Introduction
Appendages of the fetus, included the amnion, are
normally discarded after delivery as medical waste. A large
quantity of human amniotic epithelial and mesenchymal stem cells
can be obtained non-invasively from the amniotic membrane, which
represents an advantageous source of cells for cell therapy
(1).
In vivo studies have previously reported the
therapeutic potential of stem cells using various animal models
including hindlimb ischemia (2,3),
wound healing (4,5) and myocardial infarction (6,7).
However, in many cases, the frequency of stem cell engraftment and
the number of newly generated adult cells, either by
transdifferentiation or cell fusion, appear to be too low to
explain the significant improvement described (8,9).
Meanwhile, tissue concentrations of proteins, including vascular
endothelial growth factor (VEGF) and basic fibroblast growth factor
(bFGF) are increased in the injured areas treated with stem cells
(10). There is a growing body of
evidence supporting the hypothesis that paracrine mechanisms
mediated by factors released by pluripotent stem cells play an
essential role in the reparative process (11,12). This paracrine effect renders these
cells an attractive therapeutic source for regenerative
medicine.
Stem cells may be beneficial in various
cell-therapeutic approaches where they function by promoting the
survival of endothelial cells (13,14), the stabilization of pre-existing
vessels (15), and the
revascularization of ischemic tissues (2,3).
Given that the natural response to tissue repair is such a complex
process, many growth factors may be involved. Thus, a great deal of
interest has arisen in angiogenetic factors found in stem cells,
such as hepatocyte growth factor (HGF), epidermal growth factor
(EGF), heparin binding EGF like growth factor (HB-EGF) and insulin
growth factor-1 (IGF-1), and the paracrine effects which are
significantly related to the angiogenesis of endothelial cells
(3,16–18).
The aim of the present study was: i) to isolate and
characterize cells from human amnions; ii) to investigate the
biological potential and behavior of these cells in regards to the
function of endothelial cells in vivo and in vitro;
and iii) to examine the variations in the expression profile of
growth factors in different human amnion-derived cell types.
Materials and methods
Ethics
The amnion samples discarded after Caesarean
sections were collected from the Department of Obstetrics and
Gynecology, The First at Hospital of China Medical University
(Shenyang, China). This study was approved by the Ethics Committee
of the First Affiliated Hospital of China Medical University.
Written informed consent was obtained from all of the patients
prior to their participation. C57BL/6J mice were obtained from the
Experimental Animal Centre of China Medical University. All
experiments and animal care were approved by the Ethics Committee
of the First Affiliated Hospital of China Medical University.
Cells and cell culture
Human aorta endothelial cells
(hAoECs)
hAoECs were purchased from ScienCell (Carlsbad, CA,
USA) and cells at passage 4–6 were used for in vitro
experiments. The cells were cultured in Endothelial Basal Media-2
(EBM-2) with 5% fetal bovine serum (FBS) and Endothelial Cell
Growth Supplement (ECGS) (EGM-2; ScienCell).
Human amniotic epithelial cells
(hAECs)
Primary cell culture was performed as described
previously (5). Briefly, amnions
were manually separated and washed with phosphate-buffered saline
(PBS) supplemented with 100 U/ml penicillin and streptomycin.
Amnions were then incubated with 0.25% trypsin solution for 30 min.
This process was repeated three times. Supernatants were collected
and centrifuged for 5 min at 1,000 rpm to obtain a cell pellet.
Those cells were plated on a culture flask (designated as hAEC P0)
in Dulbecco's modified Eagle's medium (DMEM; HyClone, Logan, UT,
USA), and 100 U/ml penicillin and streptomycin. In this study,
hAECs at passage 2–3 were used.
Human amniotic mesenchymal stem cells
(hAMSCs)
The amnion tissue was cut into small pieces, and
then incubated with 1 mg/ml collagenase IV (Sigma-Aldrich, St.
Louis, MO, USA) and 0.1 mg/ml DNase (Takara Bio, Inc., Shiga,
Japan) at 37°C for 20 min. FBS was then added to stop digestion,
and supernatants were filtered through a cell strainer (200
μm) and centrifuged for 5 min at 1,000 rpm. Cells were
plated on a culture flask (designated as hAMSC P0) in DMEM
containing 10% FBS (both from HyClone), 10 mmol/ml FGF-2
(PeproTech, Rocky Hill, NJ, USA), and 100 U/ml penicillin and
streptomycin. The culture medium was changed 48 h later. In this
study, hAMSCs at passages 3–6 were used for the functional
experiment.
The differentiation capacity of
amniotic cells
hAECs (passages 2) and hAMSCs (passage 3) were
tested for their ability to differentiate into osteocytes,
chondrocytes and adipocytes.
To induce differentiation into osteocytes, the cells
were cultured in osteocyte differentiation medium: 1 μM
dexamethasone, 50 μg/ml L-ascorbate, and 10 mM
β-glycerophosphate (Sigma-Aldrich) in DMEM supplemented with 10%
FBS. After 14 days of differentiation, the cells were fixed and
stained with Alizarin Red S (Cyagen, Guangzhou, China).
To induce differentiation into adipocytes, the cells
were cultured with adipocyte differentiation medium: 0.5 mM
3-isobutyl-1-methyl xanthine, 1 μM dexamethasone, 200
μM indomethacin, and 10 μg/ml insulin (Sigma-Aldrich)
in DMEM supplemented with 10% FBS. After 14 days of
differentiation, the cells were stained with Oil Red O
(Cyagen).
To induce differentiation into chondrocytes, the
cells were cultured with chondrocyte differentiation medium: 0.1
μM dexamethasone, 50 μg/ml L-ascorbate, 100
μg/ml sodium pyruvate (Sigma-Aldrich), and 10 ng/ml
transforming growth factor (TGF)-β1 (PeproTech) in DMEM
supplemented with 10% FBS. After 14 days of differentiation, the
cells were stained with Alcian blue (Cyagen).
Flow cytometry and immunofluorescence
of cells
Flow cytometry and immunofluorescence were used to
identify the characteristics of the cells and detect stem
cell-related cell surface markers. For flow cytometry, the cells
(106 cells/100 μl) were collected and incubated
with monoclonal phycoerythrin (PE)-conjugated antibodies for CD29
(cat. no. 303003), CD31 (cat. no. 303105), CD34 (cat. no. 343505),
CD44 (cat. no. 338807), CD45 (cat. no. 368509), CD49d (cat. no.
304303), CD73 (cat. no. 344003), CD90 (cat. no. 32810), CD105 (cat.
no. 323205), HLA-DR (cat. no. 307605), SSEA-4 (cat. no. 330405),
SOX-2 (cat. no. 656103) and OCT-4 (cat. no. 653703) (BioLegend, San
Diego, CA, USA) for 30 min on ice. Appropriate isotype-matched
antibodies were used as negative controls (BD Biosciences, San
Jose, CA, USA). Data from 10,000 viable cells were acquired. List
mode files were analyzed by FCS Express software (BD Biosciences).
For immunofluorescence, cells growing on the glass slide were
stained with anti-cytokeratin 19 (cat. no. ab52625, 1:200; Abcam,
Cambridge, MA, USA) and secondary antibody (cat. no. A-11034,
1:200; Alexa 488, donkey anti-rabbit; Life Technologies, Carlsbad,
CA, USA). Nuclei were stained with DAPI (Beyotime, Shanghai,
China). Cells on the glass slide were photographed using an
inverted fluorescence microscope (Carl Zeiss, Oberkochen,
Germany).
