Introduction
Multiple myeloma (MM), also known as plasma cell
myeloma, myelomatosis or Kahler's disease (1), is a cancer of plasma cells (the
white blood cells that produce antibodies), and is considered to be
incurable (2). The incidence of
MM is ~4/100,000 and it accounts for 10% of all hematological
malignancies (3). The treatment
of MM is complex due to rapid advances in stem cell
transplantation, medications and supportive care, which have
improved the survival times of MM patients over the past 30 years
(4). The principal treatments
include stem cell (bone marrow) transplantation,
non-chemotherapeutic drugs that target cancer cells, standard
chemotherapeutic drugs, and corticosteroids (5). However, despite various treatment
options for MM, advanced combination therapies with effective
agents are required for the efficient treatment of the disease in
the first-line and relapsed settings, and this has resulted in the
development of novel agents (6,7).
Traditional Chinese medicine (TCM) is considered to
be an important source of therapies for various diseases. In 2015,
a Chinese scientist won the Nobel Prize for her work in drug
development from TCM; indeed, numerous drug candidate compounds
from TCM are under investigation (8). Triptolide is a diterpenoid epoxide
that can be isolated from the traditional Chinese medicinal herb
Tripterygium wilfordii Hook F., which is used to treat
inflammatory and autoimmune diseases (9). Following the initial identification
of triptolide in 1972, in vivo and in vitro
experiments have revealed it to exhibit a variety of bioactivities,
including immunosuppression and antiinflammatory effects (10,11). Triptolide has a complex mechanism
of action, which involves inhibition of transcription factor
nuclear factor κB (NF-κB) activation (12,13), suppression of the production of
prostaglandin E2, and reduction of cytokine levels
(13,14).
In addition to the aforementioned activities, it was
recently reported that triptolide has antitumor effects in
pancreatic, gastric, breast, adrenal and thyroid cancer, as well as
neuroblastoma (1,15–19). Likewise, a number of studies on
the effects of triptolide on various types of cancer cells,
including MM, are currently underway. Although there have been
numerous attempts to explain the molecular mechanisms underlying
the effect of triptolide, the mechanism in MM cells remains
unclear. It has been proposed that triptolide induces epigenetic
alterations via the regulation of histone methylation (20), and induces apoptosis via the
activation of caspases-8, -9 and -3 (cysteine proteases) in MM
cells (21). However, a greater
understanding of its mechanism of action in MM cells is necessary
in order to develop novel anticancer drugs from triptolide, due to
its potential multi-organ toxicity and narrow therapeutic
window.
Notably, triptolide inhibits the Janus kinase (JAK)2
pathway in myeloproliferative disorder cells, which is a similar
disease to MM (22), and it
inhibits the JAK1/signal transducer and activator of transcription
3 (STAT3) and JAK2/STAT3 pathways in colon cancer cells and in a
rat neuropathic pain model, respectively (23,24). STATs consist of a family of six
transcription factors that regulate tumor development (25). Ligand-induced activation of
cell-surface cytokine receptors activates the JAK family, which
phosphorylates and activates latent cytoplasmic STAT3 protein to
form an active dimer (26).
Activated STAT3 translocates to the nucleus and induces
transcription of STAT3-regulated genes (26). Several cytokines, including IL-6
(27), transforming growth
factor-α (28) and epidermal
growth factor (EGF) (27) have
been shown to activate STAT3. Numerous studies have proposed that
the activation of STAT3 can suppress apoptosis, and promote
proliferation, angiogenesis, chemoresistance and inflammation
(29–32). Thus, the STAT3 pathway must be
considered to gain an understanding of oncogenesis (25). Therefore, in the present study,
the effect of triptolide on MM cells via the JAK and JAK2/STAT3
pathways was investigated.
The present study aimed to investigate the effects
of triptolide on the STAT3 activation pathway and the underlying
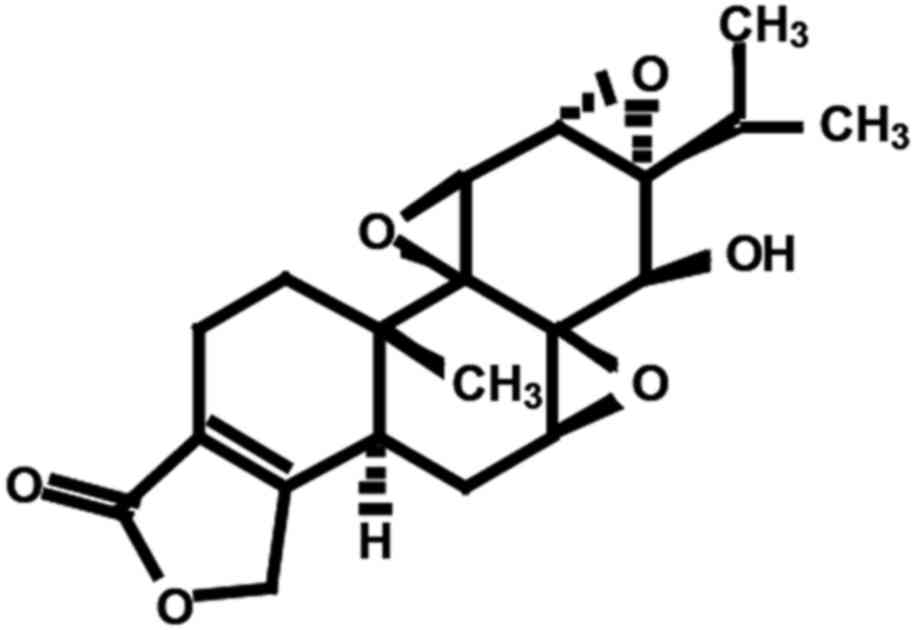
mechanism in MM cells. The structure of triptolide is shown in
Fig. 1. The effect of triptolide
on constitutive and IL-6-induced STAT3 activation was evaluated.
The effect of triptolide on STAT3-mediated gene products associated
with cellular proliferation, survival and apoptosis was also
investigated. Triptolide was found to suppress STAT3 activation by
activating protein tyrosine phosphatase non-receptor type 6
(SHP-1), and down-regulated the expression of STAT3-regulated
antiapoptotic, proliferative and angiogenic proteins.
Materials and methods
Materials
A 50-mM solution of triptolide with purity >95%
was kindly provided by Dr Aggarwal at MD Anderson Cancer Center
(Houston, TX, USA). Hoechst 33342,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
Tris, glycine, NaCl, SDS, and bovine serum albumin (BSA) were
purchased from Sigma-Aldrich (Merck Millipore, Darmstadt, Germany).
