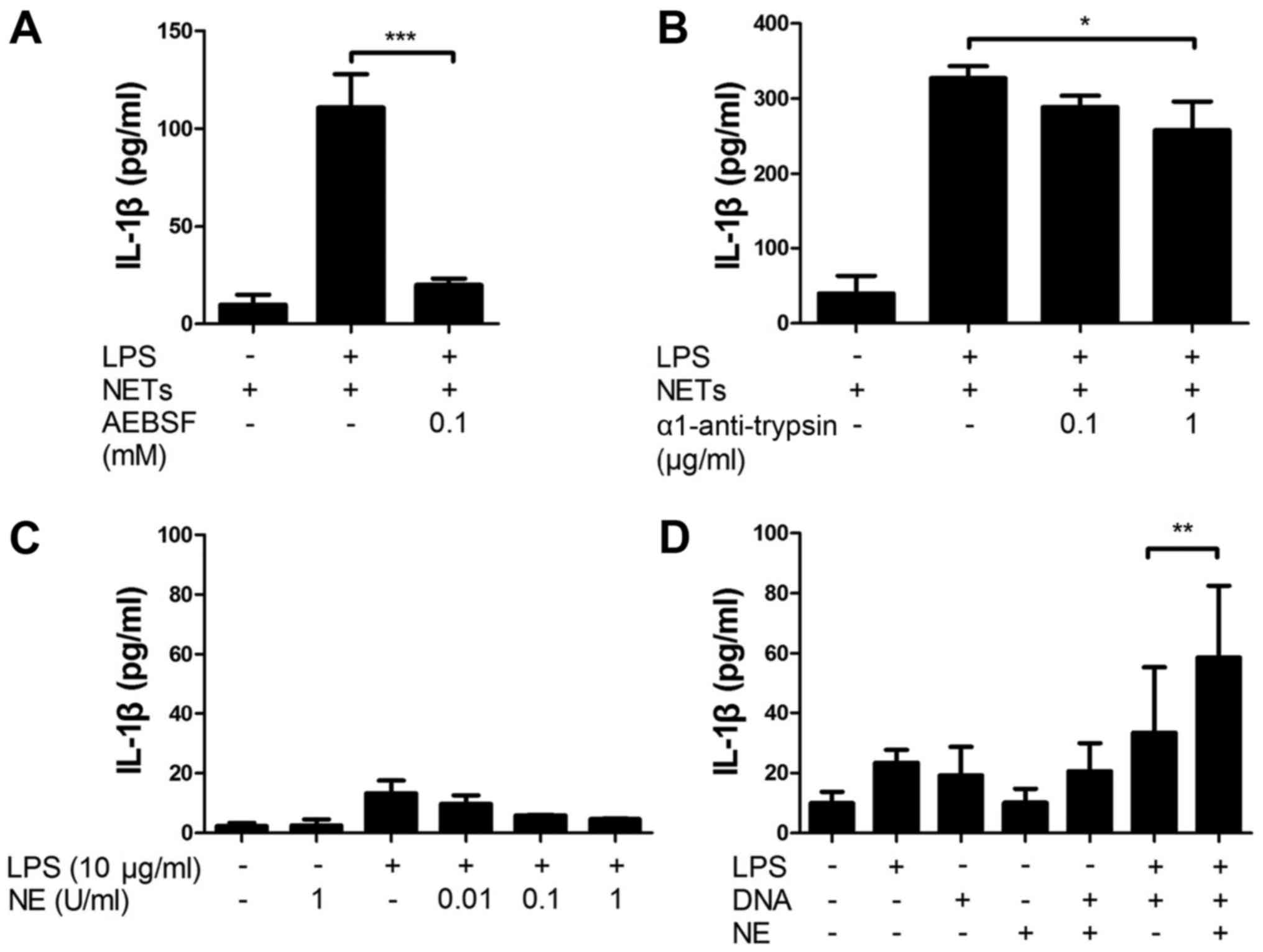
NET-associated serine proteases are
important components of NET/LPS-induced IL-1β production
Next, we investigated the involvement of
NET-associated serine proteases in the NET/LPS-induced IL-1β
production using serine protease inhibitors. Thus, the J774 cells
were treated with NETs/LPS in the presence of serine protease
inhibitors (AEBSF, a permeable serine inhibitor or α1-anti-trypsin,
an impermeable serine protease inhibitor) for 24 h. Importantly,
both AEBSF and α1-anti-trypsin significantly inhibited the
NET/LPS-induced IL-1β production by J774 cells (Fig. 6A and B). These results indicate
that NET-associated serine proteases are important components of
the NET/LPS-induced IL-1β production by macrophages. To confirm the
involvement of serine proteases in IL-1β production, the J774 cells
were incubated with NE. Unexpectedly, neither NE alone nor the
NE/LPS treatment essentially induced the production of IL-1β
(Fig. 6C). Notably, however, the
NE/DNA treatment significantly induced the IL-1β production by the
J774 cells in the presence of LPS (Fig. 6D), suggesting that NE induces
IL-1β production from macrophages in combination with DNA.
