Introduction
Osteoarthritis (OA) is the most common age-related
degenerative disease. It is a metabolically dynamic process that
involves all joint structures (loss of cartilage matrix,
inflammation of synovium, formation of osteophytes, sclerosis of
subchondral bone) (1,2). These pathologies lead to chronic
joint pain, limited movement and eventually disability.
Osteoarthritis to varying degrees happened in an approximate 10–15%
of adults >60 all around the world (3). The cost of OA is estimated to make
up of 0.50% of a country's gross domestic product (4). Currently, no agent has been approved
for OA management by the FDA or any other agencies worldwide.
Available drugs provide only temporarily symptomatic relief but
most involve numerous side effects. Therefore, there is a dire need
for effective and widely available approaches in OA management,
which is partly hindered by our limited understanding of the
pathomechanisms of OA.
In diarthrodial joints, articular cartilage serves a
critical weight-bearing role. Chondrocytes, the unique cells in
articular cartilage, account for maintaining the balance of
degradation and synthesis of extracellular matrix (ECM) (5). In progression of articular cartilage
degradation in OA, an increased synthesis of pro-inflammatory
cytokines such as interleukin-1β (IL-1β), matrix metalloproteinases
(MMPs) and cyclooxygenase-2 (COX-2) results in loss of the major
components of the ECM (6).
Chondrocyte death, as a consequence of direct damage as well as
apoptotic cell death, may be a result of the lack of certain ECM
components bounding to the cell surface, which in turn implicate
cartilage matrix degradation and further joint inflammation
(7). It is known that IL-1β is
one of the major cytokines in the pathogenesis of OA (8), which inhibits the synthesis of ECM
proteins and induces the releases of MMPs in chondrocytes (9). In particular, IL-1β also has been
shown to induce the nuclear factor-κB (NF-κB) signaling pathway in
chondrocytes, which was considered as the key molecular pathway in
progression of cartilage degradation in OA (10). The NF-κB molecules not only could
enhance the articular damage through promote secretion of many
degradative enzymes, including MMP-1, MMP-2 and MMP-3, but also
promote synthesis of catabolic factors, such as COX2, nitric oxide
(NO) and NO synthase (NOS), which aggravate apoptosis of OA
chondrocytes and cartilage inflammation (11).
Traditional Chinese medicine is widely used in Asian
societies. Many herbs were employed for the treatment of arthritis
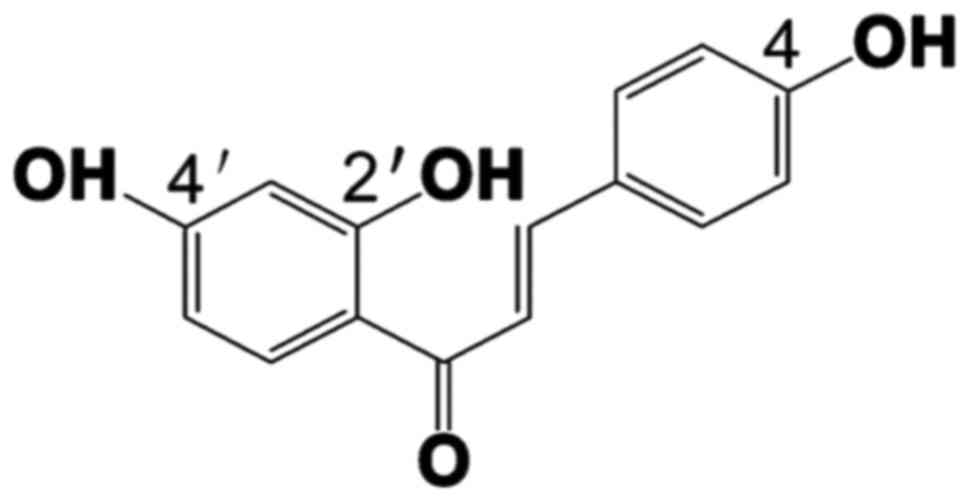
and other inflammatory diseases for centuries (12,13). Isoliq uiritigenin (ISL) (Fig. 1), a natural flavonoid extracted
from licorice, has drawn wide attention due to its lots of
biological activities, including vasorelaxant, anti-viral,
anti-oxidant, anti-platelet aggregation as well as
anti-inflammatory properties and its proven pharmacologic safety
(14,15). The licorice is also used in
Western countries widely for culinary purpose. Although Zhu et
al (16) reported that
licorice be used in inhibition of osteoclast differentiation, which
is a significant physiopathological mechanism of OA (17), a search of Medline, PubMed
(carried out at in February, 2017) revealed no article on the
subject of licorice be used in treatment of OA. Conversely, it has
been previously reported that ISL could prevent the progression of
psoriasis-like symptoms in mice and inhibit LPS-stimulated COX-2
expression in RAW 264.7 macrophages, which both as a result of
attenuation of the NF-κB signaling pathway (18), which is a central regulator of the
inflammatory cytokine-induced catabolic actions in chondrocytes
(19). A further literature
review indicated that the effects of ISL on chondrocyte-like ATDC5
cells have not been investigated at the cellular or molecular
levels yet. Accordingly, the aim of the present study was to access
whether ISL could inhibit IL-1β-stimulated inflammation and
apoptosis by decreasing NF-κB activation in chondrocyte-like ATDC5
cells. In addition, the authors determined whether ISL had
potential protective effects on cartilage of anterior cruciate
ligament transaction models in mice.
Materials and methods
Ethics approval
The experimental schemes were approved by the
Institutional Animal Care and use Committee of First Affiliated
Hospital of Xinjiang Medical University (protocol no.
IACUC20160616-08).
Materials and methods
ISL (purity >98%) was purchased from
Aladdin® (Shanghai, China). Fetal bovine serum (FBS),
Dulbecco's modified Eagle's minimum essential medium/Ham's F12
medium (DMEM/F12), penicillin, streptomycin, insulin, transferrin,
selenium (ITS) and Trypsin were purchased from Invitrogen; Thermo
Fisher Scientific, Inc. (Waltham, MA, USA). Alcian Blue 8GX was
purchased from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). The
primary antibodies against GADPH, Bax, Bcl-2, NF-κB p65,
phospho-p65, caspase-3, cleaved-caspase-3, caspase-9 and
cleaved-caspase-9 were bought from Cell Signaling Technology, Inc.
(Danvers, MA, USA); COL II, MMP-13, COX-2 were purchased from Abcam
(Cambridge, MA, USA) and C57BL/6 male mice (n=80) of 3 months old
were purchased from Vital River Laboratories (Beijing, China).
Cell differentiation and treatment
Cultures of undifferentiated ATDC5 cells (Riken Cell
Bank, Tsukuba, Japan) were maintained in DMEM/F12 supplemented with
5% (v/v) FBS, 100 U/ml penicillin and 100 µg/ml streptomycin
in a humidified incubator with 5% CO2 at 37°C. Once the
culture was confluent 70–80%, the medium (as described above)
supplemented with 1% ITS additionally. The differentiation medium
was changed every two days for 3 weeks to induce differentiation
into chondrocyte-like cells. Differentiation was confirmed by the
expression level of mRNA for COL II and COL X. The production of
glycosaminoglycan was visualized by 1% Alcian Blue staining. The
cells were used for the experimental procedures after
differentiation for 14 days. The serum-starved chondrocyte-like
cells (cultivated in 1% FBS) were incubated alone for 24 h and
pretreated with ISL (2.5, 5, 10, 20 and 40 µmol/l) for 1 h,
then co-treated with IL-1β (10 ng/ml) for a further 48 h in
monolayer cultures. The cells treated with IL-1β only were used as
the model group and the chondrocyte-like cells which only exposed
to serum-starved medium served as the normal control.
Cell viability assay
Chondrocyte-like ATDC5 cells were seeded in 96-well
plates with 4×103 cells/well and cultured with 100 ml
maintenance medium for 24 h. The following day, the cells
pretreated with ISL (2.5, 5, 10, 20 and 40 µmol/l) for 1 h,
then co-incubated with IL-1β (10 ng/ml) for a further 24, 48 and 72
h, respectively and Cell Counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was used to monitor cell
viability. The number of viable cells was assessed by measurement
of absorbance at 450 nm after additional 2-h incubation with CCK-8
by a microplate reader Multiskan Go (Thermo Fisher Scientific,
Inc.).
