Introduction
Liver transplantation has become the most effective
treatment for end-stage liver diseases. However, a shortage of
donor organs remains the major impediment to the further
development of liver transplantation. Reduced-size liver
transplantation (RSLT), either living-related liver transplantation
or split-liver transplantation, can meet the shortage of livers to
some extent (1) and has been
applied widely in clinics. However, ischemia-reperfusion injury
weakens the regeneration of the remnant liver after RSLT (2,3).
In addition, the risk of primary graft dysfunction caused by
microvascular dysfunction and immune-mediated allograft rejection
also increases following RSLT (4). Therefore, it is necessary to find
treatments that inhibit the death and stimulate the regeneration of
hepatocytes, alleviate the rejection of the transplanted liver and
induce immune tolerance.
Bone marrow-derived mesenchymal stem cells (BM-MSCs)
are pluripotent, can differentiate into endothelial cells for
endothelial repair (5,6), secrete a variety of cytokines,
chemokines and growth factors promoting cell proliferation and
differentiation and can migrate to the damaged tissue to repair it
(7). In addition, because of
their low cell surface expression of major histocompatibility
complex molecules, BM-MSCs show immunomodulatory activity, which
plays an important role in allograft rejection, and can induce
immune tolerance and graft regeneration (8,9).
However, following transfusion, BM-MSCs present at a low
concentration and short duration of activity in the target tissues,
and thus, are not widely applied. Heme oxygenase-1 (HO-1) is the
rate-limiting enzyme of heme catalyzation, whose activity can be
induced by oxidative stress. HO-1 has antioxidant and homeostasis
maintenance effects (10). HO-1
can increase the activity and prolong the duration of BM-MSCs
(11). Previous studies of the
authors have demonstrated that HO-1-transduced BM-MSCs
(HO-1/BM-MSCs) may protect the transplanted liver by participating
in the regulation of transplantation immunity and repair of the
damaged liver tissue (12,13).
However, the mechanism of this protective effect remains
unknown.
Autophagy is an important lysosomal
pathway-dependent biological process in eukaryotes, which degrades
cytoplasmic components to maintain cell homeostasis and provide
substrates for energy metabolism (14). Autophagy plays an important role
in a variety of liver functions. Firstly, as the liver has a unique
regenerative ability, autophagy can clear the damaged cell
organelles, and oxidized or accumulated proteins, during liver
regeneration. Second, autophagy participates in and regulates
metabolic pathways of proteins, carbohydrates and lipids in the
liver. Furthermore, autophagy may have metabolic or proliferative
effects during rapid regeneration of hepatocytes, although this
requires further study (14).
Studies have demonstrated that, as a cytoprotective mechanism,
autophagy could inhibit oxidative stress, reduce the amount of
reactive oxygen species (ROS) generated by Kupffer cells and
inhibit the death of hepatocytes (14–17). Autophagy also participates in the
positive and negative selection of CD4+ T cells, and
plays an important role in the central and peripheral immune
tolerance to self-antigens (18).
Is autophagy involved in the protection of HO-1/BM-MSCs on RSLT?
The aim of the present study was to determine the participation of
autophagy in the protective effects of HO-1/BM-MSCs on RSLT and to
explore its possible mechanism.
Materials and methods
Animals and ethics
Specific-pathogen-free healthy adult inbred male
Brown-Norway (BN) rats (n=45) and male Lewis rats (n=35) were
purchased from the Vital River Laboratories Animal Technology, Co.,
Ltd. (Beijing, China). The rats were housed individually in
standard animal facilities at 18–26°C with a 12-h light/dark cycle
and were provided with commercially available chow and tap water
ad libitum. BM-MSCs were extracted from syngeneic inbred
male BN rats (80–100 g; 4–5 weeks old). Inbred male Lewis rats
(210–250 g; 8–10 weeks old) were the liver transplantation donors,
and inbred male BN rats (210–250 g; 8–10 weeks old) were the
recipients. The difference in body weight between the donor and the
recipient of each pair did not exceed 10 g. Food was withheld from
the recipient animals for 12 h before surgery, but was not withheld
from the donor animals. The study protocol was approved by the
Animal Care and Research Committee of Tianjin First Central
Hospital (Tianjin, China). All surgeries and sacrifices were
performed under 5% chloral hydrate anesthesia (0.5 ml/100 g). Every
effort was made to minimize animal suffering.
Isolation, characterization and
differentiation induction of BM-MSCs
BM-MSCs were isolated aseptically from the femur and
tibia of 10 syngeneic male BN rats following sacrifice by cervical
dislocation. After cutting off both ends of the epiphyseal, the
marrow cavity was rinsed by DMEM/F12 (Gibco, Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (FBS). Red blood cells were lysed using 0.1 mol/l
NH4Cl, and the remaining cells were washed, resuspended,
and cultured 1×106/T75 culture flask at 37°C with 5%
CO2 in Dulbecco's modified Eagle's medium (DMEM)/F12
containing 100 U/ml penicillin, 100 mg/ml streptomycin and 15% FBS
(13). The well-grown
third-passage BM-MSCs were resuspended at 1×106/ml and
then labeled with antibodies against CD29 (1:80, PE, 102207), CD90
(1:200, FITC, 202503), CD45 (1:80, PE, 202207), RT1A (1:80, PE,
205208), RT1B (1:200, FITC, 205305) (both from BioLegend, Inc., San
Diego, CA, USA) and CD34 (1:5, FITC, sc-7324; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 30 min for flow
cytometric analysis (BD FACSAria III; BD Biosciences, Franklin
Lakes, NJ, USA) of the expression of cell surface markers.
Adipogenic differentiation medium was prepared as
DMEM/F12 containing 10% FBS, 200 mM indomethacin (Sigma-Aldrich,
Merck KGaA, Darmstadt, Germany), 0.5 mM 1-methyl-3-isobutylxanthine
(Sigma-Aldrich, Merck KGaA), 40 U/ml insulin (Sigma-Aldrich, Merck
KGaA) and 1 mM dexa-methasone (Sigma-Aldrich, Merck KGaA).
Following staining with Oil Red O (Beijing Dingguo Changsheng
Biotechnology Co., Ltd., Beijing, China), BM-MSCs were observed
under a light microscope to identify red lipid droplets in the
cytoplasm. Osteogenic differentiation medium was prepared as
DMEM/F12 containing 10% FBS, 1 mM dexamethasone, 1 M sodium
glycerophosphate (Sigma-Aldrich, Merck KGaA) and 100 ng/ml vitamin
C (Sigma-Aldrich, Merck KGaA), with a pH value adjusted to 7.3–7.4.
The medium was changed every 72 h. Following staining with the von
Kossa cell staining kit (Shanghai Genmed Gene Pharmaceutical
Technology Co., Ltd., Shanghai, China), BM-MSCs were observed under
a light microscope to identify black calcium salt in the
cytoplasm.
Transduction of BM-MSCs with HO-1-bearing
recombinant adenovirus
When BM-MSCs were completely adherent, HO-1-bearing
recombinant adenovirus (Shanghai GeneChem Co., Ltd., Shanghai,
China) at a multiplicity of infection of 10 were added into the
flask to transduce Adv/HO-1/BM-MSCs; the reaction was carried out
in the dark. Adenoviruses expressing the green fluorescence protein
(GFP) were also added into the flask to transduce Adv/GFP/BM-MSCs
to verify the expression of the target gene. After culturing for
6–8 h, the supernatant was discarded and replaced with complete
culture medium DMEM/F12 containing 10% FBS for continued
cultivation. The cells were observed under a fluorescence
microscope (Olympus IX71; Olympus Corp., Tokyo, Japan) to evaluate
the expression of GFP fluorescence and the morphology of nucleus at
room temperature, and then were photographed using Image-Pro Plus
6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Establishment of a rejection model in 50%
RSLT of rats and the experimental protocol
A 50% RSLT rejection model was established with
Lewis donor rats and BN recipient rats, as described by Zhao et
al (19). The donor livers
were perfused via the portal vein (PV) with 4°C lactated Ringer's
solution containing heparin sodium (50 U/ml). The harvested graft
was preserved in a bath of lactated Ringer's solution at 4°C. The
cuff was slipped over the PV using microforceps, and the distal end
of the vein was everted over the cuff and secured with a
circumferential 5–0 silk ligature. The same procedure was performed
for the infrahepatic vena cava (IHVC) cuff preparation. After the
recipient liver was removed, the donor liver was placed
orthotopically in the abdominal cavity of the recipient. The
suprahepatic vena cava (SHVC) was anastomosed end-to-end using a
continuous 7–0 nylon suture. A cuff anastomosis of the PV and IHVC
was then performed. The graft was reperfused by opening the PV,
IHVC and SHVC in turn. The bile duct was connected by telescoping a
tube in the bile duct of the donor into that of the recipient.
Experimental animals were divided into three groups, which received
normal saline (NS) (group 1), BM-MSCs (group 2) or HO-1/BM-MSCs
(group 3). HO-1/BM-MSCs or BM-MSCs were injected through the
superficial dorsal veins immediately following the surgery. Rats of
the cell-treated group received 5×106/ml (1 ml), while
the control groups were given the equivalent volume of NS. Five
animals per time-point were euthanized on post-operative day (POD)
0, 1, 3, 5, 7, 10 or 14 for further analysis of the transplanted
liver.
Histopathological analysis
After fixation in 10% formalin at 37°C for at least
48 h, recipients' hepatic tissues were embedded in paraffin, cut
into 5 µm thick sections, and stained with hematoxylin and eosin
(H&E). Pathological changes and the extent of rejection were
evaluated under a light microscope. Acute cellular rejection was
classified according to the Banff criteria (20).
Apoptosis detection
Paraffin sections of liver tissue were routinely
dewaxed, dehydrated and digested by proteinase K (Promega Corp.,
Madison, WI, USA). Terminal deoxynucleotidyl transferase dUTP nick
end labeling (TUNEL) reaction mixture (Promega) was added dropwise
and incubated at 37°C for 1 h. The average number of apoptotic
cells was analyzed in 10 randomly selected visual fields using
fluorescence microscopy (magnification, ×200; Nikon Ni-U; Nikon
Corp., Tokyo, Japan), and the mean number of apoptotic cells was
measured by staining cell nuclei wih DAPI (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China) to determine the total
number of cells.