Preparation of conditioned medium
To generate conditioned medium (CdM), hAoECs, hAMSCs
and hAECs were cultured with EGM-2. After the cells had reached
~50–60% confluence (~4×105 cells in 25 cm2
flask), cultures were gently rinsed three times with PBS and the
medium was replaced with EGM-2 or EBM-2. After 48 h, the CdM
(EGM-2) obtained from each plate was then collected, pooled for
each cell type, centrifuged at 1,000 × g for 5 min, filtered (0.2
μm) to remove cellular debris, stored at −80°C and
supernatants were used as CdM-hAoEC, CdM-hAEC and CdM-hAMSC for
cell assays. Positive control, non-conditioned medium (non-CdM) was
generated in the same way as above, except that no cells were
cultured in the plates. Batches of 2X concentrated CdM (EBM-2) were
also prepared for the in vivo Matrigel plug assay. In this
way, a final concentration of 1X CM after 1:1 dilution in Matrigel
was obtained.
Cell viability assays
For the growth curves of hAECs and hAMSCs, cells
(5×103/well) were plated in 96-well plates with EGM-2.
Cells were cultured for 7 days, and cell proliferation was measured
using Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto, Japan) every
day according to the manufacturer's protocol. For determining the
effect of CdM on endothelial cell viability, hAoECs were cultured
in EGM-2 without FBS for 24 h to arrest mitosis. Then, hAoECs
(2×104/well) were plated in 96-well plates, the medium
was replaced with CdM-hAoEC (control), CdM-hAEC, CdM-hAMSC and
EGM-2 (positive control). Cells were cultured for 24 h, after which
hAoEC proliferation was measured using the CCK-8 (Dojindo). In
brief, cells were incubated with CCK-8 for 1.5 h at 37°C. The
staining intensity in the medium was measured by determining the
absorbance at 450 nm.
Cell cycle analysis
The effect of CdM on cell cycle distribution was
determined by flow cytometry. Briefly, hAoECs were treated with
different CdM for 24 h. Cells were suspended, washed with PBS,
centrifuged, and fixed in 70% ethanol at −20°C overnight. Cells
were then resuspended in 500 μl of dyeing buffer containing
10 μl RNase A and 25 μl PI (Beyotime). Cells were
incubated in the dark for 30 min at room temperature. A total of
1×104 cells were subjected to cell cycle analysis using
a flow cytometer (BD Biosciences).
Scratch wound closure assay
hAoECs (500,000 cells/insert) were plated in 6-well
plates, and at 80–90% confluence, at 12 h after plating, a scratch
of ~0.5 mm was created using a sterile pipette tip. Each well was
washed twice with PBS and then the cell culture medium was either
replaced with CdM-hAoEC (control), CdM-hAEC, CdM-hAMSC or EGM-2
(positive control). Cell migration into the scratch was
photographed at 0, 6 and 24 h using an inverted microscope
(Olympus, Tokyo, Japan). Results were analyzed with Image-Pro Plus
software 6.0 (IPP; Media Cybernetics, Inc., Rockville, MD, USA).
The results are presented as the percentage of wound healing, which
was calculated as follows: [Wound area (initial) − Wound area
(final)]/Wound area (initial) × 100% (5).
Transwell migration assay
Cell migration assays were performed using inserts
with 8-μm pore-sized membranes in a 24-well plate (Corning
Costar, Lowell, MA, USA). CdM-hAoEC (control), CdM-hAEC, CdM-hAMSC
and EGM-2 (positive control) were placed in the bottom chamber.
hAoECs were resuspended in serum-free EGM-2, transferred onto the
filter of the insert (50,000 cells/insert) and incubated at 37°C in
5% CO2 for 6 or 24 h. Non-migratory cells were removed
from the upper side of the filter. Migratory cells at the bottom
side of the filter were fixed with paraformaldehyde (PFA), stained
with hematoxylin and eosin (H&E) and photographed. Cells were
counted from eight randomly selected regions/well.
Matrigel tube formation assay
To evaluate the tube formation potential, hAoECs
were seeded with each CdM derived from different cells, at a
concentration of 2.5×104 cells/well in growth
factor-reduced basement membrane matrix gel (Matrigel; BD
Biosciences)-coated 96-well plates. After 6, 24 and 48 h of
incubation, representative fields were photographed using inverted
microscopy (Olympus), and branching points from each sample were
examined with Image-Pro Plus software 6.0.
Matrigel plug assays
The CdM (EBM-2) was mixed with 250 μl of
liquid Matrigel-reduced growth factor (BD Matrigel 356230) at a
ratio of 1:1 at 4°C. Mice (8-weeks old) received a total of 500
μl of this mixture subcutaneously in the dorsal region,
generating Matrigel plugs when warmed to body temperature. Plugs
were recovered 1 week later.
qPCR assay for growth factor and
cytokine detection
Total RNA was extracted, using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA), from hAECs and hAMSCs cultured
with EGM-2 complete medium. RNA concentration was determined by
NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, DE, USA). cDNA
was synthesized using PrimeScript™ RT reagent (Takara Bio, Inc.).
Reactions were performed using the SYBR PrimeScript RT-PCR kit
(Takara Bio, Inc.) with an ABI 7500 Sequence Detection system
(Applied Biosystems, Foster City, CA, USA). As an internal control,
the β-actin level was quantified in parallel with the target genes.
Normalization and fold-changes were calculated using the ΔΔCq
method. The primers used for real-time PCR are the following:
5′-CTGTCTAATGCCCTGGAGCC/ACGCGAGTCTGTGTTTTTGC-3′ for VEGFA;
5′-TCAGCCAGCAGATGGGAATG/TCAGGGCTGTATGG GCAAAG-3′ for EGF;
5′-GGCTGTACTGCAAAAACGGG/TAGCTTGATGTGAGGGTCGC-3′ for bFGF;
5′-CAATGCCTCTGGTTCCCCTT/TGTTCCCTTGTAGCTGCGTC-3′ for HGF;
5′-AGTTCTCTCGGCACTGGTGA/TAGCAGCTG GTCCGTGGATA-3′ for HB-EGF;
5′-ATCAGCAGTCTTCCAACCCA/GAGATGCGAGGAGGACATGG-3′ for IGF-1;
5′-AGGATTCCTATGTGGGCGAC/ATAGCACAGCCTGGATAGCAA-3′ for β-actin.
Enzyme-linked immunosorbent assay
(ELISA) detection of angiogenetic growth factors
Conditioned medium from hAoECs, hAECs and hAMSCs
were collected after 48 h incubation. The concentration of
cytokines in the different CdM was measured using sandwich ELISA
kits (VEGFA, EGF, bFGF, HGF, HB-EGF and IGF-1; R&D Systems,
Minneapolis, MN, USA). After media were collection, the cells were
counted. ELISA values were corrected for total cell numbers.
Positive control-conditioned medium was also assayed.
Statistical analysis
All experiments were performed 3 times on amniotic
cells and with CdM from 3 different donors. The data are shown as
the means ± SDs. Comparisons between groups were analyzed using
t-test. Comparisons of parameters for more than three groups were
made by one-way analysis of variance (ANOVA) followed by the
Bonferroni test. All statistical analyses were performed using SPSS
17.0 computer software. P-values <0.05 were considered to
indicate statistically significant differences.
