1. Introduction
Cell-penetrating peptides (CPPs), also known as
protein transduction domains (PTDs) or Trojan peptides, are
peptides of diverse structure and physicochemical properties. They
share two common features. First, they are short peptides
containing 5–30 amino acids (1).
Furthermore, they have the ability to cross the cell membrane,
which is not restricted by their covalent or noncovalent binding of
molecules such as DNA (2), RNA
(3), antisense oligonucleotides
(4), plasmids (5), liposomes (6), proteins (3) or nanoparticles (7). The cellular uptake of CPPs may occur
via energy-dependent or -independent mechanisms (1). Due to their low cytotoxicity and
immunogenicity, CPPs have been applied in in vitro (8) and in vivo (9) tests with good results. At present,
promising results have been obtained primarily for their topical
application, while little information on the effects of intravenous
administration is currently available. However, predictions are not
favorable due to the lack of selectivity of the action of peptides,
which may lead to penetration of the cell membrane components of
healthy tissue (10).
The discovery of the first CPP was made in 1988 by
two independent research groups - Frankel and Pabo (11) and Green and Loewenstein (12). This peptide was derived from the
human immunodeficiency virus-1 (HIV-1) TAT protein, a
transcriptional transactivator that is essential for HIV-1
replication (13). In 1994,
Derossi et al (14)
reported that a Drosophila homeobox protein fragment shares
the same properties. These studies initiated research into the use
of proteins as vectors capable of efficient transport of molecules
into the cell and led to the recognition of numerous novel
molecules with similar characteristics.
2. Classification of CPPs
Based on their origin, CPPs may be divided into
three groups: i) Naturally occurring peptides produced by living
organisms, ii) chimeric peptides, which are modified natural
proteins and iii) synthetic peptides entirely designed and
synthesized in the laboratory (15). Specific examples are listed in
Table I (11,12,16–20).
 | Table IExamples of cell-penetrating peptides
classified on the basis of their origin with indication of their
sequence and key feature. |
Table I
Examples of cell-penetrating peptides
classified on the basis of their origin with indication of their
sequence and key feature.
| Protein group | Protein name | Sequence |
Characteristics | Refs. |
|---|
| Natural | TAT | GRKKRRQRRRPPQ | Transcriptional
regulator of HIV. | (11,12) |
| pVEC |
LLIILRRRIRKQAHAHSK | Mouse's catherin
sequence. | (16) |
| Chimeric | Transportan |
GWTLNSAGYLLGKINLKALAALAKKIL | Protein formed by
the combination of neuropeptide galanin and wasp's botulinum toxin,
mastoparan, through a lysine residue. | (17) |
| MPG |
GALFLGFLGAAGSTMGAWSQPKKKRKV | Protein obtained by
the fusion of the transmembrane glycoprotein of HIV, gp41, with
SV40 virus T-antigen. | (18) |
| Synthetic | MAP |
KLALKLALKALKAALKLA | Amphipathic protein
created de novo from lysine, arginine and leucine
residues. | (19) |
|
R6W3 | RRWWRRWRR | Artificial peptide
created de novo based on the structure of penetrin. | (20) |
Based on their physicochemical properties, CPPs may
be also divided into three groups as follows: i) Cationic peptides
with an overall positive charge, ii) hydrophobic protein with a
high content of hydrophobic amino acids such as alanine, methionine
or valine and iii) amphipathic proteins, which contain a
hydrophobic as well as a hydrophilic fragment (15). Examples for peptides in this
classification system are listed in Table II (14,21–25).
| Table IIExamples of cell-penetrating peptides
classified on the basis of their physicochemical properties with
indication of their sequence and key features. |
Table II
Examples of cell-penetrating peptides
classified on the basis of their physicochemical properties with
indication of their sequence and key features.
| Protein group | Protein name | Sequence |
Characteristics | Refs. |
|---|
| Cationic | R9 | RRRRRRRRR | Synthetically
created sequence of nine arginines. | (21) |
| Antp |
RQIKIWFQNRRMKWKK | Homeobox gene of
Drosophila melanogaster, which determines the development of
the morphological differences between the segment of head and torso
of the insect. | (14) |
| Hydrophobic | VP22 |
DAATATRGRSAASRPTERPRAPARSASRPRRVD | A component of a
capsid of HSV-1 virus. | (22) |
| K-FGF | AAVLLPVLLAAP | Artificial peptide
containing the penetrating motif and locating the cell nucleus
sequence. | (23) |
| Amphipathic | VT5 |
DPKGDPKGVTVTVTVTVTGKGDPKPD | Capsid protein of
rotaviruses | (24) |
| SynB1 |
RGGRLSYSRRRFSTSTGR | The peptide derived
from protegrin. It has the ability to cross the blood-brain
barrier. | (25) |
Although hundreds of CPPs have already been
described in literature, only a few of them are recently used in
commercial applications due to insufficient knowledge regarding
their properties and mechanism of uptake into cells (26).
3. Methods for producing CPP-cargo
conjugates
There are two main approaches to introduce CPP-cargo
bonds. The first involves the creation of covalent bonds, while the
second one comprises the formation of non-covalent bonds. Even
though covalent bonding of CPP and cargo is more widespread, it has
several disadvantages. First, the chemical reaction, which occurs
during the formation of the bond, may modify the properties of the
cargo or CPP (27). Furthermore,
this method cannot be used to bind plasmids. The solution may be
introduction of non-covalent bonds that have been successfully used
in the case of combining nucleic acid and protein molecules with
short amphipathic penetrating peptides such as MPG (28) or Pep-1 (29). The formation of non-covalent bonds
is based on the creation of electrostatic and hydrophobic
interactions, which eliminates the risk of changing the biological
activity of the bonded molecule by chemical reactions and enables
the delivery of plasmid molecules (30). In addition, the non-covalent
strategy is simpler in execution, since it only requires incubation
of the protein with the transported molecule. It also appears to be
an effective and safe means of multiple cargo delivery (31). The effectiveness of interaction
between CPP and cargo depends on the peptide/cargo charge ratio,
which is the proportion of moles of positively charged amino acid
groups in the CPP to those of phosphate ones in the cargo (Fig. 1). In general, a negative to
positive (N/P) ratio that is too high results in precipitation or
formation of larger particles whose capacity to enter cells is
poor, while an N/P ratio that is too low is not effective due to
the small capacity of cell internalization (31).
4. Mechanism of internalization of CPPs
Despite numerous years of research, the detailed
mechanisms of CPP internalization remain to be fully elucidated
(15). According to various
mechanistic models, it appears that several factors influencing the
intake: i) Cell type, ii) mass and structure of the cargo, iii)
method of binding, iv) concentration of CPP and v) incubation time
and temperature. The diversity in the structure of the
cell-penetrating proteins also results in differences in the manner
in which the CPP is taken and its effectiveness (32).
Endocytosis and direct translocation are probably
the major mechanisms of CPPs entering cells. Endocytosis may occur
according to two mechanisms (Fig.