Electron microscope analysis
Transplanted liver tissues were obtained from the
recipient freshly, cut into 2 × 3 mm samples, and fixed in 2.5%
glutaraldehyde solution. Embedded sections were observed under a
transmission electron microscopy (Hitachi H-600; Hitachi, Ltd.,
Tokyo, Japan) for the ultrastructures of the transplanted liver
tissue.
Immunohistochemical analysis
Transplanted liver tissues were fixed on glass
slides, heated at 70°C for 1 h, deparaffinized by dimethylbenzene,
hydrated by gradient ethanol, subjected to antigen retrieval, and
blocked with 10% goat serum (Shanghai GeneChem; www.genechem.com.cn.html). Following incubation
with primary antibodies Beclin-1 (1:500, ab55878) and LC3 A/B
(1:500, #12741), staining by diaminobenzidine and hematoxylin, and
dehydration by gradient ethanol, the sections were covered with
neutral balsam (Shanghai Yiyang Instrument Co., Ltd., Shanghai,
China; http://yiyang17.com/index.php)
according to the manufacturer's instructions, and then observed
under a light microscope.
Detection of protein levels by western
blotting
Radioimmu-noprecipitation assay lysis buffer
(Beijing Solarbio Science & Technology) was added to frozen
liver tissues, which were then homogenized, and centrifuged at
12,000 × g for 4 min at 4°C to extract the proteins. A
bicinchoninic acid assay (Wuhan Boster Biological Technology, Ltd.,
Wuhan, China; http://www.boster.com.cn/) was used to determine the
protein content. SDS-PAGE (10%) was used for Beclin-1, ERK and
p-ERK; 8% SDS-PAGE was used for mammalian target of rapamycin
(mTOR) and p-mTOR; and 15% SDS-PAGE was used for LC3 A/B. A total
of 30 µg proteins each well in the gels were wet transferred to
nitrocellulose membranes for 2 h, blocked by 5% skimmed milk for 2
h, and then incubated at 4°C overnight with antibodies recognizing
Beclin-1 (ab55878) and GAPDH (ab8245) (both from Abcam, Cambridge,
UK), ERK (#4695) and LC3 A/B (#12741) (both from Cell Signaling
Technology, Inc., Danvers, MA, USA), p-ERK (RT1206; Hangzhou HuaAn
Biotechnology Co., Ltd., Hangzhou, China), mTOR (ab32028; Abcam)
and p-mTOR (sc-293132; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA). The antibodies were used at the following dilutions: Beclin-1
(1:500), GAPDH (1:5,000), ERK (1:1,000), p-ERK (1:250), mTOR
(1:1,000), p-mTOR (1:100) and LC3 A/B (1:1,000). The membranes were
then rinsed, incubated with secondary antibodies HRP-conjugated
anti-rabbit IgG (1:5,000, ab191866) and HRP-conjugated anti-rat
(1:2,000, ab131368) (both from Abcam) for 2 h, rinsed again, and
then visualized using the enhanced chemiluminescence system (Wuhan
Boster Biological Technology). The abundance of the target protein
was calculated relative to the abundance of the internal control
protein, GAPDH, using a gel imaging analysis system (Alpha Innotech
FluorChem FC2; Alpha Innotech Corp., San Leandro, CA, USA).
Detection of gene levels by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the liver tissue using
RNAiso Plus reagents (Takara Bio, Inc., Shiga, Japan). cDNA was
reverse transcribed using reverse transcription kits (Tiangen
Biotech Co., Ltd., Beijing, China) and 2 µl cDNA was added to 20
μl of a PCR reaction system using a fluorescence
quantitative PCR kit (Sangon Biotech Co., Ltd., Shanghai, China;
http://www.sangon.com/) for amplification.
Primers were synthesized by Sangon Biotech. The primer sequences
were as follows: ERK sense, 5′-GGCAACCGCCATTTCTCG-3′ and antisense,
5′-GCTTCGCTTCTGTTTAGCATCAC-3′; mTOR sense,
5′-TTTGGACGGTGTAGAACTTGGAG-3′ and antisense,
5′-CGAACCCTGTTAATAATCTGAATAGC-3′; and GAPDH sense,
5′-CGTATCGGACGCCTGGTTAC-3′ and antisense,
5′-GGATCTCGCTCCTGGAAGATG-3′. The reaction conditions were 95°C
pre-denaturation for 30 sec, and 40 cycles of PCR amplification
comprising 95°C for 10 sec, 58°C for 30 sec and 72°C for 30 sec.
The results were analyzed by a LightCycler® 96 real-time
fluorescence quantitative PCR detection system (Roche Diagnostics
GmbH, Basel, Switzerland) (21).
Statistical analysis
The SPSS software (version, 17.0; SPSS, Inc.,
Chicago, IL, USA) was used for statistical analysis. The GraphPad
Prism 5.0 software (GraphPad Software, Inc., La Jolla, CA, USA) was
used to plot data for presentation. Normally distributed data were
presented as means ± standard deviation. Different groups of data
were compared by the t-test or analysis of variance (ANOVA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Morphology, differentiation induction and
phenotypic analysis of BM-MSCs
Isolated primary BM-MSCs gradually became adherent
after 12 h, and impurities such as hematopoietic cells became fewer
and fewer with each medium exchange and passage. The third-passage
cells were obtained after ~15 days, and grew as long spindle-shaped
whirlpool or paralleled pattern cells, which are the typical
morphological characteristics of BM-MSCs (Fig. 1A). Meanwhile, isolated BM-MSCs
could be induced to differentiate into adipocytes and osteoblasts
in adipogenic and osteogenic differentiation medium, respectively
(Fig. 1B and C). Phenotypic
examination of the third-passage BM-MSCs surface markers by flow
cytometry demonstrated that >92% of these cells were positive
for CD29, CD90 and RT1A, and negative for CD34, CD45 and RT1B
(Fig. 1E–G). These results
suggested that, following three passages of culture, the BM-MSCs
were pure and had typical characteristics.
 | Figure 1Morphological characteristics,
differentiation induction and phenotypic identification of BM-MSCs.
(A) Bright field image of third-passage BM-MSCs following
transduction with HO-1, growing as long spindle-shapes with a
whirlpool or paralleled pattern (magnification, ×100). (B)
Adipogenic induction (magnification, ×200), the arrow presents the
orange lipid droplets in the cytoplasm after staining with Oil Red
O. (C) Osteogenic induction (magnification, ×200), the arrow
indicates the black calcium salt deposit after von Kossa staining.
(D) Fluorescent image of third-passage BM-MSCs following
transduction with HO-1, showing that ~85% of the cells are
transduced with HO-1 (emitting green fluorescence; magnification,
×100). (E) 98.3% of the cells CD29+CD34−
cells. (F) 97.6% of the cells are CD90+CD45−
cells. (G) 92.3% of the cells are RT1A+RT1B−
cells. BM-MSCs, bone marrow mesenchymal stem cells; HO-1; heme
oxygenase-1. |
Transduction of BM-MSCs with HO-1-bearing
recombinant adenovirus
BM-MSCs were transduced with HO-1-bearing
recombinant adenoviruses carrying GFP, at a multiplicity of
infection of 10. After 48 h, the GFP fluorescence emitted by
Adv-BM-MSCs was observed under a fluorescence microscope, and the
proportion of adenovirus-infected BM-MSCs emitting green
fluorescent was ~85% (Fig. 1D),
which suggested that most BM-MSCs were successfully transduced with
HO-1.
Histopathological characteristics and
rejection activity index of transplanted liver after RSLT
The recipients treated with NS reported minor damage
immediately after RSLT, which then progressed with little
inflammation periportally (mainly lymphocyte infiltration) without
significant expansion of the portal tracts, and lymphocyte
infiltration beneath venous endothelium was not obvious on POD 1.
Periportal inflammation was obvious on POD 3, primarily because of
lymphocyte infiltration, and lymphocyte infiltration beneath the
venous endothelium was still not evident. The acute rejection
increased on POD 5, with mixed inflammatory infiltration (including
lymphoblasts and eosinophils) of the portal tracts spreading to the
surrounding parenchyma; most interlobular bile ducts were
infiltrated by inflammatory cells, and subendothelial lymphocytic
infiltration involved the interlobular vein. Acute rejection
increased sharply on POD 7, with moderate to severe acute rejection
being observed on POD 7, POD 10 and POD 14. Most of the portal
areas were infiltrated with a large number of mixed lymphocytes,
which extended to the surrounding parenchyma significantly.
Interlobular bile ducts presented inflammation and luminal
disruption; subendothelial lymphocytic infiltration involving the
interlobular vein and central vein was significant and perivenular
hepatocyte necrosis was observed. The rejection was mild in
BM-MSCs-treated group on POD 0, POD 1, POD 3 and POD 5 compared
with that of the NS-treated group, with little visible lymphocyte
infiltration periportally, and cholangitis and degeneration of the
biliary epithelium was scarcely seen; interlobular and central vein
phlebitis was uncommon. However, acute rejection in the
BM-MSCs-treated group increased sharply on POD 7, but was less
severe than that of the NS-treated group. The liver deteriorated to
serious injury on POD 14, with no significant difference compared
with the NS-treated group. The rejection was mild in
HO-1/BM-MSCs-treated group on POD 0, POD 1, POD 3 and POD 5
compared with that of the NS-treated group and the BM-MSCs-treated
group. The condition of the livers deteriorated gradually after POD
7, but was less severe than that of the NS-treated group and the
BM-MSCs-treated group on POD 7, POD 10 and POD 14. These results
suggested that the rejection model after RSLT was constructed
successfully, and that the acute rejection in the
HO-1/BM-MSCs-treated group was less severe compared with that of
the BM-MSCs-treated group and the NS-treated group at different
time-points after RSLT and the duration of action was long-lasting
(Fig. 2A).
