Introduction
Melanin is a natural pigment synthesized and stored
in specialized organelles, termed melanosomes, in melanocytes
(1). In the skin and hair, the
melanosomes are transported to dendrites of melanocytes and then
transferred to the neighboring keratinocytes. Insufficient
synthesis of melanin leads to several human hypopigmentation
disorders, such as hair graying and vitiligo (2). A recent study showed that 74% of
people between 45 and 65 years of age have grey hair (3). Vitiligo is a frequent cause of
depigmentation worldwide with an estimated prevalence of 1%
(4). Although there are several
treatments for hypopigmentation disorders, the efficacy and safety
remain unsatisfied and need to be improved (5). Therefore, it is urgent to develop
novel agents for these diseases.
Melanogenesis is the process of melanin synthesis in
melanocytes. Two types of melanin are present in mammals: a
black/brown eumelanin and a red/yellow pheomelanin. The ratio of
eumelanin and pheomelanin determines the diversity of skin and hair
pigmentation in humans. Synthesis of both types of melanins begins
with the amino acid L-tyrosine. Tyrosinase catalyzes L-tyrosine to
L-3,4-dihydroxyphenyla lanine (L-DOPA), the rate-limiting step, for
both eumelanin and pheomelanin. Eumelanin synthesis additionally
requires the enzymes tyrosinase-related protein 2 (Trp2) and Trp1,
whereas pheomelanin additionally requires the amino acid cysteine
(6). A well-known factor that can
induce tyrosinase expression is α-melanocyte stimulating hormone
(α-MSH), which binds the melanocortin 1 receptor (MC1R) to activate
adenylyl cyclase to produce cAMP. The cAMP activates cAMP-dependent
kinase A (PKA) and increases the expression of melanocyte-specific
microphthalmia-associated transcription factor (Mitf), a master
regulator for the expression of melanogenic enzymes tyrosinase,
Trp1 and Trp2 (6). Identifying
molecules that can modulate tyrosinase expression and activity is
recognized to be crucial for developing agents to treat
pigmentation disorders.
Lactoferrin (Lf) is a multifunctional glycoprotein
in mammal milk which is known to exhibit a wide range of biological
activities including antimicrobial, antiviral, antioxidant, and
immunomodulatory activities. Lactoferricin B (LfcinB) is a
25-residue peptide (aa17-41) derived from pepsin-digested bovine
lactoferrin (LfB).
LfcinB-related peptides share some activities with
full-length Lf, including antimicrobial, antiviral, antitumor,
immunological, antihypertensive and anti-catabolic properties
(7–11). LfB has been reported to inhibit
pigmentation of melanoma cells (12). However, the effects of
LfcinB-related peptides on melanogenesis remain unknown.
In this study, we found that three LfcinB-related
peptides enhanced pigmentation in B16F10 melanoma cells. Among
them, the 18-residue peptide LfB17-34, FKCRRWQWRM KKLGAPSI,
strongly increased melanin synthesis, which is associated with
elevated expression of melanogenic enzymes tyrosinase and Trp1.
Moreover, LfB17-34 was able to decrease phosphorylation of Erk, a
well-known signaling pathway to regulate melanogenesis. These
findings suggest that LfB17-34 is a novel inducer of melanogenesis,
whose stimulatory effect is at least partly through decreased
phosphorylation of Erk and increased expression of tyrosinase and
Trp1.
Materials and methods
Chemicals
Isobutylmethylxanthine (IBMX), L-DOPA, melanin,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
solution (MTT), paraformaldehyde, and all other chemicals were
purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise
indicated.
Preparation of peptides
All peptides were synthesized by MDBio, Inc.
(Taipei, Taiwan) and the purity and composition of these peptides
was confirmed by high performance liquid chromatography (HPLC) and
mass spectrometry. Peptide stocks were prepared by dissolving 10 mg
of lyophilized peptide powder in 1 ml of double deionized water
(ddH2O), and then stored at -20°C.
Cell culture
B16F10 and B16F0 murine melanoma cells (American
Type Culture Collection, Manassas, VA, USA) were cultured in phenol
red-free Dulbecco's modified Eagle's medium (DMEM) (Thermo
Scientific, Barrington, IL, USA) with 10% fetal bovine serum (FBS)
(Gibco, Grand Island, NY, USA) and penicillin/streptomycin (100
IU/50 g/ml) in a humidified atmosphere containing 5% CO2
at 37°C. B16F10 cells (2×105) were transfected with 1
µg of empty vector or vector coding constitutively active
MEK (13) using Lipofectamine
2000 (Invitrogen Life Technologies, Carlsbad, CA, USA) for 48 h and
then used for the assays.
Cell viability assay
Cells (2×103) were seeded in 96-well
plates. After culture for different time periods, MTT was added to
a final concentration of 0.5 mg/ml and incubated for 4 h at 37°C to
allow MTT reduction. The formazan crystals were then dissolved in
10% sodium dodecyl sulfate (SDS) containing 0.01 N HCl and
absorbance was measured at the dual wavelengths of 570 and 630 nm
with a spectrophotometer (VersaMax; Molecular Devices, Silicon
Valley, CA, USA).
Melanin content assay
Melanin contents of cultured cells were measured
according to the method (14)
with a slight modification. Briefly, cells were seeded in a 6-well
plate (2×104 cells/well) and incubated overnight to
allow cells to adhere. After treating with various test samples for
4 or 5 days, cells were washed with phosphate-buffered saline
(PBS), trypsinized, counted, and then lysed in 1 N NaOH at 100°C
for 1 h. The melanin content in each sample was calculated by
comparison of absorbance at 400 nm of unknown samples with a
standard curve obtained using synthetic melanin. The results were
converted to means ± SE pg/cell from 3 independent experiments.
Tyrosinase activity assay
Tyrosinase enzyme activity of cultured cells were
measured according to the a previous study (15) with a slight modification. Briefly,
cells were seeded in a 6-well plate (2×104 cells/well)
and incubated for overnight. Cells were treated with peptides for 4
or 5 days. Cells were harvested and lysed with phosphate buffer (pH
6.8) containing 1% Triton X-100, followed by freezing and thawing.
The supernatants were collected by centrifugation at 10,000 × g for
10 min and the protein contents were determined using the BCA
protein assay kit (Thermo Scientific, Rockford, IL, USA). For
reactions, 100 µl of cell lysates with the same amount of
proteins were aliquoted into each wells of a 96-well plate, and
then 100 µl of 5 mM L-DOPA were added to each wells.
Following a 30-min incubation period at 37°C, the absorbance was
measured spectrophotometrically at 475 nm.
L-DOPA staining
L-DOPA staining was performed to measure tyrosinase
enzyme activity in cultured cells. Cells were seeded in a 6-well
plate (2×104 cells/well) and incubated overnight. After
treating with various test samples in an incubator for 4 or 5 days,
cells were washed with PBS and fixed with 2% paraformaldehyde for
20 min. After washing with PBS for three times, the cells were
incubated with 0.1% L-DOPA in phosphate buffer (pH 6.8) at 37°C for
5 h and then photographed under a light microscope (16).