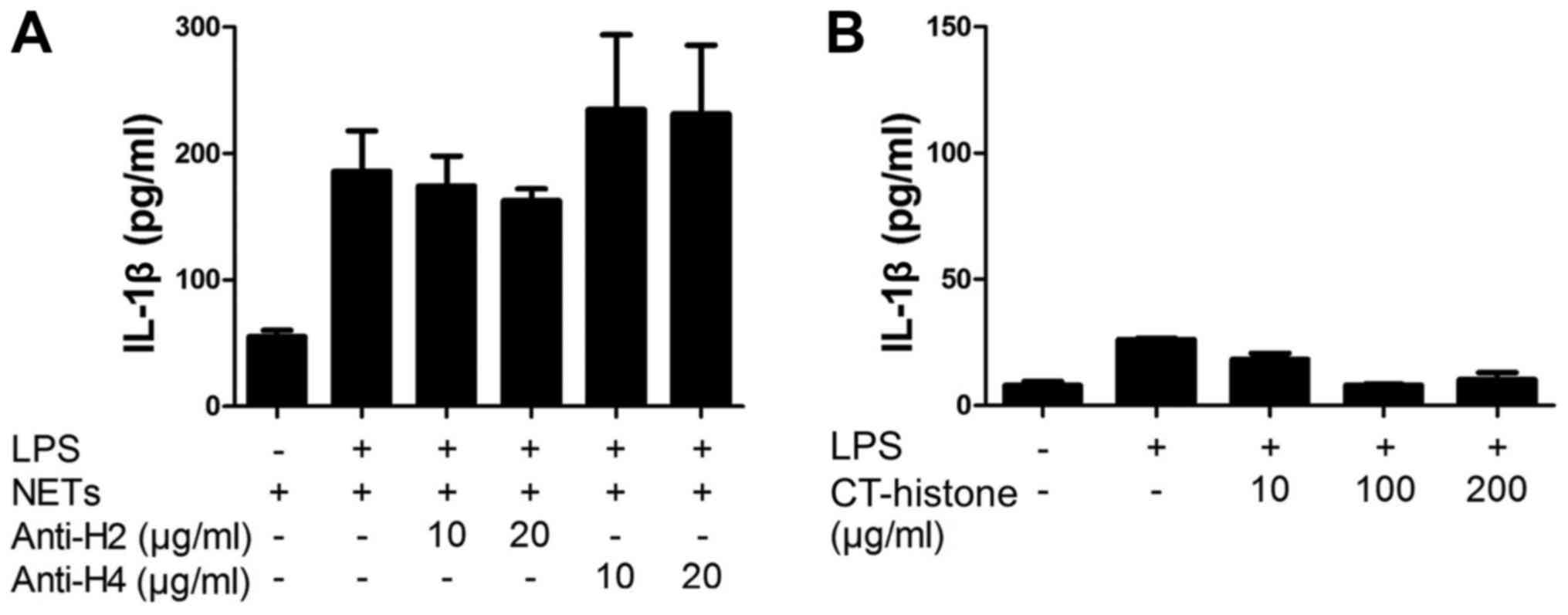
NET-associated histones are unlikely
involved in NET/LPS-induced IL-1β production
Finally, we investigated the involvement of
NET-associated histones in the NET/LPS-induced IL-1β production by
macrophages using histone antibodies. Thus, the J774 cells were
treated with NETs/LPS in the presence of histone antibodies
(anti-histone H2 mAb or anti-histone H4 pAb) for 24 h. Notably,
neither anti-histone H2 mAb nor anti-histone H4 pAb inhibited the
NET/LPS-induced IL-1β production by J774 cells (Fig. 7A). Further, we examined the direct
effect of histones on the IL-1β production by J774 cells using
CT-histones. As a result, CT-histones did not induce IL-1β
production by the J774 cells in the presence of LPS (Fig. 7B). These observations indicate
that NET-associated histones were unlikely involved in the
NET/LPS-induced IL-1β production by macrophages in our
experiments.