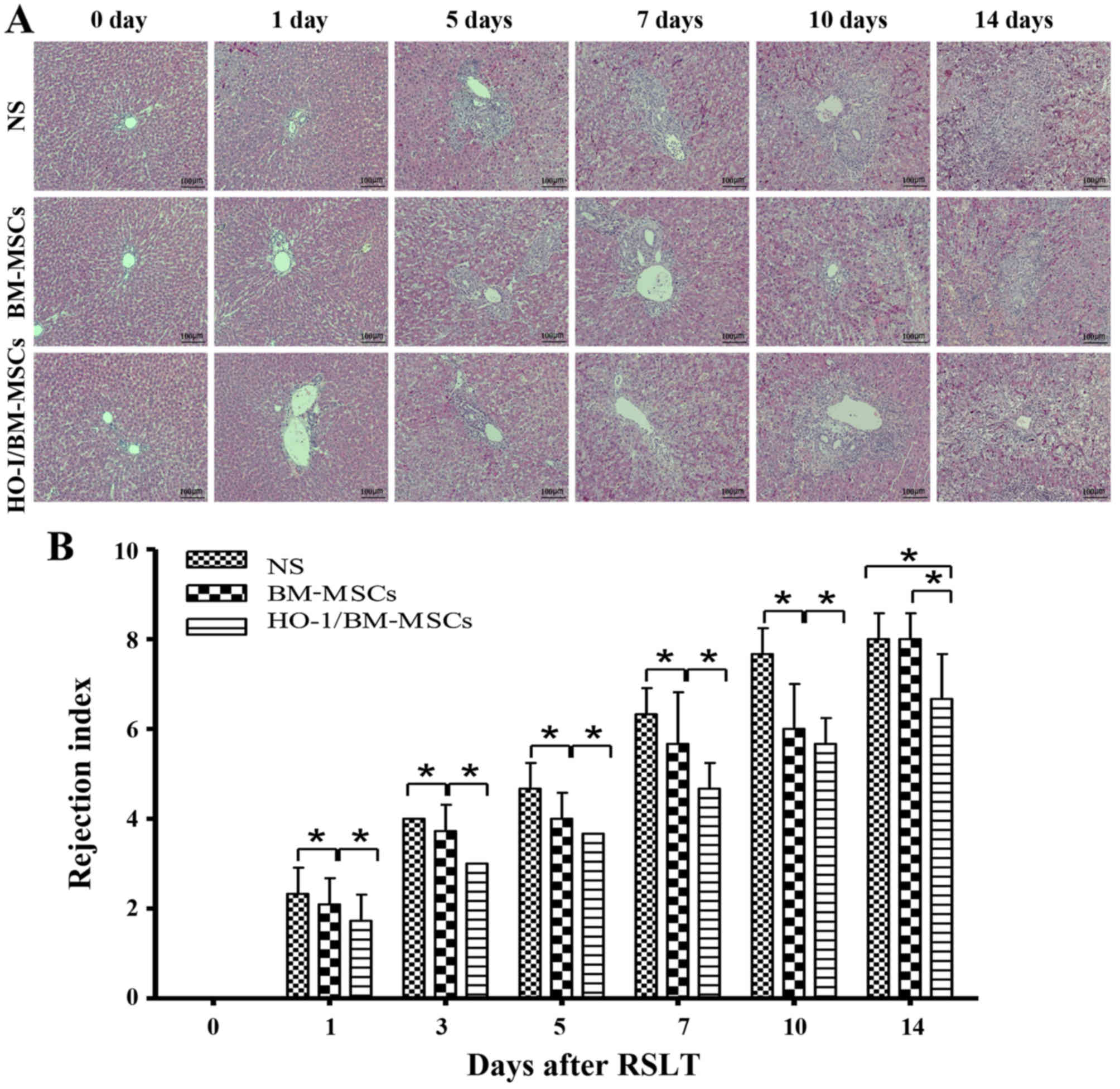 | Figure 2Histopathological characteristics and
rejection activity index of transplanted livers after RSLT
(magnification, ×100). (A) The acute rejection in the normal saline
(NS)-treated group became progressively aggravated post
transplantation. Periportal lymphocytes infiltration increased
gradually to significant levels, interlobular bile ducts showed
inflammation and subendothelial lymphocytic infiltration involving
the interlobular vein and central vein progressed significantly.
The acute rejection was moderate to severe on POD 7, POD 10 and POD
14. Portal areas were infiltrated with a large number of mixed
lymphocytes, which extended significantly to the surrounding
parenchyma, interlobular bile ducts were infiltrated with
inflammatory cells with severe biliary epithelial injury and
subendothelial lymphocytic infiltration involving the interlobular
vein and central vein was significant. The acute rejection in the
BM-MSCs-treated group also became progressively aggravated post
transplantation, however, it was less severe than that of the
NS-treated group at each time-point, except on POD 14. Little
lymphocyte infiltration was visible periportally on POD 7. The
portal areas were infiltrated with a large number of lymphocytes,
with aggravated biliary inflammation and periphlebitis on POD 14.
The acute rejection in the HO-1/BM-MSCs-treated group also became
increasingly aggravated post transplantation; however, it was less
severe than that of the NS-treated group and the BM-MSCs-treated
group. The rejection was mild at POD 7, with little lymphocyte
infiltration periportally, scarcely any biliary epithelium
degeneration and cholangitis. The acute rejection in the
HO-1/BM-MSCs-treated group deteriorated after POD 7, but was still
less severe than that of the NS-treated group and the
BM-MSCs-treated group. (B) RAI in the different groups. The RAI of
the BM-MSCs-treated group was significantly lower than that of the
NS-treated group at each time-point except on POD 14 and the RAI of
the HO-1/BM-MSCs-treated group was significantly lower than that of
the NS-treated group and the BM-MSCs-treated group at each
time-point. n=5 at each time-point for each group.
*P<0.05 as indicated. RSLT, reduced-size liver
transplantation; POD, post-operative day; BM-MSCs, bone marrow
mesenchymal stem cells; HO-1/BM-MSCs, HO-1 transduced BM-MSCs; RAI,
rejection activity index. |
When the degree of rejection was classified by the
rejection activity index (RAI) using the Banff scheme, the RAIs of
the HO-1/BM-MSCs-treated group were significantly lower than those
of the NS and BM-MSCs-treated groups at each time-point
(P<0.05). The RAIs of the BM-MSCs-treated group were
significantly lower than those of the NS-treated group at each
time-point, except on POD 14 (P<0.05). These results suggested
that HO-1/BM-MSCs treatment could reduce acute rejection injury
after RSLT, and the duration of action could extend to POD 14,
which was longer than that achieved by simple BM-MSCs treatment
(Fig. 2B).
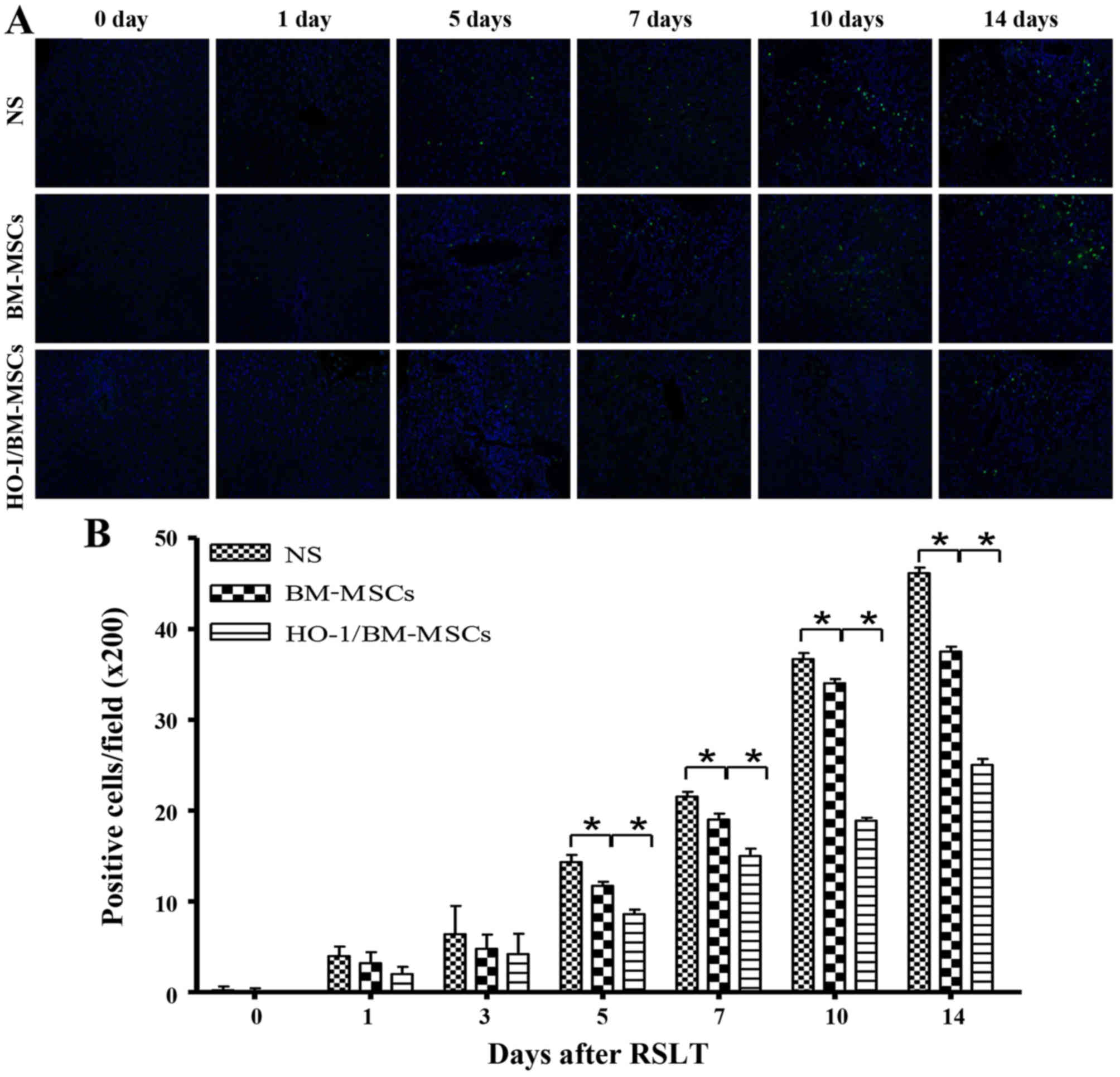
Apoptosis in transplanted liver
tissue
Apoptotic cells were scattered in all of the three
groups of transplanted livers on POD 0 and 1. The numbers of
apoptotic cells increased progressively on POD 5, POD 7, POD 10 and
POD 14, but were significantly lower in the HO-1/BM-MSCs-treated
group than in the NS and BM-MSCs-treated groups. These results
indicated that HO-1/BM-MSCs could reduce the apoptosis of
transplanted liver tissue (Fig.