RPMI-1640 medium, fetal bovine serum (FBS), 0.4% trypan blue vital
stain, and antibiotic/antimycotic mixture were purchased from
Invitrogen (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The
rabbit polyclonal anti-STAT3 antibody (#sc-482) and mouse
monoclonal antibodies against phospho (p)-STAT3 (Tyr-705;
#sc-8059), Bcl-xL (#sc-8392), SHP-1 (#sc-7289), cyclin D1
(#sc-753), procas-pase-3 (#sc-373730), JAK2 (#sc-278), VEGF
(#sc-53462), Mcl-1 (#sc-53951) and poly(ADP-ribose) polymerase
(PARP; #sc-7150) were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). The horseradish peroxidase (HRP)-conjugated
goat anti-rabbit antibody (#1662408) was purchased from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA). Antibodies against
phospho-specific Src (Tyr-416; #2101S), Src (#2108),
phospho-specific JAK1 (Tyr-1022/1023; #3331S), p-JAK2 (#3771S), and
JAK1 (#3332) were obtained from Cell Signaling Technology, Inc.
(Beverly, MA, USA). The HRP-conjugated goat anti-mouse antibody
(#554002) was purchased from Transduction Laboratories (Lexington,
KY, USA). Bacteria-derived recombinant human IL-6 was obtained from
Novartis Pharmaceuticals (East Hanover, NJ, USA).
Cell lines
The human MM cell lines U266 and
dexamethasone-sensitive MM.1S were purchased from the American Type
Culture Collection (ATCC; Manassas, VA, USA). Cell lines were
cultured in RPMI-1640 medium containing 10% FBS and 1X
antibiotic/antimycotic solution. Cells were maintained at 37°C in
an atmosphere of 5% CO2, 95% air.
Western blot analysis
For the detection of p-STAT3 (Tyr-705) and STAT3
proteins, triptolide-treated whole-cell extracts were lysed in
lysis buffer [20 mM Tris (pH 7.4), 250 mM NaCl, 2 mM EDTA (pH 8.0),
0.1% Triton X-100, 0.01 mg/ml aprotinin, 0.005 mg/ml leupeptin, 0.4
mM phenylmethylsulfonyl fluoride, and 4 mM
Na3VO4]. Lysates were then centrifuged at
16,025 × g for 10 min to remove insoluble material. The extracted
proteins were resolved on a 7.5% SDS polyacrylamide gel. Following
electrophoresis, the proteins were electrotransferred to a
nitrocellulose membrane, blocked with 5% non-fat milk, and probed
with anti-p-STAT3 antibodies (1:500) and anti-STAT3 antibodies
(1:1,000) overnight at 4°C. The blot was then washed, exposed to
HRP-conjugated secondary antibodies (1:10,000) at room temperature
for 1 h, and finally examined by enhanced chemiluminescence
(Amersham Biosciences, Piscataway, NJ, USA).
To detect the expression of STAT3-regulated proteins
and caspase-3, U266 cells (2×106/ml) were treated with
60 μM triptolide for the indicated times. The cells were
then washed and proteins were extracted by incubation for 30 min on
ice in 0.05 ml of buffer containing 20 mM HEPES (pH 7.4), 2 mM
EDTA, 250 mM NaCl, 0.1% Nonidet P-40, 2 μg/ml leupeptin, 2
μg/ml aprotinin, 1 mM phenylmethylsulfonyl fluoride, 0.5
μg/ml benzamidine, 1 mM DTT, and 1 mM sodium orthovanadate.
The lysate was centrifuged, and the supernatant was collected.
Whole-cell protein extract (50 μg) was resolved by 10%
SDS-PAGE; electrotransferred onto a nitrocellulose membrane;
blotted with antibodies against Bcl-xL (1:500), myeloid cell
leukemia-1 (Mcl-1; 1:500), cyclin D1 (1:1,000), VEGF (1:1,000), and
caspase-3 (1:1,000) overnight at 4°C. The blots were washed,
exposed to HRP-conjugated goat anti-mouse antibodies for 1 h at
room temperature, and then detected by enhanced
chemiluminescence.
Electrophoretic mobility shift assay
(EMSA)
DNA binding of STAT3 was analyzed by EMSA using a
32P-labeled high-affinity sis-inducible element probe,
as previously described (33).
Briefly, nuclear extracts were prepared from γ-T3-treated cells and
incubated with high-affinity sis-inducible element probes
(5′-CTTCATTTCCCGTAAATCCCTAAAGCT-3′ and
5′-AGCTTTAGGGATTTACGGGAAATGA-3′). The DNA-protein complexes that
formed were separated from free oligonucleotides on 5% native
polyacrylamide gels. The dried gels were visualized, and the
radioactive bands were quantitated with a Storm 820 Phosphorimager
and ImageQuant TL 7.0 (both from Amersham; GE Healthcare Life
Sciences, Little Chalfont, UK).
Western blot analysis of PARP
degradation
Triptolide-induced apoptosis was examined by
proteolytic cleavage of PARP. Briefly, cells (2×106/ml)
were treated with 60 μM triptolide for the indicated times
at 37°C. The cells were then washed and subjected to protein
extraction by incubation for 30 min on ice in 0.05 ml buffer
containing 20 mM HEPES (pH 7.4), 2 mM EDTA, 250 mM NaCl, 0.1%
Nonidet P-40, 2 ng/ml leupeptin, 2 ng/ml aprotinin, 1 mM
phenylmethylsulfonyl fluoride, 0.5 ng/ml benzamidine, 1 mM DTT, and
1 mM sodium orthovanadate. The lysate was centrifuged, and the
supernatant was collected. Protein extract (40 μg) was
resolved by 10% SDS-PAGE, electrotransferred onto a nitrocellulose
membrane, blotted with anti-PARP antibody, overnight at 4°C. The
blot was washed, exposed to HRP-conjugated goat anti-mouse antibody
for 1 h at room temperature, and then detected by enhanced
chemiluminescence.