Discussion
Neutrophils play an important role as effectors of
inflammation, tissue injury and host defense against microbial
infection (9,10). Besides the well-known
antimicrobial potential, neutrophils trap and kill invading
microorganisms by extracellularly releasing NETs via NETosis, a
recently identified type of programmed cell death (11–13). Importantly, the components of NETs
(such as DNA and NE) are demonstrated to be potential mediators for
cytokine production in sepsis (11). For example, degradation of DNA (by
administration of recombinant DNase I) was found to inhibit
over-produced cytokines in septic mice (23–25); and knockout of NE significantly
reduced the production of IL-1β in response to Pseudomonas
aeruginosa infection in mice (26). In the present study, we revealed
that NETs, as a complex of DAMPs (containing DNA, histone and
serine proteases) induced the production of IL-1β by
macrophage-like J774 cells in the presence of LPS via the action of
caspase-1 and caspase-8, and that the NET-associated DNA and serine
proteases were involved in the production of IL-1β by the
cells.
IL-1β is a prototypical inflammatory cytokine, which
stimulates both local and systemic inflammatory responses (3), and acts synergistically with other
cytokines to cause tissue injury in sepsis (27). The production of IL-1β is mediated
mainly by the activation of caspase-1 (27–29), and requires two distinct stimuli,
microbial pathogen-associated molecular patterns (PAMPs, e.g.,
lipoproteins and LPS) and endogenous DAMPs (e.g., DNA and ATP)
(28,29). Stimulation by PAMPs initiates a
signaling cascade that leads to cellular activation (including the
upregulation of inflammatory cytokine genes) (30). In contrast, stimulation by DAMPs
activates caspase-1, which is involved in the processing and
release of IL-1β (30).
Additionally, recent studies have revealed that caspase-8 acts as