Reverse transcription-quantitative
PCR(RT-qPCR)
Total cellular RNA was isolated by using PureLink™
RNA mini kit (Invitrogen Life Technologies). For cDNA synthesis, 5
µg of total RNA was used as templates in a 20 µl
reverse transcription reaction. The quantitative PCR system Mx3000P
(Stratagene, La Jolla, CA, USA) was used for real-time PCR
reactions. Reactions were performed in a 20-µl volume with 1
µl cDNA and 10 µl Brilliant®
SYBR®-Green qPCR master mix (Stratagene). Samples were
analyzed in triplicate, and product purity was checked by
dissociation curves at the end of real-time PCR cycles. Relative
quantity of gene expression normalized to β-actin was analyzed with
MxPro software (Stratagene) (17). The following primer pairs were
used: Mitf sense, 5′-AACAAGGGAACCATTCTCAAGG-3′ and anti-sense,
5′-AGATCAGGCGAGCAGAGACC-3′; Tyr sense, 5′-CTCTGGGCTTAGCAGTAGGC-3′
and antisense, 5′-GCAAGCTGTGGTAGTCGTCT-3′; Trp1 sense,
5′-GCTGCAGGAGCCTTCTTTCTC-3′ and antisense,
5′-AAGACGCTGCACTGCTGGTCT-3′; Trp2 sense,
5′-GTCCTCCACTCTTTTACAGACG-3′ and antisense,
5′-ATTCGGTTGTGACCAATGGGT-3′; β-actin sense,
5′-AATCGTGCGTGACATCAAAG-3′ and antisense,
5′-GAAAAGAGCCTCAGGGCAT-3′.
Western blot analysis
Protein lysates (40 µg) were resolved by 8%
SDS-PAGE, transferred to polyvinylidene difluoride (PVDF) membranes
and then exposed to the appropriate antibodies. The primary
antibodies used were p-Akt (#4060), Akt (#4691), p-Erk (#9101), Erk
(#9102), p38 (#9212), and p-p38 (#9216) from Cell Signaling
Technology (Danvers, MA, USA), β-actin from Sigma-Aldrich, and
tyrosinase (H-109) (#sc-15341), Trp1 (H-90) (#sc-25543), Trp2
(H-150) (#sc-25544), and Mitf (H-50) (#sc-25386) antibodies were
supplied by Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
The secondary antibodies used were horseradish peroxidase
(HRP)-conjugated anti-mouse (115-035-003) or anti-rabbit
(111-035-144) antibody (Jackson ImmunoResearch Laboratories, West
Grove, PA, USA). Protein signals were quantified by optical density
ratios using β-actin expression as a control.
Statistical analyses
Quantitative data from at least three independent
experiments are expressed as the means ± standard deviation (SD).
Student's t-tests were used to compare the differences between
groups. p<0.05 is considered statistically significant.
Results
LfcinB-related peptides increase
pigmentation in B16F10 cells
To investigate whether LfcinB-related peptides
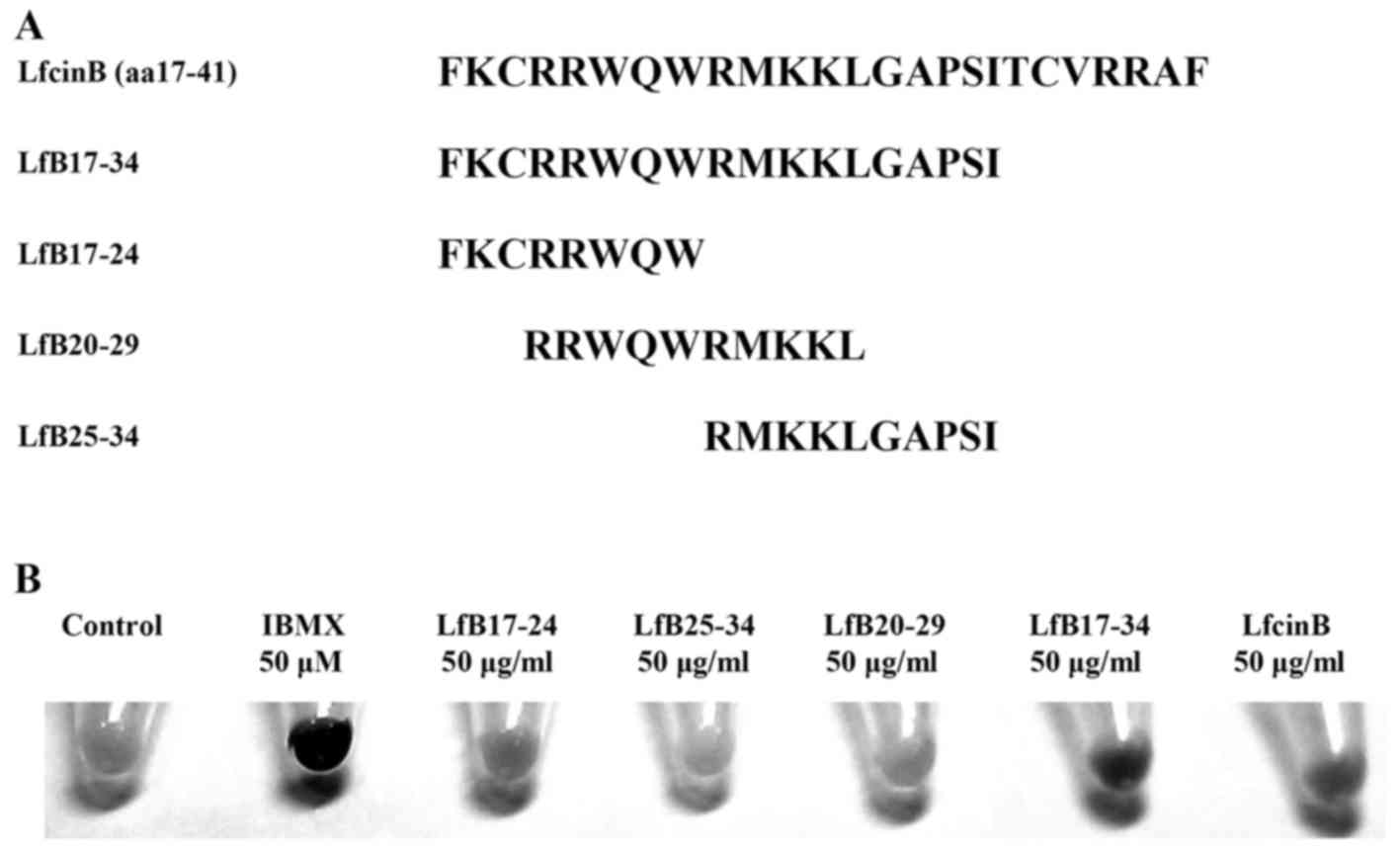
affect pigmentation, we synthesized 5 LfcinB-related peptides. The
amino acid sequences of these peptides are shown in Fig. 1A. The peptides and a chemical 50
µM IBMX were used to treat mouse B16F10 melanoma cells for 5
days and their effects on pigmentation in cells were observed.