2). The first mechanism is the clathrin-dependent way, which
involves the coating of transported molecules by polymerized
clathrin. The second mechanism, known as the clathrin-independent
mechanism, does not require the presence of clathrin and proceeds
through macropinocytosis or via caveolae (33).
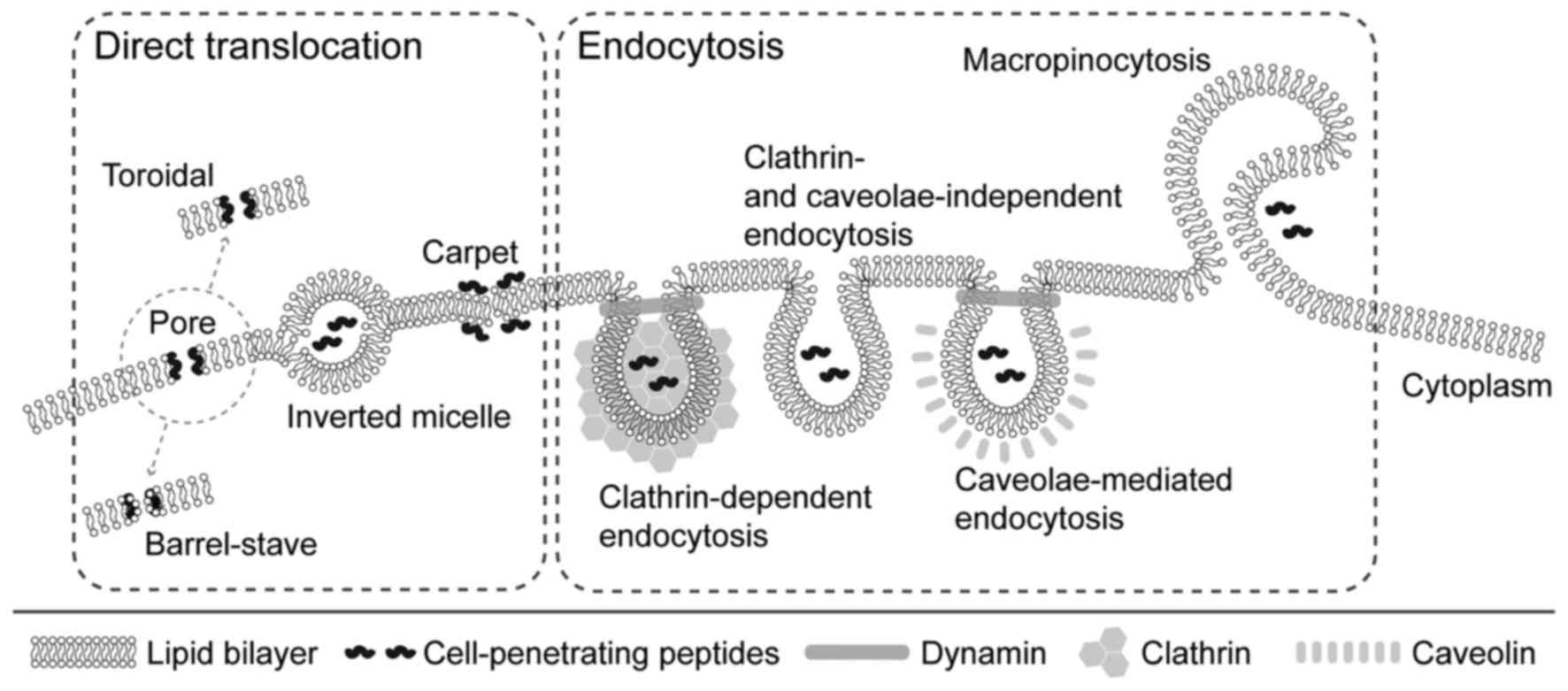 | Figure 2Mechanisms of cellular
internalization of CPPs. Different mechanisms of cellular uptake of
CPPs have been proposed. These mechanisms include direct
translocation and endocytosis. Several models have been proposed
for direct translocation: i) Formation of transient pores (the
toroidal pore model, where CPPs interact with polar groups of
membrane phospholipids, and the barrel stave model, where CPPs
assume an amphipathic α-helix structure when inserted into the
cellular membrane); ii) inverted micelles, where CPPs disturb the
lipid bilayer, leading to the formation of inverted hexagonal
structures, and iii) the carpet model, where CPPs transiently
destabilize the cellular membrane by their association to its
surface, leading to the reorganization of phospholipids. Uptake of
CPPs through the cell membrane was demonstrated to proceed via
several endocytotic pathways: i) Clathrin-dependent, ii) clathrin-
and caveolae-independent, and iii) caveolae-mediated. In addition,
CPPs may be internalized by macropinocytosis. CPPs,
cell-penetrating peptides. |
Direct translocation, as an energy-independent
transport method, is the third of the proposed models. Its validity
was confirmed under conditions that prevented endocytosis, e.g.
using endocytosis inhibitors (34) and low temperature (35). Despite these factors, effective
CPP internalization has been observed (36). The process begins with the
interaction of positively charged amino acid residues of unfolded
penetrating protein with membrane phospholipids, which interferes
with standard interactions between cell membrane components and
allows for cell penetration. The protein is then folded again by
chaperone action (37). Three
alternative mechanisms of direct translocation across the lipid
bilayer have also been proposed as follows: i) Inverted micelles,
ii) carpet model and iii) pore formation. An inverted micelles
variant was described by Derossi et al (16) based on the results of nuclear
magnetic resonance imaging of penethrin internalization. This
hypothetical mechanism starts with the interaction of cationic
amino acid residues with negatively charged phospholipid groups,
but in this case, it leads to the production of micelles by the
phospholipids which are enclosing the peptide. The presence of
hydrophobic amino acids in the peptide is also required for this
process. The micelle then reopens inside the cell and releases the
protein (38). In the carpet
model, internalization takes place in three stages. The first
involves interactions of cationic residues of proteins with
phospholipids, leading to changes in membrane structure. In the
next step, a rotation of the peptide occurs, enabling the
interaction of hydrophobic protein residues with hydrophobic tails
of phospholipids. Finally, CPP penetrates the membrane by the
slight disturbances caused during the second stage. It is also a
mechanism by which certain antibacterial substances, including
magainins, exert their toxic effects (39). The third model comprises pore
formation, which may occur due to creation of bonds between the
hydrophobic part of an amphipathic α-helical penetrating peptide
with the lipid part of the membrane. As a result, the hydrophilic
residues produce pores that allow this molecule to penetrate the
membrane (40).
So far, it has not been determined which of these
models represent the true processes of CPP uptake. It is also
likely that conditions of the cellular environment affect the way
in which CPPs internalize the cell. It is possible that several
modes of internalization occur at the same time (32).
5. Applications of CPPs
CPPs are widely used in studies on methods for
transporting therapeutic particles through the cell membrane.
Numerous studies have confirmed their potential in vitro and
in vivo (10,41). Using CPPs, molecules including
proteins (3), liposomes (6) and nanoparticles (7) have already been introduced into
cells with satisfying results. However, CPPs may also be used for
the internalization of nucleic acids (2,4,5).