either a direct enzyme for the processing and release of IL-1β or
as an initiator for the activation of caspase-1, in response to
PAMPs and DAMPs (e.g., LPS and ATP) (31–34). In the present study, LPS and NETs
were regarded as PAMPs and DAMPs, respectively. Importantly, LPS or
NET treatment alone did not essentially elicit IL-1β production
from J774 cells, and treatment with both LPS and NETs significantly
induced IL-1β production (Fig.
3A). Importantly, the NET/LPS-induced IL-1β production was
inhibited by not only Ac-YVAD-CHO (a caspase-1-specific inhibitor)
but also Ac-IEAD-CHO (a caspase-8-specific inhibitor) (Fig. 3A and B). Moreover, we confirmed
that the NET/LPS treatment activated both caspase-1 and caspase-8
(Fig. 3D). These observations
suggest that the NET/LPS treatment induced the production of IL-1β
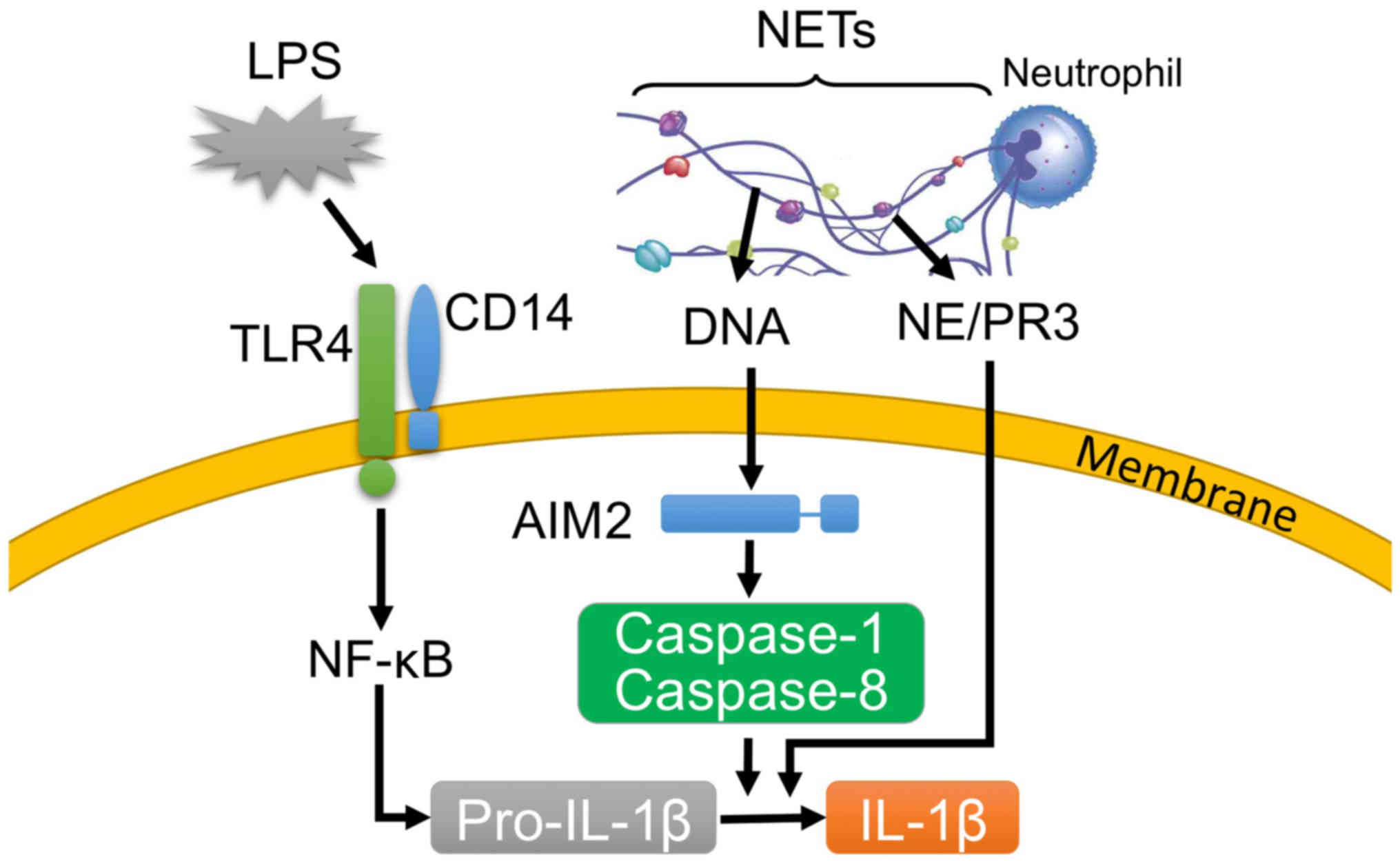
via the action of caspase-1 and caspase-8 (Fig. 8). Furthermore, it has been
recently reported that ROS can be common upstream activators of the
caspase-1 and caspase-8 pathways (35,36). Thus, we investigated the effect of
NAC (an ROS scavenger) on the NET/LPS-induced IL-1β production.
Notably, NAC inhibited the NET/LPS-induced IL-1β production by J774
cells (Fig. 3E), supporting the
involvement of ROS in the NET/LPS-induced IL-1β production by
macrophages. Furthermore, it has been reported that LPS alone can
efficiently induce the production of other cytokines (e.g., IL-6
and TNF-α), and the additional treatment of DAMPs (e.g., ATP)
cannot augment the LPS-induced production of these cytokines
(37,38). In the present study, we confirmed
that LPS alone significantly increased the levels of IL-6 and TNF-α
compared with the NETs alone (Fig.
4), and the NET/LPS treatment did not further increase the
levels of IL-6 and TNF-α production compared with LPS alone,
suggesting that NETs may not be important for the production of
these cytokines in sepsis compared with PAMPs and other DAMPs.