Results
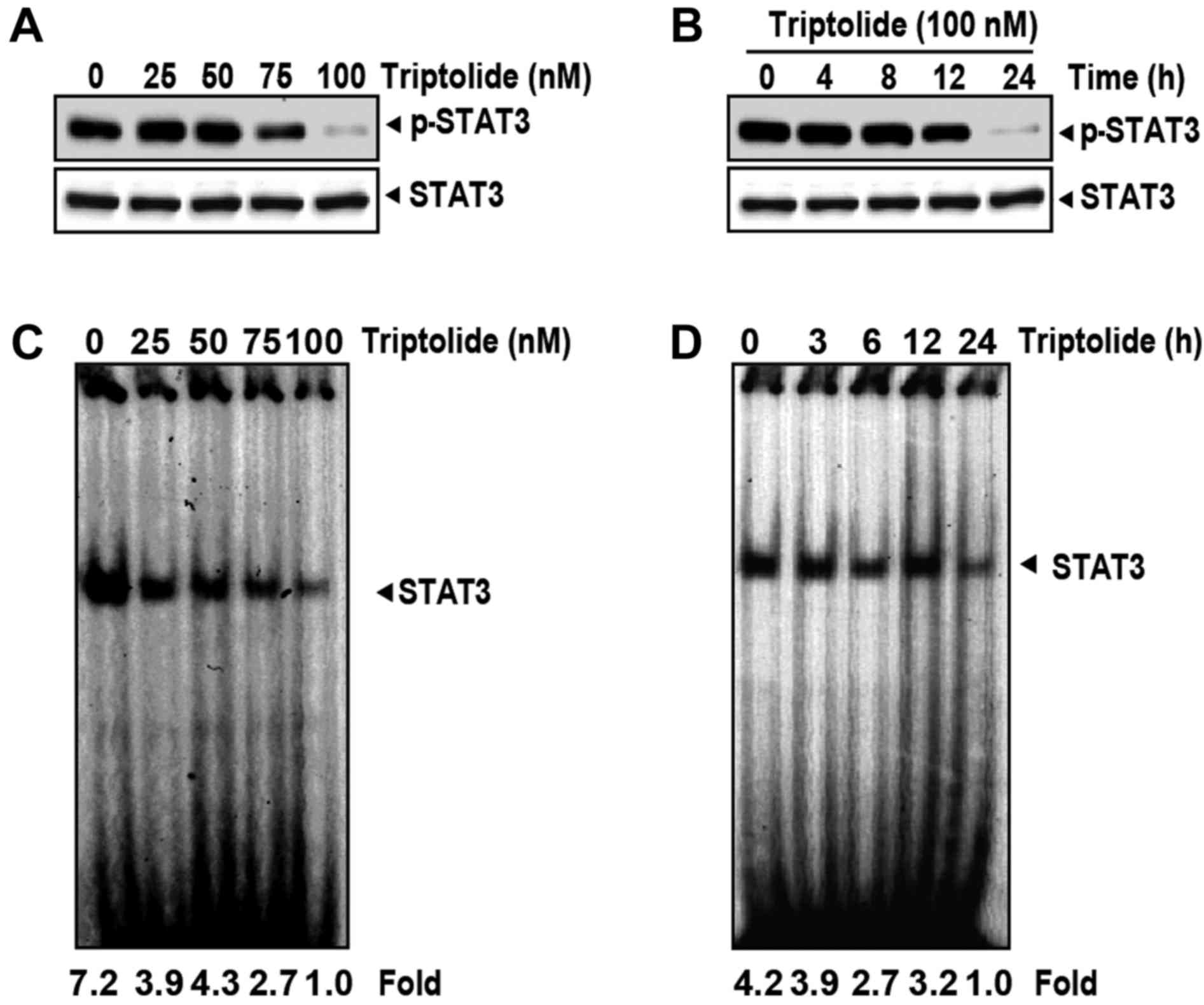
Triptolide downregulates constitutive
STAT3 activation in a dose- and time-dependent manner
Whether triptolide regulates constitutive STAT3
activation in MM cells was examined by determination of STAT3
phosphorylation. Whole-cell extracts were prepared from U266 cells
pretreated for 6 h with various concentrations of triptolide and
STAT3 phosphorylation was examined by western blot analysis using
an anti-p-STAT3 (Tyr705) antibody. Triptolide inhibited the
constitutive activation of STAT3 in U266 cells, with maximum
inhibition at 100 nM (Fig. 2A).
Under these conditions, triptolide had no effect on the STAT3
protein level (Fig. 2A). The
incubation time required for suppression of STAT3 activation by
triptolide was also examined in U266 cells. Triptolide inhibited
the activation of STAT3 in a time-dependent manner, with maximum
inhibition occurring at 24 h, again with no effect on the STAT3
protein level (Fig. 2B).
Triptolide inhibits DNA binding of
STAT3
It was investigated whether triptolide reduces the
DNA-binding activity of STAT3 based on the fact that
phosphorylation of STAT3 regulates gene transcription via its
dimerization, nuclear translocation and DNA binding. An EMSA using
nuclear extracts of U266 cells was performed; the results indicated
that triptolide decreased the DNA-binding activity of STAT3 in a
dose-dependent (Fig. 2C) and
time-dependent (Fig. 2D) manner.
Thus, triptolide is capable of eliminating the DNA-binding ability
of STAT3.
Triptolide suppresses constitutive
activation of Src, JAK1, and JAK2
STAT3 may be constitutively activated by soluble
tyrosine kinases of the Src kinase (34) and JAK (35) families. Therefore, the effects of
triptolide on the constitutive activation of Src kinase, JAK1 and
JAK2 in U266 cells were investigated. Triptolide was found to
suppress the constitutive phosphorylation of JAK1 (Fig. 3A) and JAK2 (Fig. 3B) in a dose-dependent manner,
while the total levels of JAK1 and JAK2 remained unchanged under
the same conditions (Fig. 3A and
B). As shown in Fig. 3C,
triptolide suppressed the constitutive phosphorylation of Src
kinase in a dose-dependent manner, while the total Src kinase
protein levels remained unchanged (Fig. 3C).