After numerous successful preclinical studies, certain CPPs,
including TAT and its conjugates, have entered phase-I, phase-II or
even phase-III clinical trials. Data obtained from these studies
proved that the use of CPPs for clinical therapy is possible, as
they are well tolerated and directed to intracellular targets. So
far, penetrating peptides have been used in clinical trials,
including those on the treatment of cardiovascular diseases
(42), pain (43–45), hearing loss (46) and even facial wrinkles (47). One of the first phase-III clinical
trials that reached completion included the use of TAT in
combination with a c-Jun N-terminal kinase inhibitor. In 2012, a
TAT-coupled dextrogyre peptide inhibiting the c-Jun N-terminal
kinase, named XG-102, successfully passed a phase-I clinical trial,
which was designed to determine the safety of its use, and did not
produce any adverse effects other than those induced by the placebo
(48). The phase-III trial was
aimed at assessing the ability of the substance to inhibit
intraocular inflammation and reduce pain in patients undergoing
cataract surgery. That phase was completed in 2015 and demonstrated
the absence of anterior chamber cells and pain (49). Another member of the c-Jun
N-terminal kinase inhibitor family conjugated with TAT has been
used in a study on the treatment of hearing loss. After the success
of the phase-I trial (46),
compound AM-111 created by Auris Medical is currently being
investigated in two phase-III clinical trials (50,51).
Transport of nucleic acids
Nucleic acid molecules such as small interfering RNA
(siRNA), antisense oligonucleotides, decoy DNA or plasmids are
widely used in the gene therapy of numerous diseases. The current
drawbacks include low efficiency of cell uptake and low
bioavailability of the therapeutic nucleic acid molecules alone
(52). These inconveniences may
be overcome by the use of penetrating proteins (53).
Numerous available CPPs have been used to transport
siRNA particles; these include MPG (28), transportan (54) and TAT (55). Penetrating peptides may be
covalently or non-covalently conjugated with siRNA. The covalent
approach is suitable to provide only one siRNA particle conjugated
to one peptide particle. However, there are several problems
associated with this method. First, the bonding is required to be
reversible within the cell's environment. Furthermore, cationic
CPPs form non-covalent bonds with siRNA and the large aggregates
are created involuntarily (27).
In addition, the efficacy of the uptake is insufficient, probably
due to neutralization of the positive charge of the cationic
peptide, which is necessary to disrupt cell membrane, by the
negative charge of the siRNA (30). The problem of aggregation may be
overcome by addition of denaturing agent, e.g. formamide (56). Only a few studies on CPP, siRNA
conjugates have been published. The most widely used method of
preparation is the chemical synthesis of disulfide-linked CPP
conjugates of oligonucleotides, the bond of which is impermanent in
the reducing environment of the cytosol. Muratovska and Eccles
(57) confirmed that delivery of
siRNA against luciferase or enhanced green fluorescent protein
(EGFP) mRNA fused with penetrin or transportan via a disulfide bond
reduced the expression of these genes in transfected cell lines
more efficiently than Lipofectamine/siRNA complex. The same type of
bond was successfully used by Davidson et al (58). They demonstrated the introduction
of siRNA-targeting caspases and superoxide dismutase into primary
hippocampal neuronal cells using penetratin 1. This study was
particularly important because of the small selection of tools for
introducing molecules into highly specialized neuronal cells
(58). One problem with the above
studies is the lack of a purification step for conjugates prepared
by a non-specific oxidation method (56). Another approach was applied by
Chiu et al (59). They
used a stable thiolmaleimide linkage and also purification of
conjugates by denaturing gels or reverse-phase high-performance
liquid chromatography. The study demonstrated that TAT 47–57
peptide or TAT 47–57-derived oligocarbamate and siRNA against EGFP
or endogenous cyclin-dependent kinase 9 inhibited the expression of
reporter as well as endogenous genes with satisfying results, but
again, as indicated by Turner et al (60), the efficiency of the purification
process was not evaluated. Uptake of the conjugates was similar to
that obtained by the application of Lipofectamine (59). The purification step may also be
performed by using anion exchange chromatography with addition of a
denaturing agent (60).
Although covalent conjugation of CPP to cargo has
proven to be effective, a much simpler and equally effective method
is available: Non-covalent binding, which includes formation of
electrostatic and hydrophobic interactions between cargo and CPP.
This approach has several advantages. First, it is easy to use, as
it only requires simple incubation of penetrating peptide and
cargo. In addition, there is no chemical reaction, and therefore,
there is no risk of any undesirable cargo structure or activity
modification (61). While
covalent conjugates appear to cross the membrane via an endosomal
pathway, the mechanism of internalization of non-covalent complexes
still remains to be elucidated (62). The most widely used class of CPPs
for non-covalent binding of siRNA is an amphipathic class, which
includes MPG (63), Pep-1
(64) or CADY (62). The MPG-8 protein, which is a
shorter version of MPG, was used to deliver siRNA against cyclin B1
mRNA in a murine model of human prostate tumor (63). Cyclin B1 is responsible for
cyclin-dependent kinase 1 activation. The delivery of siRNA against
cyclin B1 conjugated to MPG-8 led to the blockage of tumor cell
proliferation as a result of cell cycle arrest in G2 phase
(63). siRNA is also an effective
tool to inhibit HIV-1 replication within infected cells, but this
requires an effective delivery system. It was proven that a
chimeric peptide composed of CPP as an RNA-binding domain and cell
fusion peptide domain combined with siRNA entered the cells and
inhibited HIV-1 replication (65).
CPPs are not only of use for inhibiting the
expression of genes by delivering siRNA, but also for introducing
additional genetic material, e.g. plasmid DNA. The widely used CPP
Arg nonapeptide (R9) was demonstrated to be a more efficient tool
for plasmid delivery than the commercially available transfection
reagent TurboFect (66). Another
study also reported that TAT is capable of transferring plasmids
into cells (67).
CPPs in genome editing
In addition to positive or negative regulation of
gene products, CPPs may be used for genome editing. Cre
recombinase, zinc-finger nucleases (ZFNs), transcription
activator-like effector nucleases (TALENs) and clustered regularly
interspaced short palindromic repeats (CRISPR)/Cas9 are currently
the most widespread tools for genome engineering. CPPs may be used
to deliver all of them into cells (68).
The Cre/locus of X(cross)-over in P1 (LoxP) system
is a promising tool for editing specific DNA sequences. The
cleavage location is determined by the short LoxP sequences below
and above the removed sequence in reverse orientation. The reaction
of cleavage is catalyzed by Cre recombinase derived from
bacteriophage P. This tool is attractive, as insertion of Cre
recombinase under the control of an inducible promoter may allow
the site-specific excretion of a particular gene. However, the
problem with this method is the selection of an appropriate
Cre/LoxP delivery system (69).