3).
Ultrastructure and autophagic vacuoles of
the transplanted liver on POD 7
The autophagic vacuoles were surrounded by a bilayer
membrane, and the bilayer was parallel, with a narrow gap. The
initial autophagic vacuoles contained complete mitochondria or
endoplasmic reticulum, and the degraded autophagic vacuoles
contained degraded rough endoplasmic reticulum or mitochondria,
which could be recognized by the different inclusions. In the
NS-treated group on POD 7, the nucleus was condensed significantly,
and the endoplasmic reticulum and mitochondria showed obvious
edema, without significant autophagic vacuoles. In the
BM-MSCs-treated group, the nucleus was condensed slightly, with
visible autophagic vacuoles. By contrast, in the
HO-1/BM-MSCs-treated group, there was no nuclear condensation or
distortion, and a large number of autophagic vacuoles were
observed. These results suggested that the ultrastructural damage
of HO-1/BM-MSCs-treated livers on POD 7 was milder than that of the
NS and BM-MSCs-treated groups, and the number of initial and
degraded autophagic vacuoles in the HO-1/BM-MSCs-treated group was
higher than that in the NS and BM-MSCs-treated groups (Fig. 4).
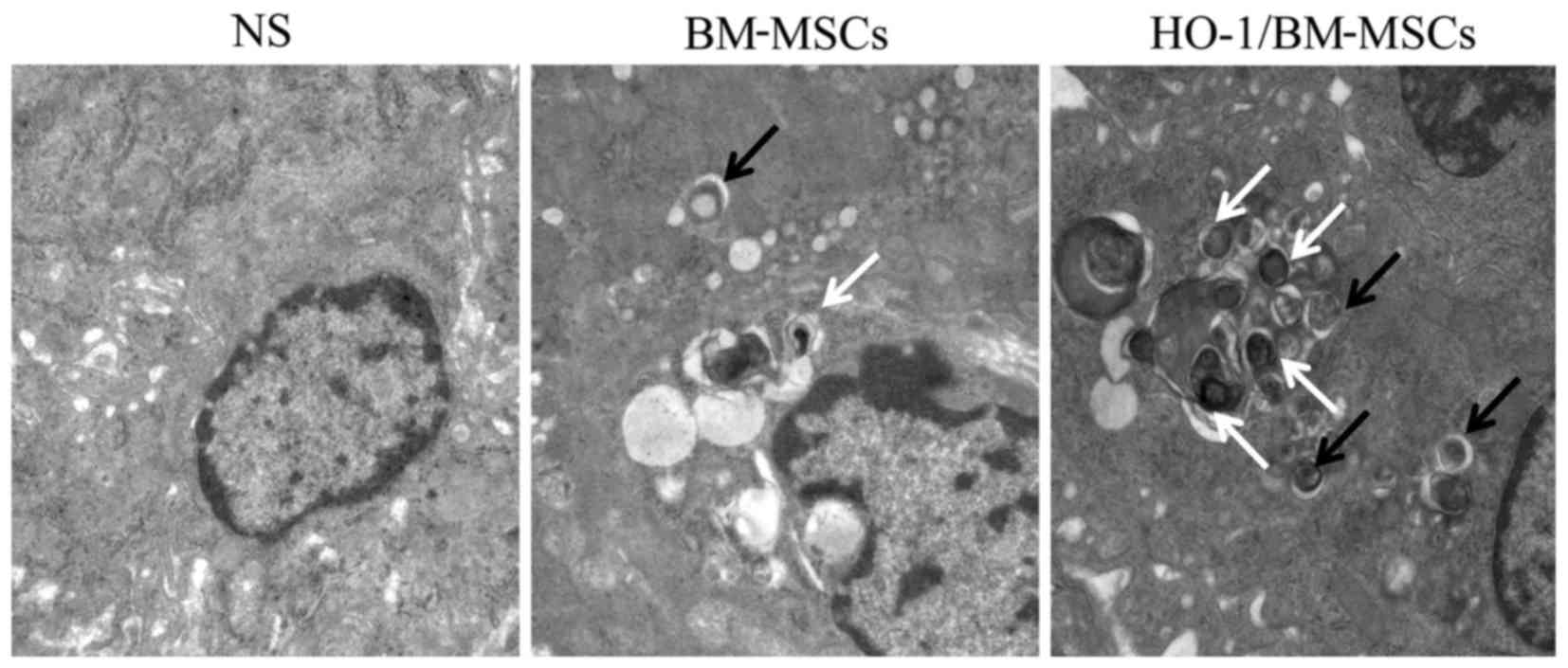 | Figure 4Ultrastructure and autophagic
vacuoles of the transplanted liver on POD 7 observed under a
transmission electron microscope (magnification, ×25,000). The
normal saline (NS)-treated group presented nuclear pycnosis, an
irregular nuclear membrane with deformation, significantly swollen
endoplasmic reticulum and mitochondria and no typical autophagic
vacuoles. The BM-MSCs-treated group showed slight nuclear pycnosis
and deformation, no obvious edema of the endoplasmic reticulum,
slight edema of the mitochondria, and visible initial and degraded
autophagic vacuoles. The HO-1/BM-MSCs-treated group reported slight
nuclear pycnosis without deformation, no edema in endoplasmic
reticulum or mitochondria, and a large number of initial autophagic
vesicles and degraded autophagic vesicles. The black arrow shows
initial autophagic vesicles, containing complete rough endoplasmic
reticulum and mitochondria. The white arrow shows degraded
autophagic vesicles, containing degraded rough endoplasmic
reticulum and mitochondria. POD, postoperative day; BM-MSCs, bone
marrow mesenchymal stem cells; HO-1/BM-MSCs, HO-1 transduced
BM-MSCs. |
Levels of autophagy-related proteins LC3
I/II and Beclin-1 after RSLT
Levels of the LC3 I/II protein
Immunohistochemical tests presented only a small
amount of LC3 I/II protein in the endothelia of the central vein in
all three groups on POD 0, with no significant difference between
the three groups. The LC3 I/II protein was visible in the cytoplasm
of both the central venous endothelia and hepatocytes on POD 1,
with the highest level of the protein in the HO-1/BM-MSCs-treated
group. The amount of LC3 I/II protein increased after POD 5 as the
time after RSLT was prolonged, and the overall level in the
HO-1/BM-MSCs-treated group was much higher than that in the NS and
the BM-MSCs-treated groups (Fig.
5A).
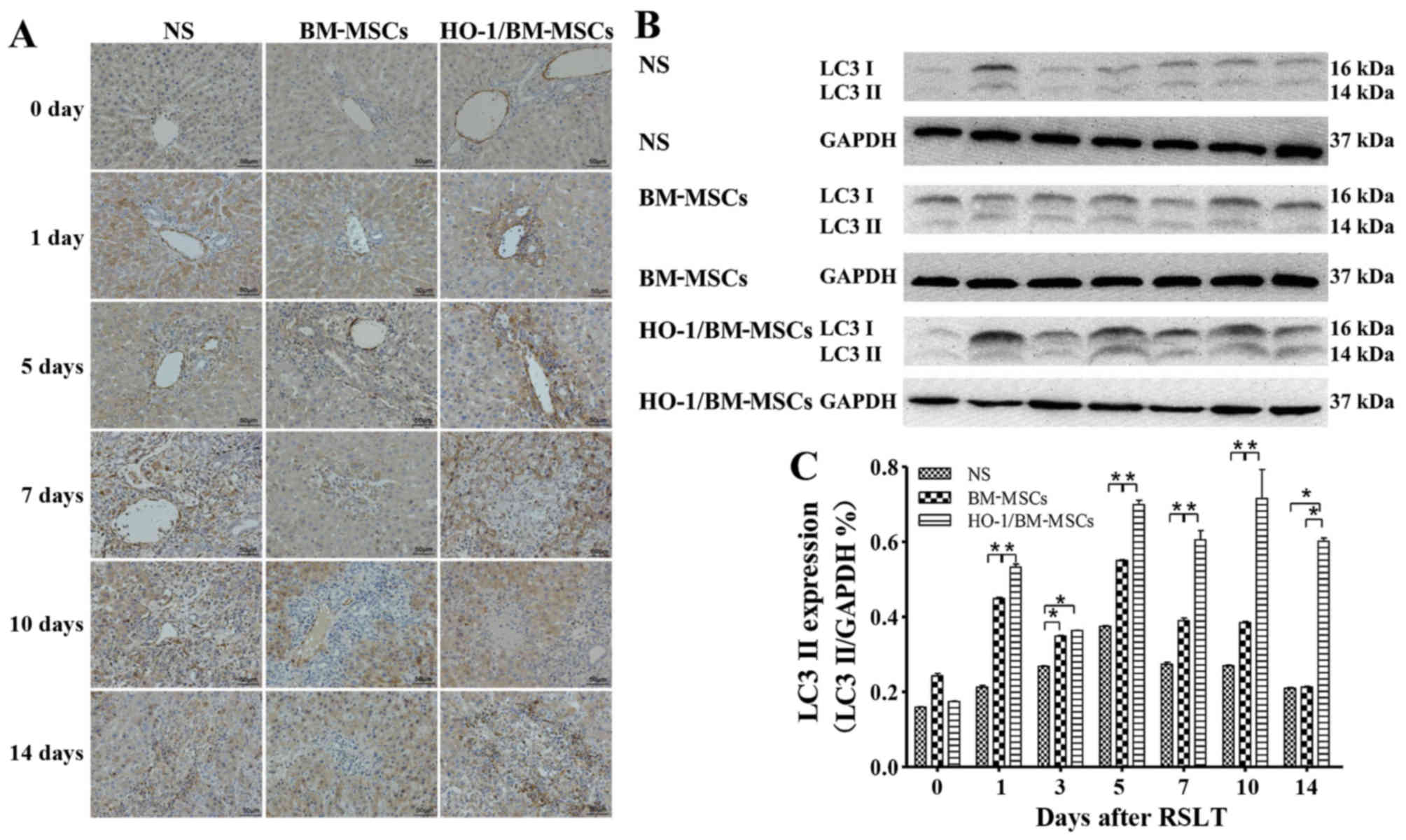 | Figure 5Expression of autophagy-related
protein LC3 I/II following RSLT. (A) Expression of LC3 I/II protein
immunohistochemically on POD 0, POD 1, POD 5, POD 7, POD 10 and POD
14. Only a small amount of LC3 I/II protein was present in
endothelia of the central vein on POD 0, which was similar in all
three groups. The LC3 I/II protein was also visible in cytoplasm of
hepatocytes on POD 1. The protein abundance increased significantly
after POD 5, appearing in the cytoplasm of most central venous
endothelia and hepatocytes. The overall expression in the
HO-1/BM-MSCs-treated group was the highest and was much higher than
that in the normal saline (NS)-treated group and in the
BM-MSCs-treated group. (B and C) Expression of the LC3 II protein
as demonstrated by western blotting on POD 0, POD 1, POD 3, POD 5,
POD 7, POD 10 and POD 14. The protein level in the
HO-1/BM-MSCs-treated group was significantly higher than that of
the NS-treated group and BM-MSCs-treated group at each time-point,
except on POD 0 and POD 3. *P<0.05 as indicated.