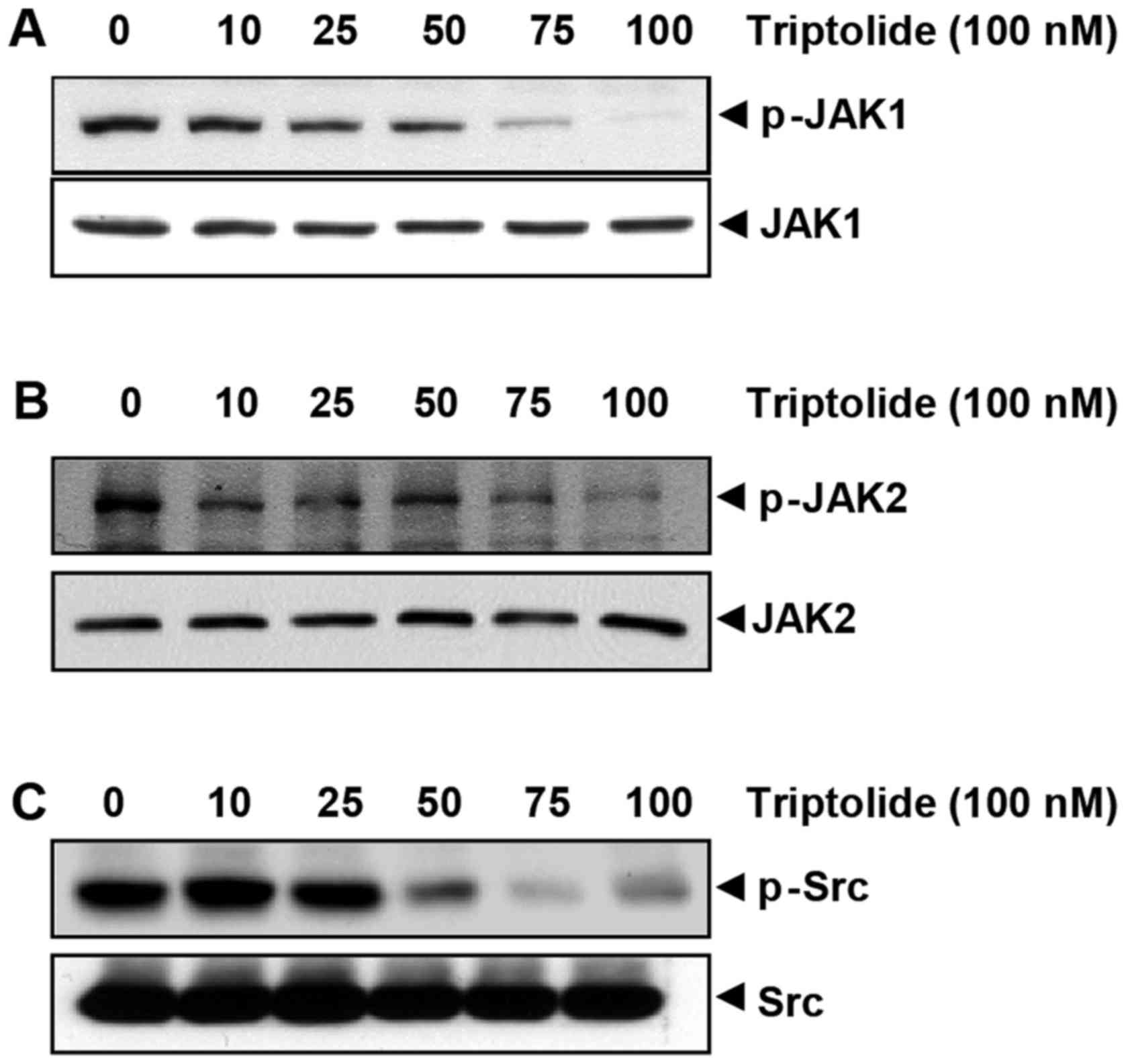 | Figure 3Triptolide downregulates
constitutively active JAK1, JAK2 and Src. Triptolide downregulates
(A) p-JAK1 expression, (B) p-JAK2 expression, and (C) p-Src levels
in a dose-dependent manner. In all experiments, U266 cells
(2×106/ml) were treated with the indicated doses of
triptolide for 24 h, after which whole-cell extracts were prepared,
and 40 μg of those extracts were resolved by SDS-PAGE,
electrotransferred onto nitrocellulose membranes, and probed with
p-JAK1, p-JAK2 and p-Src, respectively. JAK, Janus kinase; p-,
phospho-. |
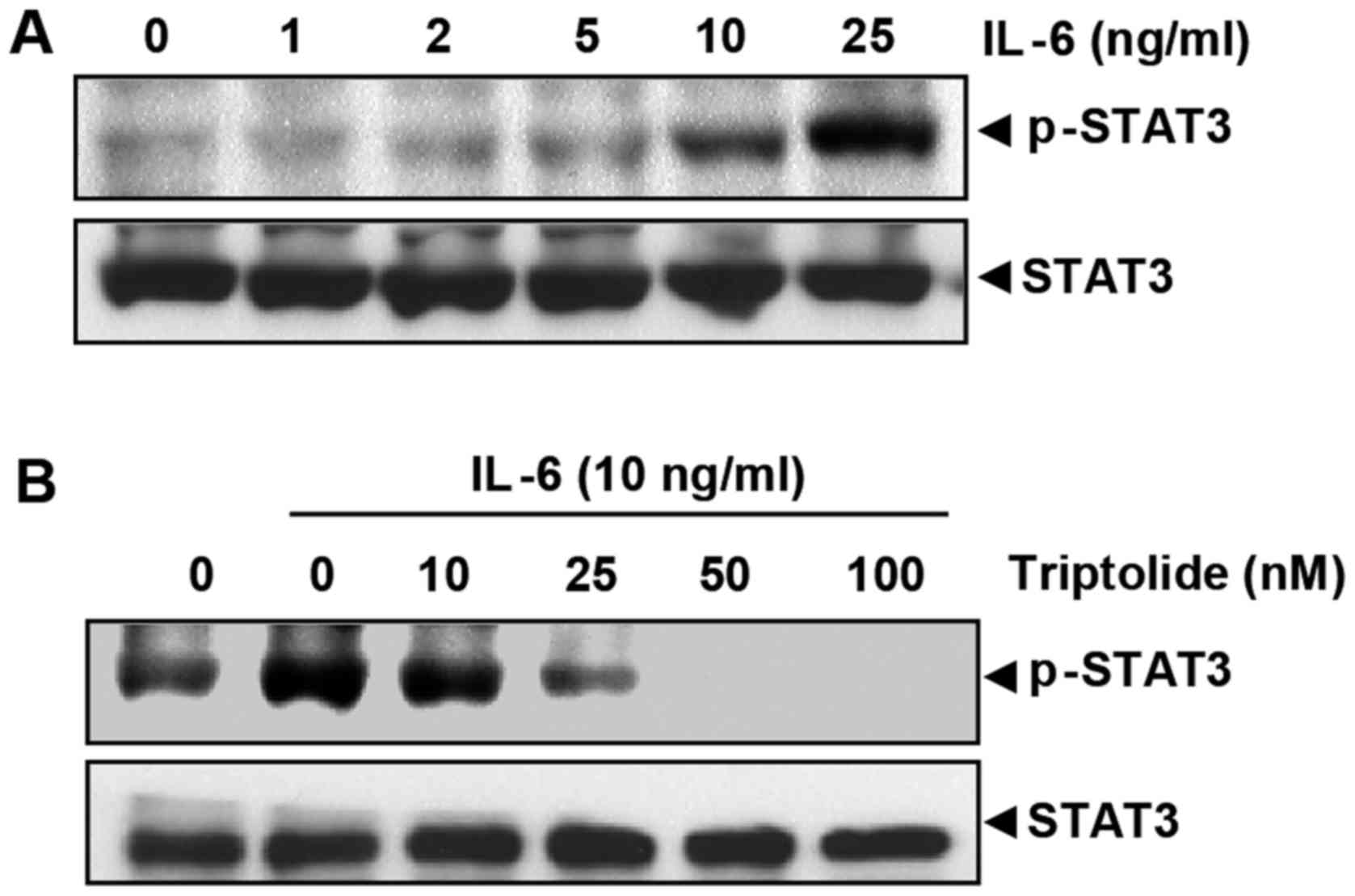
Triptolide inhibits inducible STAT3
phosphorylation in MM cells
Because STAT3 is also activated by phosphorylation
at tyrosine residues in response to IL-6 (27), it was investigated whether
triptolide could inhibit IL-6-induced STAT3 phosphor-ylation in
MM.1S cells, which lack constitutively active STAT3. As shown in
Fig. 4A, IL-6 induced
phosphorylation of STAT3 in a dose-dependent manner. IL-6-induced
STAT3 phosphorylation was suppressed by triptolide in a
dose-dependent manner (Fig. 4B).