Results
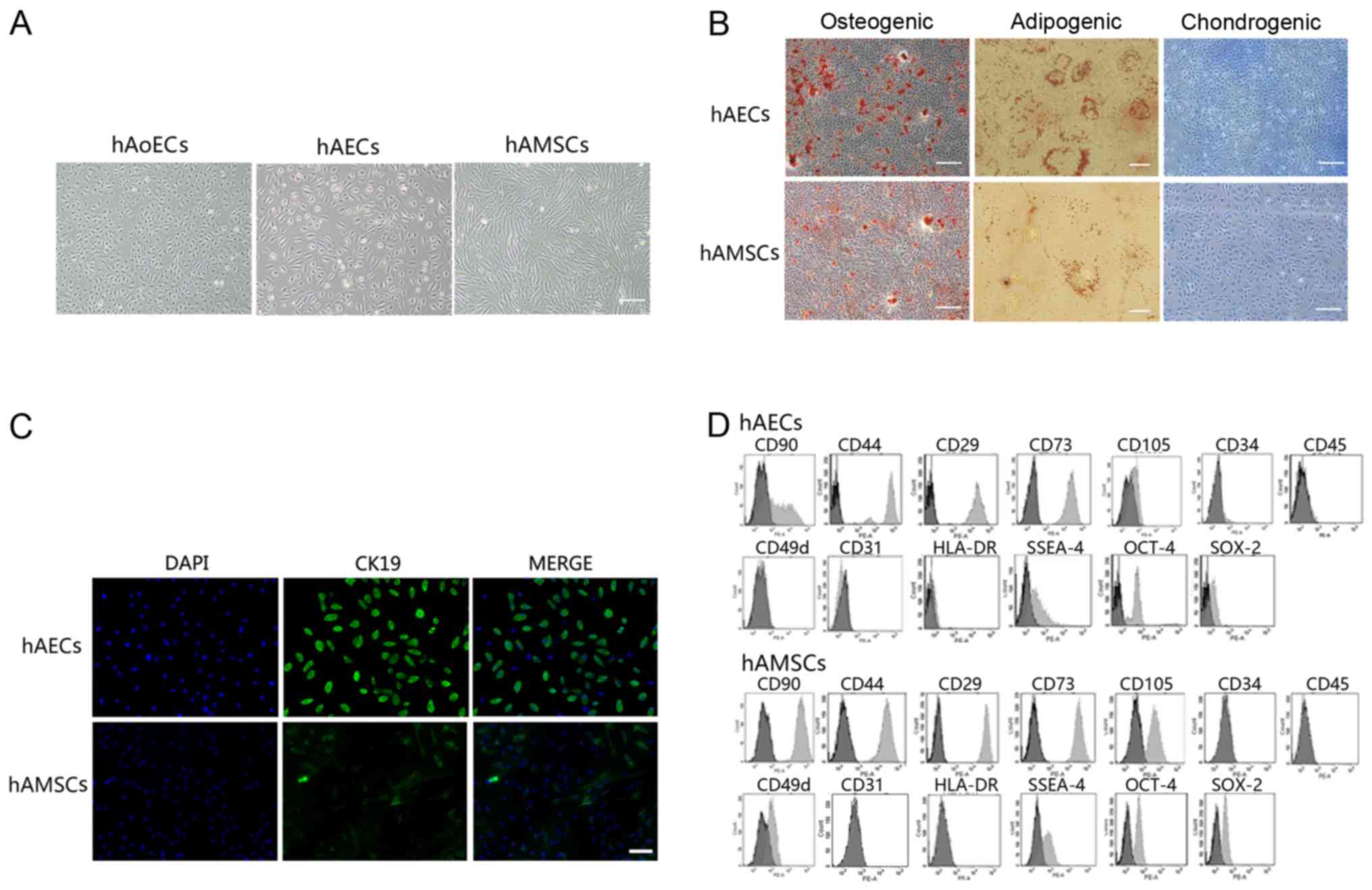
Characterization of hAECs and hAMSCs
hAECs exhibited a cobblestone-like morphology,
similar to hAoECs. Cultured hAMSCs showed a spindle fibroblast-like
morphology (Fig. 1A). Flow
cytometry and immunofluorescence revealed the expression of surface
markers. hAECs were positive for CK19, CD29, CD44, CD73, CD90 and
CD105, but were negative for CD31, CD34, CD45 and CD49d. hAMSCs
were positive for CD29, CD44, CD49d, CD73, CD90 and CD105, but were
negative for CK19, CD31, CD34 and CD45 (Fig. 1C and D). In addition, amniotic
cells were all negative for HLA-DR, indicating that these cells
possess low immunogenicity. To confirm the stem cell
characteristics of hAECs and hAMSCs, we performed FACS analysis
using embryonic stem and germ cell markers. Amniotic cells were all
found to express SSEA-4, SOX2 and OCT-4 (Fig. 1D). These results are consistent
with previously reported data (1,19,20). In addition, hAECs and hAMSCs could
differentiate into osteocytes, adipocytes and chondrocytes, as
demonstrated by positive Alizarin Red, Oil Red O and Alcian blue
staining, respectively (Fig. 1B),
which indicated that cultured amniotic cells possess stem cell
characteristics.
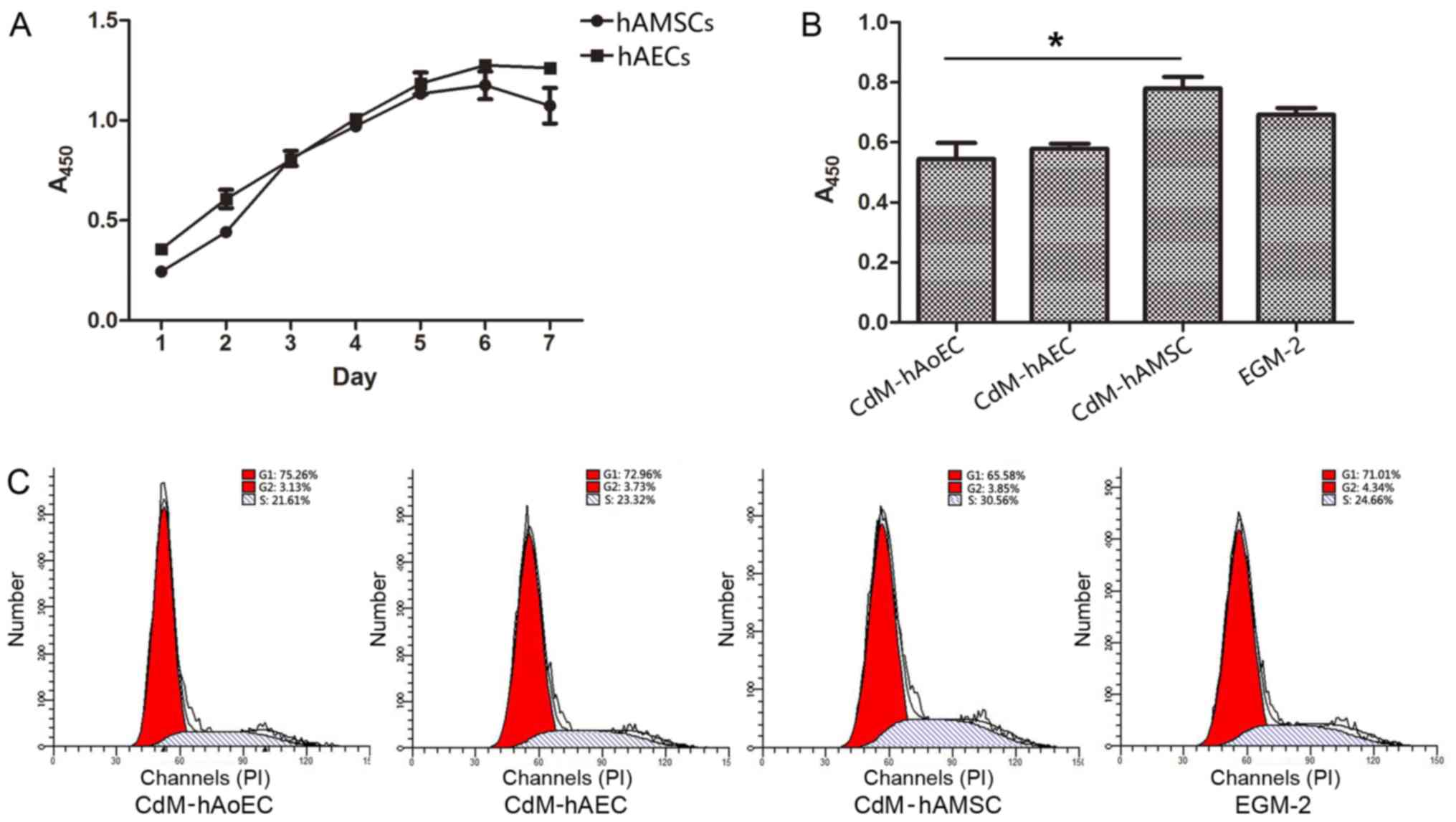
Culture medium from hAMSCs enhances
proliferation ability
We tested the growth kinetics of hAECs and hAMSCs.