The first successful attempt to introduce Cre recombinase into the
cell using penetrating peptides was made in 2001 by Jo et al
(70). They used purified
recombinant fusion proteins bearing the 12 amino acid membrane
translocation sequence (MTS) from Kaposi fibroblast growth factor
to deliver enzymatically active Cre proteins directly into NIH3T3
and S4R murine embryonic stem cells with a single loxP-modified
sulfonylurea receptor gene, and the Tex.loxP.EG cell line was
generated by Cre-mediated recombination activating the expression
of a GFP reporter gene following delivery via this system. Of the
four recombinant proteins investigated glutathione-S-transferase
(GST)-Cre-MTS, GST-nuclear localization signal (NLS)-Cre-MTS,
maltose-binding protein-NLS-Cre-MTS and His6-NLS-Cre-MTS), the best
result was obtained using His6-NLS-Cre-MTS. Recombination was
induced in 82% of Tex.loxp.EG cells following three consecutive 2-h
treatments with 10 µM His6-NLS-Cre-MTS. In the same study, the
in vivo activity of His6-NLS-Cre-MTS was tested in ROSA26R
mice. ROSA26R is a transgenic mouse strain in which a
β-galactosidase reporter gene is activated by Cre-mediated
recombination. It was demonstrated that recombination induced by
His6-NLS-Cre-MTS occurred in ~50% of splenocytes cultured in the
absence of lipopolysaccharide. In addition, in mice injected with
His6-NLS-Cre-MTS once a day for three days, Cre-mediated
recombination without any side effect was observed in all examined
tissues, including the brain, heart, kidney, lung, spleen and liver
(70). In another study, reporter
T cells were also used to determine the mechanism of TAT-Cre
internalization. Despite the strong binding of TAT to the cell
surface, indicating a direct penetration mechanism, the protein was
revealed to enter the cell via multistep mechanism. Wadia et
al (71) suggested that TAT
CPP-mediated cellular entry occurs by interaction with lipid rafts
in a receptor-independent manner and subsequent rapid
internalization by macropinocytosis, followed by a pH drop and
disintegration of the lipid bilayer of the macropinosome, resulting
in cargo release from TAT into the cytosol (71). Furthermore, Hashimoto et al
(72) reported that Cre-mediated
recombination of genomic targets takes places principally during
the S-phase of the cell cycle. This is probably due to the
relaxation of the chromatin structure during S-phase, which
enhances the attachment of Cre to the DNA molecule and increases
the Cre/loxP recombination reaction. Xu et al (73) have confirmed the safety of Cre
recombinase in combination with TAT, as there were no significant
changes in cell proliferation or the karyotype of transgenic goat
fibroblasts after incubation with TAT-Cre. In addition, TAT-Cre did
not alter the developmental competence of embryos reconstructed by
nuclear transfer from TAT-Cre-transduced cells. Furthermore, they
obtained two live transgenic goats with no evident abnormalities in
development or behavior for at least 3 months after birth (73).
The CPP and Cre conjugates have been successfully
applied to numerous cell lines, including HeLa, Rp250 and HEK-283
(74), and also in organotypic
cultures derived from adult mice and embryos carrying a reporter
gene flanked by LoxP sequences (75). In vivo studies
demonstrating their usefulness are also available. Sonsteng et
al (76) proved that Cre
fused with TAT enters hepatocytes of mice following tail vein
injection. For this purpose, two types of mice were used. The first
one was ROSAmT-mG, whose cells exhibit red membrane
fluorescence in the absence of Cre or strong membrane expression of
EGFP in the presence of Cre. The second one was
ROSAnT-nG, which differs from ROSAmT-mG in
nuclear localization of red- and GFP in the absence or presence of
Cre, respectively. They demonstrated that after TAT-Cre
administration, 5–20% of hepatocytes were converted, which provided
a more effective conversion of centrolobular hepatocytes than in
mice hydrodynamically inoculated with pMC1-Cre plasmid.
Recombination occurred only in hepatocytes, and was not observed in
endothelial other non-hepatocyte cells in the liver or in any other
organs. However, in animals inoculated with TAT-Cre cells
exhibiting green fluorescence were identified near the injection
site in the tail (76).
Although in engineered site-directed nucleases, such
as ZFNs, delivery of components into the cells is not a significant
problem, as it has been proven that purified ZFNs have the ability
to be internalized by cells independently (77); however, the use of CPPs enhances
the uptake efficiency (78). The
major problems with using ZFNs are their cytotoxicity and
off-target effects (79). These
may also be potentially reduced by using CPP (80). So far, CPP-ZFN conjugates have
been applied in approaches towards treating breast cancer (81) and malaria (82). CPPs conjugated to mammalian target
of rapamycin (mTOR)-specific ZFN have been proposed as a novel
therapeutic approach to disrupt the gene function of mTOR (81). The phosphoinositide-3
kinase/Akt/mTOR pathway is essential for growth and proliferation
of breast cancer cells. It has been suggested that elimination of
the effect of mTOR in sustaining the survival of cancerous cells
may inhibit this pathway and result in slower tumor growth. ZFN-CPP
has also been proposed as a safe drug which may help to eliminate
parasites from infected cells (82). However, only an in vitro
study is currently available.
Introducing transcription activator-like effector
nucleases (TALENs) into cells is slightly more challenging due to
reasons including its molecular weight. TALENs do not contain the
Cys2-His2 zinc-finger domain, which allows for direct penetration
of the cell membrane by ZFNs (83). For this purpose, CPPs have already
been adapted with satisfying results in in vitro studies and
they are a promising alternative to current methods for the
delivery of TALENs into mammalian cells. Liu et al (83) demonstrated that TALEN conjugated
to poly-Arg peptides enters the cells and, without any obvious
toxicity, is able to induce gene knockout in human cell lines. They
proved that HeLa cells incubated for 2 h with R9-conjugated C-C
chemokine receptor type 5 (CCR5) targeting TALEN protein resulted
in CCR5 gene disruption at a frequency that was three times higher
than that achieved by transient transfection of TALEN expression
vectors, while the process did not cause any significant decrease
in cell viability. The reaction was most effective with
peptide/protein ratios of 8:1 and 15:1. At lower ratios, R9 was
probably unable to penetrate the cell membrane efficiently, whereas
at higher peptide/protein ratios, no complete breakdown of bonds
between CPP and cargo occurred and these ratios were not effective
during internalization of TALEN designed to knock down the bone
morphogenetic protein receptor type IA gene in the HEK293 cell line
(83). This suggests that
different types of these nucleases require different
peptide-to-protein ratios. However, the penetrating abilities of
penetrin, transportan and human growth hormone-1 connected with
TALEN were not observed. Another CPP with proven efficacy in TALEN
delivery was TAT. It was used for disruption of the CCR5 gene in
HeLa cells and human induced pluripotent stem cells under
hypothermic conditions (83).