RSLT, reduced-size liver transplantation; POD, post-operative day;
HO-1/BM-MSCs, HO-1 transduced BM-MSCs; BM-MSCs, bone marrow
mesenchymal stem cells. |
Western blotting indicated that the LC3 II protein
level was relative low in all three groups on POD 0, with no
significant difference between them. The protein level in the
HO-1/BM-MSCs-treated group on POD 1 was significantly higher than
that in the NS and the BM-MSCs-treated groups (P<0.05). The
protein level in the HO-1/BM-MSCs-treated group on POD 3 was
significantly higher than that of the NS-treated group (P<0.05),
but reported no statistically significant difference with that of
the BM-MSCs-treated group. The protein level increased
significantly on POD 5, POD 7, POD 10 and POD 14, especially in the
HO-1/BM-MSCs-treated group. The protein level in the
BM-MSCs-treated group on POD 5, POD 7 and POD 10 was significantly
higher than that of the NS-treated group (P<0.05), and the
protein level in the HO-1/BM-MSCs-treated group was significantly
higher than that of the BM-MSCs-treated group (P<0.05) except on
POD 0 and POD 3. These results demonstrated that the level of
autophagy-related protein LC3 I/II in the HO-1/BM-MSCs-treated
group was the highest (Fig. 5B and
C).
Levels of the Beclin-1 protein
Immunohistochemical tests presented only a small
amount of Beclin-1 protein in the cytoplasm of hepatocytes around
the central vein in all three groups on POD 0, with no significant
difference between the three groups. Beclin-1 could also be
observed in the biliary epithelia on POD 1. The Beclin-1 level
increased after POD 5 as the time after RSLT prolonged, and the
overall Beclin-1 level in the HO-1/BM-MSCs-treated group was much
higher than that in the NS and the BM-MSCs-treated groups (Fig. 6A).
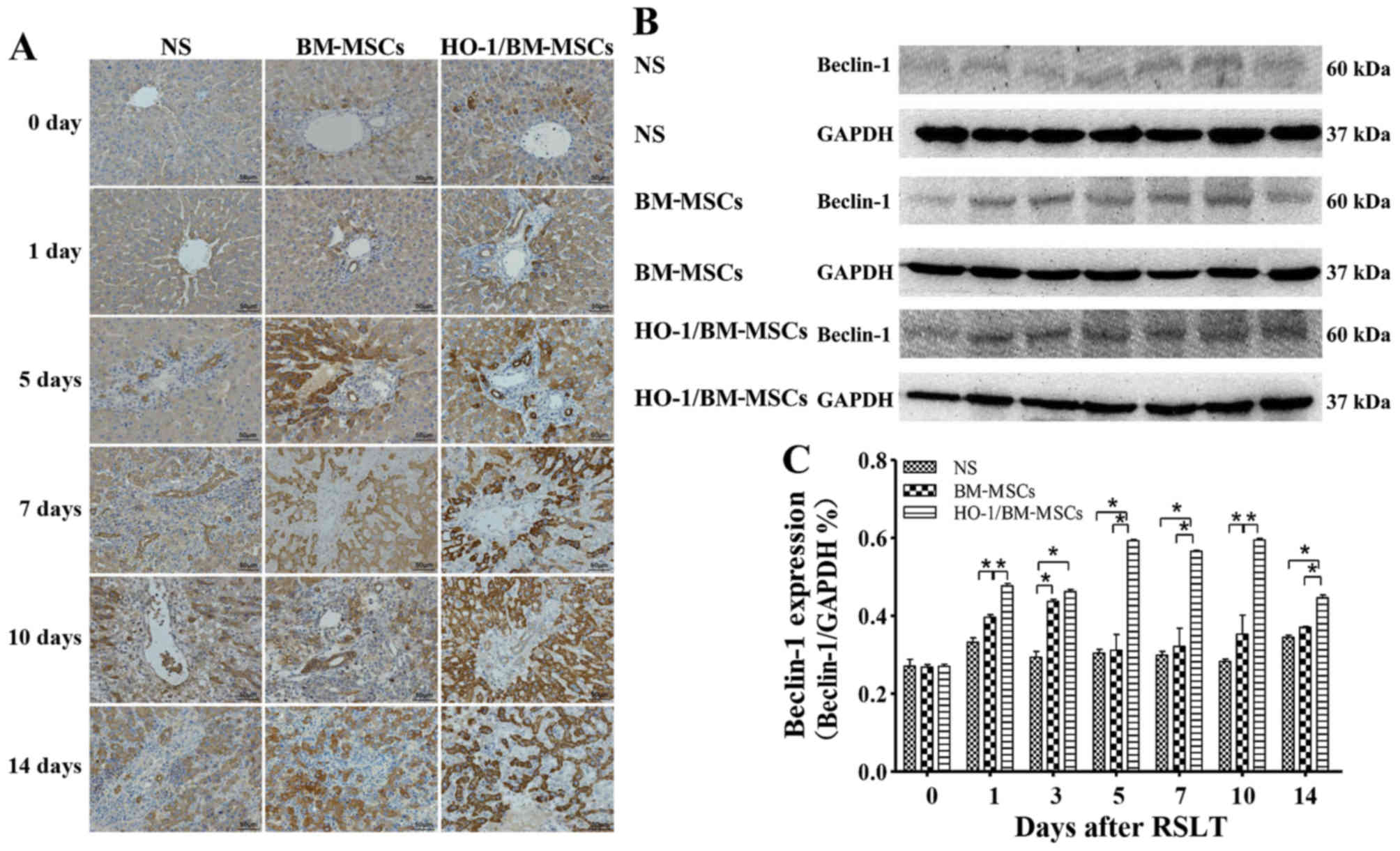 | Figure 6Levels of autophagy-related protein
Beclin-1 after RSLT. (A) Levels of Beclin-1 protein as assessed
immunohistochemically on POD 0, POD 1, POD 5, POD 7, POD 10 and POD
14. Only a small amount of Beclin-1 protein was present in the
cytoplasm of hepatocytes around the central vein on POD 0, which
was similar in all three groups. Beclin-1 could also be observed in
the biliary epithelia on POD 1. The protein expression increased
after POD 5, extending to the cytoplasm of most hepatocytes and
biliary endothelia. The overall protein level in the
HO-1/BM-MSCs-treated group was the highest and was much higher than
that in the normal saline (NS)-treated group and the
BM-MSCs-treated group. (B and C) Levels of the Beclin-1 protein as
demonstrated by western blotting on POD 0, POD 1, POD 3, POD 5, POD
7, POD 10 and POD 14. The protein abundance in the BM-MSCs-treated
group was significantly higher than that in the NS-treated group at
each time-point, except on POD 5, POD 7 and POD 14. The protein
abundance in the HO-1/BM-MSCs-treated group was significantly
higher than that in the NS-treated group and BM-MSCs-treated group
at each time-point, except on POD 0. *P<0.05 as
indicated. RSLT, reduced-size liver transplantation; POD,
post-operative day; HO-1/BM-MSCs, HO-1 transduced BM-MSCs; BM-MSCs,
bone marrow mesenchymal stem cells. |
Western blotting demonstrated that the amount of
Beclin-1 protein was relatively low in all three groups on POD 0,
with no significant difference between them. The Beclin-1 level in
the HO-1/BM-MSCs-treated group on POD 1 was significantly higher
than that in the NS and the BM-MSCs-treated groups (P<0.05). The
Beclin-1 level in the HO-1/BM-MSCs-treated group on POD 3 was
significantly higher than that in the NS-treated group (P<0.05),
but reported no statistically significant difference with that of
the BM-MSCs-treated group. The Beclin-1 level increased
significantly on POD 5, POD 7, POD 10 and POD 14, especially in the
HO-1/BM-MSCs-treated group. The Beclin-1 level in the
HO-1/BM-MSCs-treated group was significantly higher than that of
the NS and BM-MSCs-treated group (P<0.05). These results
indicated that the HO-1/BM-MSCs-treated group produced the highest
amount of the autophagy-related protein Beclin-1 (Fig. 6B and C).