Quantitative analysis of apoptosis
cells
PE-Annexin V/7-amino-actinomycin (7-ADD)
double-fluorescence labeling and flow cytometry were performed to
quantify changes of cell apoptosis. Following co-treatment with
IL-1β (10 ng/ml) and ISL (2.5, 5 and 10 µmol/l) for 48 h,
1×106 cells were harvested and washed 3 times with
pre-cold phosphate-buffered saline (PBS), then re-suspended in
binding buffer followed by apoptosis detection kit (BD Biosciences,
Franklin Lakes, NJ, USA). The cells were also treated with only ISL
(5, 10, 20 and 40 µmol/l) for 48 h and received the same
pretreatment. The samples were analyzed using FACSAria™ II flow
cytometer (BD Biosciences) after PE-Annexin V and 7-ADD marking at
indoor temperature for 15 min in the dark.
Gene expression analysis
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) was
used according to the manufacturer's protocols to extract total RNA
from chondrocyte-like ATDC5 cells. A spectrophotometer at 260 nm
was selected to quantify total RNA and purity was evaluated by
determining the ratio of A260/A280. All samples had ratios above
1.90. cDNA was synthesized using ~2 µg RNA with an
PrimeScript™ RT Master Mix (Takara Bio, Inc., Otsu, Japan) and was
subjected to reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) reactions with SyBR® Fast qPCR Mix
(Takara Bio, Inc.) using a CFX96 real-time PCR system (Bio-Rad
Laboratories, Inc., Hercules, MA, USA) under the condition of 94°C
for 30 sec, followed by 40 cycles at 95°C for 5 sec, 60°C for 10
sec, and finally the dissociation curve of each primer pair was
analyzed to determine the primer specificity. The primers were
synthesized and supplied by Sangon Biotech Co., Ltd. (Shanghai,
China). Sequences of the primers used are listed in Table I. All of the PCR reactions were
performed 3 times for each gene. The data were analyzed by the
2−ΔΔCq method (20)
using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the
internal control.
 | Table IPrimer sequences for different
genes. |
Table I
Primer sequences for different
genes.
| COL II | F:
5′-ACGAAGCGGCTGGCAACCTCA-3′ |
| R:
5′-CCCTCGGCCCTCATCTCTACATCA-3′ |
| COX-2 | F:
5′-TTCCAATCCATGTCAAAACCGT-3′ |
| R:
5′-AGTCCGGGTACAGTCACACTT-3′ |
| MMP-13 | F:
5′-TGTTTGCAGAGCACTACTTGAA-3′ |
| R:
5′-CAGTCACCTCTAAGCCAAAGAAA-3′ |
| GAPDH | F:
5′-AGGTCGGTGTGAACGGATTTG-3′ |
| R:
5′-GGGGTCGTTGATGGCAACA-3′ |
Western blot analysis
Radioimmunoprecipitation assay lysis buffer with
protease inhibitors (Sigma-Aldrich; Merck KGaA) were used to lyse
treated cells on ice for 30 min according to the manufacturer's
protocols. The protein concentration was determined by the
bicinchoninic acid protein assay kit (Sangon Biotech Co., Ltd.).
Equal amounts (20 µg/load) of protein samples were resolved
on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) (Bio-Rad Laboratories, Inc.) electrophoresis and
transferred on to polyvinylidene fluoride difluoride membranes
(Merck KGaA). 5% bovine serum albumin (BSA) were selected to block
the membranes for 1 h at room temperature and then incubated with
the primary antibodies COL II (1:5,000, cat. no. ab34712), MMP-13
(1:3,000, cat. no. ab39012), COX-2 (1:1,000, cat. no. ab62331) (all
from Abcam), GAPDH (1:1,000, cat. no. 2118S), Bcl-2 (1:1,000, cat.
no. 3498), Bax (1:1,000, cat. no. 14796), caspase-3 (1:1,000, cat.
no. 14220), cleaved caspase-3 (1:1,000, cat. no. 9654), caspase-9
(1:1,000, cat. no. 9508), cleaved caspase-9 (1:1,000, cat. no.
7237), NF-κB p65 (Cell Signaling Technology Inc., 1:1,000, cat. no.
6956), phospho-NF-κB p65 (1:1,000, cat. no. 3036) (all from Cell
Signaling Technology, Inc.) at 4°C overnight with gentle rocking.
The membranes were then washed 3 times, and then cultured with the
secondary antibody (peroxidase-conjugated AffiniPure goat
Anti-Rabbit IgG (H+L), 1:5,000, cat. no. ZB-2301, Origene
Technologies, Inc., Beijing, China) for 2 h, and immunoreactivity
was detected with the ECL substrate (Thermo Fisher Scientific,
Inc.). GADPH was used as an internal control in all western blot
analyses.
Anterior cruciate ligament transection
(ACLT) model in mice
The experimental schemes were approved by the
Institutional Animal Care and use Committee of the First Affiliated
Hospital of Xinjiang Medical University. The animals were kept in a
controlled envivonment (temperature, 22–25°C; light/dark cycle, 12
h; relative humidity, 60%) and were allowed free access to drinking
water and fodder) The 3-month-old male C57BL/6J mice were
anesthetized with intravenous injection of 1% pentobarbitone (40
mg/kg). After a routine sterilize, the patella was laterally
dislocated through medially parapatellar arthrotomy. The ACL was
visualized and transected with a scissors. The articular cavity was
irrigated and sutured layer-by-layer, then the mice underwent
penicillin treatment (400,000 IU/Kg) for one week. Sham operation
was conducted by opening the joint capsule and suturing the
incision in the right knee of independent rodents.
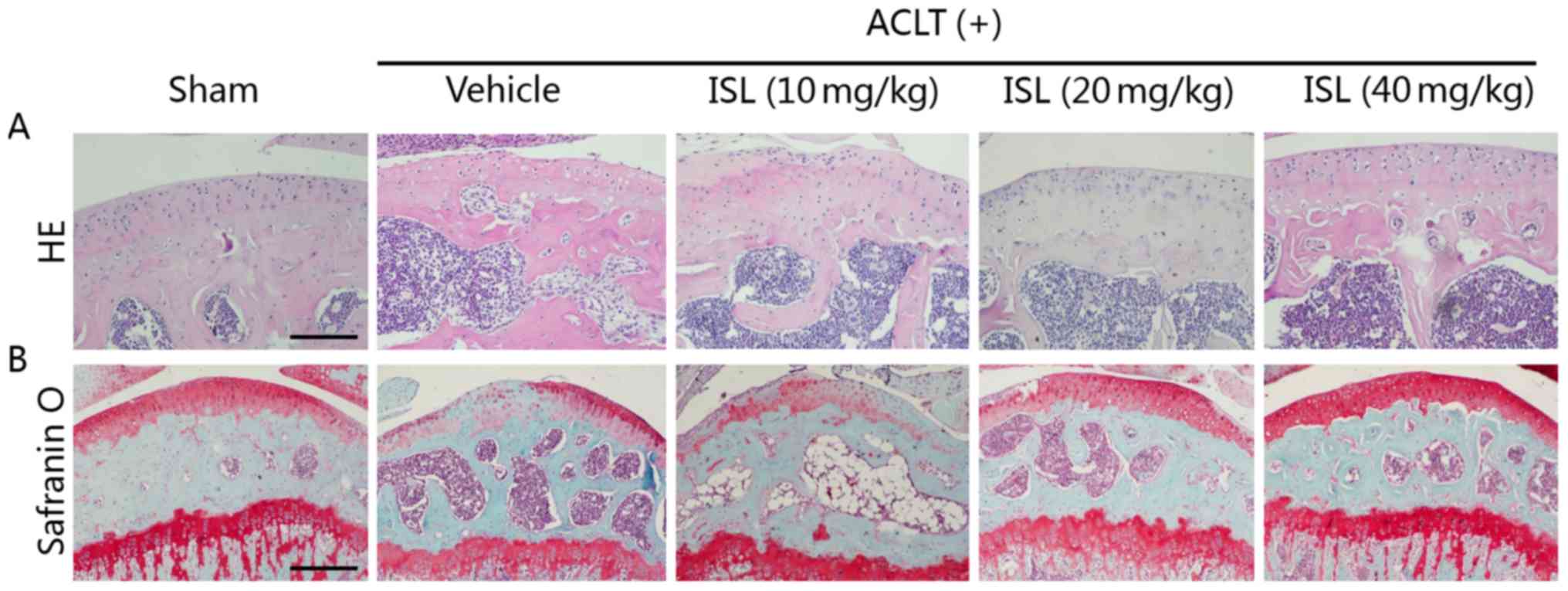
Preliminary experiment was performed firstly, the
optimal dose (40 mg/kg) was identified using multiple
concentrations of ISL (10, 20 and 40 mg/kg) injected every other
day for 2 months post-operation (Fig.