IBMX, a non-specific inhibitor of cyclic AMP and cyclic GMP
phosphodiesterases (PDEs), is a positive control that is commonly
used compound to induce melanin synthesis. We found that three
LfcinB-related peptides including LfB17-24, LfB17-34 and LfcinB,
but not LfB25-34 and LfB20-29, were able to enhance pigmentation in
B16F10 cells (Fig. 1B). Notably,
LfB17-34 exhibited the most prominent effect on the enhancement of
pigmentation.
LfB17-34 enhances melanin synthesis and
secretion in a dose-dependent manner without significant
toxicity
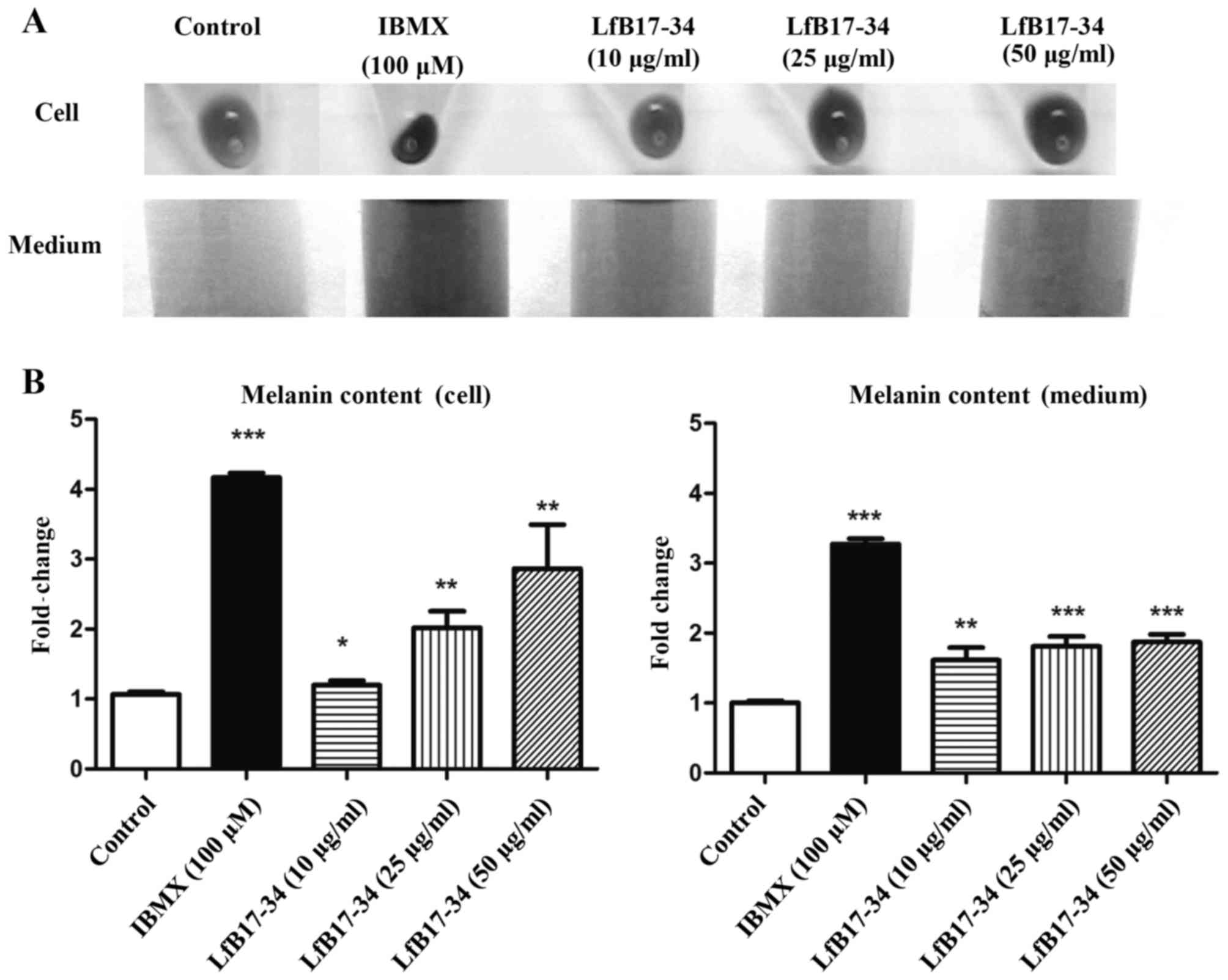
To extend our preliminary finding on LfB17-34, we
treated B16F10 cells with different concentrations of LfB17-34
ranging from 10 to 50 µg/ml and analyzed melanin contents in
cells and conditioned medium. We found that LfB17-34 was able to
enhance pigmentation of B16F10 cells in a dose-dependent manner
(Fig. 2A). To further quantify
the effect of LfB17-34 on pigmentation, we measured melanin content
in both cells and conditioned medium. The results revealed that
LfB17-34 significantly increased melanin content in both cells and
conditioned medium in a dose-dependent manner (Fig. 2B). The concentration of LfB17-34