Autophagy regulated by the ERK/mTOR
signaling pathway is involved in the regulation of the protective
effects of HO-1/BM-MSCs on RSLT
The amount of ERK mRNA in the HO-1/BM-MSCs-treated
group was significantly higher than that of the NS-treated group at
each time-point, and was significantly higher than that in the
BM-MSCs-treated group, except on POD 0 and POD 10 (P<0.05). The
amount of p-ERK protein in the HO-1/BM-MSCs-treated group on POD 0
was significantly higher than that in the NS-treated group
(P<0.05), but demonstrated no significant difference with the
BM-MSCs-treated group. The amount of p-ERK protein in the
HO-1/BM-MSCs-treated group was significantly higher than that in
the NS and BM-MSCs-treated groups on POD 1 and POD 3 (P<0.05).
The overall amount of p-ERK protein increased significantly on POD
5, POD 7, POD 10 and POD 14, especially in the HO-1/BM-MSCs-treated
group. The amount of p-ERK in the HO-1/BM-MSCs-treated group was
significantly higher than that of the NS-treated group at each
time-point (P<0.05). The amount of p-ERK in the
HO-1/BM-MSCs-treated group was significantly higher than that of
the BM-MSCs-treated group at each time-point, except on POD 7
(P<0.05; Fig. 7A).
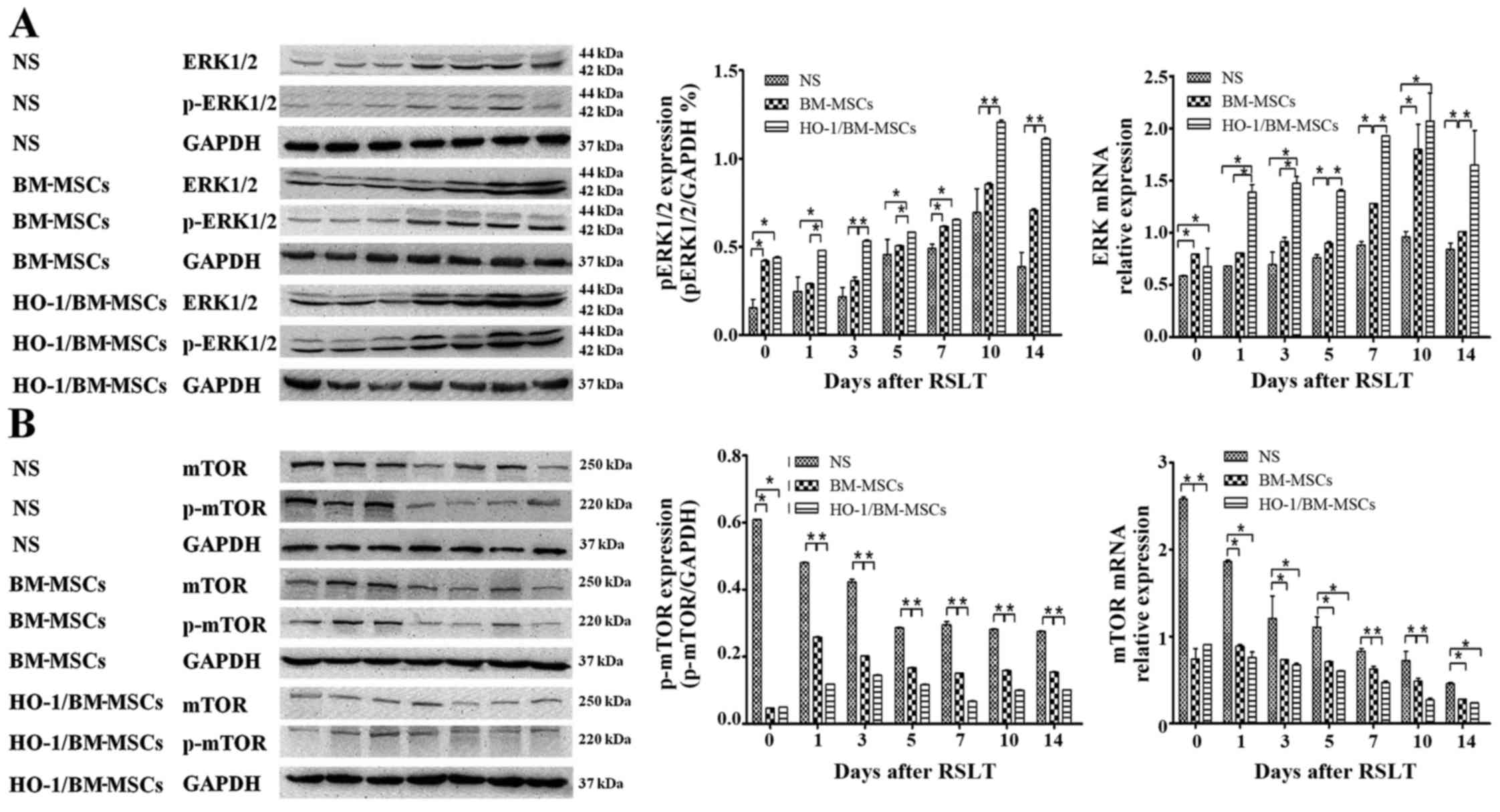 | Figure 7Expression of ERK1/2, p-ERK1/2, mTOR
and p-mTOR after reduced-size liver transplantation. (A) Relative
content of ERK1/2 to p-ERK1/2, as measured by RT-qPCR and western
blotting. The amount of ERK mRNA in the HO-1/BM-MSCs-treated
group was significantly higher than that of the normal saline
(NS)-treated group at each time-point, and that in the
HO-1/BM-MSCs-treated group was significantly higher than that of
the BM MS treated-group except on POD 0 and POD 10. The amount of
p-ERK1/2 in the BM-MSCs-treated group was significantly higher than
that in the NS-treated group on POD 0, POD 3, POD 7, POD 10 and POD
14. The amount of p-ERK1/2 in the HO-1/BM-MSCs-treated group was
significantly higher than that of the BM-MSCs-treated group at each
time-point, except on POD 0 and POD 7, and was significantly higher
than that in the NS-treated group at each time-point. (B) Relative
content of mTOR to p-mTOR, as measured by RT-qPCR and western
blotting. The amount of mTOR mRNA in the
HO-1/BM-MSCs-treated group was significantly lower than that of the
NS-treated group at each time-point, and that in the
HO-1/BM-MSCs-treated group was significantly lower than the
BM-MSCs-treated group except on POD 7 and POD 10. The amount of
p-mTOR in the BM-MSCs-treated group was significantly lower than
that in the NS-treated group at each time-point. The level of
p-mTOR in the HO-1/BM-MSCs-treated group was significantly lower
than that in the BM-MSCs-treated group at each time-point, except
on POD 0, and was significantly lower than that in the NS-treated
group at each time-point. *P<0.05 as indicated. ERK,
extracellular signal-regulated kinase; mTOR, mammalian target of
rapamycin; HO-1/BM-MSCs, HO-1 transduced BM-MSCs; POD,
post-operative day; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction. |
The amount of mTOR mRNA in the
HO-1/BM-MSCs-treated group was lower than that of the NS-treated
group at each time-point, and that in the HO-1/BM-MSCs-treated
group was lower than the BM-MSCs-treated group on POD 7 and POD 10.
The amount of p-mTOR protein in the HO-1/BM-MSCs-treated group on
POD 0 was significantly lower than that of the NS and
BM-MSCs-treated groups (P<0.05). The amount of p-mTOR protein in
the HO-1/BM-MSCs-treated group was also significantly lower than
that in the NS and BM-MSCs-treated groups on POD 1 and POD 3
(P<0.05). The overall amount of p-mTOR protein decreased
significantly on POD 5, POD 7, POD 10 and POD 14, especially in the
HO-1/BM-MSCs-treated group. The amount of p-mTOR in the
HO-1/BM-MSCs-treated group was significantly lower than that of the
NS and BM-MSCs-treated groups (P<0.05). These results suggested
that the amount of p-ERK protein increased, while the amount of
p-mTOR protein decreased in the HO-1/BM-MSCs-treated group, which
suggested that the protective effects of HO-1/BM-MSCs on RSLT may
involve autophagy regulated by the ERK/mTOR signaling pathway
(Fig. 7B).
Discussion
Although RSLT could alleviate the problem of organ
shortage for liver transplantation to some extent (1), it also weakens the regenerative
ability of the transplanted liver (2,3).
Previous studies have indicated that BM-MSCs could inhibit the
death of hepatocytes via paracrine or direct differentiation into
hepatocytes, which promotes the regeneration of the liver after
injury (22–24). HO-1, also known as a 'stop signal'
of inflammation, has anti-inflammatory and anti-apoptotic effects
(25). HO-1 is the inducible form
of the HO family, and is the rate-limiting enzyme that catalyzes
heme degradation to CO, iron and biliverdin. The main function of
HO-1 is to protect cells under oxidative stress and other stimuli.
Many factors, such as infection, inflammation and hypoxia induce
HO-1 (26). The pathological and
TUNEL analysis of transplanted livers in the current study
indicated that HO-1/BM-MSCs could promote liver regeneration and
suppress rejection of the transplanted liver significantly, with
better protective effects, which was consistent with our previous
studies (12,13); however, the exact mechanism
remained unclear. The authors observed autophagic vesicles in the
ultrastructure of transplanted livers after RSLT, which were more
obvious in the HO-1/BM-MSCs-treated group than in the
BM-MSCs-treated group and NS-treated group. Therefore, to expand on
the previous studies, the authors investigated whether autophagy is
involved in the protective effects of HO-1/BM-MSCs on the
transplanted liver following RSLT.