8). Lower concentration (10 or 20 mg/kg) had minimal effects on
chondroprotection. The authors did not continue to increase the
dose because nearly half of mice died in higher concentration (80
mg/kg group) during the 2 months post-operation. Therefore, in the
formal experiment, the mice were randomized to sham group, ACLT +
vehicle group, or ACLT + ISL (40 mg/kg) group. Beginning the second
day after surgery, ISL or equivalent volume of vehicle (10%
Tween-80) was injected intraperitoneally every other day for 60
days. Mice were sacrificed by CO2 inhalation at 60 days
after surgery. Knee joints of mice were dissected, fixed in 10%
buffered formalin for 48 h, and decalcified in 10% EDTA (pH 7.4)
for 3 weeks. Specimens were embedded in paraffin and cut for
hematoxylin and eosin (H&E) and Safranin O and fast green
staining. Osteoarthritis Research Society International-modified
Mankin criteria (OARSI) scores were calculated for evaluating the
state of articular cartilage in each group.
Statistical analysis
If the data met homogeneity of variance, statistical
significance between different groups was evaluated by one-way
analysis of variance followed by Tukey's post hoc test. If the data
did not meet homogeneity of variance, Dunnett's T3 post hoc test
was used. Student's t-test were performed to compare mean values
from different samples. Statistical tests were carried out using
SPSS version 18.0 software (SPSS Inc., Chicago, IL, USA). Data are
expressed as mean ± standard deviation from three separate
experiments.
Results
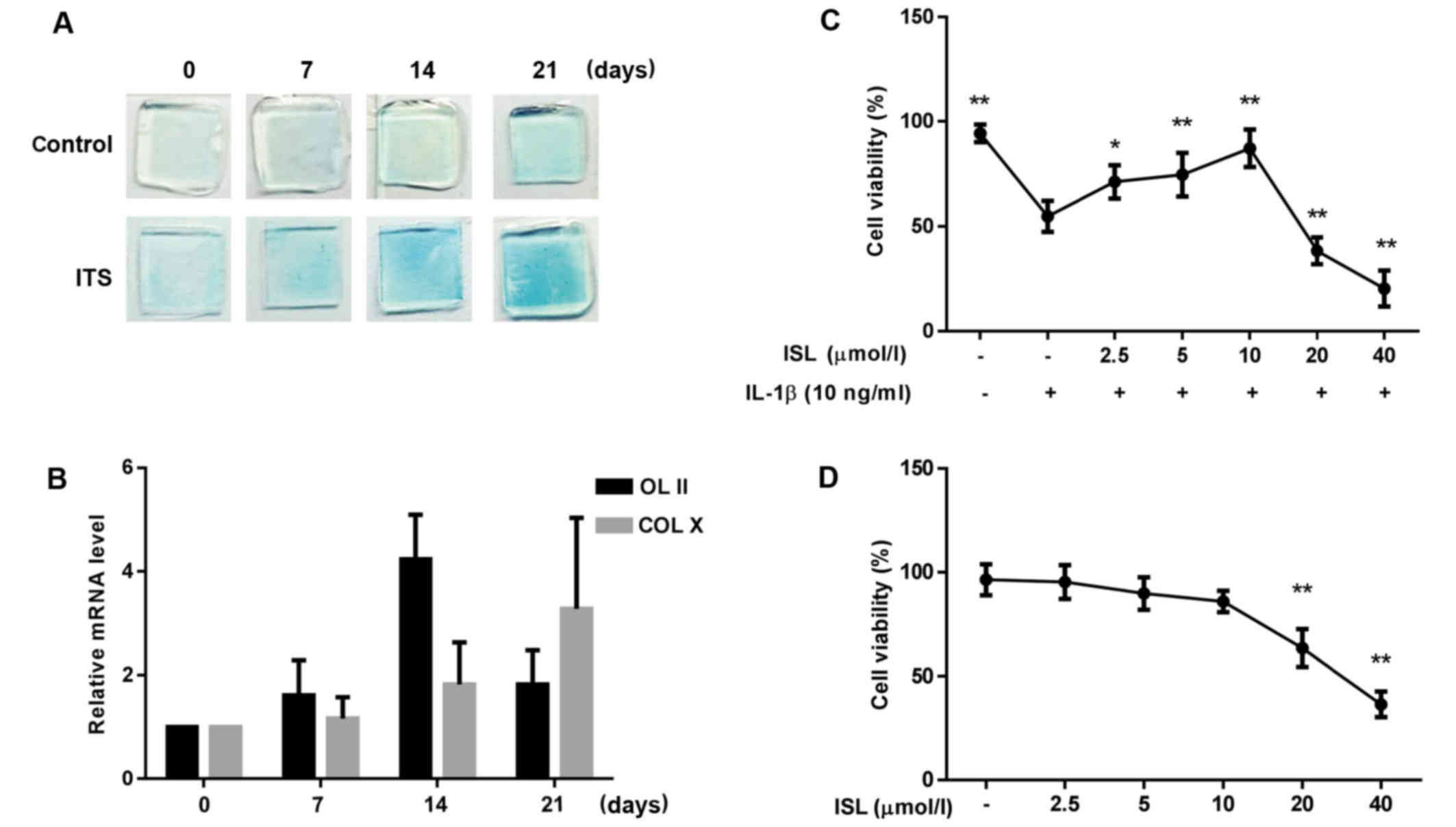
Differentiation of ATDC5 cells
The authors evaluated whether treatment with ITS
induces formation of cartilage nodules in ATDC5 cells. The cells
were stimulated with ITS for 21 days and 1% Alcian blue staining
were performed at 0, 7, 14 and 21 days. Fig. 2A presents that staining
intensities gradually increased in a time-dependent manner from 0
to 21 days in ATDC5 cells cultured with ITS. The authors then
assessed the expression of chondrogenic differentiation markers
including COL II and COL X by using RT-qPCR. As Fig. 2B indicates, COL II mRNA increased
significantly after 7 days of chondrogenic induction and further
increased with a maximum elevation at 14 days, which indicates
early-stage differentiation of chondrocytes. mRNA expression of
collagen X gradually increased and exceeded COL II at 21 days,
indicating late-stage differentiation of chondrocytes. These
results show that undifferentiated ATDC5 cells differentiate in
culture from proliferative chondrocytes to hypertrophic
chondrocytes. Therefore, ATDC5 cells with differentiation of 14
days were selected as following experimental cells.
Effects of ISL on cell viability
CCK-8 analysis was performed to study effects of ISL
on viability of chondrocyte-like ATDC5 cells incubated with IL-1β.
The results demonstrated a significant decrease of IL-1β-induced
chondrocytes viability compared with the normal control, which was
reversed by ISL at low concentrations (2.5, 5 and 10 µmol/l)
in a dose-dependent manner (Fig.
2C). However, higher doses of ISL (20 and 40 µmol/l),
did not reverse the viability of chondrocyte-like ATDC5 cells.
Correspondingly, the cells were treated with only ISL at different
concentrations indicated that the viability of the cells was not
remarkable change within the scope of 2.5–10 µmol/l, while
significant decrease at high concentrations (20 and 40
µmol/l) (Fig. 2D). As a
result, 2.5, 5 and 10 µmol/l ISL were selected as the low,
medium and high concentrations, respectively. The criterion was
selected in subsequent experiments.