CRISPR, which is associated with nucleases from Cas
family, naturally occurs as a part of the adaptive immune system
against bacteria and archaea. The effect of CRISPR is based on
storing fragments of exogenous DNA harvested, e.g. during
bacteriophage infection, from the bacterial genome for quick
recognition and destruction of DNA molecules containing this
fragment by nucleases in case of further infection (84). This method, as an RNA-guided way
of DNA cutting, has been successfully applied in the genome
modification of bacteria (85),
cell cultures (3), animals
(86) and plants (87). Derived from the CRISPR/Cas system,
RNA-guided nuclease consists of two major components - Cas9 protein
and guide RNA (gRNA). Efficient delivery of these two types of
particle is necessary for gene editing and these components may be
introduced into the cell using lentiviral and adeno-associated
viral systems, which have the potential for random integration of
vector sequence into the host genome, leading to undesirable
changes (88,89). For this reason, a virus- and
plasmid-free approach has been developed. Cas9 and gRNA have been
delivered separately as CPP-conjugated Cas9 and CPP-complexed gRNA.
This approach led to a lesser degree of off-target mutations and
immune responses in the host, and also induced lower cytotoxicity
(89). Ramakrishna et al
(3) reported on the preparation
of a modified sequence of the Cas9 gene, its ligation within
plasmid pET-28a(+), introduction of the plasmid into Escherichia
coli BL21, and isolation and purification of the resulting
protein. The modified Cas9 protein contained cysteine at the
N-terminus, which allowed for the attachment of
4-maleimidobutyryl-4G9R4L (m9R) via a thioether bond between the
free SH residue of C-terminal cysteine of Cas9 and the primary
amine residue (−NH2)of m9R, whereas the single guide RNA
(sgRNA)-9R complex was formed by incubation of the components at
room temperature for 30 min (3).
In addition, the introduction of sgRNA as a complex with CPP may
potentially enable the simultaneous introduction of several of its
type, and thus the system's functioning at several locations in the
genome.
6. Toxicity of CPPs
The cytotoxic effects of peptides depend on their
amino acid sequence, secondary structure and summary charge
(90). Abnormalities that may be
caused by CPPs include cytoplasmic leakages due to changes in
membrane permeability, and dysfunction in membrane proteins
resulting from interactions with amphipathic CPPs (91). Cellular membrane permeability was
examined by measuring the leakage of lactate dehydrogenase (LDH) in
several cell lines, K562 (erythroblastic leukemia), MDA-MB-231
(breast cancer) and immortalized endothelial cells from an aorta
(91). Saar et al
(91) reported that incubation
with the model amphipathic protein and trasportan 10 resulted in a
significant increase of LDH leakage in the two tumor cell lines but
did not produce any statistically significant changes in the
endothelial cell line. In addition, cationic peptides, including
penetrin, an 18 amino acid CPP derived from murine vascular
endothelial-cadherin (pVEC) and TAT, did not induce any
cytotoxicity (91). The
mechanisms of the selective action of amphipathic CPPs on tumor
cells have remained to be elucidated. However, they appear to be a
good choice for the treatment of oncological patients. Of note,
none of the proteins caused any significant hemolysis, which
confirms the relative safety of their use (91).
In general, in vitro and in vivo
experiments demonstrated that CPPs did not induce any significant
cytotoxic effect, even at high concentrations (92). The behavior, appearance and eating
habits of the animals were examined prior to and after
administration of various doses of CPP-phosphorodiamidate
morpholino oligomer conjugates (93). The body weight and serum levels of
urea were also measured. In rats receiving the conjugate at 15
mg/kg body weight, no changes in any of the parameters were
observed. In those receiving 30 mg/kg body weight, a slight
decrease in body weight and an increase in urea levels in the serum
were observed, but these changes were not health-threatening. No
dose lower than 150 mg/kg body weight caused any weight loss or
significant deterioration of any biochemical indicators.
Approximately 50% mortality of animals occurred in the groups
receiving 210 and 250 mg/kg and 100% mortality was observed in
those receiving 400 mg/kg body weight of the conjugate. These
studies also confirm the safety of the use of CPP, as no
significant adverse effects were noticeable at doses sufficient for
therapeutic purposes (≤15 mg/kg) (93). The toxicity of CPPs may also be
affected by the presence of the cargo. However, Maiolo et al
(94) demonstrated that free R7
and R7W exhibited slightly higher toxicity than those associated
with cargo.
7. Conclusion
Gene modification systems, e.g. Cre recombinase,
ZFNs, TALENs or CRISPRs, provide the possibility of effective
treatment of diseases that have been so far considered as
incurable. One of the problems with using these methods is to
identify an appropriate system for their transport through the cell
membrane. Viral delivery systems, in spite of being most effective,
generate abnormalities such as immunogenicity or cytotoxicity.
Another disadvantage is the limited size of the insert. On the
other hand, non-viral systems, including electroporation or
liposomes, do not cause any immune reactions, and within certain
limits, the size of the transported molecule has no effect;
however, they also have certain disadvantages. The major
disadvantage is a lower transduction efficiency (95,96). The lack of an ideal tool for
introducing molecules into cells motivated researchers to search
for novel and improved methods. In this light, CPPs were discovered
as a promising alternative to current methods. As a non-viral
system, CPP-cargo complexes are characterized by safety of use,
while maintaining a relatively high efficiency. For this reason,
they are used in genome editing studies with better results than
those achieved for other non-viral methods (57,66). However, certain problems remain to
be solved prior to any clinical trials being performed on delivery
of genome modification systems by CPPs, especially after promising
trials with other cargoes. First of all, the mechanisms by which
CPPs penetrate the cell membrane requires to be fully elucidated.
It would also be useful to determine the impact of CPPs on
organisms after intravenous administration, most results available
were obtained by studies applying local administration. Further
research on the selection of suitable conditions for the production
of conjugates or complexes of CPP and cargo, as well as appropriate
cell incubation protocols should be performed to fully exploit the
potential of CPPs and to achieve maximum efficiencies.
Acknowledgments
This study was supported by a grant from the
National Science Centre, Poland (grant no. 2015/17/D/NZ7/00809 to
M.G.).
References
|
1
|
Zorko M and Langel U: Cell-penetrating
peptides: Mechanism and kinetics of cargo delivery. Adv Drug Deliv
Rev. 57:529–545. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karro K, Männik T, Männik A and Ustav M:
DNA transfer into animal cells using stearylated CPP based
transfection reagent. Methods Mol Biol. 1324:435–445. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ramakrishna S, Kwaku Dad A-BB, Beloor J,
Gopalappa R, Lee SK and Kim H: Gene disruption by cell-penetrating
peptide-mediated delivery of Cas9 protein and guide RNA. Genome
Res. 24:1020–1027. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zatsepin TS, Turner JJ, Oretskaya TS and
Gait MJ: Conjugates of oligonucleotides and analogues with cell
penetrating peptides as gene silencing agents. Curr Pharm Des.
11:3639–3654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Segovia N, Dosta P, Cascante A, Ramos V
and Borrós S: Oligopeptide-terminated poly(β-amino ester)s for
highly efficient gene delivery and intracellular localization. Acta
Biomater. 10:2147–2158. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu C, Luo Q, Tu Y, Wang G, Liu Y and Xie
Y: Drug-carrier interaction analysis in the cell penetrating
peptide-modified liposomes for doxorubicin loading. J
Microencapsul. 32:745–754. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Y, Wen G, Wang D, Zhang X, Lu Y, Wang
J, Zhong L, Cai H, Zhang X and Wang Y: A complementary strategy for
enhancement of nanoparticle intracellular uptake. Pharm Res.