Soluble autophagy microtubule-associated protein 1
light chain 3 (LC3 I) is converted to LC3 II during the induction
of the autophagosome, and LC3 II is involved in the formation of
the autophagosome membrane. Thus, LC3 II may represent the number
of autophagosomes and is a good marker for their formation
(27). The process of autophagy
includes induction, nucleation, extension and formation of the
membrane, formation of autophagosomes and fusion with lysosomes
(28,29). Autophagy-related protein Beclin-1,
encoded by the BECN1 gene, is a major protein involved in the
nucleation of autophagosomes, and is the key target for regulation
of autophagy (30). Therefore,
the authors investigated the levels of these two proteins in the
model of RSLT. HO-1/BM-MSCs-treated group had the highest level of
autophagy-related proteins LC3 II and Beclin-1, which suggested
that significant amounts of autophagy occurred in the
HO-1/BM-MSCs-treated group, and was consistent with ultrastructure
results of the transplanted livers. Previous studies indicated that
autophagy, as a cytoprotective mechanism, is involved in biological
process such as growth, development and immune regulation (14,31). In addition, autophagy could
relieve ischemia-reperfusion injury and induce immune tolerance in
the organ transplantation (16,17,32–37). On the one hand,
ischemia-reperfusion injury involves many mechanisms, among which
autophagy can remove damaged mitochondria to prevent the
accumulation of abnormal mitochondria and toxic products in the
cytoplasm (mitochondrial autophagy) (32). Swelling and structural damage of
mitochondria in the liver of transgenic mice with autophagic
defects have been reported (33).
Autophagy could also inhibit the release of
proinflammatory cytokines, such as tumor necrosis factor (TNF)-α,
interleukin (IL)-2, IL-6 and IL-1, and the high mobility group
proteins produced by activated Kupffer cells, neutrophils and
platelets, to prevent the death of hepatocytes (15–17). TNF-α could lead to swelling of the
endothelia and activation of ROS; IL-6 could damage hepatocytes,
and induce them to produce C-reactive protein, α-trypsin and
fibrinogen (34). These
observations were consistent with the authors' preliminary
experimental results that HO-1/BM-MSCs could reduce the levels of
IL-2 and TNF-α in the transplanted liver (12,13).
On the other hand, autophagy may be involved in the
induction of immune tolerance, among which thymic epithelial cells,
professional and non-professional antigen presenting cells from the
thymus and its periphery, could present autologous antigens to MHC
II molecules directly via autophagy, subsequently promoting the
central tolerance and peripheral immune tolerance of
CD4+ T cells (35).
Autophagy also inhibits the maturation of DCs, induces the
expression of CD4+CD25+Foxp3+ T
cells and CD8+CD25+Foxp3+ T cells,
and increases the secretion of IL-10 to induce allogeneic T cell
anergy (36,37), which were consistent with the
preliminary results suggesting that HO-1/BM-MSCs could increase
IL-10 and regulatory T cells in the transplanted liver (12,13). All of these effects of autophagy
in the field of organ transplantation were similar to those induced
by BM-MSCs.
BM-MSCs could also reduce ischemia-reperfusion
injury by secretion of cytokines and showed immunomodulatory
effects (8,9,38,39). A previous study demonstrated a
link between the effects of BM-MSCs and autophagy (40). In addition, induction of HO-1 is
an adaptive response to recover cell homeostasis (such as
autophagy) (26). HO-1 increases
the signals of autophagy, and inhibition of HO-1 activity or HO-1
knockout led to reduced autophagy in hepatocytes, with increased
cell death; inhibition of autophagy also weakened the
anti-inflammatory effects of HO-1 (41–43). Therefore, autophagy and expression
of autophagy-related proteins are associated closely with BM-MSCs
and HO-1.
In the present study, the authors further found that
autophagy was enhanced and apoptosis was weakened in the
HO-1/BM-MSCs-treated group, which suggested that autophagy is
involved in the protective effects of HO-1/BM-MSCs on transplanted
livers following RSLT. In addition, the amount of LC3 II protein
was higher on POD 1 than on POD 3, while it was lower on POD 14
than POD 10. The authors further analyzed the mechansim. Previous
studies have found that starvation is a potent inducer of antophagy
(44). In response to starvation,
autophagy in the liver of rodents increased, and the rate of
protein degradation was also increased to maintain cell function
(14). As a result, the authors
believe that this phenomenon probably correlates with starvation of
the transplanted liver, since the recipient eats less food on POD
1. As the degree of rejection increased over time, the autophagy of
remnant hepatocytes was not sufficient to counter the severe
rejection on POD 14, so the amount of LC3 II protein was lower on
POD 14 than POD 10.
Autophagy is a complex process that is tightly
regulated by >30 autophagy-related proteins, and by signaling
pathways such as the mammalian target of rapamycin (mTOR),
adenosine monophosphate-activated protein kinase and
hypoxia-inducible factor pathways (45). Among them, mTOR is an important
receptor of intracellular energy and nutritional status. mTOR is an
evolutionarily conserved serine/threonine protein kinase, and is a
negative regulator of autophagy proteins. It comprises two
different complexes: mTORC1 and mTORC2 (46). mTORC1 is the focus of many
upstream stimuli and signals, including MEK/ERK and PI3K/AKT, which
regulate autophagy and other cellular activities (46). Among them, ERK is a member of the
mitogen-activated protein kinase family and responds to both
intracellular and extracellular stimuli. p-ERK regulates
cytoskeletal proteins, kinases and transcription factors, leading
to altered gene expression, cell proliferation and differentiation
(47–49). Therefore, the ERK/mTOR signaling
pathway was chosen to explore its possible involvement in the
mechanism of autophagy associated with the protective effects of
HO-1/BM-MSCs on transplanted livers after RSLT. The present study
demonstrated increased p-ERK and decreased p-mTOR in the
BM-MSCs-treated and HO-1/BM-MSCs-treated groups. The results in the
HO-1/BM-MSCs-treated group were more dramatic. Activation of
MEK/ERK and inhibition of mTORC1 could enhance the expression of
autophagy-related proteins, leading to increased autophagic
activity, which could protect cells (50,51). The results of the present study
demonstrated that ERK is a negative regulator of the upstream
mTORC1, and autophagy may have protective effects on cells via this
signaling pathway. Therefore, it is hypothesized that autophagy is
involved in the protective effects of HO-1/BM-MSCs on the
transplanted liver after RSLT through the ERK/mTOR signaling
pathway.
To the best of the authors' knowledge, the present
study is the first to demonstrate that autophagy is involved in the
protective effects of HO-1/BM-MSCs on the transplanted liver after
RSLT. Regulation of autophagy is primarily mediated through the
ERK/mTOR signaling pathway. In a future study, the authors intend
to carry out related experiments (such as the use of inhibitors,
inducers and pathway blockers of autophagy, autophagy-related gene
knockout animal models). Future studies will also focus on
additional in vitro experiments to provide a theoretical
basis for the reduction of transplanted liver damage after RSLT and
its wider clinical application.
Acknowledgments
The authors would like to thank the Key Laboratory
of the Emergency and Care Medicine of Ministry of Health and
Tianjin Key Laboratory of Organ Transplantation (Tianjin, China)
for allowing this work to progress in their laboratories. The
present study was supported by the National Natural Science
Foundation of China (grant nos. 81670574, 81441022 and 81270528);
the Natural Science Foundation of Tianjin, China (grant nos.
08JCYBJC08400, 11JCZDJC27800 and 12JCZDJC25200); and the Technology
Foundation of the Health Bureau in Tianjin, China (grant no.
2011KY11).
Abbreviations:
|
RSLT
|
reduced-size liver transplantation
|
|
BM-MSCs
|
bone marrow-derived mesenchymal stem
cells
|
|
HO-1
|
heme oxygenase-1
|
|
HO-1/BM-MSCs
|
HO-1-transduced BM-MSCs
|
|
ROS
|
reactive oxygen species
|
|
GFP
|
green fluorescence protein
|
|
POD
|
post-operative day
|
|
LC3-I
|
soluble autophagy
microtubule-associated protein 1 light chain 3
|
|
PV
|
portal vein
|
|
IHVC
|
infrahepatic vena cava
|
|
SHVC
|
suprahepatic vena cava
|
|
mTOR
|
mammalian target of rapamycin
|
References
|
1
|
Müller SA, Mehrabi A, Schmied BM, Welsch
T, Fonouni H, Engelmann G, Schemmer P, Weitz J and Schmidt J:
Partial liver transplantation-living donor liver transplantation
and split liver transplantation. Nephrol Dial Transplant. 22(Suppl
8): viii13–viii22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dahm F, Georgiev P and Clavien PA:
Small-for-size syndrome after partial liver transplantation:
Definition, mechanisms of disease and clinical implications. Am J
Transplant. 5:2605–2610. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Taki-Eldin A, Zhou L, Xie HY and Zheng SS:
Liver regeneration after liver transplantation. Eur Surg Res.
48:139–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goss JA, Yersiz H, Shackleton CR, Seu P,
Smith CV, Markowitz JS, Farmer DG, Ghobrial RM, Markmann JF,
Arnaout WS, et al: In situ splitting of the cadaveric liver for
transplantation. Transplantation. 64:871–877. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Davani S, Marandin A, Mersin N, Royer B,
Kantelip B, Hervé P, Etievent JP and Kantelip JP: Mesenchymal
progenitor cells differentiate into an endothelial phenotype,
enhance vascular density, and improve heart function in a rat
cellular cardiomyoplasty model. Circulation. 108(Suppl 1):
II253–II258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Popp FC, Piso P, Schlitt HJ and Dahlke MH:
Therapeutic potential of bone marrow stem cells for liver diseases.
Curr Stem Cell Res Ther. 1:411–418. 2006. View Article : Google Scholar
|
|
7
|
Zhou QI, Yang C and Yang P: The
promotional effect of mesenchymal stem cell homing on bone tissue
regeneration. Curr Stem Cell Res Ther. 2015.
|
|
8
|
Ren G, Zhang L, Zhao X, Xu G, Zhang Y,
Roberts AI, Zhao RC and Shi Y: Mesenchymal stem cell-mediated
immunosuppression occurs via concerted action of chemokines and
nitric oxide. Cell Stem Cell. 2:141–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Spaggiari GM, Abdelrazik H, Becchetti F
and Moretta L: MSCs inhibit monocyte-derived DC maturation and
function by selectively interfering with the generation of immature
DCs: Central role of MSC-derived prostaglandin E2. Blood.