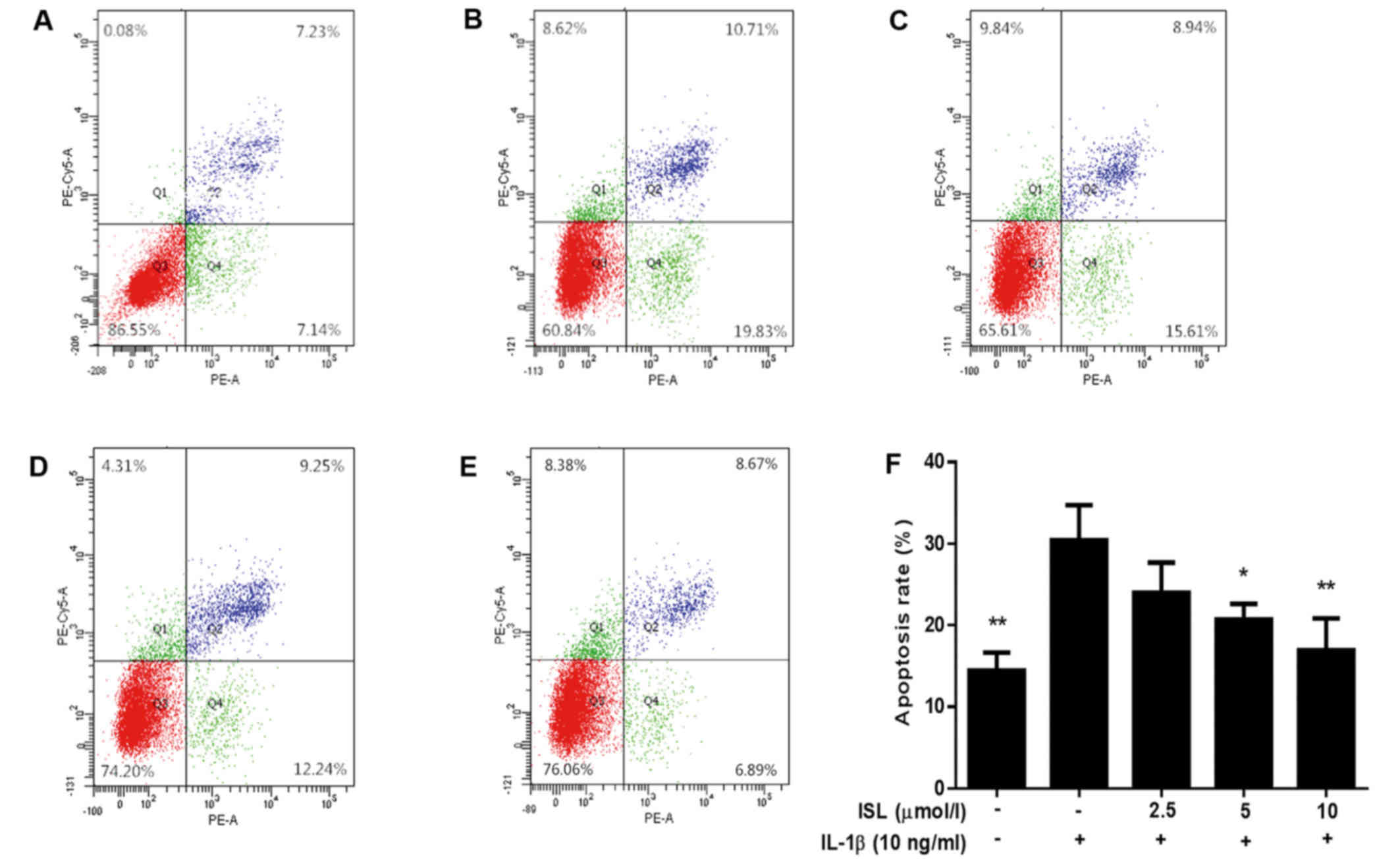
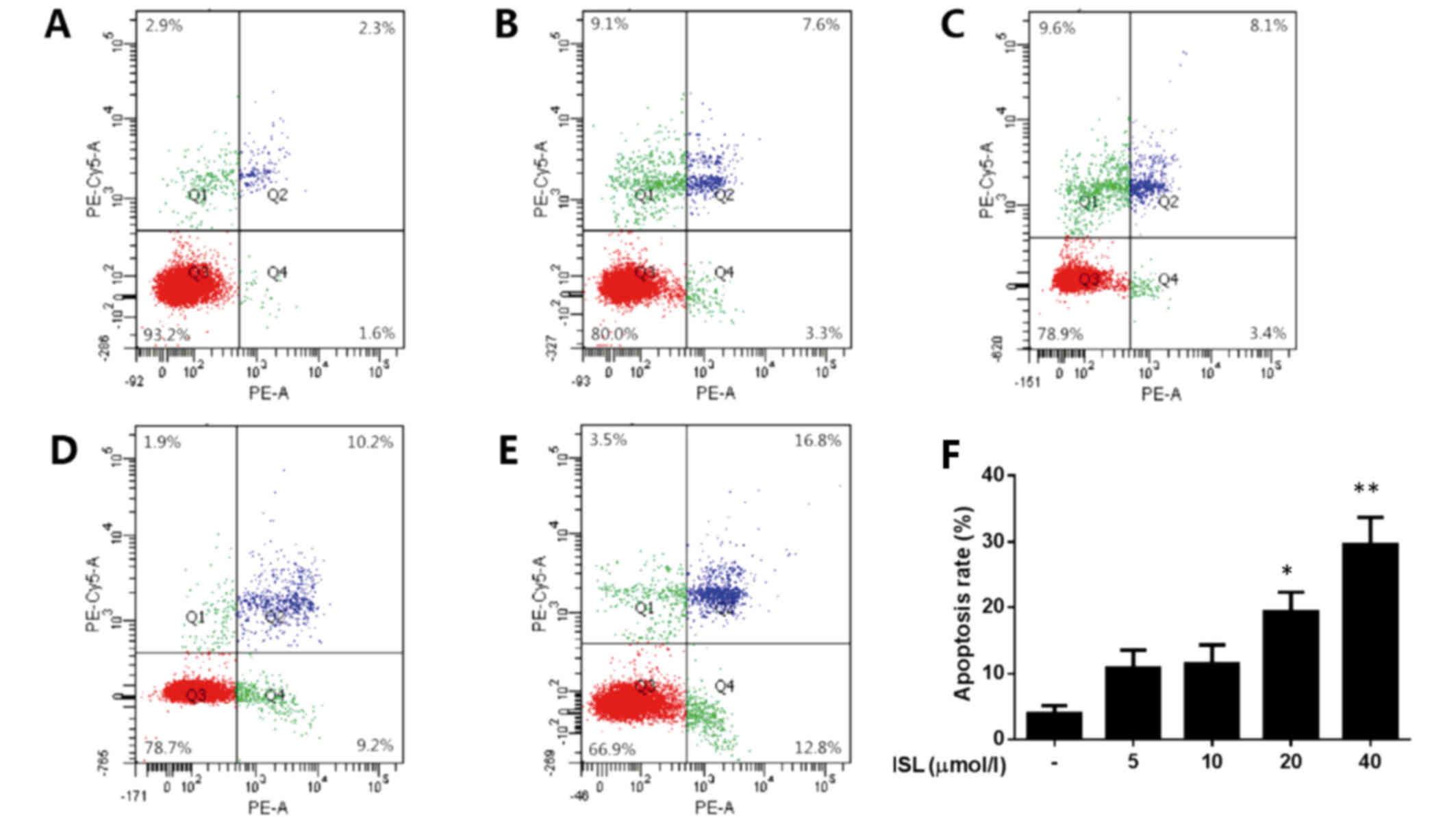
ISL decreased the apoptotic rate of
chondrocyte-like ATDC5 cells
The effects of ISL on cell apoptosis was determined
by flow cytometry using PE-Annexin V/7-ADD double-fluorescence
labeling. The percentage of apoptotic cells in IL-1β treated group
was prominently increased compared with the normal control.
Chondrocyte-like ATDC5 cells co-treated with ISL and IL-1β showed a
decrease apoptotic rate, in a dose-independent manner (Fig. 3). Additionally, ISL induced only
mild cytotoxicity at lower concentrations (5 and 10 µmol/l),
but it exhibited apparent cytotoxicity at higher concentrations (20
and 40 µmol/l) (Fig.
4).