Our data showed that when cultured with EGM-2, cells obtained from
the same amnion had a similar proliferation ability/day (Fig. 2A). Since the proliferation of
hAoECs is an important aspect of angiogenesis, we compared the
proliferation ability of hAoECs when stimulated with different
CdMs. The CCK-8 assay revealed that hAoECs cultured in CdM-hAMSC
showed enhanced proliferation compared with the other CdMs, even
the positive control EGM-2. However, there was no significant
difference in hAoEC proliferation between CdM-hAEC and CdM-hAoEC
(Fig. 2B). In order to further
confirm the effect of CdM on the proliferation ability of hAoECs,
we examined the effect of CdM on cell cycle distribution using flow
cytometry. Compared to the basal level (21.61±1.54%), hAoECs
treated with CdM-hAMSC led to a marked increase in the number of
cells in the S phase (30.56±1.91%) (Fig. 2C). There was no statistical
difference, however, when hAoECs were treated with CdM-hAEC or
CdM-hAoEC.
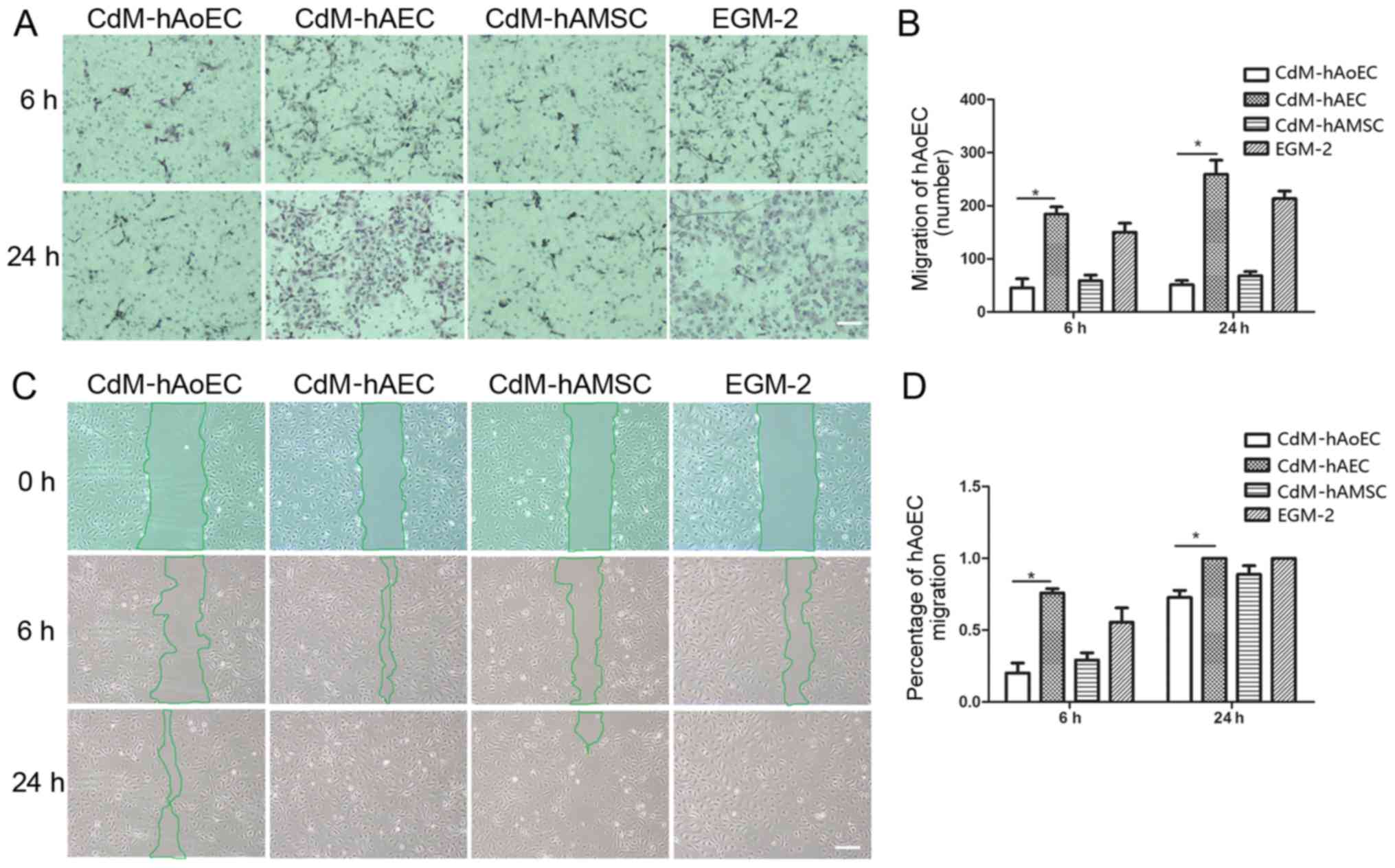
Culture medium from hAECs enhances
migration ability
In order to examine whether CdMs exhibited
biological effects relevant to hAoEC migration, we compared the
effects of the different CdMs on migration by means of scratch and
Transwell assays. The images showed that hAoEC migration into the
scratch wound area was accelerated when cultured with CdM-hAEC
(75.86±3.06% CdM-hAEC vs. 29.16±5.12% CdM-hAMSC; P<0.05,
20.11±7.04% CdM-hAoEC; n=3/group) (Fig. 3C). In Transwell cell migration
assays, our results revealed that CdM-hAEC significantly increased
the rate of hAoEC migration compared with CdM-hAMSC and CdM-hAoEC
(184.01±33.66 CdM-hAEC vs. 58.82±23.99 CdM-hAMSC; P<0.05,
45.20±30.04 CdM-hAoEC; n=3/group) (Fig. 3A).