A triptolide concentration >50 nM was sufficient to suppress
IL-6-induced STAT3 phosphorylation.
Triptolide induces the expression of
SHP-1
SHP-1, a non-receptor-type protein tyrosine
phosphatase, is known to negatively regulate the STAT3 signaling
pathway (36,37). Therefore, it was examined whether
the inhibition of STAT3 phosphorylation by triptolide was due to
the increased expression of SHP-1. As shown in Fig. 5A, triptolide upregulated SHP-1
expression in a time-dependent manner.
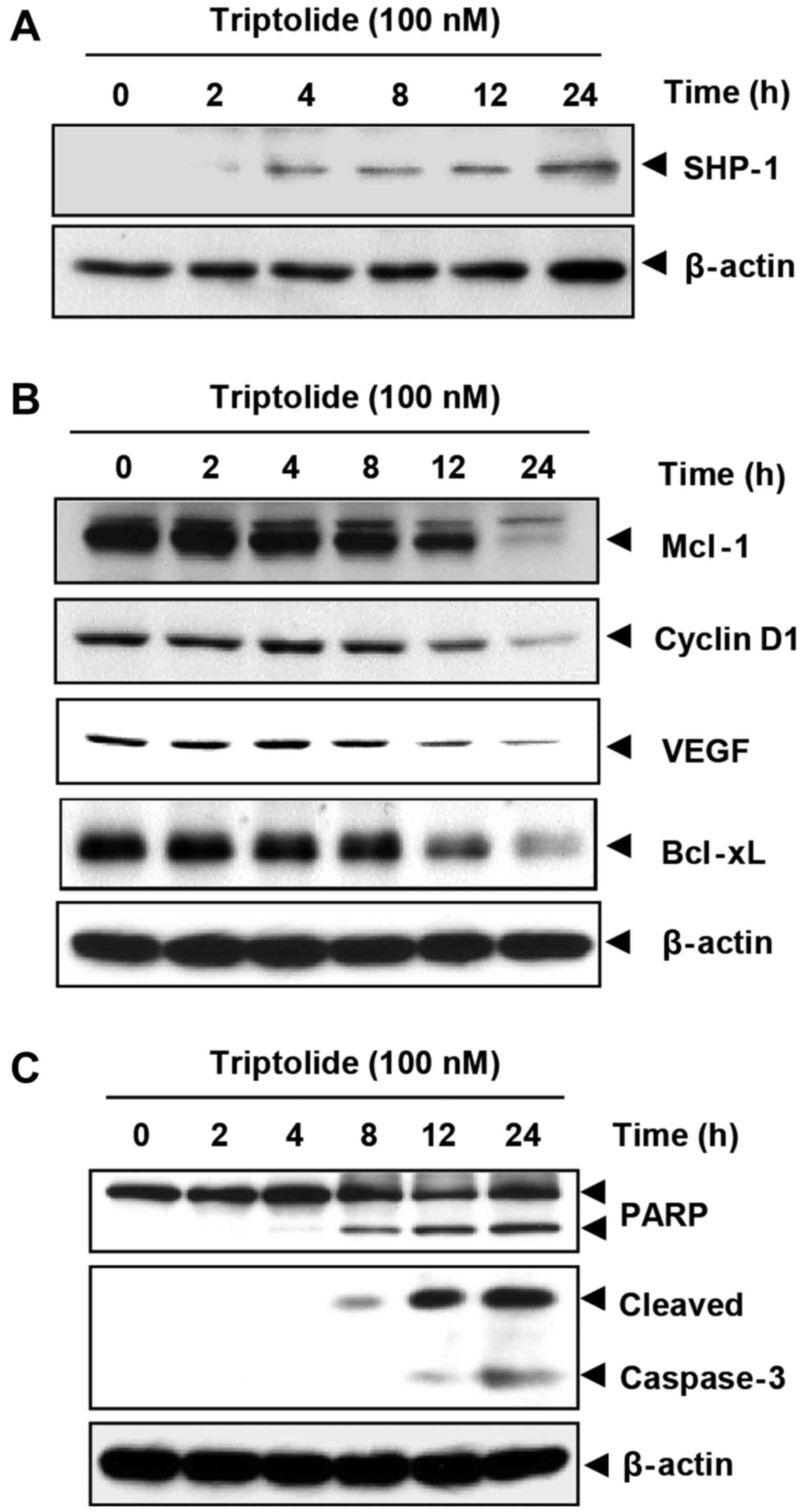 | Figure 5Triptolide upregulates SHP-1 and
downregulates STAT3-regulated antiapoptotic proteins. (A)
Triptolide induces the expression of SHP-1 protein in U266 cells.
U266 cells (2X106/ml) were treated with triptolide (100
nM) for the indicated times. Whole-cell extracts were prepared, and
40 μg of those extracts were resolved by 10% SDS-PAGE,
electrotransferred onto nitrocellulose membranes, and probed with
anti-SHP-1 antibody. (B) Triptolide suppresses STAT3-regulated gene
products. U266 cells (2×106/ml) were treated with 100 nM
triptolide for the indicated time intervals. Whole cell extracts
were prepared, 40 μg of those extracts were resolved by 10%
SDS-PAGE and probed against Mcl-1, cyclin D1, VEGF and Bcl-xL
antibodies. (C) Triptolide induces caspase-3-dependent PARP
cleavage. U266 cells were treated with 100 nM triptolide for the
indicated times, and whole cell extracts were prepared, separated
by SDS-PAGE and subjected to western blot analysis with
anti-caspase-3 and anti-PARP antibodies. In all experiments, the
same blots were stripped and reprobed with anti-β-actin antibody to
verify equal protein loading. SHP-1, protein tyrosine phosphatase
non-receptor type 6; STAT3, signal transducer and activator of
transcription 3; Mcl-1, myeloid cell leukemia-1; VEGF, vascular
endothelial growth factor; PARP, poly(ADP-ribose) polymerase. |
Triptolide downregulates the expression
of genes associated with cell survival, proliferation, and
angiogenesis
The expression of the antiapoptotic proteins Mcl-1
and Bcl-xL, the cell cycle regulator protein cyclin D1, and the
angiogenic protein VEGF, all of which have been reported to be
regulated by STAT3 (25,38), were downregulated by triptolide
treatment (Fig. 5B). Tript olide
downregulated the expression of these proteins in a time-dependent
manner, with maximum suppression at 24 h after the beginning of
treatment (Fig. 5B).