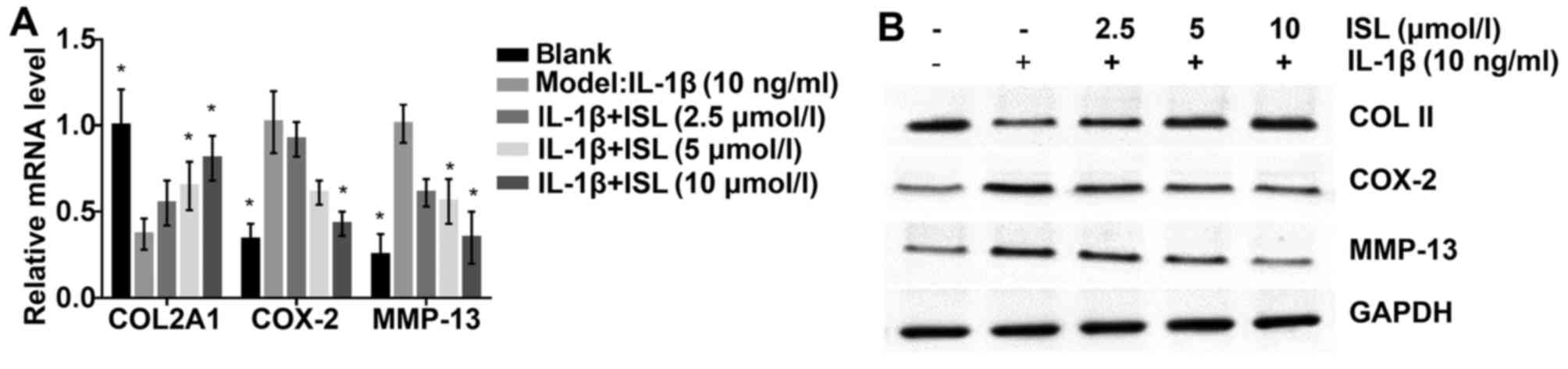
Protective effects of ISL against
catabolic responses on mRNA and protein expression
Because IL-1β contributes to the synthesis of more
pro-inflammatory factors and matrix-degraded enzymes such as MMPs
and COX-2. The authors further examined the anti-inflammatory
effect of ISL on chondrocyte-like ATDC5 cells. The mRNA expressions
of COL, COX-2 and MMP-13 gene were assessed by RT-qPCR (Fig. 5A). The results showed that in the
IL-1β-induced ATDC5 cells treated by ISL, the mRNA levels of COX-2
and MMP-13 were evidently decreased dose-dependently, whereas the
level of COL was increased significantly. The results of western
blotting consistently demonstrated the anti-inflammatory effect of
ISL (Fig. 5B).
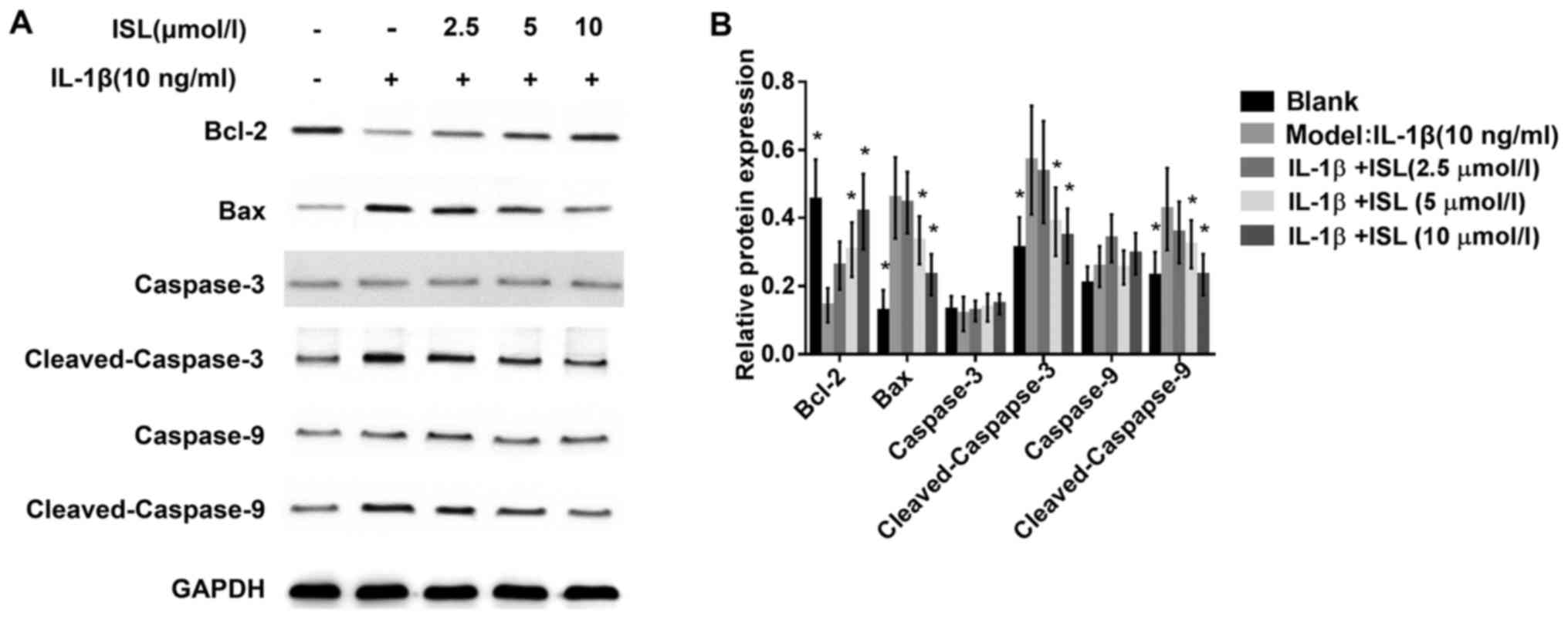
Anti-apoptotic effects of ISL on
IL-1β-stimulated chondrocyte-like ATDC5 cells
In order to evaluate whether ISL could modulate the
expressions of anti-apoptotic and promoting-apoptotic genes,
western blotting were performed and the results demonstrated that
the expression of anti-apoptotic protein Bcl-2 dose-dependent
increased with the increasing concentration of ISL while the
expression level of pro-apoptotic protein Bax decreased in a
concentration manner when ADTC5 cells were stimulated with IL-1β
(Fig. 6A). These results
indicated ISL exerted anti-apoptotic effects in IL-1β-induced
chondrocyte-like ATDC5 cells.
To provide further evidence for anti-apoptotic
effects of ISL on IL-1β-stimulated chondrocyte-like ATDC5 cells,
the downstream protein levels of caspase-3 cleavage and caspase-9
cleavage that are in the mitochondria apoptosis pathway were
assessed. Western blot data revealed that ISL had few influence on
expression of caspase-3 and caspase-9. However, the expression of
cleaved-caspase-3 and cleaved-caspase-9 was prominent in
chondrocyte-like ATDC5 cells stimulated with IL-1β and was blocked
in the cells treated with ISL (Fig.
6B).
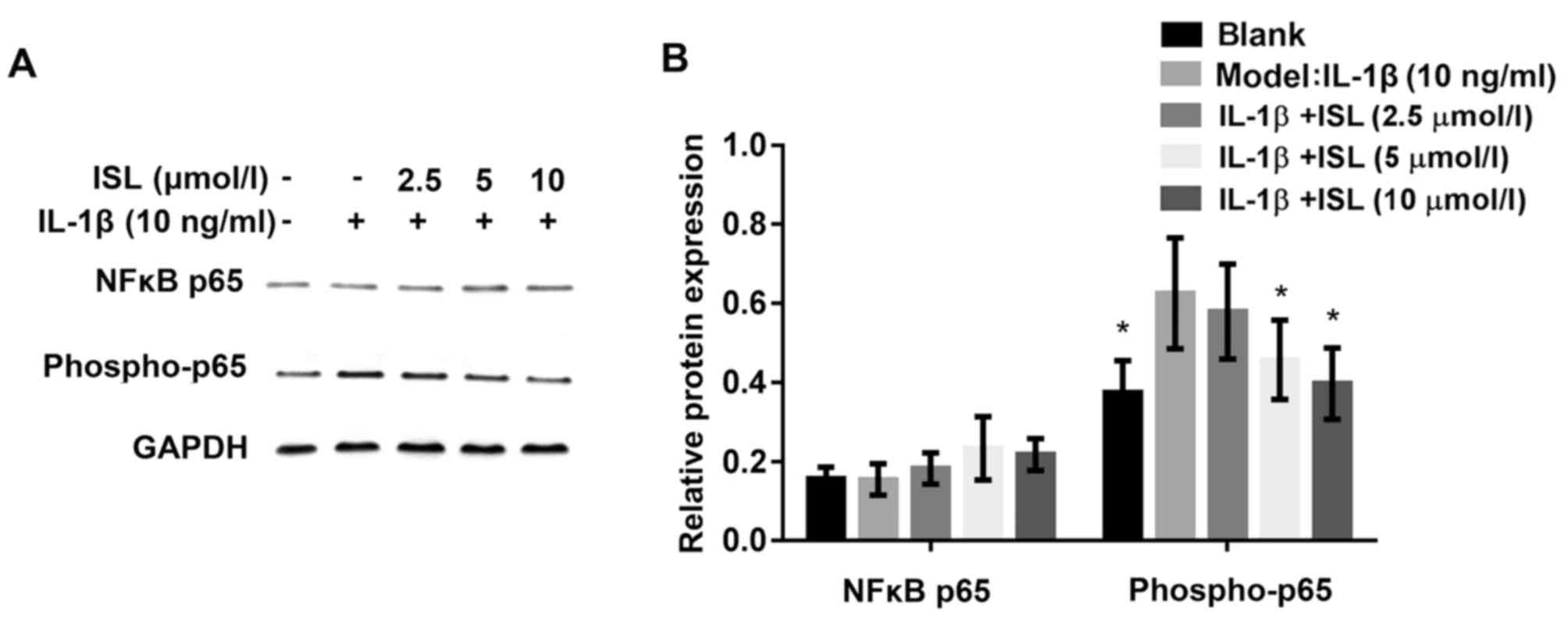
ISL inhibits NF-κB p65 phosphorylation in
IL-1β-stimulated chondrocytes
Based on the data above, the authors speculated that
the NF-κB signaling pathway may be inhibited by ISL. To determine
whether ISL inhibited IL-1β-induced NF-κB p65 phosphorylation,
nuclear protein extracts from serum-starved chondrocyte-like ATDC5
cells were probed for the phosphorylated form of NF-κB after
pretreatment with ISL for 1 h followed by IL-1β stimulation for a
further 48 h. The western blot data indicated that pretreatment of
ISL attenuated IL-1β-stimulated NF-κB phosphorylation in a
dose-dependent manner (Fig.