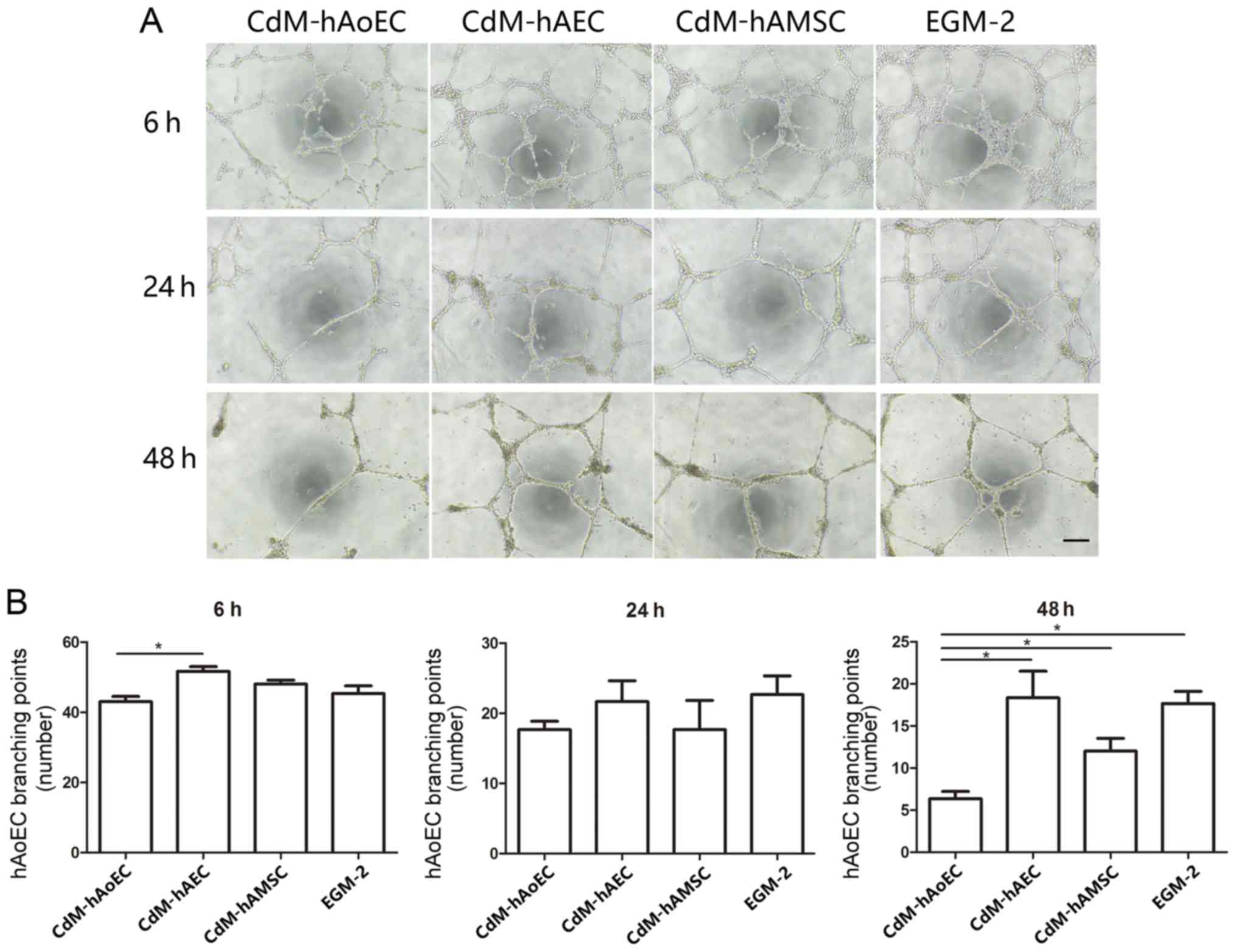
Angiogenesis assay: network
formation
The Matrigel assay is a commonly used method to
evaluate network formation by endothelial cells and was applied to
investigate whether induced CdM were also involved in forming
networks. After 6 h, the addition of CdM-hAEC to hAoECs supported
the formation of network-like structures in the Matrigel assay, to
a greater extent than CdM-hAoEC. After 24 h, the addition of
CdM-hAEC to hAoECs still supported the formation of network-like
structures, but this was not statistically significant. While
networks formed by endothelial cells in CdM-hAoEC had disintegrated
after 48 h, networks formed by hAoECs cultured with CdM-hAEC and
CdM-hAMSC were still stable (Fig.
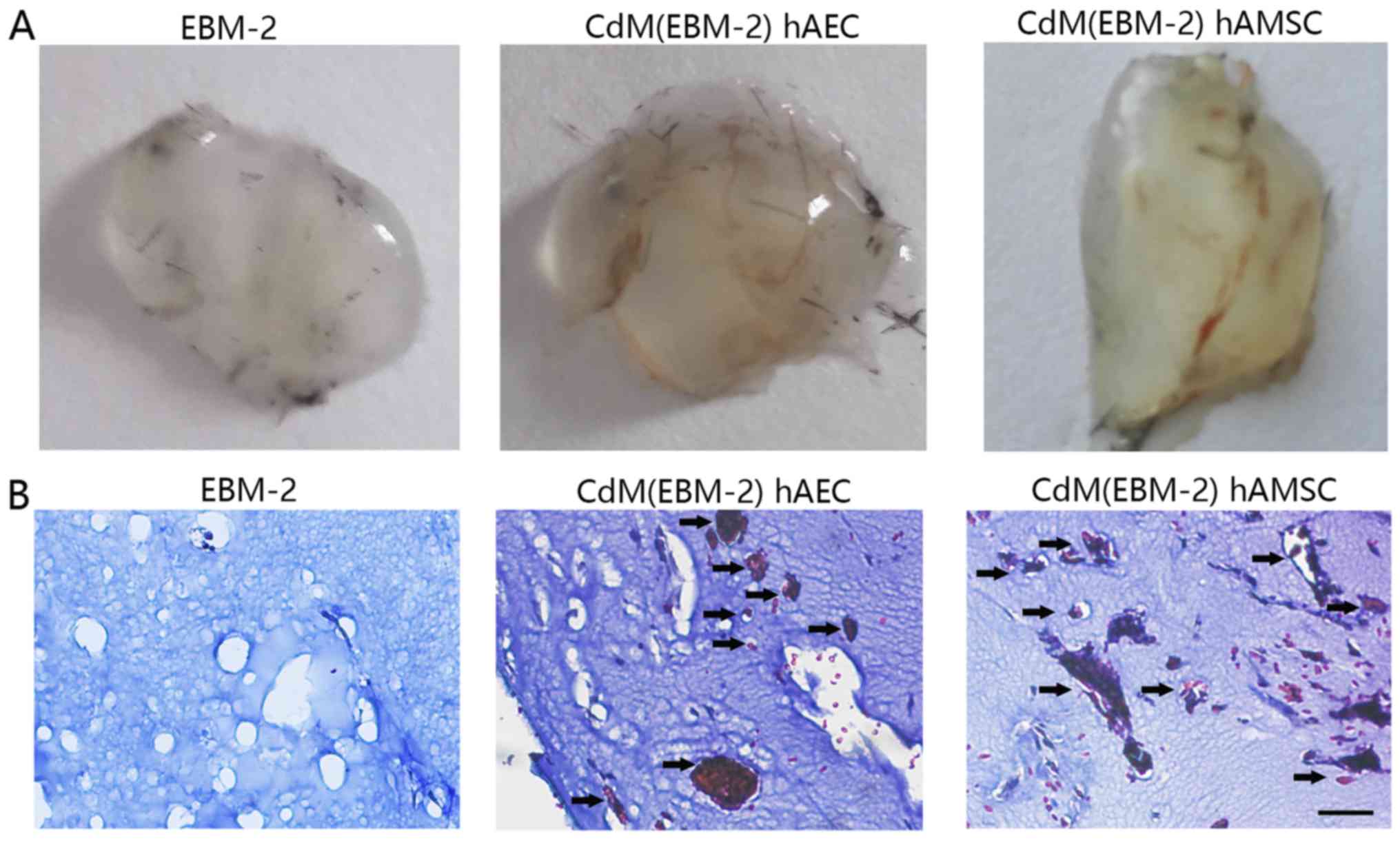
4). To examine the angiogenic potential of CdM in vivo,
we used the murine Matrigel plug assay. At 1 week after
implantation, the Matrigel plug containing CdM-hAEC and CdM-hAMSC
formed a blood vessel network connected with the host vasculature.