Genomic DNA is the main component of NETs (14). In this study, nucleases (DNase I
and MNase) inhibited the NET/LPS-induced production of IL-1β
(Fig. 5A and B), suggesting that
DNA is an important component of the NET/LPS-induced production of
IL-1β. We also confirmed that extracellular DNA (CT-DNA) induced
the production of IL-1β in combination with LPS (Fig. 5C). Previous studies demonstrated
that cytosol absent in melanoma 2 (AIM2) is essential for the
recognition of cytosol DNA to induce the activation of
caspase-1/caspase-8 and production of IL-1β (39,40). In our system, we did not transfect
DNA into the cytosol; however, extracellular DNA could be
phagocytosed by macrophages in the presence of LPS, since LPS can
potently augment the phagocytic ability of macrophages (41). Importantly, NETs are phagocytosed
by macrophages either in the presence or absence of invading
microorganisms (20,24). Thus, it is reasonable to speculate
that the NETs were phagocytosed by macrophages, and then the
NET-associated DNA induced the production of IL-1β via the
recognition of DNA by AIM2 (Fig.
8).
Notably, genomic DNA is normally localized in the
nuclei, and is extracellularly released during cell death (e.g.,
NETosis and necrosis) (42).
Importantly, the serum levels of extracellular DNA in septic
patients are significantly higher than that in healthy individuals
(43). Notably, degradation of
DNA by the administration of recombinant DNase I was found to
inhibit the over-produced cytokines in septic mice (23–25). Importantly, NET-associated DNA
(characterized by DNA-histone, DNA-MPO or DNA-NE complex) is
detected in blood, and its level is correlated with that of
extracellular DNA in blood in sepsis (44,45). These observations suggest that the
NET-associated DNA may contribute to the cytokine production in
sepsis.
Neutrophil serine proteases (including proteinase 3,
NE and cathepsin G) are important regulators of inflammation
(14,46). Importantly, NET-associated serine
proteases mediate the production of cytokines and chemokines [e.g.,
IL-1β and monocyte chemotactic protein-1 (MCP-1)] by inflammatory
cells (47). In the present
study, both AEBSF (a cell permeable serine protease inhibitor) and
α1-anti-trypsin (a cell impermeable serine protease inhibitor)
significantly inhibited the NET/LPS-induced production of IL-1β
(Fig. 6A and B), suggesting that
NET-associated serine proteases are important for the production of
IL-1β. Notably, the inhibitory effect of α1-anti-trypsin was
significantly lower than that of AEBSF (Fig. 6A and B), suggesting a possibility
that intracellular NET-associated serine proteases are more
important for the production of IL-1β. Notably, the possibility may
be supported by the findings that NET-associated serine proteases
are phagocytosed by macrophages accompanied with NETs (20,24), as discussed above, and that
neutrophil serine proteases (e.g., proteinase 3 and NE) can process
IL-1β (48). Furthermore, we
aimed to evaluate the effect of serine proteases on the production
of IL-1β using NE. Unexpectedly, NE (0.01 U and 0.1 U) did not
essentially induce the production of IL-1β by J774 cells in the
absence or presence of LPS (Fig.
6C). Thus, based on the high affinity of NE for DNA (516–712
nM) (49), we evaluated the
combined effect of NE (0.1 U/ml) and CT-DNA (1 µg/ml) on
IL-1β production. The results revealed that NE and CT-DNA induced
the production of IL-1β in the presence of LPS (Fig. 6D), suggesting that neutrophil
serine proteases such as NE can be an important component of NETs
for the production of IL-1β by macrophages in combination with DNA
(Fig. 8).
NETs are described as a complex of antimicrobial
components with the ability to trap and kill bacteria (11); however, NETs exacerbate the
pathophysiology of sepsis via the actions of inducing cell death
and thrombus formation (15,19). Furthermore, we revealed that NETs,
as a complex of DAMPs, induce the production of IL-1β by
macrophages in combination with LPS via the action of caspase-1 and
caspase-8, and that DNA and serine proteases are important
components of NETs for IL-1β production. Thus, our findings suggest
a possibility that NETs play a role in cytokine production and have
an adverse impact on sepsis, and provide a novel insight that the
degradation and/or inhibition of NET-associated effective
components may be a therapeutic target of sepsis.