7A).
ISL exerts chondroprotective effects on
knee joint tissue of mice
At last, the in vivo therapeutic effects of
ISL in OA were evaluated by using a mouse ACLT model. The medial of
the tibia plateau from the operative hind leg was sectioned and
observed 8 weeks after operation. H&E staining demonstrated
decreased thickness of calcified cartilage zone in ISL (40
mg/kg)-treated ACLT mice relative to vehicle-treated ACLT controls
(P<0.05) (Fig. 9A and B).
Specifically, in the vehicle group, the surface of the articular
cartilage was rough, and the intensity of Safranin O staining in
the matrix was low. However, the superficial layer of the cartilage
in ISL-treated ACLT mice was smooth. There was no disruption of
surface integrity and strong staining with Safranin O was observed
in these ACLT mice (Fig. 9A,
lower right panel). OARSI scores in ISL-treated ACLT mice were
improved compared to the vehicle-treated ACLT controls, whereas no
difference was noted in ISL versus sham controls (Fig. 9C).
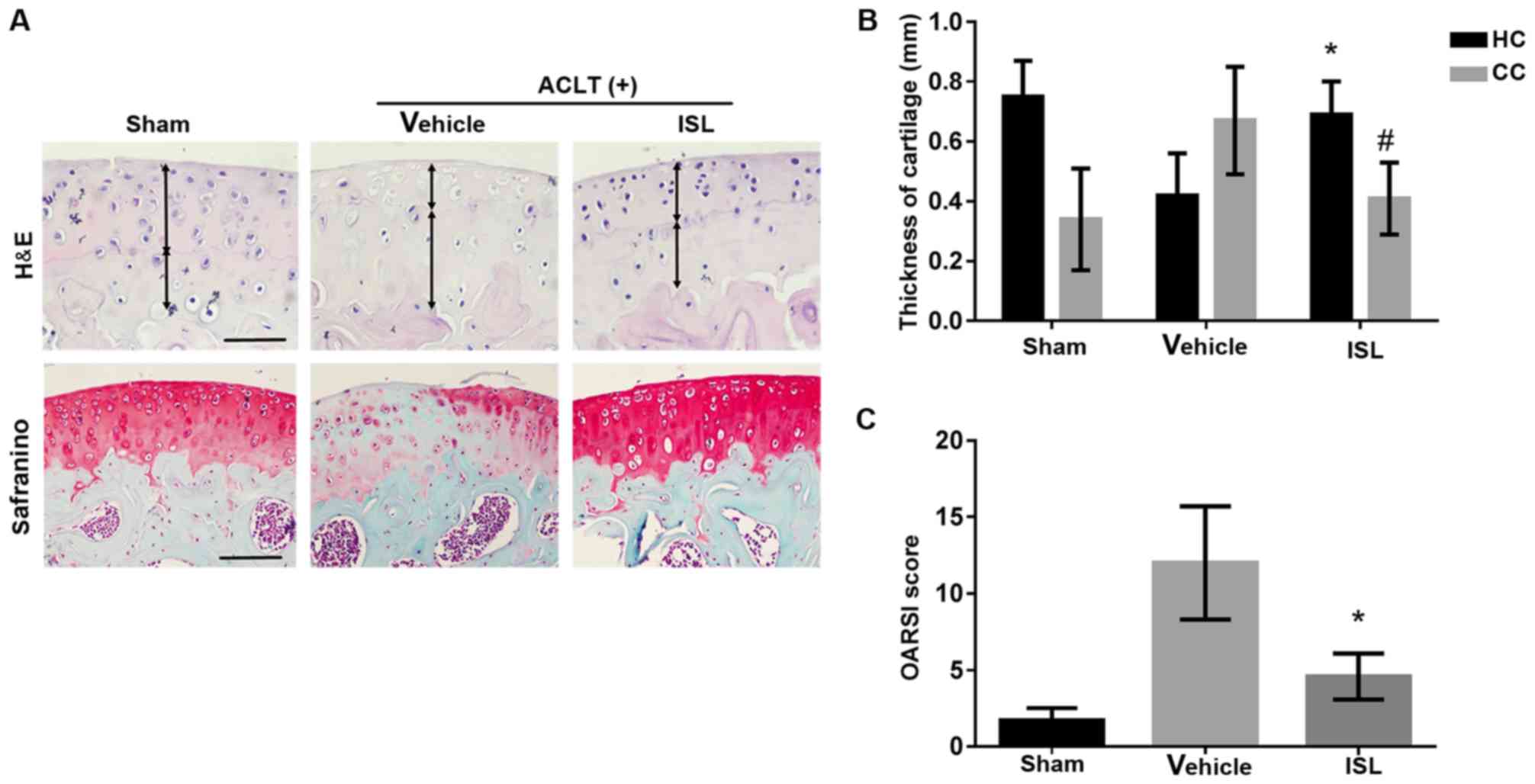 | Figure 9ISL exhibited chondroprotective
effects on a mice ACLT model. Mice underwent ACLT operation and
received intraperitoneal injections with 10% Tween-80 or ISL as
described in Materials and methods. The mice were sacrificed and
their knee joints were excised eight weeks after ACLT operation.
(A) H&E staining (top) where the thickness of CC and HC in each
group were measured (double-headed arrows) and (B) quantitative
analyzed. Scale bars, 100 µm. Safranin O and fast green
staining (bottom): Solid arrows indicate proteoglycan loss and
cartilage destruction. Scale bars, 100 µm. (C) OARSI scores
of articular cartilage at 8 weeks after surgery. Sham,
sham-surgery; vehicle, ACLT-surgery treated with 10% Tween-80;
ISL,ACLT-surgery treated with isoliquiritigenin. n=6/group. The
values represent the mean ± standard deviation.
#P<0.05 and *P<0.05 vs. vehicle group.
ISL, isoliquiritigenin; ACLT, anterior cruciate ligament
transection; H&E, hematoxylin and eosin; CC, calcified
cartilage zone; HC, hyaline cartilage zone; OARSI, Osteoarthritis
Research Society International-modified Mankin. |
Discussion
Historically, different types of therapeutic agents
and drug leads have been extracted from natural products (21). A great deal of effort has recently
been put toward finding bioactive small molecules from natural
herbage that may be suitable for treatment of OA, especially those
with minimal or no adverse side effects (22–24). ISL, a bioactive small molecules
isolated from licorice, which is one of the most commonly used
herbs in traditional Chinese medicine, has been used to treat many
kinds of ailments ranging from skin diseases to peptic ulcers and
rheumatoid arthritis (23). In
the present study, the anti-apoptotic and anti-inflammatory effects
of ISL on chondrocyte-like ATDC5 cells were investigated. The
present study is the first to demonstrate that ISL can prevent
IL-1β-induced chondrocyte-like ATDC5 cells apoptosis and
inflammation by inhibiting NF-κB activation. These
chondroprotective effects were also observed in an ACLT model of
osteoarthritis.