These vessels contained blood. while there was no blood vessel in
the negative group (Fig. 5).
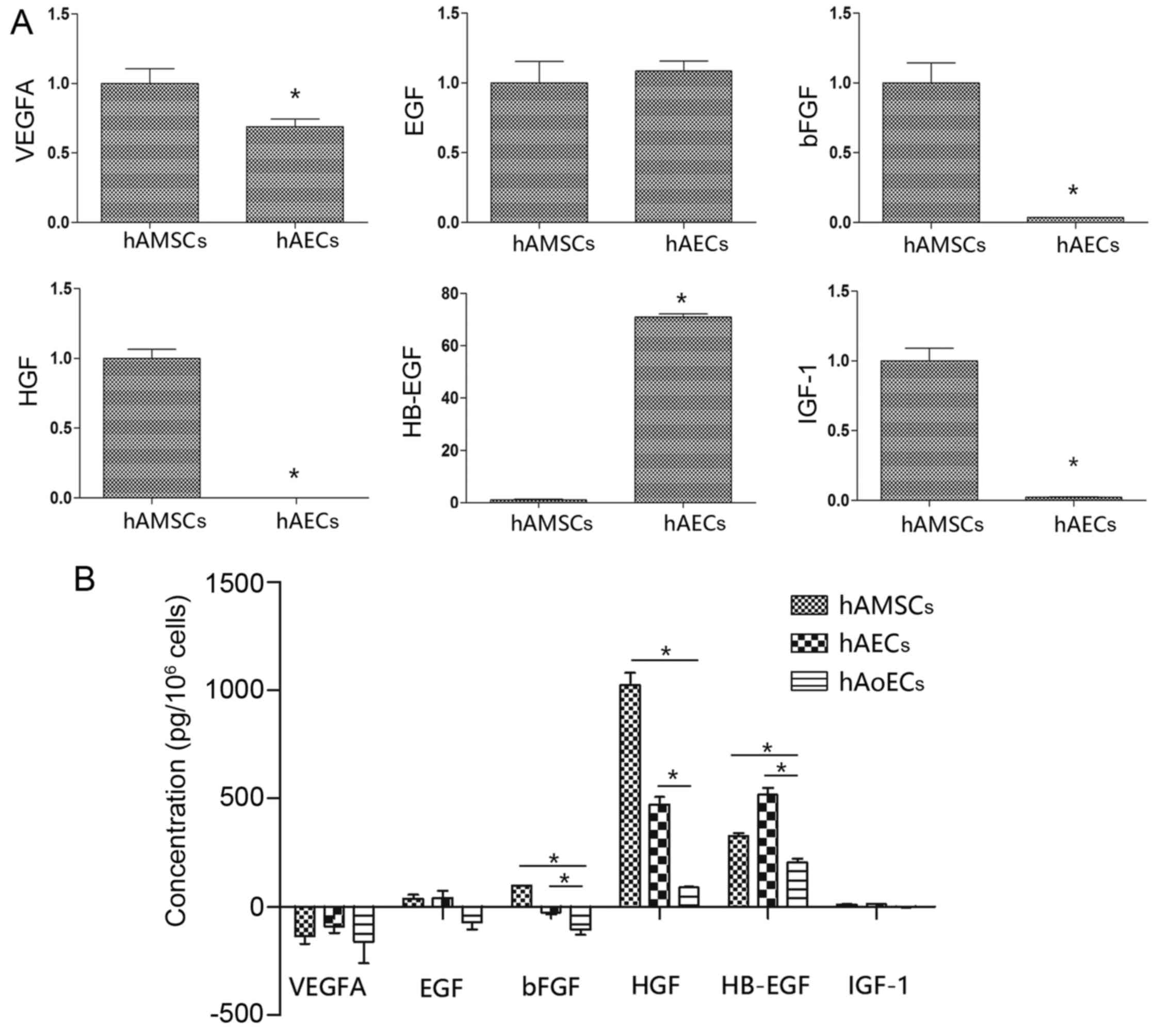
Expression of angiogenesis-specific mRNAs
in hAMSCs and hAECs
We used qRT-PCR to investigate the
angiogenic-related mRNA expression in the hAMSCs and hAECs. hAECs
showed significantly higher expression of the angiogenic gene
HB-EGF, which was 71-fold higher than the expression in the hAMSCs.
Notably, hAMSCs had a higher expression of bFGF, HGF and IGF-1
compared to these levels in the hAECs (>30-fold higher). In
addition, EGF and VEGFA, angiogenic factors that are pivotal in
neovascularization, were expressed to a similar extent in the hAECs
and hAMSCs (Fig. 6A).
Expression of angiogenic proteins in
CdMs
Collected CdMs were analyzed for the presence of
angiogenic proteins using an ELISA kit (Fig. 6B). The EGM-2 value was taken as a
background level. Compared with CdM collected from hAoECs (HB-EGF,
205.2±25.3 pg/106 cells; EGF, −72.9±43.26
pg/106 cells; bFGF, −106.4±33.3 pg/106 cells;
HGF, 89.9±2.8 pg/106 cells), amniotic cells secreted
higher levels of angiogenic factors. In line with the different
mRNA expression levels, compared with CdM-hAMSC (HB-EGF, 326.8±25.4
pg/106 cells; bFGF, 97.0±2.8 pg/106 cells;
HGF, 1024.5±98.3 pg/106 cells), CdM-hAEC had a higher
level of the proangiogenic factor HB-EGF (518.0±53.5
pg/106 cells), and lower levels of bFGF (−30.0±14.4
pg/106 cells) and HGF (472.1±49.8 pg/106
cells). There was no difference between hAECs and hAMSCs in regards
to VEGFA expression (−135.1±61 and −91.1±51.9 pg/106
cells, respectively) or EGF expression (38.3±29.7 and 41.5±57.9
pg/106 cells, respectively). Notably, although there was
an obvious difference in IGF-1 mRNA levels, there was no difference
in IGF-1 protein levels (10.5±3.8 and 12.8±2.1 pg/106
cells).
Discussion
Many studies have previously demonstrated the
therapeutic potential of stem cells using animal models including
wound healing (4,5), limb ischemia (2,3),
and myocardial infarction (6,7).
The proliferation, migration and angiogenic properties of
endothelial cells are important in the revascularization of
ischemic tissue and the reperfusion of myocardial infarction. While
recent studies have revealed the angiogenic properties of human
amniotic membrane and mesenchymal stem cells (16,21), information concerning hAECs is
rare, and comparative studies of the biological effects between
hAECs and hAMSCs are lacking. Therefore, in the present study, we
reported that amniotic cells possess high biological potential for
endothelial cells and we compared the differences in the cellular
function and biological properties between these cells. The main
findings of this study were: i) hAECs and hAMSCs display similar
growth kinetics, and express stem cell markers; ii) CdM-hAEC
significantly promoted endothelial cell migration, CdM-hAMSC
promoted endothelial cell proliferation, and they both promoted the
stabilization of angiogenesis; iii) there was high expression of
HB-EGF in hAECs; and iv) high expression of bFGF and HGF in hAMSCs.