at 10 µg/ml was sufficient to significantly enhance melanin
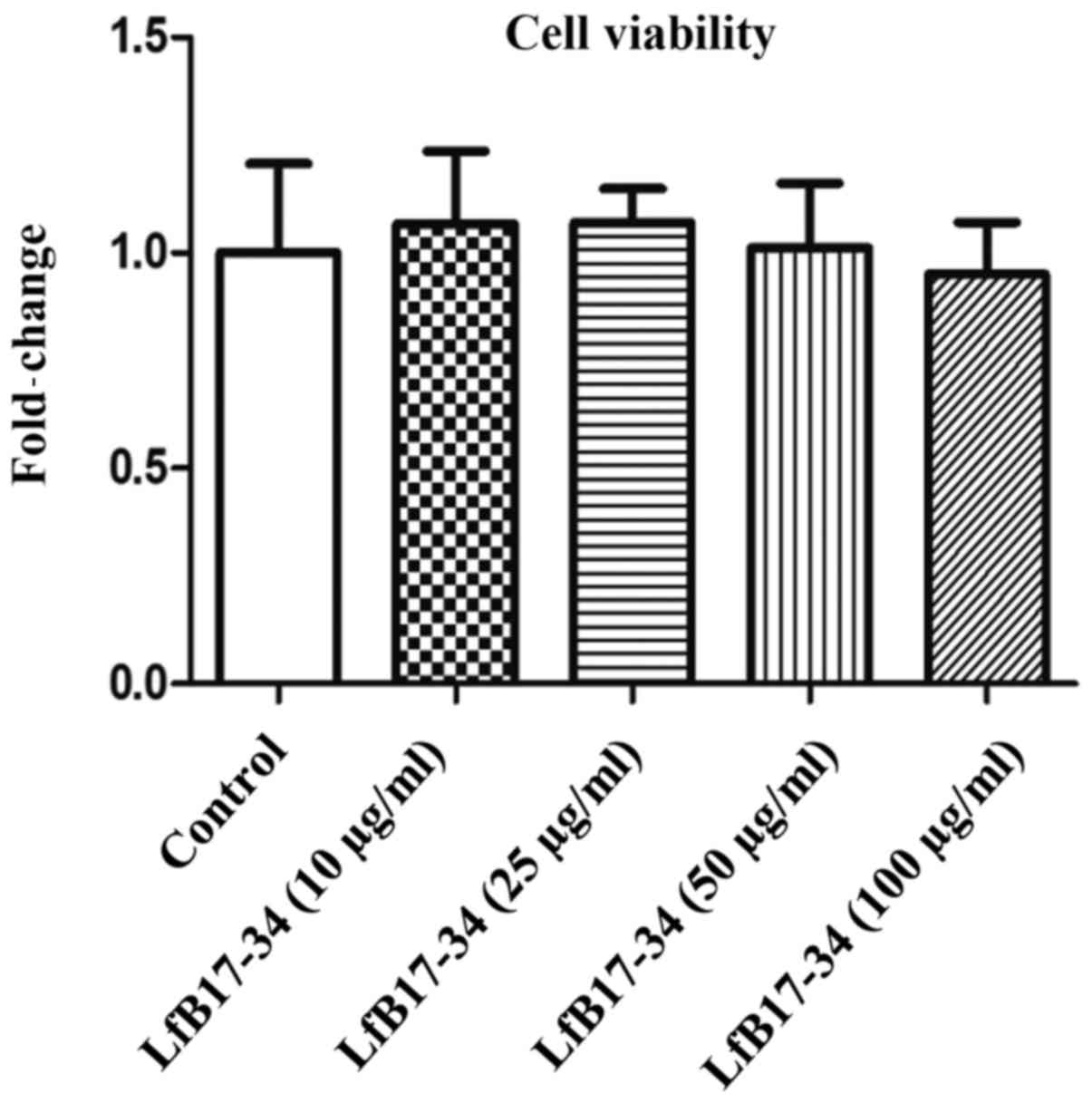
synthesis. To assess the toxicity of the LfB17-34 peptide, cell
viability was analyzed by MTT assay. The results revealed that
treatment of the B16F10 cells with 100 µg/ml of LfB17-34 for
48 h did not exert any significant effect on cell viability
(Fig. 3). These results suggest
that LfB17-34 enhances melanin synthesis and secretion in a
dose-dependent manner without significantly affecting cell
viability.
LfB17-34 increases tyrosinase
activity
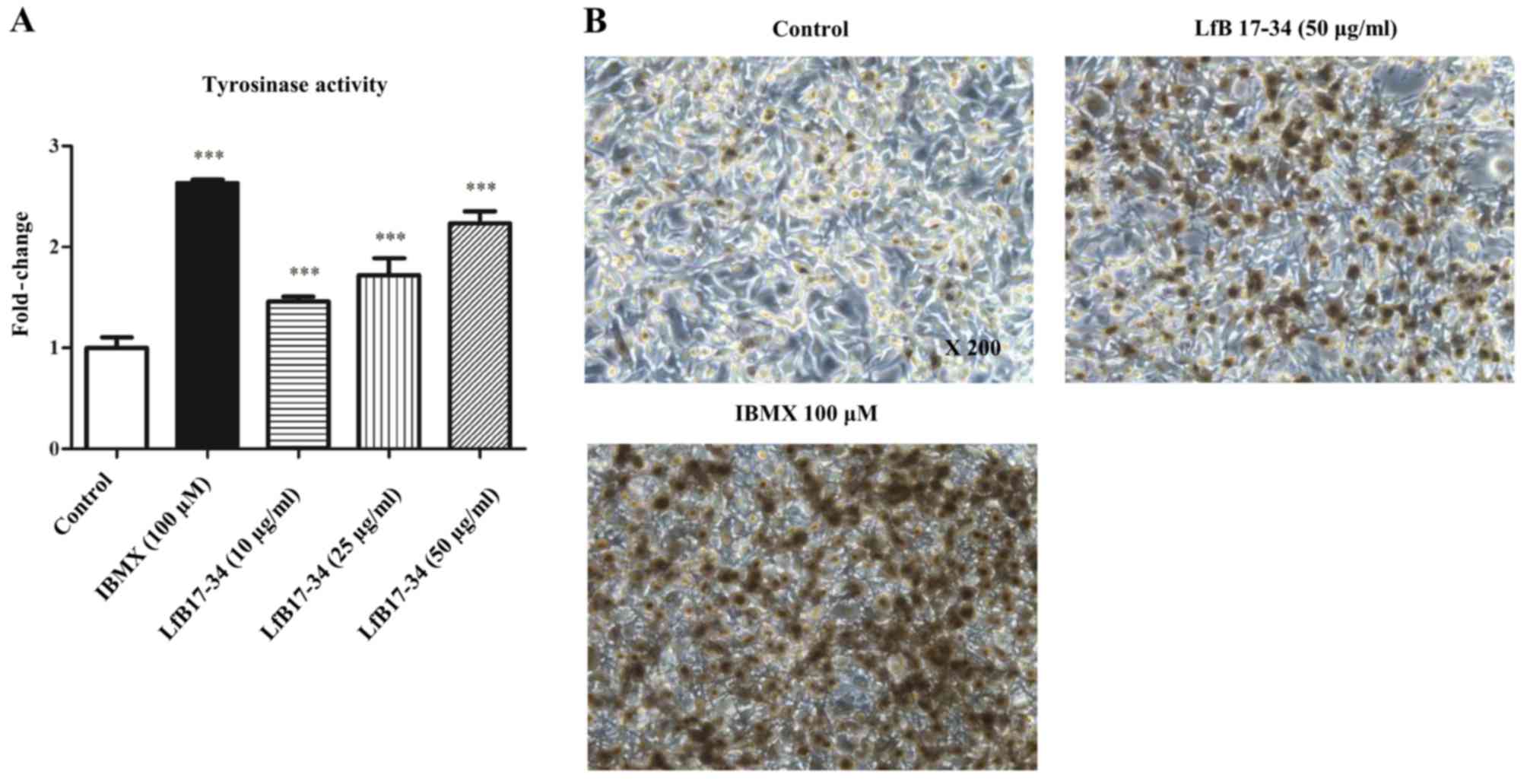
Since tyrosinase has been demonstrated to be the
most critical molecule to regulate melanin synthesis, we therefore
analyzed whether LfB17-34 could increase tyrosinase enzyme activity
by tyrosinase activity assay and L-DOPA staining. Consistent with
our findings in the effect on melanin synthesis, LfB17-34 was able
to enhance tyrosinase activity in a dose-dependent manner (Fig. 4A). L-DOPA staining of the cells
treated with LfB17-34 also confirmed this effect (Fig. 4B). Our results suggest that
LfB17-34 enhances tyrosinase activity in a dose-dependent
manner.