Pro-inflammatory cytokines have been identified to
play a crucial role in the progression of OA, especially IL-1β and
TNF-α, which are considered as prominent cytokines for destruction
of articular cartilage (8). One
of the major effects of IL-1β seems to be the induction of
catabolic processes that affects cartilage matrices. It has been
reported that IL-1β not only induces the release of degenerative
enzymes, such as MMPs, from synoviocytes and chondrocytes, but also
inhibits the expression of a large number of genes related with the
differentiated chondrocyte phenotypes such as COL II (8). IL-1β was also found to activate
cytokine receptors on surfaces of articular chondrocytes, which
leads to induction of the NF-κB signaling pathway (25). This pathway, acting either by
itself, or in cooperation with bone morphogenetic protein, Wnt or
other signaling cascades, interferes with the anabolic activity of
chondrocytes (26) and leads to
increases in expression of the inflammatory gene products COX-2
(25). Expression of inducible
NOS, which is induced by a single stimulation of IL-1β (27), was detected in synovial tissue and
articular cartilage in joints affected by OA (28,29). In this way, IL-1β may cause
apoptotic death of chondrocytes via overproduction of NO. Moreover,
increased levels of apoptosis were observed in human chondrocytes
from normal and OA cartilage that had been incubated with human
IL-1β (30). For this reason,
IL-1β was selected to treat ATDC5 cells in cell viability assay
(Fig. 2C) and following
experiment, which made this in vitro study more
reliable.
In the cell viability assay, it is noteworthy that
instead of reverse the viability of chondrocyte-like ATDC5 cells,
higher doses of ISL (20 and 40 µmol/l) lead to significant
decrease of IL-1β-induced chondrocytes viability. Following that,
cytotoxicity of ISL was examined by CCK-8 assay and flow cytometry
using PE-Annexin V/7-ADD double-fluorescence labeling. The ATDC5
cells were treated with only ISL at different concentrations, and
results indicated that ISL induced only mild cytotoxicity at lower
concentrations (2.5, 5 and 10 µmol/l), but it exhibited
apparent cytotoxicity at higher concentrations (20 and 40
µmol/l) (Figs. 2D and
4). These data indicated the
stacking effect of toxicity in cells treated with 10 ng/ml IL-1β
plus ISL at different concentrations and help explain the biphasic
pattern in the viability of IL-1β-stimulated chondrocytes depending
on the concentration of ISL. Beyond that, ISL is one kind of
phytoestrogen. Dual-directional regulation effect of phytoestrogen
(31), which means that the
contrary effect would be present depending on the concentration of
treatment, may contribute to this complex mechanism. The results
also indicated that IL-1β promoted the levels of MMP-13 and COX-2,
and decreased the levels of COL II in chondrocyte-like ATDC5 cells.
This effect was also reversed by ISL in a dose-independent manner
(Fig. 5).
Chondrocytes are the only cell type in articular
cartilage, and they play a significant role in dynamic balance
between anabolism and catabolism of cartilage ECM (5). A regular balance between tissue
destruction and repair is essential to maintaining joint function.
However, this balance is broken towards overall tissue destruction
in pathological process of OA (19). This death of chondrocytes is the
most prominent feature of arthritis. Natural chondrocytes survival
factors have potential to be developed for therapeutic application.
Our results from Annexin V/7-ADD double-fluorescence staining
indicated that ISL inhibited the apoptotic rates of IL-1β-induced
chondrocyte-like ATDC5 cells in a dose-independent manner (Fig. 3).
Inflammatory cytokines simulate a series of
downstream signaling pathways, including MAPK and NF-κB pathways,
which are aberrantly activated in chondrocytes of OA (18). The NF-κB molecules are a family of
widely expressed transcription factors involved in stress
responses, immunity, cell proliferation, inflammatory diseases and
cell apoptosis (32). The NF-κB
pathway is a central regulator of the inflammatory cytokine-induced
catabolic actions in chondrocytes and triggers the secretion of
several matrix-degrading proteinases, including the MMPs and the
aggrecanases, ADAMTS4 and ADAMTS5, leading to articular cartilage
breakdown (33). Moreover, the
NF-κB pathway leads to upregulation of the inflammatory gene
products COX-2, which rapidly induces the production of
pro-inflammatory cytokines and MMPs during the early stage of OA,
upregulates the expression of pro-apoptotic genes, and
downregulates the expression of anti-apoptotic genes at the late
stage of OA (30). Meanwhile,
NF-κB has been shown to regulate the expression of several members
of the Bcl-2 gene family transcriptionally, which remains the
prototypic anti-apoptotic protein (34). Phosphorylation of NF-κB p65 is one
of major signaling mechanisms associated with cell apoptosis. The
present results indicated ISL had no obvious influence on the
expression of p65, but it could inhibit phosphorylation of p65
(Fig. 6) and markedly alleviated
the increase in cleaved-caspase-3 and cleaved-caspase-9 activity in
IL-1β-stimulated chondrocyte-like ATDC5 cells (Fig. 5).
In addition, the authors found that ISL exerted
protective effects on cartilage in ACLT mice. Hyaline cartilage is
the primary degenerative substance during OA, linking the calcified
cartilage by tidemark. Calcified cartilage is a highly mineralized
region in the articular cartilage, which is the end stage of
hyaline cartilage (35). It is
here reported that the activation of the NF-κB signaling pathway
can facilitate the transformation of articular chondrocytes from a
pre-hypertrophic state to a terminal differentiation state,
hypertrophy. The outcome of this hypertrophic transformation is the
calcification of chondrocytes from hyaline cartilage zone and the
formation of osteophytes at the joint periphery in the progression
of OA (36). In mild OA, the
thickness of calcified cartilage first increased at first and then
decreased, but it increased progressively in cases of moderate OA,
which suggests that the early pathological changes in calcified
cartilage are reversible (37).
The data collected here indicated that the calcified cartilage zone
was significantly thicker 8 weeks after the ACLT operation, which
was reversed by ISL. The OARSI scores as determined using Safranin
O staining, demonstrated retention of proteoglycan in ISL-treated
ACLT mice relative to vehicle-treated ACLT controls. These outcomes
above suggest that ISL may work at the early pathological change of
OA, which could serve as a potential preventive therapy for OA.
According to treatment of ISL in IL-1β induced
chondrocyte-like ATDC5 cells and mouse ACLT models, ISL was found
to suppress IL-1β-induced apoptosis and inflammation in
chondrocyte-like ATDC5 cells by inhibiting the NF-κB signaling
pathway. These results also indicated that ISL exerts protective
effects on articular cartilage in vivo, indicating that ISL
may be a potential novel preventive therapy for OA. These results
may in part explain the mechanisms by which ISL exerts its
beneficial effects in OA, and they have certain limits. The
mechanism underlying the effect of ISL on the NF-κB pathway in
chondrocytes has not yet been fully determined and requires further
study.
Acknowledgments
The present study was supported by grants from the
Major Science and Technology Projects in Xinjiang Uygur Autonomous
Region (grant no. 201430123-3), the National Natural Science
Foundation of China (Youth Fund) (grant no. 81303065) and the
Science and Technology Projects in Urumqi, Xinjiang (grant no.
Y151310024).
References
|
1
|
Lawrence RC, Felson DT, Helmick CG, Arnold
LM, Choi H, Deyo RA, Gabriel S, Hirsch R, Hochberg MC, Hunder GG,
et al: National Arthritis Data Workgroup: Estimates of the
prevalence of arthritis and other rheumatic conditions in the
United States. Part II. Arthritis Rheum. 58:26–35. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Clouet J, Vinatier C, Merceron C,
Pot-vaucel M, Maugars Y, Weiss P, Grimandi G and Guicheux J: From
osteoarthritis treatments to future regenerative therapies for
cartilage. Drug Discov Today. 14:913–925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Haq I, Murphy E and Dacre J:
Osteoarthritis. Postgrad Med J. 79:377–383. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Puig-Junoy J and Ruiz Zamora A:
Socio-economic costs of osteoarthritis: A systematic review of
cost-of-illness studies. Semin Arthritis Rheum. 44:531–541. 2015.
View Article : Google Scholar
|
|
5
|
Wang M, Shen J, Jin H, Im HJ, Sandy J and
Chen D: Recent progress in understanding molecular mechanisms of
cartilage degeneration during osteoarthritis. Ann NY Acad Sci.