31:2054–2064. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jo J, Hong S, Choi WY and Lee DR:
Cell-penetrating peptide (CPP)-conjugated proteins is an efficient
tool for manipulation of human mesenchymal stromal cells. Sci Rep.
4:43782014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khafagy S, Morishita M, Isowa K, Imai J
and Takayama K: Effect of cell-penetrating peptides on the nasal
absorption of insulin. J Control Release. 133:103–108. 2009.
View Article : Google Scholar
|
|
10
|
Skotland T, Iversen TG, Torgersen ML and
Sandvig K: Cell-penetrating peptides: Possibilities and challenges
for drug delivery in vitro and in vivo. Molecules. 20:13313–13323.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Frankel AD and Pabo CO: Cellular uptake of
the tat protein from human immunodeficiency virus. Cell.
55:1189–1193. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Green M and Loewenstein PM: Autonomous
functional domains of chemically synthesized human immunodeficiency
virus tat trans-activator protein. Cell. 55:1179–1188. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ruben S, Perkins A, Purcell R, Joung K,
Sia R, Burghoff R, Haseltine WA and Rosen CA: Structural and
functional characterization of human immunodeficiency virus tat
protein. J Virol. 63:1–8. 1989.PubMed/NCBI
|
|
14
|
Derossi D, Joliot AH, Chassaing G and
Prochiantz A: The third helix of the Antennapedia homeodomain
translocates through biological membranes. J Biol Chem.
269:10444–10450. 1994.PubMed/NCBI
|
|
15
|
Milletti F: Cell-penetrating peptides:
Classes, origin, and current landscape. Drug Discov Today.
17:850–860. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Derossi D, Chassaing G and Prochiantz A:
Trojan peptides: The penetratin system for intracellular delivery.
Trends Cell Biol. 8:84–87. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pooga M, Hällbrink M, Zorko M and Langel
U: Cell penetration by transportan. FASEB J. 12:67–77.
1998.PubMed/NCBI
|
|
18
|
Kwon SJ, Han K, Jung S, Lee JE, Park S,
Cheon YP and Lim HJ: Transduction of the MPG-tagged fusion protein
into mammalian cells and oocytes depends on amiloride-sensitive
endocytic pathway. BMC Biotechnol. 9:732009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mo RH, Zaro JL and Shen WC: Comparison of
cationic and amphipathic cell penetrating peptides for siRNA
delivery and efficacy. Mol Pharm. 9:299–309. 2012. View Article : Google Scholar :
|
|
20
|
Bechara C, Pallerla M, Burlina F, Illien
F, Cribier S and Sagan S: Massive glycosaminoglycan-dependent entry
of Trp-containing cell-penetrating peptides induced by exogenous
sphingomyelinase or cholesterol depletion. Cell Mol Life Sci.
72:809–820. 2015. View Article : Google Scholar
|
|
21
|
Melikov K, Hara A, Yamoah K, Zaitseva E,
Zaitsev E and Chernomordik LV: Efficient entry of cell-penetrating
peptide nona-arginine into adherent cells involves a transient
increase in intracellular calcium. Biochem J. 471:221–230. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zavaglia D, Favrot MC, Eymin B, Tenaud C
and Coll JL: Intercellular trafficking and enhanced in vivo
antitumour activity of a non-virally delivered P27-VP22 fusion
protein. Gene Ther. 10:314–325. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin YZ, Yao SY, Veach RA, Torgerson TR and
Hawiger J: Inhibition of nuclear translocation of transcription
factor NF-kappa B by a synthetic peptide containing a cell
membrane-permeable motif and nuclear localization sequence. J Biol
Chem. 270:14255–14258. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oehlke J, Krause E, Wiesner B, Beyermann M
and Bienert M: Extensive cellular uptake into endothelial cells of
an amphipathic beta-sheet forming peptide. FEBS Lett. 415:196–199.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rousselle C, Clair P, Temsamani J and
Scherrmann JM: Improved brain delivery of benzylpenicillin with a
peptide-vector-mediated strategy. J Drug Target. 10:309–315. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Järver P and Langel U: Cell-penetrating
peptides - a brief introduction. Biochim Biophys Acta.
1758:260–263. 2006. View Article : Google Scholar
|
|
27
|
Meade BR and Dowdy SF: Exogenous siRNA
delivery using peptide transduction domains/cell penetrating
peptides. Adv Drug Deliv Rev. 59:134–140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Simeoni F, Morris MC, Heitz F and Divita
G: Insight into the mechanism of the peptide-based gene delivery
system MPG: Implications for delivery of siRNA into mammalian
cells. Nucleic Acids Res. 31:2717–2724. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muñoz-Morris MA, Heitz F, Divita G and
Morris MC: The peptide carrier Pep-1 forms biologically efficient
nanoparticle complexes. Biochem Biophys Res Commun. 355:877–882.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gros E, Deshayes S, Morris MC,
Aldrian-Herrada G, Depollier J, Heitz F and Divita G: A
non-covalent peptide-based strategy for protein and peptide nucleic
acid transduction. Biochim Biophys Acta. 1758:384–393. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Deshayes S, Morris M, Heitz F and Divita
G: Delivery of proteins and nucleic acids using a non-covalent
peptide-based strategy. Adv Drug Deliv Rev. 60:537–547. 2008.
View Article : Google Scholar
|
|
32
|
Guo Z, Peng H, Kang J and Sun D:
Cell-penetrating peptides: Possible transduction mechanisms and
therapeutic applications. Biomed Rep. 4:528–534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mäger I, Langel K, Lehto T, Eiríksdóttir E
and Langel U: The role of endocytosis on the uptake kinetics of
luciferin-conjugated cell-penetrating peptides. Biochim Biophys
Acta. 1818:502–511. 2012. View Article : Google Scholar
|
|
34
|
Dutta D and Donaldson JG: Search for
inhibitors of endocytosis: Intended specificity and unintended
consequences. Cell Logist. 2:203–208. 2012. View Article : Google Scholar
|
|
35
|
Tomoda H, Kishimoto Y and Lee YC:
Temperature effect on endocytosis and exocytosis by rabbit alveolar
macrophages. J Biol Chem. 264:15445–15450. 1989.PubMed/NCBI
|
|
36
|
Bode SA1, Thévenin M, Bechara C, Sagan S,
Bregant S, Lavielle S, Chassaing G and Burlina F: Self-assembling
mini cell-penetrating peptides enter by both direct translocation
and glycosaminoglycan-dependent endocytosis. Chem Commun (Camb).
48:7179–7181. 2012. View Article : Google Scholar
|
|
37
|
Cleal K, He L, Watson PD and Jones AT:
Endocytosis, intracellular traffic and fate of cell penetrating
peptide based conjugates and nanoparticles. Curr Pharm Des.