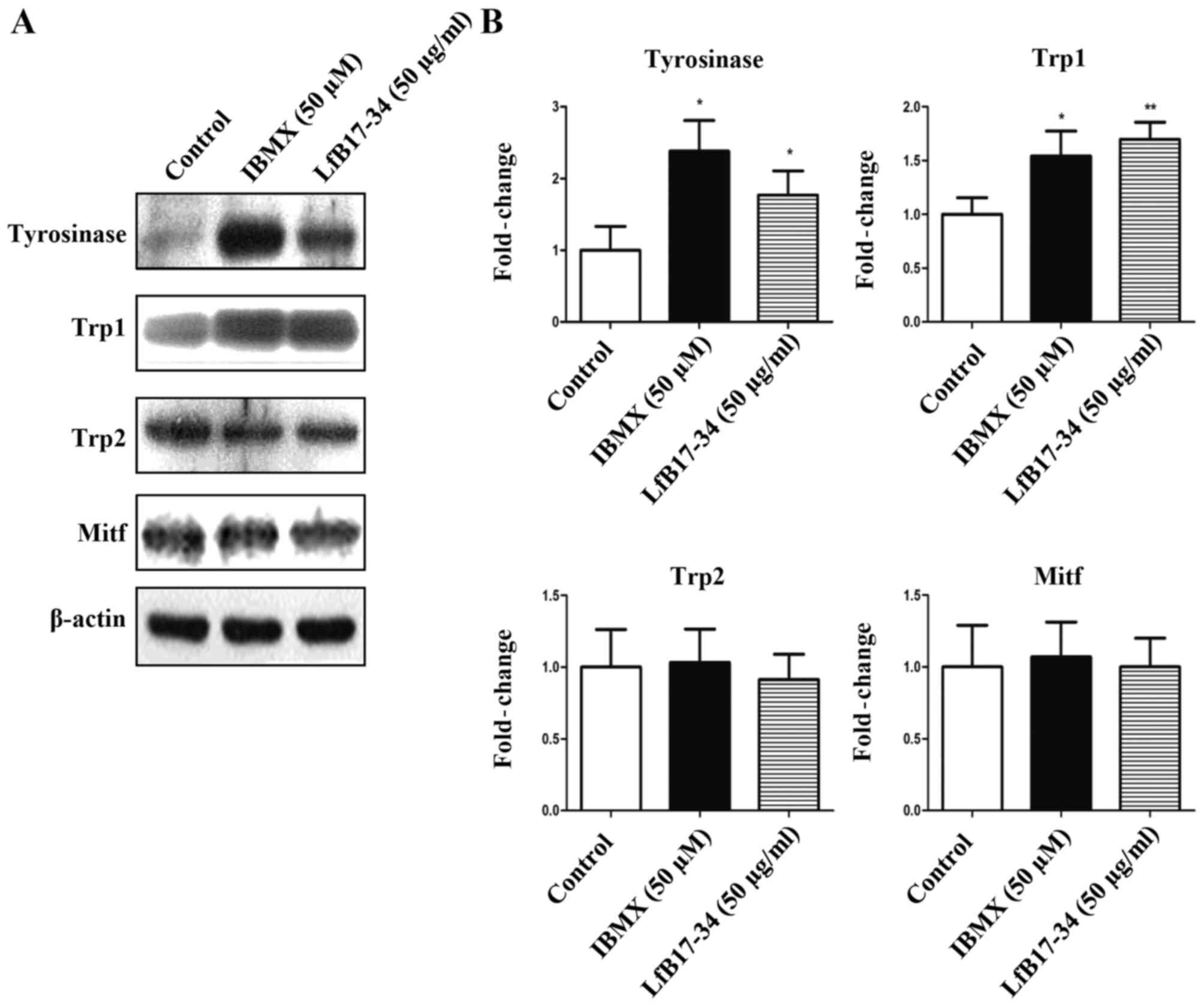
LfB17-34 increases the expression of
tyrosinase and Trp1
To investigate whether tyrosinase and other
melanogenesis-related proteins were induced by LfB17-34, we
performed western blot analysis and RT-qPCR. The results from
western blot analysis revealed that LfB17-34 significantly
increased the expression of tyrosinase and Trp1, but not that of
Trp2 and Mitf (Fig. 5). We
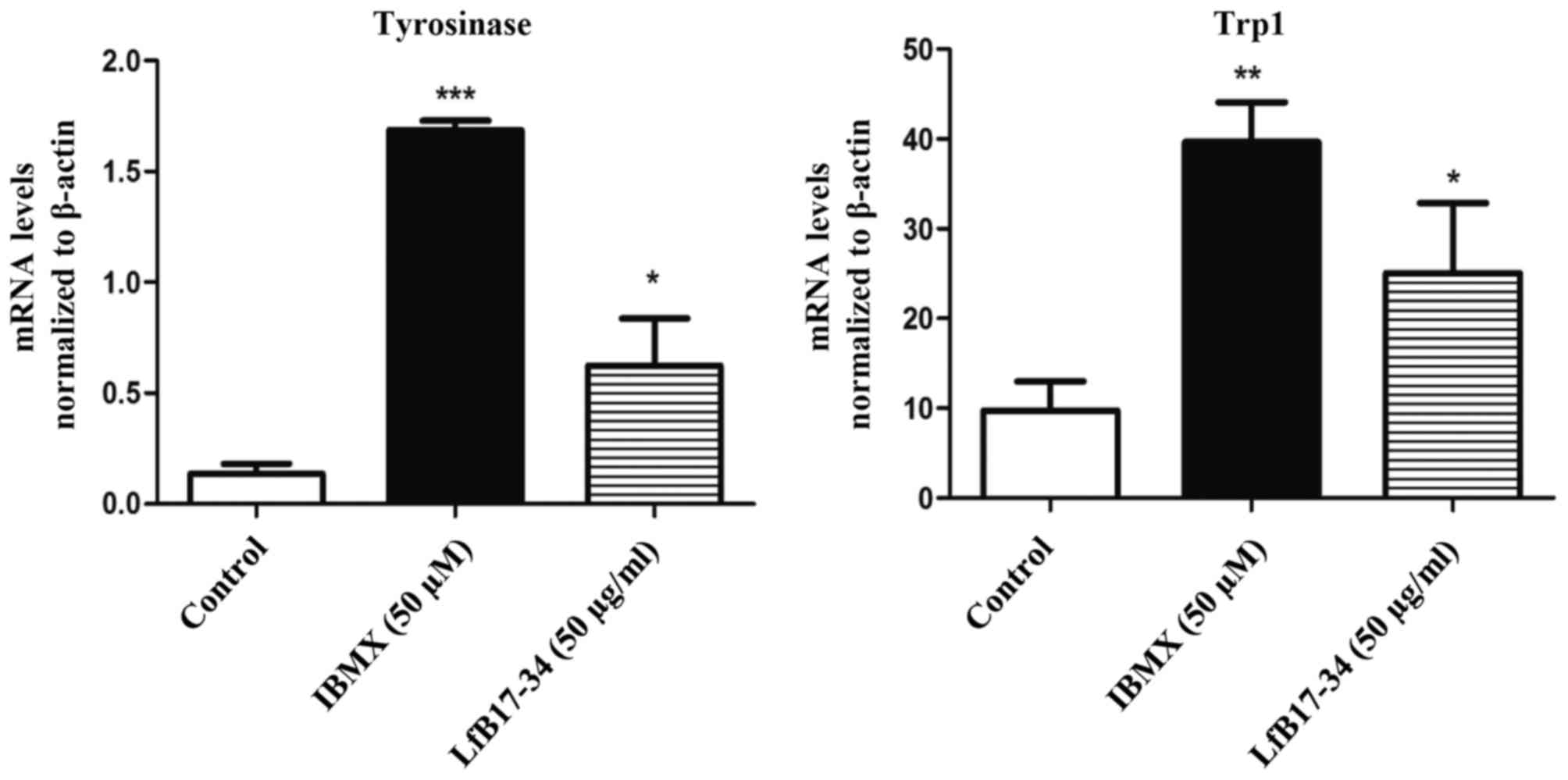
further assessed whether the mRNA levels of tyrosinase and Trp1
were also induced by LfB17-34. The results from RT-qPCR indicated
that LfB17-34 significantly increased the mRNA levels of both
tyrosinase and Trp1 (Fig. 6).