Triptolide causes caspase-3 activation
and PARP cleavage
As caspase-3 is critically involved in the
regulation of antiapoptotic proteins (39) and VEGF (40), it was determined whether
triptolide could activate caspase-3. Treatment of U266 cells with
100 nM triptolide induced caspase-3-dependent cleavage of a 118-kDa
PARP protein into an 87-kDa fragment in a time-dependent manner
(Fig. 5C).
Discussion
The Chinese herb T. wilfordii has been used
for centuries as a traditional medicine for treating fever, chills
and edema. Triptolide, a component extracted from T.
wilfordii, has been intensively researched in recent years as
it exhibits potent antitumor activity in various cancerous cells,
such as human promyelocytic leukemia, T cell lymphoma, human
hepatocellular carcinoma, cervical adenocarcinoma, pancreatic
carcinoma, oral cancer cell and cholangiocarcinoma (41–43). However, triptolide has a serious
drawback as an antitumor agent: Its toxicity (44). For this reason, the mechanism of
action of triptolide in cancerous cells must be determined, as this
would allow its toxicity to be reduced through chemical
modification.
STAT3 is an important signaling pathway that has
been associated with inflammation, survival, proliferation,
chemoresistance, and angiogenesis (26). The results of the present study
indicated that triptolide suppressed the phosphorylation of JAK1
and JAK2, which are upstream regulators of STAT3. Two STAT3
activation pathways were considered: i) IL-6-induced STAT3
activation via the IL-6/JAK/STAT3 cell signaling pathway, in which
IL-6 binding to its receptor leads to phosphorylation of JAK, and
phosphorylated JAK sequentially induces STAT3 phosphorylation; and
ii) constitutive STAT3 activation involving various protein kinases
(26,45,46). The IL-6/JAK/STAT3 signaling
pathway serves a critical role in the growth, invasion and
metastasis of cancerous cells (47). Constitutively active STAT3 is also
important in the survival and proliferation of myeloma, as ~48% of
MM patients have a constitutively active form of STAT3 (48). Regarding the mechanisms of STAT3
inhibition, it was observed in the present study that triptolide
inhibited IL-6-induced and constitutive STAT3 activation in MM
cells. It was also demonstrated that the mechanism of action of
triptolide involves inhibition of induction of SHP-1, as well as
activation of JAK1, JAK2 and c-Src. This is closely associated with
the inhibition of the expression of various STAT3-regulated
proteins and the induction of apoptosis in MM cells.
SHP-1, a negative regulator of the JAK/STAT
signaling pathway, is hypermethylated in 79% of MM patients
(46), and demethylation of SHP-1
decreases STAT3 activation (49).
In the present study, it was demonstrated that triptolide could
induce SHP-1 expression, suggesting that the induction of SHP-1 may
have inhibited STAT3 activation. Abnormal methylation of suppressor
of cytokine signaling 1, another regulator of the JAK/STAT3
signaling pathway, has also been observed in MM patients (50). However, this was not considered in
the present study, as methylation of this gene is not strongly
correlated with the clinical outcome of MM patients (50).
The present study also demonstrated that triptolide
can suppress the activation of Src. Phosphorylation of Src serves a
critical role in tumor cell survival. The majority of
Src-transformed cell lines have persistently activated STAT3 and
dominant-negative STAT3 blocks transformation (51). Furthermore, constitutive STAT3
activation has been implicated in the increased resistance to
apoptosis, which leads to radiation resistance and chemoresistance
(51). This may be mediated by
the expression of the STAT3-regulated gene products including
Bcl-xL, Mcl-1, and cyclin D1. Antiapoptotic proteins such as Bcl-xL
and Mcl-1 inhibit the induction of cell death by various
chemotherapeutic drugs, in parallel with an increase in
chemoresistance (52). The
present study demonstrated that triptolide downregulated Bcl-xL,
Mcl-1 and cyclin D1, which implies that the suppression of STAT3
activation by triptolide may facilitate the apoptosis of MM cells.
It was also found that triptolide downregulated the expression of
VEGF, which is involved in angiogenesis of cancer cells, suggesting
that triptolide may exert an antiangiogenic effect by
downregulating VEGF.
Triptolide was previously reported to inhibit the
transcription factor, NF-κB (21). However, it is unclear whether the
inhibition of NF-κB activation is linked to the inhibition of STAT3
activation by triptolide. NF-κB and STAT3 are activated by
different cytokines; for example, tumor necrosis factor is a major
activator of NF-κB, whereas IL-6 activates STAT3. However, notably,
JAK activation has been reported to regulate both STAT3 and NF-κB
activation (53). Therefore,
triptolide is a potent chemotherapeutic agent targeting JAK
regulation, which regulates NF-κB and STAT3 activation.
Triptolide has been widely used in East Asia for
centuries for the treatment of inflammatory and autoimmune
diseases, and the antitumor activity of triptolide has been
investigated in various cancer cell types. The present study
demonstrated that triptolide inhibits both inducible and
constitutive STAT3 activation in MM cells, suggesting that it
suppresses tumor cell survival, proliferation and angiogenesis.
Although several barriers to the clinical use of triptolide remain
to be overcome, further development of triptolide derivatives and
clinical studies may result in the development of novel anticancer
agents.
Acknowledgments
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF), funded by the Ministry of Education (grant no.
NRF-2016R1A6A1A03011325)
Glossary
Abbreviations
Abbreviations:
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
JAK
|
Janus kinase
|
|
SHP-1
|
protein tyrosine phosphatase
non-receptor type 6
|
|
Mcl-1
|
myeloid cell leukemia-1
|
|
VEGF
|
vascular endothelial growth factor
|
|
IL-6
|
interleukin-6
|
|
PARP
|
poly(ADP-ribose) polymerase
|
|
MM
|
multiple myeloma
|
References
|
1
|
Yang S, Chen J, Guo Z, Xu XM, Wang L, Pei
XF, Yang J, Underhill CB and Zhang L: Triptolide inhibits the
growth and metastasis of solid tumors. Mol Cancer Ther. 2:65–72.