113:6576–6583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang HF, Zeng Z, Wang KH, Zhang HY, Wang
S, Zhou WX, Wang ZB, Xu WG and Duan J: Heme oxygenase-1 protects
rat liver against warm ischemia/reperfusion injury via
TLR2/TLR4-triggered signaling pathways. World J Gastroenterol.
21:2937–2948. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeng B, Lin G, Ren X, Zhang Y and Chen H:
Overexpression of HO-1 on mesenchymal stem cells promotes
angiogenesis and improves myocardial function in infarcted
myocardium. J Biomed Sci. 17:802010. View Article : Google Scholar
|
|
12
|
Shen ZY, Wu B, Liu T, Yang Y, Yin ML,
Zheng WP, Zhang BY and Song HL: Immunomodulatory effects of bone
marrow mesenchymal stem cells overexpressing heme oxygenase-1:
Protective effects on acute rejection following reduced-size liver
transplantation in a rat model. Cell Immunol. 313:10–24. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu B, Song HL, Yang Y, Yin ML, Zhang BY,
Cao Y, Dong C and Shen ZY: Improvement of liver transplantation
outcome by heme oxygenase-1-transduced bone marrow mesenchymal stem
cells in rats. Stem Cells Int. 2016:92350732016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Czaja MJ, Ding WX, Donohue TM Jr, Friedman
SL, Kim JS, Komatsu M, Lemasters JJ, Lemoine A, Lin JD, Ou JH, et
al: Functions of autophagy in normal and diseased liver. Autophagy.
9:1131–1158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li L, Tan J, Miao Y, Lei P and Zhang Q:
ROS and autophagy: Interactions and molecular regulatory
mechanisms. Cell Mol Neurobiol. 35:615–621. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu J, Hao H, Huang H, Tong C, Ti D, Dong
L, Chen D, Zhao Y, Liu H, Han W, et al: Hypoxia regulates the
therapeutic potential of mesenchymal stem cells through enhanced
autophagy. Int J Low Extrem Wounds. 14:63–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bhogal RH, Weston CJ, Curbishley SM, Adams
DH and Afford SC: Autophagy: A cyto-protective mechanism which
prevents primary human hepatocyte apoptosis during oxidative
stress. Autophagy. 8:545–558. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Klein L, Münz C and Lünemann JD:
Autophagy-mediated antigen processing in CD4+ T cell
tolerance and immunity. FEBS Lett. 584:1405–1410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao Q, Ren H, Zhu D and Han Z:
Stem/progenitor cells in liver injury repair and regeneration. Biol
Cell. 101:557–571. 2009.PubMed/NCBI
|
|
20
|
No authors listed. Banff schema for
grading liver allograft rejection: An international consensus
document. Hepatology. 25:658–663. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
van Poll D, Parekkadan B, Cho CH,
Berthiaume F, Nahmias Y, Tilles AW and Yarmush ML: Mesenchymal stem
cell-derived molecules directly modulate hepatocellular death and
regeneration in vitro and in vivo. Hepatology. 47:1634–1643. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rabani V, Shahsavani M, Gharavi M, Piryaei
A, Azhdari Z and Baharvand H: Mesenchymal stem cell infusion
therapy in a carbon tetrachloride-induced liver fibrosis model
affects matrix metalloproteinase expression. Cell Biol Int.
34:601–605. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fouraschen SM, Wolf JH, van der Laan LJ,
de Ruiter PE, Hancock WW, van Kooten JP, Verstegen MM, Olthoff KM
and de Jonge J: Mesenchymal stromal cell-derived factors promote
tissue repair in a small-for-size ischemic liver model but do not
protect against early effects of ischemia and reperfusion injury. J
Immunol Res. 2015:2029752015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Otterbein LE, Soares MP, Yamashita K and
Bach FH: Heme oxygenase-1: Unleashing the protective properties of
heme. Trends Immunol. 24:449–455. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Waltz P, Carchman EH, Young AC, Rao J,
Rosengart MR, Kaczorowski D and Zuckerbraun BS: Lipopolysaccaride
induces autophagic signaling in macrophages via a TLR4, heme
oxygenase-1 dependent pathway. Autophagy. 7:315–320. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tanida I, Minematsu-Ikeguchi N, Ueno T and
Kominami E: Lysosomal turnover, but not a cellular level, of
endogenous LC3 is a marker for autophagy. Autophagy. 1:84–91. 2005.
View Article : Google Scholar
|
|
28
|
Loos B, Engelbrecht AM, Lockshin RA,
Klionsky DJ and Zakeri Z: The variability of autophagy and cell
death susceptibility: Unanswered questions. Autophagy. 9:1270–1285.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Komatsu M: Liver autophagy: Physiology and
pathology. J Biochem. 152:5–15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu Q, Sun Y, Lv Y, Le Z, Xin Y, Zhang P
and Liu Y: TERT alleviates irradiation-induced late rectal injury
by reducing hypoxia-induced ROS levels through the activation of
NF-κB and autophagy. Int J Mol Med. 38:785–793. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee S and Kim JS: Mitophagy: Therapeutic
potentials for liver disease and beyond. Toxicol Res. 30:243–250.
2014. View Article : Google Scholar
|
|
33
|
Takamura A, Komatsu M, Hara T, Sakamoto A,
Kishi C, Waguri S, Eishi Y, Hino O, Tanaka K and Mizushima N:
Autophagy-deficient mice develop multiple liver tumors. Genes Dev.
25:795–800. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shen M, Lu J, Dai W, Wang F, Xu L, Chen K,
He L, Cheng P, Zhang Y, Wang C, et al: Ethyl pyruvate ameliorates
hepatic ischemia-reperfusion injury by inhibiting intrinsic pathway
of apoptosis and autophagy. Mediators Inflamm. 2013:4615362013.
View Article : Google Scholar
|
|
35
|
Lünemann JD and Münz C: Autophagy in
CD4+ T-cell immunity and tolerance. Cell Death Differ.
16:79–86. 2009. View Article : Google Scholar
|
|
36
|
Li X, Li JJ, Yang JY, Wang DS, Zhao W,
Song WJ, Li WM, Wang JF, Han W, Zhang ZC, et al: Tolerance
induction by exosomes from immature dendritic cells and rapamycin
in a mouse cardiac allograft model. PLoS One. 7:e440452012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li XC, Strom TB, Turka LA and Wells AD: T
cell death and transplantation tolerance. Immunity. 14:407–416.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Isbambetov A, Baimakhanov Z, Soyama A,
Hidaka M, Sakai Y, Takatsuki M, Kuroki T and Eguchi S: Equal
distribution of mesenchymal stem cells after hepatic
ischemia-reperfusion injury. J Surg Res. 203:360–367. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jin G, Qiu G, Wu D, Hu Y, Qiao P, Fan C
and Gao F: Allogeneic bone marrow-derived mesenchymal stem cells
attenuate hepatic ischemia-reperfusion injury by suppressing
oxidative stress and inhibiting apoptosis in rats. Int J Mol Med.
31:1395–1401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nuschke A, Rodrigues M, Stolz DB, Chu CT,
Griffith L and Wells A: Human mesenchymal stem cells/multipotent
stromal cells consume accumulated autophagosomes early in
differentiation. Stem Cell Res Ther. 5:1402014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yun N, Cho HI and Lee SM: Impaired
autophagy contributes to hepatocellular damage during
ischemia/reperfusion: Heme oxygenase-1 as a possible regulator.
Free Radic Biol Med. 68:168–177. 2014. View Article : Google Scholar
|
|
42
|
Carchman EH, Rao J, Loughran PA, Rosengart
MR and Zuckerbraun BS: Heme oxygenase-1-mediated autophagy protects
against hepatocyte cell death and hepatic injury from
infection/sepsis in mice. Hepatology. 53:2053–2062. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang Y, Xiong X, Guo H, Wu M, Li X, Hu Y,
Xie G, Shen J and Tian Q: ZnPP reduces autophagy and induces
apoptosis, thus aggravating liver ischemia/reperfusion injury in
vitro. Int J Mol Med. 34:1555–1564. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Deretic V, Saitoh T and Akira S: Autophagy
in infection, inflammation and immunity. Nat Rev Immunol.
13:722–737. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
White E, Mehnert JM and Chan CS:
Autophagy, metabolism, and cancer. Clin Cancer Res. 21:5037–5046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang Z and Klionsky DJ: Mammalian
autophagy: Core molecular machinery and signaling regulation. Curr
Opin Cell Biol. 22:124–131. 2010. View Article : Google Scholar :
|
|
47
|
Ma K, Huang MY, Guo YX and Hu GQ:
Matrine-induced autophagy counteracts cell apoptosis via the ERK
signaling pathway in osteosarcoma cells. Oncol Lett. 12:1854–1860.
2016.PubMed/NCBI
|
|
48
|
Lu Z and Xu S: ERK1/2 MAP kinases in cell
survival and apoptosis. IUBMB Life. 58:621–631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wen J, Zhao Y and Guo L: Orexin A induces
autophagy in HCT-116 human colon cancer cells through the ERK
signaling pathway. Int J Mol Med. 37:126–132. 2016. View Article : Google Scholar
|
|
50
|
Wang PR, Wang JS, Zhang C, Song XF, Tian N
and Kong LY: Huang-Lian-Jie-Du-Decotion induced protective
autophagy against the injury of cerebral ischemia/reperfusion via
MAPK-mTOR signaling pathway. J Ethnopharmacol. 149:270–280. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang J, Chen Q, Tian S, Song S, Liu F,
Wang Q and Fu Z: The role of 1,25-dyhydroxyvitamin D3 in mouse
liver ischemia reperfusion injury: Regulation of autophagy through
activation of MEK/ERK signaling and PTEN/PI3K/Akt/mTORC1 signaling.
Am J Transl Res. 7:2630–2645. 2015.
|