These results suggest that LfB17-34 increases both the mRNA and
protein levels of tyrosinase and Trp1.
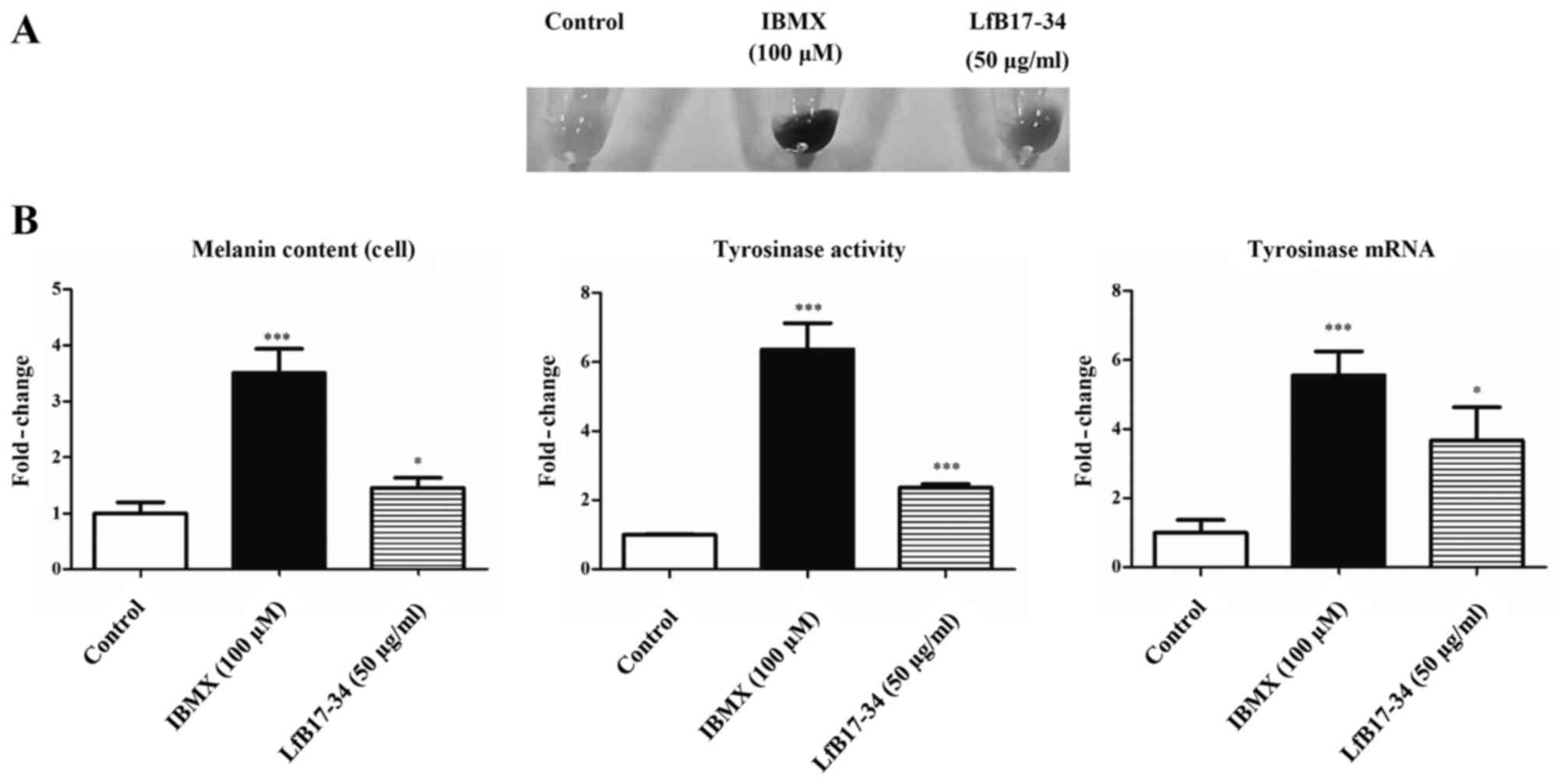
LfB17-34 enhances melanogenesis in B16F0
cells
To further confirm the effect of LfB17-34, we used
another melanoma cell line, B16F0. We found that LfB17-34 was also
able to increase cell pigmentation, melanin content, tyrosinase
activity, and tyrosinase mRNA expression (Fig. 7). These data further confirm that
LfB17-34 enhances melanogenesis.