2003.PubMed/NCBI
|
|
2
|
Raab MS, Podar K, Breitkreutz I,
Richardson PG and Anderson KC: Multiple myeloma. Lancet.
374:324–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kumar SK, Dispenzieri A, Lacy MQ, Gertz
MA, Buadi FK, Pandey S, Kapoor P, Dingli D, Hayman SR, Leung N, et
al: Continued improvement in survival in multiple myeloma: changes
in early mortality and outcomes in older patients. Leukemia.
28:1122–1128. 2014. View Article : Google Scholar :
|
|
5
|
Kyle RA and Rajkumar SV: Treatment of
multiple myeloma: a comprehensive review. Clin Lymphoma Myeloma.
9:278–288. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kyle RA: Targeted therapy of multiple
myeloma. Hematology. 17(Suppl 1): 125–128. 2012.
|
|
7
|
Anderson KC: The 39th David A. Karnofsky
Lecture: bench-to- bedside translation of targeted therapies in
multiple myeloma. J Clin Oncol. 30:445–452. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kong LY and Tan RX: Artemisinin, a miracle
of traditional Chinese medicine. Nat Prod Rep. 32:1617–1621. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding GS: Important Chinese herbal
remedies. Clin Ther. 9:345–357. 1987.PubMed/NCBI
|
|
10
|
Chen BJ: Triptolide, a novel
immunosuppressive and anti-inflammatory agent purified from a
Chinese herb Tripterygium wilfordii Hook F. Leuk Lymphoma.
42:253–265. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu R, Li Y, Guo Z, Gong J, Zhu W, Li N and
Li J: Triptolide ameliorates ileocolonic anastomosis inflammation
in IL-10 deficient mice by mechanism involving suppression of
miR-155/SHIP-1 signaling pathway. Mol Immunol. 56:340–346. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wei X, Gong J, Zhu J, Wang P, Li N, Zhu W
and Li J: The suppressive effect of triptolide on chronic colitis
and TNF-alpha/TNFR2 signal pathway in interleukin-10 deficient
mice. Clin Immunol. 129:211–218. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qiu D and Kao PN: Immunosuppressive and
anti-inflammatory mechanisms of triptolide, the principal active
diterpenoid from the Chinese medicinal herb Tripterygium wilfordii
Hook. f. Drugs R D. 4:1–18. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin N, Liu C, Xiao C, Jia H, Imada K, Wu H
and Ito A: Triptolide, a diterpenoid triepoxide, suppresses
inflammation and cartilage destruction in collagen-induced
arthritis mice. Biochem Pharmacol. 73:136–146. 2007. View Article : Google Scholar
|
|
15
|
Zhu W, He S, Li Y, Qiu P, Shu M, Ou Y,
Zhou Y, Leng T, Xie J, Zheng X, et al: Anti-angiogenic activity of
triptolide in anaplastic thyroid carcinoma is mediated by targeting
vascular endothelial and tumor cells. Vascul Pharmacol. 52:46–54.
2010. View Article : Google Scholar
|
|
16
|
Banerjee S, Sangwan V, McGinn O, Chugh R,
Dudeja V, Vickers SM and Saluja AK: Triptolide-induced cell death
in pancreatic cancer is mediated by O-GlcNAc modification of
transcription factor Sp1. J Biol Chem. 288:33927–33938. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Phillips PA, Dudeja V, McCarroll JA,
Borja-Cacho D, Dawra RK, Grizzle WE, Vickers SM and Saluja AK:
Triptolide induces pancreatic cancer cell death via inhibition of
heat shock protein 70. Cancer Res. 67:9407–9416. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu PP, Liu KC, Huang WW, Ma CY, Lin H,
Yang JS and Chung JG: Triptolide induces apoptosis in human adrenal
cancer NCI-H295 cells through a mitochondrial-dependent pathway.
Oncol Rep. 25:551–557. 2011.
|
|
19
|
Zheng Y, Zhang WJ and Wang XM: Triptolide
with potential medicinal value for diseases of the central nervous
system. CNS Neurosci Ther. 19:76–82. 2013. View Article : Google Scholar
|
|
20
|
Zhao F, Chen Y, Li R, Liu Y, Wen L and
Zhang C: Triptolide alters histone H3K9 and H3K27 methylation state
and induces G0/G1 arrest and caspase-dependent apoptosis in
multiple myeloma in vitro. Toxicology. 267:70–79. 2010. View Article : Google Scholar
|
|
21
|
Yinjun L, Jie J and Yungui W: Triptolide
inhibits transcription factor NF-kappaB and induces apoptosis of
multiple myeloma cells. Leuk Res. 29:99–105. 2005. View Article : Google Scholar
|
|
22
|
Chen Q, Lu Z, Jin Y, Wu Y and Pan J:
Triptolide inhibits Jak2 transcription and induces apoptosis in
human myeloproliferative disorder cells bearing Jak2V617F through
caspase-3-mediated cleavage of Mcl-1. Cancer Lett. 291:246–255.
2010. View Article : Google Scholar
|
|
23
|
Tang J, Li ZH, Ge SN, Wang W, Mei XP, Wang
W, Zhang T, Xu LX and Li JL: The inhibition of spinal astrocytic
JAK2-STAT3 pathway activation correlates with the analgesic effects
of triptolide in the rat neuropathic pain model. Evid Based
Complement Alternat Med. 2012:1851672012. View Article : Google Scholar
|
|
24
|
Wang Z, Jin H, Xu R, Mei Q and Fan D:
Triptolide downregulates Rac1 and the JAK/STAT3 pathway and
inhibits colitis-related colon cancer progression. Exp Mol Med.
41:717–727. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aggarwal BB, Sethi G, Ahn KS, Sandur SK,
Pandey MK, Kunnumakkara AB, Sung B and Ichikawa H: Targeting signal
-transducer-and-activator-of-transcription-3 for prevention and
therapy of cancer: modern target but ancient solution. Ann N Y Acad
Sci. 1091:151–169. 2006. View Article : Google Scholar
|
|
26
|
Yu H, Kortylewski M and Pardoll D:
Crosstalk between cancer and immune cells: role of STAT3 in the
tumour microenvironment. Nat Rev Immunol. 7:41–51. 2007. View Article : Google Scholar
|
|
27
|
Zhong Z, Wen Z and Darnell JE Jr: Stat3: a
STAT family member activated by tyrosine phosphorylation in
response to epidermal growth factor and interleukin-6. Science.