In conclusion, the present results demonstrated that
NETs significantly induced the IL-1β production by mouse
macrophage-like J774 cells in combination with LPS. Importantly,
the NET/LPS-induced IL-1β production was inhibited by both
caspase-1 and caspase-8 inhibitors. Moreover, nucleases and serine
protease inhibitors significantly inhibited the NET/LPS-induced
IL-1β production. These observations suggest that NETs induce the
production of IL-1β from macrophages in combination with LPS via
the caspase-1 and caspase-8 pathways, and NET-associated DNA and
serine proteases are likely involved in the NET/LPS-induced IL-1β
production as essential components (Fig. 8).
Acknowledgments
We are grateful to Dr Kaori Suzuki (Department of
Host Defense and Biochemical Research, Juntendo University,
Graduate School of Medicine) for the helpful discussion. This study
was supported by a Grant-in-Aid (grant no. 26460538) for Scientific
Research from the Japan Society for the Promotion of Science, a
Grant-in-Aid (grant no. S0991013) and a Grant-in-Aid (grant no.
S1311011) from the Ministry of Education, Culture, Sport, Science,
and Technology, Japan (MEXT) for the Foundation of Strategic
Research Projects in Private Universities.
References
|
1
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The Third International Consensus
Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA.
315:801–810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
O'Brien JM Jr, Ali NA, Aberegg SK and
Abraham E: Sepsis. Am J Med. 120:1012–1022. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gabay C, Lamacchia C and Palmer G: IL-1
pathways in inflammation and human diseases. Nat Rev Rheumatol.
6:232–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cohen J: The immunopathogenesis of sepsis.
Nature. 420:885–891. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pinheiro da Silva F and Nizet V: Cell
death during sepsis: Integration of disintegration in the
inflammatory response to overwhelming infection. Apoptosis.
14:509–521. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hotchkiss RS and Nicholson DW: Apoptosis
and caspases regulate death and inflammation in sepsis. Nat Rev
Immunol. 6:813–822. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sarkar A, Hall MW, Exline M, Hart J, Knatz
N, Gatson NT and Wewers MD: Caspase-1 regulates Escherichia coli
sepsis and splenic B cell apoptosis independently of interleukin-1β
and interleukin-18. Am J Respir Crit Care Med. 174:1003–1010. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu Z, Murakami T, Suzuki K, Tamura H,
Reich J, Kuwahara-Arai K, Iba T and Nagaoka I: Antimicrobial
cathelicidin peptide LL-37 inhibits the pyroptosis of macrophages
and improves the survival of polybacterial septic mice. Int
Immunol. 28:245–253. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mantovani A, Cassatella MA, Costantini C
and Jaillon S: Neutrophils in the activation and regulation of
innate and adaptive immunity. Nat Rev Immunol. 11:519–531. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nathan C: Neutrophils and immunity:
Challenges and opportunities. Nat Rev Immunol. 6:173–182. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Camicia G, Pozner R and de Larrañaga G:
Neutrophil extracellular traps in sepsis. Shock. 42:286–294. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Remijsen Q, Kuijpers TW, Wirawan E,
Lippens S, Vandenabeele P and Vanden Berghe T: Dying for a cause:
NETosis, mechanisms behind an antimicrobial cell death modality.
Cell Death Differ. 18:581–588. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McDonald B, Urrutia R, Yipp BG, Jenne CN
and Kubes P: Intravascular neutrophil extracellular traps capture
bacteria from the bloodstream during sepsis. Cell Host Microbe.
12:324–333. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Urban CF, Ermert D, Schmid M, Abu-Abed U,
Goosmann C, Nacken W, Brinkmann V, Jungblut PR and Zychlinsky A:
Neutrophil extracellular traps contain calprotectin, a cytosolic
protein complex involved in host defense against Candida albicans.
PLoS Pathog. 5:e10006392009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saffarzadeh M, Juenemann C, Queisser MA,
Lochnit G, Barreto G, Galuska SP, Lohmeyer J and Preissner KT:
Neutrophil extracellular traps directly induce epithelial and
endothelial cell death: A predominant role of histones. PLoS One.
7:e323662012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Alfaidi M, Wilson H, Daigneault M, Burnett
A, Ridger V, Chamberlain J and Francis S: Neutrophil elastase
promotes interleukin-1β secretion from human coronary endothelium.