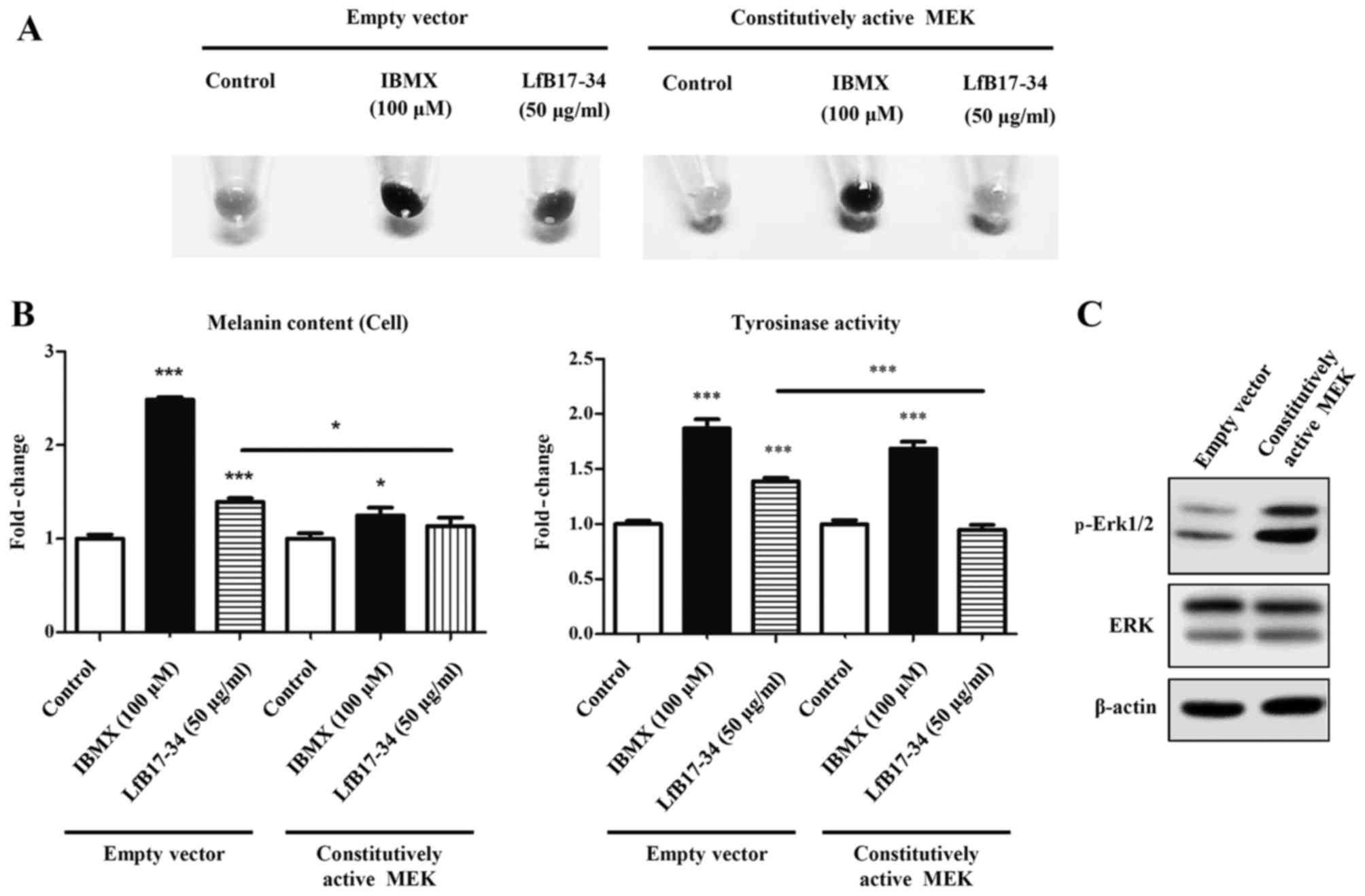
LfB17-34 inhibits the phosphorylation of
Erk
It is well known that the MAPK and Akt signaling
pathways are involved in tyrosinase expression and melanogenesis.
We therefore analyzed the phosphorylation of MAPK/Erk1/2, MAPK/p38
and Akt by western blot analysis. Of note, we found that LfB17-34
was able to decrease the phosphorylation levels of Erk1/2, but not
those of p38 and Akt (Fig. 8). It
has been demonstrated that Erk1/2 signaling negatively regulates
melanin synthesis in B16F10 cells (19,20). To further determine whether
p-Erk1/2 is involved in LfB17-34-induced melanogenesis, we
transfected the B16F10 cells with an expression vector coding for
constitutively active MEK. As expected, we found that
constitutively active MEK reversed the LfB17-34-increased
pigmentation, melanin content and tyrosinase activity (Fig. 9). Although the fold-changes in
melanin content and tyrosinase activity varied in different
experiments, the effects of LfB17-34 showed a similar tendency.
Therefore, these findings suggest that the mechanism by which
LfB17-34 enhances melanin synthesis may be mediated by the decrease
in p-Erk1/2 levels.
Discussion
Bovine Lf has been reported to exhibit a
depigmenting effect on mouse melanoma cells (12). Unexpectedly, in this study, we
found that LfB17-34, a bovine Lf-derived peptide, increased
pigmentation in mouse melanoma cells. To the best of our knowledge,
this is the first study showing that a Lf-derived peptide can
stimulate pigmentation. We found that LfB17-34 significantly
induced the expression of pigmentation-related genes, such as
tyrosinase and Trp1. Moreover, LfB17-34 significantly inhibited Erk
signaling, which negatively regulates melanogenesis. Therefore, we
propose that LfB17-34 enhances pigmentation through increased
expression of tyrosinase and Trp1, which is probably via decreased
phospho-Erk.
Tyrosinase has been well demonstrated to be the
crucial enzyme responsible for melanin synthesis. Hence,
identifying molecules that can regulate tyrosinase expression and
activity has been considered to be the key strategy to develop
agents for treating pigmentation disorders. In this study, we found
that LfB17-34 enhanced the expression of tyrosinase mRNA and
protein as well as tyrosinase activity without significant
cytotoxicity. Furthermore, the effect of LfB17-34 on tyrosinase
activity and melanin content was dose-dependent. These results
suggest that LfB17-34 induces pigmentation primarily through
increased tyrosinase expression and activity and that LfB17-34 has
great potential for drug development.
Mitf is regulated both transcriptionally and
post-translationally (6). It is
well known that cAMP activates PKA and in turn phosphorylates CREB,
leading to activate the transcription of Mitf. However, in this
study LfB17-34 did not significantly change phospho-CREB levels
(data not shown). At the post-translational level, it has been
reported that Mitf is phosphorylated at Ser73 by MAPK/Erk (18), which is known to facilitate
ubiquitination and degradation of Mitf. Consistent with this
finding, activation of Erk has been reported to decrease Mitf
expression and melanogenesis, whereas inhibition of Erk activity by
PD98059 increases melanin production in B16F10 cells (19,20). However, we consistently observed
that LfB17-34 decreased phosphorylation of Erk but did not increase
Mitf protein levels in B16F10 cells. In this case, LfB17-34 still
could induce tyrosinase and Trp1 expression and enhance
pigmentation. Similarly, we observed that 50 µM IBMX
increased pigmentation without elevated protein levels of
tyrosinase and Trp1. There could be several explanations. Firstly,
it is likely that LfB17-34 exerts its stimulatory effect on
pigmentation by increasing the Mitf activity instead of the Mitf
protein level via Erk signaling. Secondly, it is also possible that
LfB17-34 induces melanin synthesis independent of Mitf. It is well
known that tyrosinase, Trp1, and Trp2 are transcriptional targets
of Mitf. However, LfB17-34 increased tyrosinase and Trp1
expression, but not Mitf and Trp2. Therefore, LfB17-34 may modulate
other factors that can induce tyrosinase and Trp1 expression to
enhance pigmentation. To understand the complicated process of
melanogenesis, it will be of great interest to further investigate
the detailed mechanism of LfB17-34-mediated pigmentation.