264:95–98. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Grandis JR, Drenning SD, Chakraborty A,
Zhou MY, Zeng Q, Pitt AS and Tweardy DJ: Requirement of Stat3 but
not Stat1 activation for epidermal growth factor receptor-mediated
cell growth in vitro. J Clin Invest. 102:1385–1392. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zushi S, Shinomura Y, Kiyohara T, Miyazaki
Y, Kondo S, Sugimachi M, Higashimoto Y, Kanayama S and Matsuzawa Y:
STAT3 mediates the survival signal in oncogenic ras-transfected
intestinal epithelial cells. Int J Cancer. 78:326–330. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shen Y, Devgan G, Darnell JE Jr and
Bromberg JF: Consti tutively activated Stat3 protects fibroblasts
from serum withdrawal and UV-induced apoptosis and antagonizes the
proapoptotic effects of activated Stat1. Proc Natl Acad Sci USA.
98:1543–1548. 2001. View Article : Google Scholar
|
|
31
|
Bhattacharya S, Ray RM and Johnson LR:
STAT3-mediated transcription of Bcl-2, Mcl-1 and c-IAP2 prevents
apoptosis in polyamine-depleted cells. Biochem J. 392:335–344.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Aoki Y, Feldman GM and Tosato G:
Inhibition of STAT3 signaling induces apoptosis and decreases
survivin expression in primary effusion lymphoma. Blood.
101:1535–1542. 2003. View Article : Google Scholar
|
|
33
|
Yu CL, Meyer DJ, Campbell GS, Larner AC,
Carter-Su C, Schwartz J and Jove R: Enhanced DNA-binding activity
of a Stat3-related protein in cells transformed by the Src
oncoprotein. Science. 269:81–83. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schreiner SJ, Schiavone AP and Smithgall
TE: Activation of STAT3 by the Src family kinase Hck requires a
functional SH3 domain. J Biol Chem. 277:45680–45687. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ihle JN: STATs: signal transducers and
activators of transcription. Cell. 84:331–334. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chim CS, Fung TK, Cheung WC, Liang R and
Kwong YL: SOCS1 and SHP1 hypermethylation in multiple myeloma:
implications for epigenetic activation of the Jak/STAT pathway.
Blood. 103:4630–4635. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Oka T, Ouchida M, Koyama M, Ogama Y,
Takada S, Nakatani Y, Tanaka T, Yoshino T, Hayashi K, Ohara N, et
al: Gene silencing of the tyrosine phosphatase SHP1 gene by
aberrant methylation in leukemias/lymphomas. Cancer Res.
62:6390–6394. 2002.PubMed/NCBI
|
|
38
|
Yu H and Jove R: The STATs of cancer - new
molecular targets come of age. Nat Rev Cancer. 4:97–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Weinmann P, Gaehtgens P and Walzog B:
Bcl-Xl- and Bax-alpha-mediated regulation of apoptosis of human
neutrophils via caspase-3. Blood. 93:3106–3115. 1999.PubMed/NCBI
|
|
40
|
Jin K, Mao XO, Batteur SP, McEachron E,
Leahy A and Greenberg DA: Caspase-3 and the regulation of hypoxic
neuronal death by vascular endothelial growth factor. Neuroscience.
108:351–358. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chan EW, Cheng SC, Sin FW and Xie Y:
Triptolide induced cytotoxic effects on human promyelocytic
leukemia, T cell lymphoma and human hepatocellular carcinoma cell
lines. Toxicol Lett. 122:81–87. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen YW, Lin GJ, Chia WT, Lin CK, Chuang
YP and Sytwu HK: Triptolide exerts anti-tumor effect on oral cancer
and KB cells in vitro and in vivo. Oral Oncol. 45:562–568. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tengchaisri T, Chawengkirttikul R,
Rachaphaew N, Reutrakul V, Sangsuwan R and Sirisinha S: Antitumor
activity of triptolide against cholangiocarcinoma growth in vitro
and in hamsters. Cancer Lett. 133:169–175. 1998. View Article : Google Scholar
|
|
44
|
Pyatt DW, Yang Y, Mehos B, Le A, Stillman
W and Irons RD: Hematotoxicity of the chinese herbal medicine
Tripterygium wilfordii Hook f in CD34-positive human bone marrow
cells. Mol Pharmacol. 57:512–518. 2000.PubMed/NCBI
|
|
45
|
Schindler C and Darnell JE Jr:
Transcriptional responses to polypeptide ligands: the JAK-STAT
pathway. Annu Rev Biochem. 64:621–651. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hodge DR, Hurt EM and Farrar WL: The role
of IL-6 and STAT3 in inflammation and cancer. Eur J Cancer.
41:2502–2512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chang Q, Bournazou E, Sansone P, Berishaj
M, Gao SP, Daly L, Wels J, Theilen T, Granitto S, Zhang X, et al:
The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and
metastasis. Neoplasia. 15:848–862. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Quintanilla-Martinez L, Kremer M, Specht
K, Calzada-Wack J, Nathrath M, Schaich R, Höfler H and Fend F:
Analysis of signal transducer and activator of transcription 3
(Stat 3) pathway in multiple myeloma: Stat 3 activation and cyclin
D1 dysregulation are mutually exclusive events. Am J Pathol.
162:1449–1461. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Q, Raghunath PN, Vonderheid E, Odum
N and Wasik MA: Lack of phosphotyrosine phosphatase SHP-1
expression in malignant T-cell lymphoma cells results from
methylation of the SHP-1 promoter. Am J Pathol. 157:1137–1146.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Depil S, Saudemont A and Quesnel B: SOCS-1
gene methylation is frequent but does not appear to have prognostic
value in patients with multiple myeloma. Leukemia. 17:1678–1679.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Catlett-Falcone R, Landowski TH, Oshiro
MM, Turkson J, Levitzki A, Savino R, Ciliberto G, Moscinski L,
Fernández-Luna JL, Nuñez G, et al: Constitutive activation of Stat3
signaling confers resistance to apoptosis in human U266 myeloma
cells. Immunity. 10:105–115. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Heere-Ress E, Thallinger C, Lucas T,
Schlagbauer-Wadl H, Wacheck V, Monia BP, Wolff K, Pehamberger H and
Jansen B: Bcl-X(L) is a chemoresistance factor in human melanoma
cells that can be inhibited by antisense therapy. Int J Cancer.
99:29–34. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Digicaylioglu M and Lipton SA:
Erythropoietin-mediated neuro-protection involves cross-talk
between Jak2 and NF-kappaB signalling cascades. Nature.
412:641–647. 2001. View Article : Google Scholar : PubMed/NCBI
|