J Biol Chem. 290:24067–24078. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu J, Zhang X, Monestier M, Esmon L and
Esmon CT: Extracellular histones are mediators of death through
TLR2 and TLR4 in mouse fatal liver injury. J Immunol.
187:2626–2631. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu J, Zhang X, Pelayo R, Monestier M,
Ammollo CT, Semeraro F, Taylor FB, Esmon L, Lupu F and Esmon CT:
Extracellular histones are major mediators of death in sepsis. Nat
Med. 15:1318–1321. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gould TJ, Vu TT, Swystun LL, Dwivedi DJ,
Mai SH, Weitz JI and Liaw PC: Neutrophil extracellular traps
promote thrombin generation through platelet-dependent and
platelet-independent mechanisms. Arterioscler Thromb Vasc Biol.
34:1977–1984. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Braian C, Hogea V and Stendahl O:
Mycobacterium tuberculosis-induced neutrophil extracellular traps
activate human macrophages. J Innate Immun. 5:591–602. 2013.
View Article : Google Scholar
|
|
21
|
Cools-Lartigue J, Spicer J, McDonald B,
Gowing S, Chow S, Giannias B, Bourdeau F, Kubes P and Ferri L:
Neutrophil extracellular traps sequester circulating tumor cells
and promote metastasis. J Clin Invest. 123:3446–3458. 2013.
View Article : Google Scholar :
|
|
22
|
Farrera C and Fadeel B: Macrophage
clearance of neutrophil extracellular traps is a silent process. J
Immunol. 191:2647–2656. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo L, Zhang S, Wang Y, Rahman M, Syk I,
Zhang E and Thorlacius H: Proinflammatory role of neutrophil
extracellular traps in abdominal sepsis. Am J Physiol Lung Cell Mol
Physiol. 307:L586–L596. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meng W, Paunel-Görgülü A, Flohé S,
Hoffmann A, Witte I, MacKenzie C, Baldus SE, Windolf J and Lögters
TT: Depletion of neutrophil extracellular traps in vivo results in
hypersusceptibility to polymicrobial sepsis in mice. Crit Care.
16:R1372012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mai SH, Khan M, Dwivedi DJ, Ross CA, Zhou
J, Gould TJ, Gross PL, Weitz JI, Fox-Robichaud AE and Liaw PC;
Canadian Critical Care Translational Biology Group: Delayed but not
early treatment with DNase reduces organ damage and improves
outcome in a murine model of sepsis. Shock. 44:166–172. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karmakar M, Sun Y, Hise AG, Rietsch A and
Pearlman E: Cutting edge: IL-1β processing during Pseudomonas
aeruginosa infection is mediated by neutrophil serine proteases and
is independent of NLRC4 and caspase-1. J Immunol. 189:4231–4235.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Casey LC, Balk RA and Bone RC: Plasma
cytokine and endotoxin levels correlate with survival in patients
with the sepsis syndrome. Ann Intern Med. 119:771–778. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miao EA, Rajan JV and Aderem A:
Caspase-1-induced pyroptotic cell death. Immunol Rev. 243:206–214.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bergsbaken T, Fink SL and Cookson BT:
Pyroptosis: Host cell death and inflammation. Nat Rev Microbiol.
7:99–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bryant C and Fitzgerald KA: Molecular
mechanisms involved in inflammasome activation. Trends Cell Biol.
19:455–464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Antonopoulos C, Russo HM, El Sanadi C,
Martin BN, Li X, Kaiser WJ, Mocarski ES and Dubyak GR: Caspase-8 as
an effector and regulator of NLRP3 inflammasome signaling. J Biol
Chem. 290:20167–20184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maelfait J, Vercammen E, Janssens S,
Schotte P, Haegman M, Magez S and Beyaert R: Stimulation of
Toll-like receptor 3 and 4 induces interleukin-1β maturation by
caspase-8. J Exp Med. 205:1967–1973. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vince JE, Wong WW, Gentle I, Lawlor KE,
Allam R, O'Reilly L, Mason K, Gross O, Ma S, Guarda G, et al:
Inhibitor of apoptosis proteins limit RIP3 kinase-dependent
interleukin-1 activation. Immunity. 36:215–227. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shenderov K, Riteau N, Yip R, Mayer-Barber
KD, Oland S, Hieny S, Fitzgerald P, Oberst A, Dillon CP, Green DR,
et al: Cutting edge: Endoplasmic reticulum stress licenses
macrophages to produce mature IL-1β in response to TLR4 stimulation
through a caspase-8- and TRIF-dependent pathway. J Immunol.