To our knowledge, this is the first study to compare the beneficial
effects of hAECs and hAMSCs on endothelial cell function.
We are convinced that human amniotic cells are an
attractive source for cell therapy because they are free from
ethical concern, a large number of cells can be obtained, and they
display low immunogenicity, consistent with a previous study
(22). We can also obtain a large
number of cells from a small piece of amniotic membrane, thus
ensuring that there are abundant cells available for future
clinical therapy. hAECs and hAMSCs displayed similar growth
kinetics when cultured with EGM-2; and when we collected CdMs, the
number of hAECs and hAMSCs were found to be similar, thus making
the comparison of growth factors in CdMs more reasonable. After
xenogeneic transplantation into neonatal swine and rats, hAMSCs
engraft without immunosuppression (23–25). In accordance with these studies,
amniotic cells were negative for HLA-DR, which indicated that they
have low immunogenicity, which can be taken as an advantage for
in vivo therapy. OCT-3/4, SOX-2 and SSEA-4 are pluripotent
markers that are commonly expressed by stem cells (26). Consistent with previous studies
(19,26), hAMSCs and hAECs also
differentiated toward mesodermal lineages (osteogenic,
chondrogenic, and adipogenic), and expressed stem cell markers,
SSEA-4, SOX-2 and OCT-4, which indicates they have a high ability
of pluripotency and self-renewal.
In vivo studies demonstrated that there were
few differentiated cells being tested and the tissue concentrations
of growth factors were significantly increased in injured areas
treated with stem cells in transplanted models (10,27). We believe that the ability of
amniotic cells to stimulate regenerative effects is mainly induced
via paracrine routes. Furthermore, this theory is confirmed by
several studies, which showed that conditioned medium from MSCs
promoted the recovery of myocardial infarction (6,7,12).
Therefore, we collected CdM from amniotic cells, and tested the
biological effect of migration, proliferation and angiogenesis on
hAoECs. The collection time of 48 h was chosen based on the
literature (15). As an in
vitro assay of cell migration, we performed scratch and
Transwell experiments. CdM-hAEC markedly affected hAoEC migration
compared with CdM-hAMSC and CdM-hAoEC. These results are consistent
with in vivo research showing that hAECs promote
epithelialization and wound healing (4). CdM-hAMSC had a positive effect on
hAoECs, as shown by enhanced viability and proliferation ability in
the cell cycle distribution assay. CdMs from hAECs and hAMSCs
stabilized blood vessel network formation in vitro and
stimulated blood vessel formation in vivo. These results are
consistent with research showing that MSCs from bone marrow promote
angiogenesis and support blood vessel formation (3).
It has been shown that stem cells secrete a broad
variety of cytokines, chemokines, and growth factors, which may
potentially be involved in regenerative medicine (3,6,28).
The molecular processes leading to angiogenesis involve mediators
such as EGF, HB-EGF, VEGF, bFGF, HGF, IGF-1 and others, which lead
to cell migration, proliferation, vessel formation and maturation
(3,16–18,29–32). The present study demonstrates that
numerous arteriogenic cytokines are released by MSCs (11). This study demonstrated that hAECs
highly express HB-EGF; HAMSCs secreted significantly larger amounts
of HGF and bFGF, and gene results were confirmed using ELISA
assays. The difference in expression of IGF-1 mRNA was not
immediately obvious when tested by ELISA assay, perhaps due to
epigenetic regulation. We assume that these differences in the
cytokine expression profile could reflect the angiogenic and
cytoprotective properties of amniotic cells, as we observed
differences in their effects on hAoECs in our conditioned-medium
analysis.
In the present study, considering that amniotic
cells play a role in the microenvironment of endothelial cells, and
that EGM-2 functions in a similar manner, we chose EGM-2 for
conditioned medium collection to coordinate culture medium between
different cells. Cytokines not only have individual effects, but
one cytokine may potentiate (or inhibit) the effect of another,
i.e. having a synergistic or antagonistic relationship (33). Determining the nature and
mechanism(s) of the paracrine soluble molecules involved in
CdM-mediated angiogenesis stabilization is obviously a challenge
for all researchers in the field.
In conclusion, this study demonstrated the
differential effects of amniotic cells on the function of hAoECs,
via paracrine angiogenetic-related growth factors. Therefore, both
cell types may provide a convenient source for clinical
therapy.
Acknowledgments
This study was supported by the grants from the
National Basic Research Program of China (grant no. 2012CB518103),
the National Natural Science Foundation of China (grant no.
81450017), the Science and Technology Department of Liaoning
Province (grant no. 2013020200-206), the Science and Technology
Department of Liaoning Province (grant no. 2014305012), the Science
and Technology Bureau of Shenyang City (grant nos. F15-157-1-00),
the Science and Technology Planning Project of Shenyang (grant no.
F14-201-4-00).
References
|
1
|
Ilancheran S, Michalska A, Peh G, Wallace
EM, Pera M and Manuelpillai U: Stem cells derived from human fetal
membranes display multilineage differentiation potential. Biol
Reprod. 77:577–588. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim SW, Zhang HZ, Kim CE, An HS, Kim JM
and Kim MH: Amniotic mesenchymal stem cells have robust angiogenic
properties and are effective in treating hindlimb ischaemia.
Cardiovasc Res. 93:525–534. 2012. View Article : Google Scholar
|
|
3
|
Kinnaird T, Stabile E, Burnett MS, Lee CW,
Barr S, Fuchs S and Epstein SE: Marrow-derived stromal cells
express genes encoding a broad spectrum of arteriogenic cytokines
and promote in vitro and in vivo arteriogenesis through paracrine
mechanisms. Circ Res. 94:678–685. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yoshida Y, Tanaka S, Umemori H, Minowa O,
Usui M, Ikematsu N, Hosoda E, Imamura T, Kuno J, Yamashita T, et
al: Negative regulation of BMP/Smad signaling by Tob in
osteoblasts. Cell. 103:1085–1097. 2000. View Article : Google Scholar
|
|
5
|
Liu X, Wang Z, Wang R, Zhao F, Shi P,
Jiang Y and Pang X: Direct comparison of the potency of human
mesenchymal stem cells derived from amnion tissue, bone marrow and
adipose tissue at inducing dermal fibroblast responses to cutaneous
wounds. Int J Mol Med. 31:407–415. 2013.
|
|
6
|
Fidelis-de-Oliveira P, Werneck-de-Castro
JPS, Pinho-Ribeiro V, Shalom BC, Nascimento-Silva JH, Costa e Souza
RH, Cruz IS, Rangel RR, Goldenberg RC and Campos-de-Carvalho AC:
Soluble factors from multipotent mesenchymal stromal cells have
antinecrotic effect on cardiomyocytes in vitro and improve cardiac
function in infarcted rat hearts. Cell Transplant. 21:1011–1021.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Timmers L, Lim SK, Hoefer IE, Arslan F,
Lai RC, van Oorschot AA, Goumans MJ, Strijder C, Sze SK, Choo A, et
al: Human mesenchymal stem cell-conditioned medium improves cardiac
function following myocardial infarction. Stem Cell Res (Amst).
6:206–214. 2011. View Article : Google Scholar
|
|
8
|
Balsam LB, Wagers AJ, Christensen JL,
Kofidis T, Weissman IL and Robbins RC: Haematopoietic stem cells
adopt mature haematopoietic fates in ischaemic myocardium. Nature.