1240:61–69. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schuerwegh AJ, Dombrecht EJ, Stevens WJ,
Van Offel JF, Bridts CH and De Clerck LS: Influence of
pro-inflammatory (IL-1 alpha, IL-6, TNF-alpha, IFN-gamma) and
anti-inflammatory (IL-4) cytokines on chondrocyte function.
Osteoarthritis Cartilage. 11:681–687. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
López-Armada MJ, Caramés B, Lires-Deán M,
Cillero-Pastor B, Ruiz-Romero C, Galdo F and Blanco FJ: Cytokines,
tumor necrosis factor-alpha and interleukin-1beta, differentially
regulate apoptosis in osteoarthritis cultured human chondrocytes.
Osteoarthritis Cartilage. 14:660–669. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Malemud CJ, Islam N and Haqqi TM:
Pathophysiological mechanisms in osteoarthritis lead to novel
therapeutic strategies. Cells Tissues Organs. 174:34–48. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hashimoto M, Nakasa T, Hikata T and
Asahara H: Molecular network of cartilage homeostasis and
osteoarthritis. Med Res Rev. 28:464–481. 2008. View Article : Google Scholar
|
|
10
|
Roman-Blas JA and Jimenez SA: NF-kappaB as
a potential therapeutic target in osteoarthritis and rheumatoid
arthritis. Osteoarthritis Cartilage. 14:839–848. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rigoglou S and Papavassiliou Ag: The NF-κB
signalling pathway in osteoarthritis. Int J Biochem Cell Biol.
45:2580–2584. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang F, Wu L, Li L and Chen S: Monotropein
exerts protective effects against IL-1β-induced apoptosis and
catabolic responses on osteoarthritis chondrocytes. Int
Immunopharmacol. 23:575–580. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xing L, Jiang M, Dong L, Gao J, Hou Y, Bai
G and Luo G: Cardioprotective effects of the YiQiFuMai injection
and isolated compounds on attenuating chronic heart failure via
NF-κB inactivation and cytokine suppression. J Ethnopharmacol.
148:239–245. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim JY, Park SJ, Yun KJ, Cho YW, Park HJ
and Lee KT: Isoliquiritigenin isolated from the roots of
Glycyrrhiza uralensis inhibits LPS-induced iNOS and COX-2
expression via the attenuation of NF-kappaB in RAW 264.7
macrophages. Eur J Pharmacol. 584:175–184. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sekine-Osajima Y, Sakamoto N, Nakagawa M,
Itsui Y, Tasaka M, Nishimura-Sakurai Y, Chen CH, Suda G, Mishima K,
Onuki Y, et al: Two flavonoids extracts from Glycyrrhizae radix
inhibit in vitro hepatitis C virus replication. Hepatol Res.
39:60–69. 2009. View Article : Google Scholar
|
|
16
|
Zhu L, Wei H, Wu Y, Yang S, Xiao L, Zhang
J and Peng B: Licorice isoliquiritigenin suppresses RANKL-induced
osteoclastogenesis in vitro and prevents inflammatory bone loss in
vivo. Int J Biochem Cell Biol. 44:1139–1152. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kadri A, Funck-Brentano T, Lin H, Ea HK,
Hannouche D, Marty C, Lioté F, Geoffroy V and Cohen-Solal ME:
Inhibition of bone resorption blunts osteoarthritis in mice with
high bone remodelling. Ann Rheum Dis. 69:1533–1538. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marcu KB, Otero M, Olivotto E, Borzi RM
and Goldring MB: NF-kappaB signaling: Multiple angles to target OA.
Curr Drug Targets. 11:599–613. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu Y, Chen X, Ge X, Xia H, Wang Y, Su S,
Li W, Yang T, Wei M, Zhang H, et al: Isoliquiritigenin prevents the
progression of psoriasis-like symptoms by inhibiting NF-κB and
proinflammatory cytokines. J Mol Med (Berl). 94:195–206. 2016.
View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Koehn FE and Carter GT: The evolving role
of natural products in drug discovery. Nat Rev Drug Discov.
4:206–220. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cui Z, Crane J, Xie H, Jin X, Zhen G, Li
C, Xie L, Wang L, Bian Q, Qiu T, et al: Halofuginone attenuates
osteoarthritis by inhibition of TGF-β activity and H-type vessel
formation in subchondral bone. Ann Rheum Dis. 75:1714–1721. 2016.
View Article : Google Scholar
|
|
23
|
Kim KR, Jeong CK, Park KK, Choi JH, Park
JH, Lim SS and Chung WY: Anti-inflammatory effects of licorice and
roasted licorice extracts on TPA-induced acute inflammation and
collagen-induced arthritis in mice. J Biomed Biotechnol.
2010:7093782010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Blain EJ, Ali AY and Duance VC: Boswellia
frereana (frank-incense) suppresses cytokine-induced matrix
metalloproteinase expression and production of pro-inflammatory
molecules in articular cartilage. Phytother Res. 24:905–912.
2010.
|
|
25
|
Yasuda T: Activation of Akt leading to
NF-κB up-regulation in chondrocytes stimulated with fibronectin
fragment. Biomed Res. 32:209–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ge XP, Gan YH, Zhang CG, Zhou CY, Ma KT,
Meng JH and Ma XC: Requirement of the NF-κB pathway for induction
of Wnt-5A by interleukin-1β in condylar chondrocytes of the
temporomandibular joint: Functional crosstalk between the Wnt-5A
and NF-κB signaling pathways. Osteoarthritis Cartilage. 19:111–117.
2011. View Article : Google Scholar
|
|
27
|
Maier R, Bilbe G, Rediske J and Lotz M:
Inducible nitric oxide synthase from human articular chondrocytes:
cDNA cloning and analysis of mRNA expression. Biochim Biophys Acta.
1208:145–150. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Farrell AJ, Blake DR, Palmer RM and
Moncada S: Increased concentrations of nitrite in synovial fluid
and serum samples suggest increased nitric oxide synthesis in
rheumatic diseases. Ann Rheum Dis. 51:1219–1222. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Loeser RF, Carlson CS, Del Carlo M and
Cole A: Detection of nitrotyrosine in aging and osteoarthritic
cartilage: Correlation of oxidative damage with the presence of
interleukin-1beta and with chondrocyte resistance to insulin-like
growth factor 1. Arthritis Rheum. 46:2349–2357. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Héraud F, Héraud A and Harmand MF:
Apoptosis in normal and osteoarthritic human articular cartilage.
Ann Rheum Dis. 59:959–965. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu HX, Wang Y, Lu Q, Yang MZ, Fan GW,
Karas RH, Gao XM and Zhu Y: Bidirectional regulation of
angiogenesis by phytoestrogens through estrogen receptor-mediated
signaling networks. Chin J Nat Med. 14:241–254. 2016.PubMed/NCBI
|
|
32
|
Oeckinghaus A and Ghosh S: The NF-kappaB
family of transcription factors and its regulation. Cold Spring
Harb Perspect Biol. 1:a0000342009. View Article : Google Scholar
|
|
33
|
Goldring MB and Marcu KB: Cartilage
homeostasis in health and rheumatic diseases. Arthritis Res Ther.
11:2242009. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chu SH, Lim JW, Kim DG, Lee ES, Kim KH and
Kim H: Downregulation of Bcl-2 is mediated by NF-κB activation in
Helicobacter pylori-induced apoptosis of gastric epithelial cells.
Scand J Gastroenterol. 46:148–155. 2011. View Article : Google Scholar
|
|
35
|
Wang F, Ying Z, Duan X, Tan H, Yang B, Guo
L, Chen G, Dai G, Ma Z and Yang L: Histomorphometric analysis of
adult articular calcified cartilage zone. J Struct Biol.
168:359–365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Goldring MB, Otero M, Plumb DA, Dragomir
C, Favero M, El Hachem K, Hashimoto K, Roach HI, Olivotto E, Borzì
RM, et al: Roles of inflammatory and anabolic cytokines in
cartilage metabolism: Signals and multiple effectors converge upon
MMP-13 regulation in osteoarthritis. Eur Cell Mater. 21:202–220.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Deng B, Wang F, Yin L, Chen C, Guo L, Chen
H, Gong X, Li Y and Yang L: Quantitative study on morphology of
calcified cartilage zone in OARSI 0–4 cartilage from osteoarthritic
knees. Curr Res Transl Med. 64:149–154. 2016. View Article : Google Scholar : PubMed/NCBI
|