192:2029–2033. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Harijith A, Ebenezer DL and Natarajan V:
Reactive oxygen species at the crossroads of inflammasome and
inflammation. Front Physiol. 5:3522014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
El Kebir D and Filep JG: Targeting
neutrophil apoptosis for enhancing the resolution of inflammation.
Cells. 2:330–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu Z, Murakami T, Suzuki K, Tamura H,
Kuwahara-Arai K, Iba T and Nagaoka I: Antimicrobial cathelicidin
peptide LL-37 inhibits the LPS/ATP-induced pyroptosis of
macrophages by dual mechanism. PLoS One. 9:e857652014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mao K, Chen S, Chen M, Ma Y, Wang Y, Huang
B, He Z, Zeng Y, Hu Y, Sun S, et al: Nitric oxide suppresses NLRP3
inflammasome activation and protects against LPS-induced septic
shock. Cell Res. 23:201–212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Muruve DA, Pétrilli V, Zaiss AK, White LR,
Clark SA, Ross PJ, Parks RJ and Tschopp J: The inflammasome
recognizes cytosolic microbial and host DNA and triggers an innate
immune response. Nature. 452:103–107. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sagulenko V, Thygesen SJ, Sester DP, Idris
A, Cridland JA, Vajjhala PR, Roberts TL, Schroder K, Vince JE, Hill
JM, et al: AIM2 and NLRP3 inflammasomes activate both apoptotic and
pyroptotic death pathways via ASC. Cell Death Differ. 20:1149–1160.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Anand RJ, Kohler JW, Cavallo JA, Li J,
Dubowski T and Hackam DJ: Toll-like receptor 4 plays a role in
macrophage phagocytosis during peritoneal sepsis. J Pediatr Surg.
42:927–932; discussion 933. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Beyer C and Pisetsky DS: Modeling nuclear
molecule release during in vitro cell death. Autoimmunity.
46:298–301. 2013. View Article : Google Scholar
|
|
43
|
Meng W, Paunel-Görgülü A, Flohé S, Witte
I, Schädel-Höpfner M, Windolf J and Lögters TT: Deoxyribonuclease
is a potential counter regulator of aberrant neutrophil
extracellular traps formation after major trauma. Mediators
Inflamm. 2012:1495602012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Czaikoski PG, Mota JM, Nascimento DC,
Sônego F, Castanheira FV, Melo PH, Scortegagna GT, Silva RL,
Barroso-Sousa R, Souto FO, et al: Neutrophil extracellular traps
induce organ damage during experimental and clinical sepsis. PLoS
One. 11:e01481422016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tanaka K, Koike Y, Shimura T, Okigami M,
Ide S, Toiyama Y, Okugawa Y, Inoue Y, Araki T, Uchida K, et al: In
vivo characterization of neutrophil extracellular traps in various
organs of a murine sepsis model. PLoS One. 9:e11–1888. 2014.
View Article : Google Scholar
|
|
46
|
Pham CT: Neutrophil serine proteases:
Specific regulators of inflammation. Nat Rev Immunol. 6:541–550.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schauer C, Janko C, Munoz LE, Zhao Y,
Kienhöfer D, Frey B, Lell M, Manger B, Rech J, Naschberger E, et
al: Aggregated neutrophil extracellular traps limit inflammation by
degrading cytokines and chemokines. Nat Med. 20:511–517. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Netea MG, Simon A, van de Veerdonk F,
Kullberg BJ, Van der Meer JW and Joosten LA: IL-1β processing in
host defense: Beyond the inflammasomes. PLoS Pathog.
6:e10006612010. View Article : Google Scholar
|
|
49
|
Thomas MP, Whangbo J, McCrossan G, Deutsch
AJ, Martinod K, Walch M and Lieberman J: Leukocyte protease binding
to nucleic acids promotes nuclear localization and cleavage of
nucleic acid binding proteins. J Immunol. 192:5390–5397. 2014.
View Article : Google Scholar : PubMed/NCBI
|