428:668–673. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nygren JM, Jovinge S, Breitbach M, Säwén
P, Röll W, Hescheler J, Taneera J, Fleischmann BK and Jacobsen SE:
Bone marrow-derived hematopoietic cells generate cardiomyocytes at
a low frequency through cell fusion, but not transdifferentiation.
Nat Med. 10:494–501. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kinnaird T, Stabile E, Burnett MS, Shou M,
Lee CW, Barr S, Fuchs S and Epstein SE: Local delivery of
marrow-derived stromal cells augments collateral perfusion through
paracrine mechanisms. Circulation. 109:1543–1549. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Konala VBR, Mamidi MK, Bhonde R, Das AK,
Pochampally R and Pal R: The current landscape of the mesenchymal
stromal cell secretome: A new paradigm for cell-free regeneration.
Cytotherapy. 18:13–24. 2016. View Article : Google Scholar :
|
|
12
|
Yamaguchi S, Shibata R, Yamamoto N,
Nishikawa M, Hibi H, Tanigawa T, Ueda M, Murohara T and Yamamoto A:
Dental pulp-derived stem cell conditioned medium reduces cardiac
injury following ischemia-reperfusion. Sci Rep. 5:162952015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
König J, Huppertz B, Desoye G, Parolini O,
Fröhlich JD, Weiss G, Dohr G, Sedlmayr P and Lang I: Amnion-derived
mesenchymal stromal cells show angiogenic properties but resist
differentiation into mature endothelial cells. Stem Cells Dev.
21:1309–1320. 2012. View Article : Google Scholar
|
|
14
|
Sha X, Liu Z, Song L, Wang Z and Liang X:
Human amniotic epithelial cell niche enhances the functional
properties of human corneal endothelial cells via inhibiting P53
-survivin-mitochondria axis. Exp Eye Res. 116:36–46. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
König J, Weiss G, Rossi D, Wankhammer K,
Reinisch A, Kinzer M, Huppertz B, Pfeiffer D, Parolini O and Lang
I: Placental mesenchymal stromal cells derived from blood vessels
or avascular tissues: What is the better choice to support
endothelial cell function? Stem Cells Dev. 24:115–131. 2015.
View Article : Google Scholar :
|
|
16
|
Yamahara K, Harada K, Ohshima M, Ishikane
S, Ohnishi S, Tsuda H, Otani K, Taguchi A, Soma T, Ogawa H, et al:
Comparison of angiogenic, cytoprotective, and immunosuppressive
properties of human amnion- and chorion-derived mesenchymal stem
cells. PLoS One. 9:e883192014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yotsumoto F, Tokunaga E, Oki E, Maehara Y,
Yamada H, Nakajima K, Nam SO, Miyata K, Koyanagi M, Doi K, et al:
Molecular hierarchy of heparin-binding EGF-like growth
factor-regulated angiogenesis in triple-negative breast cancer. Mol
Cancer Res. 11:506–517. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim SW, Zhang HZ, Guo L, Kim JM and Kim
MH: Amniotic mesenchymal stem cells enhance wound healing in
diabetic NOD/SCID mice through high angiogenic and engraftment
capabilities. PLoS One. 7:e411052012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Miki T, Lehmann T, Cai H, Stolz DB and
Strom SC: Stem cell characteristics of amniotic epithelial cells.
Stem Cells. 23:1549–1559. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Parolini O, Alviano F, Bagnara GP, Bilic
G, Bühring HJ, Evangelista M, Hennerbichler S, Liu B, Magatti M,
Mao N, et al: Concise review: Isolation and characterization of
cells from human term placenta: Outcome of the first international
Workshop on Placenta Derived Stem Cells. Stem Cells. 26:300–311.
2008. View Article : Google Scholar
|
|
21
|
Grzywocz Z, Pius-Sadowska E, Klos P,
Gryzik M, Wasilewska D, Aleksandrowicz B, Dworczynska M, Sabalinska
S, Hoser G, Machalinski B, et al: Growth factors and their
receptors derived from human amniotic cells in vitro. Folia
Histochem Cytobiol. 52:163–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alviano F, Fossati V, Marchionni C,
Arpinati M, Bonsi L, Franchina M, Lanzoni G, Cantoni S, Cavallini
C, Bianchi F, et al: Term Amniotic membrane is a high throughput
source for multipotent Mesenchymal Stem Cells with the ability to
differentiate into endothelial cells in vitro. BMC Dev Biol.
7:112007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Manochantr S, U-pratya Y, Kheolamai P,
Rojphisan S, Chayosumrit M, Tantrawatpan C, Supokawej A and
Issaragrisil S: Immunosuppressive properties of mesenchymal stromal
cells derived from amnion, placenta, Wharton's jelly and umbilical
cord. Intern Med J. 43:430–439. 2013. View Article : Google Scholar
|
|
24
|
Magatti M, De Munari S, Vertua E, Gibelli
L, Wengler GS and Parolini O: Human amnion mesenchyme harbors cells
with allogeneic T-cell suppression and stimulation capabilities.
Stem Cells. 26:182–192. 2008. View Article : Google Scholar
|
|
25
|
Magatti M, De Munari S, Vertua E, Nassauto
C, Albertini A, Wengler GS and Parolini O: Amniotic mesenchymal
tissue cells inhibit dendritic cell differentiation of peripheral
blood and amnion resident monocytes. Cell Transplant. 18:899–914.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fatimah SS, Tan GC, Chua K, Fariha MMN,
Tan AE and Hayati AR: Stemness and angiogenic gene expression
changes of serial-passage human amnion mesenchymal cells. Microvasc
Res. 86:21–29. 2013. View Article : Google Scholar
|
|
27
|
Gnecchi M, He H, Liang OD, Melo LG,
Morello F, Mu H, Noiseux N, Zhang L, Pratt RE, Ingwall JS, et al:
Paracrine action accounts for marked protection of ischemic heart
by Akt-modified mesenchymal stem cells. Nat Med. 11:367–368. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Caplan AI and Dennis JE: Mesenchymal stem
cells as trophic mediators. J Cell Biochem. 98:1076–1084. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamakawa H, Muraoka N, Miyamoto K,
Sadahiro T, Isomi M, Haginiwa S, Kojima H, Umei T, Akiyama M,
Kuishi Y, et al: Fibroblast growth factors and vascular endothelial
growth factor promote cardiac reprogramming under defined
conditions. Stem Cell Reports. 5:1128–1142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen QH, Liu AR, Qiu HB and Yang Y:
Interaction between mesenchymal stem cells and endothelial cells
restores endothelial permeability via paracrine hepatocyte growth
factor in vitro. Stem Cell Res Ther. 6:442015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barrientos S, Stojadinovic O, Golinko MS,
Brem H and Tomic-Canic M: Growth factors and cytokines in wound
healing. Wound Repair Regen. 16:585–601. 2008. View Article : Google Scholar
|
|
32
|
Chen L, Tredget EE, Wu PY and Wu Y:
Paracrine factors of mesenchymal stem cells recruit macrophages and
endothelial lineage cells and enhance wound healing. PLoS One.
3:e18862008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang Y, Chen QH, Liu AR, Xu XP, Han JB and
Qiu HB: Synergism of MSC-secreted HGF and VEGF in stabilising
endothelial barrier function upon lipopolysaccharide stimulation
via the Rac1 pathway. Stem Cell Res Ther. 6:2502015. View Article : Google Scholar : PubMed/NCBI
|