19:2878–2894. 2013. View Article : Google Scholar
|
|
38
|
Derossi D, Calvet S, Trembleau A,
Brunissen A, Chassaing G and Prochiantz A: Cell internalization of
the third helix of the Antennapedia homeodomain is
receptor-independent. J Biol Chem. 271:18188–18193. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Matsuzaki K, Sugishita K and Miyajima K:
Interactions of an antimicrobial peptide, magainin 2, with
lipopolysaccharide-containing liposomes as a model for outer
membranes of gram-negative bacteria. FEBS Lett. 449:221–224. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Deshayes S, Plénat T, Aldrian-Herrada G,
Divita G, Le Grimellec C and Heitz F: Primary amphipathic
cell-penetrating peptides: Structural requirements and interactions
with model membranes. Biochemistry. 43:7698–7706. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Regberg J, Eriksson JN and Langel U:
Cell-penetrating peptides: From cell cultures to in vivo
applications. Front Biosci (Elite Ed). 5:509–516. 2013. View Article : Google Scholar
|
|
42
|
Safety and efficacy study of AVI-5126 when
used on vein grafts before use in heart by-pass graft surgery
(CABG). https://clinicaltrials.gov/ct2/show/NCT00451256.
2009
|
|
43
|
Safety and efficacy study of KAI-1678 to
treat pain in subjects with postherpetic neuralgia. https://clinicaltrials.gov/ct2/show/NCT01106716.
2010
|
|
44
|
Safety and efficacy study of KAI-1678 to
treat pain in subjects with spinal cord injury. https://clinicaltrials.gov/ct2/show/NCT01135108.
2010
|
|
45
|
Safety and efficacy study of KAI-1678 to
treat subjects with postoperative pain. https://clinicaltrials.gov/ct2/show/NCT01015235.
2011
|
|
46
|
Efficacy of AM-111 in patients with acute
sensorineural hearing loss. https://clinicaltrials.gov/ct2/show/NCT00802425.
2014
|
|
47
|
Safety and efficacy study of RT001 to
treat moderate to severe lateral canthal lines. https://clinicaltrials.gov/ct2/show/NCT00888914.
2013
|
|
48
|
Safety, tolerability and PK of a single iv
infusion of 10, 40, and 80 µg/kg XG-102 administered to healthy
volunteers. https://clinicaltrials.gov/ct2/show/NCT01570205.
2012
|
|
49
|
Efficacy and safety of XG-102 in reduction
of post-cataract surgery intraocular inflammation. https://clinicaltrials.gov/ct2/show/NCT02235272.
2015
|
|
50
|
AM-111 in the treatment of acute inner ear
hearing loss (HEALOS). https://clinicaltrials.gov/ct2/show/NCT02561091.
2014
|
|
51
|
Efficacy and safety of AM-111 as acute
sudden sensorineural hearing loss treatment (ASSENT). https://clinicaltrials.gov/ct2/show/NCT02809118.
2017
|
|
52
|
Bakhtiyari S, Haghani K, Basati G and
Karimfar MH: siRNA therapeutics in the treatment of diseases. Ther
Deliv. 4:45–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nakase I, Tanaka G and Futaki S:
Cell-penetrating peptides (CPPs) as a vector for the delivery of
siRNAs into cells. Mol Biosyst. 9:855–861. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wierzbicki PM, Kogut-Wierzbicka M,
Ruczynski J, Siedlecka-Kroplewska K, Kaszubowska L, Rybarczyk A,
Alenowicz M, Rekowski P and Kmiec Z: Protein and siRNA delivery by
transportan and transportan 10 into colorectal cancer cell lines.
Folia Histochem Cytobiol. 52:270–280. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Moschos SA, Jones SW, Perry MM, Williams
AE, Erjefalt JS, Turner JJ, Barnes PJ, Sproat BS, Gait MJ and
Lindsay MA: Lung delivery studies using siRNA conjugated to
TAT(48–60) and penetratin reveal peptide induced reduction in gene
expression and induction of innate immunity. Bioconjug Chem.
18:1450–1459. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Turner JJ, Jones S, Fabani MM, Ivanova G,
Arzumanov AA and Gait MJ: RNA targeting with peptide conjugates of
oligonucleotides, siRNA and PNA. Blood Cells Mol Dis. 38:1–7. 2007.
View Article : Google Scholar
|
|
57
|
Muratovska A and Eccles MR: Conjugate for
efficient delivery of short interfering RNA (siRNA) into mammalian
cells. FEBS Lett. 558:63–68. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Davidson TJ, Harel S, Arboleda VA, Prunell
GF, Shelanski ML, Greene LA and Troy CM: Highly efficient small
interfering RNA delivery to primary mammalian neurons induces
MicroRNA-like effects before mRNA degradation. J Neurosci.
24:10040–10046. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chiu YL, Ali A, Chu CY, Cao H and Rana TM:
Visualizing a correlation between siRNA localization, cellular
uptake, and RNAi in living cells. Chem Biol. 11:1165–1175. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Turner JJ, Williams D, Owen D and Gait MJ:
Disulfide conjugation of peptides to oligonucleotides and their
analogs. Current protocols in nucleic acid chemistry. Chapter 4:
Unit 4.28. 2006. View Article : Google Scholar
|
|
61
|
Huang YW, Lee HJ, Tolliver LM and Aronstam
RS: Delivery of nucleic acids and nanomaterials by cell-penetrating
peptides: Opportunities and challenges. Biomed Res Int.
2015:8340792015.PubMed/NCBI
|
|
62
|
Crowet JM, Lins L, Deshayes S, Divita G,
Morris M, Brasseur R and Thomas A: Modeling of non-covalent
complexes of the cell-penetrating peptide CADY and its siRNA cargo.
Biochim Biophys Acta. 1828:499–509. 2013. View Article : Google Scholar
|
|
63
|
Crombez L, Morris MC, Dufort S,
Aldrian-Herrada G, Nguyen Q, Mc Master G, Coll JL, Heitz F and
Divita G: Targeting cyclin B1 through peptide-based delivery of
siRNA prevents tumour growth. Nucleic Acids Res. 37:4559–4569.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kadkhodayan S, Jafarzade BS, Sadat SM,
Motevalli F, Agi E and Bolhassani A: Combination of cell
penetrating peptides and heterologous DNA prime/protein boost
strategy enhances immune responses against HIV-1 Nef antigen in
BALB/c mouse model. Immunol Lett. 188:38–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Bivalkar-Mehla S, Mehla R and Chauhan A:
Chimeric peptide-mediated siRNA transduction to inhibit HIV-1
infection. J Drug Target. 25:307–319. 2017. View Article : Google Scholar
|
|
66
|
Kato T, Yamashita H, Misawa T, Nishida K,
Kurihara M, Tanaka M, Demizu Y and Oba M: Plasmid DNA delivery by
arginine-rich cell-penetrating peptides containing unnatural amino
acids. Bioorg Med Chem. 24:2681–2687. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Rudolph C, Plank C, Lausier J, Schillinger
U, Müller RH and Rosenecker J: Oligomers of the arginine-rich motif
of the HIV-1 TAT protein are capable of transferring plasmid DNA
into cells. J Biol Chem. 278:11411–11418. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Rádis-Baptista G, Campelo IS, Morlighem
JR, Melo LM and Freitas VJ: Cell-penetrating peptides (CPPs): From
delivery of nucleic acids and antigens to transduction of
engineered nucleases for application in transgenesis. J Biotechnol.