Among 5 LfcinB-related peptides analyzed, LfcinB
(aa17-41), LfB17-24, and LfB17-34 exhibit stimulatory effect on
pigmentation. LfB20-29 and LfB25-34 do not have any effects. These
results suggest that the 8 amino acids (FKCRRWQW) in LfB17-24 could
be essential for stimulating pigmentation. Moreover, we found that
LfB17-34 exhibits stronger stimulatory effect than does LfcinB
(aa17-41), suggesting that aa35-41 (TCVRRAF) could have inhibitory
effect on pigmentation. Although the underlying mechanism remains
to be further investigated, we speculate that LfB17-34 could form a
specific conformation to interact with the surface molecules that
can stimulate pigmentation signaling, such as elevated cAMP and/or
decreased p-Erk.
LfcinB exhibits a diverse range of bioactivities.
Therefore, LfcinB-related peptides are attractive targets for
developing peptide drugs. In this study, and to the best of our
knowledge, we are the first to report LfB17-34 as a novel
stimulator of pigmentation. Increased expression of crucial
melanogenic enzymes, tyrosinase and Trp1, as well as decreased Erk
phosphorylation could be involved in the underlying mechanism of
LfB17-34-induced melanogenesis. Further investigation of LfB17-34
will provide great opportunities to develop drugs for treating
hypopigmentation disorders, such as graying of hair and vitiligo in
the future.
Acknowledgments
This study was supported by the grant from National
Taiwan University (no. 105A118).
References
|
1
|
Tobin DJ: The cell biology of human hair
follicle pigmentation. Pigment Cell Melanoma Res. 24:75–88. 2011.
View Article : Google Scholar
|
|
2
|
Dessinioti C, Stratigos AJ, Rigopoulos D
and Katsambas AD: A review of genetic disorders of
hypopigmentation: Lessons learned from the biology of melanocytes.
Exp Dermatol. 18:741–749. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Panhard S, Lozano I and Loussouarn G:
Greying of the human hair: A worldwide survey, revisiting the '50'
rule of thumb. Br J Dermatol. 167:865–873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ezzedine K, Eleftheriadou V, Whitton M and
van Geel N: Vitiligo. Lancet. 386:74–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Falabella R and Barona MI: Update on skin
repigmentation therapies in vitiligo. Pigment Cell Melanoma Res.
22:42–65. 2009. View Article : Google Scholar
|
|
6
|
Hsiao JJ and Fisher DE: The roles of
microphthalmia-associated transcription factor and pigmentation in
melanoma. Arch Biochem Biophys. 563:28–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gifford JL, Hunter HN and Vogel HJ:
Lactoferricin: A lactoferrin-derived peptide with antimicrobial,
antiviral, antitumor and immunological properties. Cell Mol Life
Sci. 62:2588–2598. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Centeno JM, Burguete MC, Castelló-Ruiz M,
Enrique M, Vallés S, Salom JB, Torregrosa G, Marcos JF, Alborch E
and Manzanares P: Lactoferricin-related peptides with inhibitory
effects on ACE-dependent vasoconstriction. J Agric Food Chem.
54:5323–5329. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fernández-Musoles R, López-Díez JJ,
Torregrosa G, Vallés S, Alborch E, Manzanares P and Salom JB:
Lactoferricin B-derived peptides with inhibitory effects on
ECE-dependent vasoconstriction. Peptides. 31:1926–1933. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ruiz-Giménez P, Ibáñez A, Salom JB, Marcos
JF, López-Díez JJ, Vallés S, Torregrosa G, Alborch E and Manzanares
P: Antihypertensive properties of lactoferricin B-derived peptides.
J Agric Food Chem. 58:6721–6727. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan D, Chen D, Shen J, Xiao G, van Wijnen
AJ and Im HJ: Bovine lactoferricin is anti-inflammatory and
anti-catabolic in human articular cartilage and synovium. J Cell
Physiol. 228:447–456. 2013. View Article : Google Scholar
|
|
12
|
Nakajima M, Shinoda I, Samejima Y,
Miyauchi H, Fukuwatari Y and Hayasawa H: A simple and quantitative
cell-blotting assay for evaluation of pigmentation of cultured
melanocytes and its use in demonstrating the depigmenting effect of
lactoferrin. Arch Dermatol Res. 289:180–183. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsu WM, Che MI, Liao YF, Chang HH, Chen
CH, Huang YM, Jeng YM, Huang J, Quon MJ, Lee H, et al: B4GALNT3
expression predicts a favorable prognosis and suppresses cell
migration and invasion via β1 integrin signaling in
neuroblastoma. Am J Pathol. 179:1394–1404. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee J, Jung E, Park J, Jung K, Park E, Kim
J, Hong S, Park J, Park S, Lee S, et al: Glycyrrhizin induces
melanogenesis by elevating a cAMP level in b16 melanoma cells. J
Invest Dermatol. 124:405–411. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bellei B, Maresca V, Flori E, Pitisci A,
Larue L and Picardo M: p38 regulates pigmentation via proteasomal
degradation of tyrosinase. J Biol Chem. 285:7288–7299. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang D, Xu X, Ma H, Yue X, Li C and Zhu W:
Optimization of the method for the culture of melanocyte precursors
from hair follicles and their activation by 1,25-dihydroxyvitamin
D3. Exp Ther Med. 6:967–972. 2013.PubMed/NCBI
|
|
17
|
Che MI, Huang J, Hung JS, Lin YC, Huang
MJ, Lai HS, Hsu WM, Liang JT and Huang MC: β1,
4-N-acetylgalactosaminyltrans ferase III modulates cancer stemness
through EGFR signaling pathway in colon cancer cells. Oncotarget.
5:3673–3684. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu M, Hemesath TJ, Takemoto CM, Horstmann
MA, Wells AG, Price ER, Fisher DZ and Fisher DE: c-Kit triggers
dual phosphorylations, which couple activation and degradation of
the essential melanocyte factor Mi. Genes Dev. 14:301–312.
2000.PubMed/NCBI
|
|
19
|
Bae JS, Han M, Yao C and Chung JH:
Chaetocin inhibits IBMX-induced melanogenesis in B16F10 mouse
melanoma cells through activation of ERK. Chem Biol Interact.
245:66–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Son YO, Lee SA, Kim SS, Jang YS, Chun JC
and Lee JC: Acteoside inhibits melanogenesis in B16F10 cells
through ERK activation and tyrosinase down-regulation. J Pharm
Pharmacol. 63:1309–1319. 2011. View Article : Google Scholar : PubMed/NCBI
|