252:15–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Nagy A: Cre recombinase: The universal
reagent for genome tailoring. Genesis. 26:99–109. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Jo D, Nashabi A, Doxsee C, Lin Q, Unutmaz
D, Chen J and Ruley HE: Epigenetic regulation of gene structure and
function with a cell-permeable Cre recombinase. Nat Biotechnol.
19:929–933. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wadia JS, Stan RV and Dowdy SF:
Transducible TAT-HA fusogenic peptide enhances escape of TAT-fusion
proteins after lipid raft macropinocytosis. Nat Med. 10:310–315.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hashimoto M, Taniguchi M, Yoshino S, Arai
S and Sato K: S Phase-preferential Cre-recombination in mammalian
cells revealed by HIV-TAT-PTD-mediated protein transduction. J
Biochem. 143:87–95. 2008. View Article : Google Scholar
|
|
73
|
Xu Y, Liu S, Yu G, Chen J, Chen J, Xu X,
Wu Y, Zhang A, Dowdy SF and Cheng G: Excision of selectable genes
from transgenic goat cells by a protein transducible TAT-Cre
recombinase. Gene. 419:70–74. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
De Coupade C, Fittipaldi A, Chagnas V,
Michel M, Carlier S, Tasciotti E, Darmon A, Ravel D, Kearsey J,
Giacca M, et al: Novel human-derived cell-penetrating peptides for
specific subcellular delivery of therapeutic biomolecules. Biochem
J. 390:407–418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Gitton Y, Tibaldi L, Dupont E, Levi G and
Joliot A: Efficient CPP-mediated Cre protein delivery to developing
and adult CNS tissues. BMC Biotechnol. 9:402009. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Sonsteng KM, Prigge JR, Talago EA, June RK
and Schmidt EE: Hydrodynamic delivery of Cre protein to
lineage-mark or time-stamp mouse hepatocytes in situ. PLoS One.
9:e912192014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Gaj T, Guo J, Kato Y, Sirk SJ and Barbas
CF III: Targeted gene knockout by direct delivery of zinc-finger
nuclease proteins. Nat Methods. 9:805–807. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chen Z, Jaafar L, Agyekum DG, Xiao H, Wade
MF, Kumaran RI, Spector DL, Bao G, Porteus MH, Dynan WS, et al:
Receptor-mediated delivery of engineered nucleases for genome
modification. Nucleic Acids Res. 41:e1822013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cornu TI, Thibodeau-Beganny S, Guhl E,
Alwin S, Eichtinger M, Joung JK and Cathomen T: DNA-binding
specificity is a major determinant of the activity and toxicity of
zinc-finger nucleases. Mol Ther. 16:352–358. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Pruett-Miller SM, Reading DW, Porter SN
and Porteus MH: Attenuation of zinc finger nuclease toxicity by
small-molecule regulation of protein levels. PLoS Genet.
5:e10003762009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Puria R, Sahi S and Nain V:
HER2+ breast cancer therapy: By CPP-ZFN mediated
targeting of mTOR? Technol Cancer Res Treat. 11:175–180. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Nain V, Sahi S and Verma A: CPP-ZFN: A
potential DNA-targeting anti-malarial drug. Malar J. 9:2582010.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Liu J, Gaj T, Patterson JT, Sirk SJ and
Barbas CF III: Cell-penetrating peptide-mediated delivery of TALEN
proteins via bioconjugation for genome engineering. PLoS One.
9:e857552014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Horvath P and Barrangou R: CRISPR/Cas, the
immune system of bacteria and archaea. Science. 327:167–170. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Selle K and Barrangou R: Harnessing
CRISPR-Cas systems for bacterial genome editing. Trends Microbiol.
23:225–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Harms DW, Quadros RM, Seruggia D, Ohtsuka
M, Takahashi G, Montoliu L and Gurumurthy CB: Mouse genome editing
using the CRISPR/Cas system. Curr Protoc Hum Genet.
83:15.7.1–15.7.27. 2014. View Article : Google Scholar
|
|
87
|
Bortesi L and Fischer R: The CRISPR/Cas9
system for plant genome editing and beyond. Biotechnol Adv.
33:41–52. 2015. View Article : Google Scholar
|
|
88
|
Park A, Hong P, Won ST, Thibault PA,
Vigant F, Oguntuyo KY, Taft JD and Lee B: Sendai virus, an RNA
virus with no risk of genomic integration, delivers CRISPR/Cas9 for
efficient gene editing. Mol Ther Methods Clin Dev. 3:160572016.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Suresh B, Ramakrishna S and Kim H:
Cell-penetrating peptide-mediated delivery of Cas9 protein and
guide RNA for genome editing. Methods Mol Biol. 1507:81–94. 2017.
View Article : Google Scholar
|
|
90
|
Langel U: Handbook of Cell-Penetrating
Peptides. 2nd edition. Taylor and Francis Group; 2006, View Article : Google Scholar
|
|
91
|
Saar K, Lindgren M, Hansen M, Eiríksdóttir
E, Jiang Y, Rosenthal-Aizman K, Sassian M and Langel U:
Cell-penetrating peptides: A comparative membrane toxicity study.
Anal Biochem. 345:55–65. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Suhorutsenko J, Oskolkov N, Arukuusk P,
Kurrikoff K, Eriste E, Copolovici DM and Langel U: Cell-penetrating
peptides, PepFects, show no evidence of toxicity and immunogenicity
in vitro and in vivo. Bioconjug Chem. 22:2255–2262. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Amantana A, Moulton HM, Cate ML, Reddy MT,
Whitehead T, Hassinger JN, Youngblood DS and Iversen PL:
Pharmacokinetics, biodistribution, stability and toxicity of a
cell-penetrating peptide-morpholino oligomer conjugate. Bioconjug
Chem. 18:1325–1331. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Maiolo JR, Ferrer M and Ottinger EA:
Effects of cargo molecules on the cellular uptake of arginine-rich
cell-penetrating peptides. Biochim Biophys Acta. 1712:161–172.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Cox DB, Platt RJ and Zhang F: Therapeutic
genome editing: Prospects and challenges. Nat Med. 21:121–131.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yin H, Song CQ, Dorkin JR, Zhu LJ, Li Y,
Wu Q, Park A, Yang J, Suresh S, Bizhanova A, et al: Therapeutic
genome editing by combined viral and non-viral delivery of CRISPR
system components in vivo. Nat Biotechnol. 34:328–333. 2016.
View Article : Google Scholar : PubMed/NCBI
|