Introduction
Intestinal ischemia reperfusion (IIR) is a
life-threatening pathological event associated with various
clinical conditions, including vessel occlusion, hernias,
necrotizing enterocolitis and septic shock, and is also an adverse
effect of small bowel transplantation (1,2).
The intestinal mucosa is particularly sensitive to IIR injury due
to the anatomical and physiological characteristics of the villus
microcirculation. A temporary interruption of blood flow (ischemia)
results in endothelial cell barrier dysfunction and proinflammatory
cytokine activation. Paradoxically, the restoration of blood flow
(reperfusion) and reoxygenation exacerbates the local
(epithelial/endothelial) damage and bacterial translocation,
leading to systemic inflammatory response syndrome (SIRS) and
multiple organ dysfunction syndrome (MODS) (3).
Accumulating evidence has demonstrated that IIR is
associated with inflammatory responses and cell death via necrosis
and apoptosis (4). Inflammatory
responses activate immunocompetent cells and release cytokines,
including interleukin-1β (IL-1β), IL-6, IL-10 and tumor necrosis
factor-α (TNF-α) (5), which in
turn aggravate the inflammatory responses to IIR by inducing
microcirculation dysfunction and aggravating cell apoptosis and by
further recruitment and accumulation of inflammatory cells.
Anti-inflammatory therapies significantly attenuate IIR injury.
Nuclear factor (NF) erythroid 2-related factor 2
(Nrf2), a member the of cap 'n' collar/basic region leucine zipper
transcription factor family, participates in the modulation of the
pathogenesis of numerous diseases by regulating the expression of
several antioxidant genes (6,7).
After exposure to oxidative stress, Nrf2 dissociates from Keap1,
translocates into the nucleus and binds to antioxidant responsive
elements (ARE). Various studies have demonstrated that Nrf2 has a
strong anti-inflammatory effect in numerous tissues (8,9).
NF-κB has a pivotal role in immune responses by regulating the
expression of multiple inflammatory genes (10). As a classical pro-inflammatory
factor, NF-κB has been implicated in the regulation of Nrf2. A
recent review summarized that Nrf2 cross-talks with NF-κB (11). However, in IIR, little is known
regarding the anti-inflammatory role of Nrf2 and the possible
counter-balancing effects of Nrf2 and NF-κB in the coordination of
the final fate of innate immune cells. Therefore, the present study
investigated the role of Nrf2 in the modulation of inflammation and
apoptosis caused by IIR.
Materials and methods
Reagents
The TNF-α (cat. no. H052), IL-1β (cat. no. H002),
IL-6 (cat. no. H007), IL-10 (cat. no. H009), D-lactic acid (D-LA;
cat. no. A019-2) and intestinal-type fatty acid-binding protein
(I-FABP; cat. no. H266) enzyme-linked immunosorbent assay (ELISA)
kits specific for mouse cytokines were obtained from Nanjing
Jiancheng Bioengineering Institute (Nanjing, China). Antibodies to
Nrf2 (cat. no. sc-722), NF-κB (cat. no. sc-71675) and
phosphorylated inhibitor of NF-κB (p-IκBα; cat. no. sc-101713) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Antibodies directed against β-actin (cat. no. 4970) and lamin B1
(cat. no. 13435) were purchased from Cell Signaling Technology,
Inc. (Danvers, MA, USA). IRDye 800CW secondary antibodies were
purchased from LI-COR Biosciences (Lincoln, NE, USA), Brusatol
(cat. no. SML1868) and all-trans retinoic acid (ATRA; cat.
no. R2625), specific antagonists of Nrf2 (12,13), were purchased from Sigma-Aldrich,
Merck KGaA (Darmstadt, Germany). t-Butylhydroquinone (t-BHQ; cat.
no. 112976), a specific activator of Nrf2 (12), was also purchased from
Sigma-Aldrich, Merck KGaA. All of the chemicals used were of the
highest grade commercially available.
Animals
This study was approved by the Animal Care Committee
of Wuhan University (Wuhan, China) and protocols were in accordance
with the National Institutes of Health (NIH) guidelines for the
care and use of experimental animals (NIH publication no. 80-23).
This study was performed at the animal center of Renmin Hospital of
Wuhan University (Wuhan, China). A total of 64 adult male C57BL/6J
mice (Hunan Slac JD Laboratory Animal Co., Ltd., Hunan, China; age,
8–10 weeks; weight, 25±3 g) were housed in individual cages (4
mice/cage) in a climate-controlled room (23±1°C; relative humidity
60±5%) with a 12-h light/dark cycle and free access to food and
water. The mice were allowed to acclimatize to the environment for
2 weeks prior to the experiments. All of the animals were fasted
for 12 h prior to the experiments but had free access to water.
Intestinal ischemia-reperfusion
model
All mice were anesthetized by intraperitoneal
injection of sodium pentobarbital (50 mg/kg). Surgery was performed
after the loss of blink and withdrawal reflexes. The mice were then
placed in the supine position and allowed to breathe spontaneously.
The IIR model was established by superior mesenteric artery (SMA)
occlusion (12). In brief, after
laparotomy, the SMA was isolated and temporarily occluded with a
microvascular clip. The mice were subjected to ischemia (45 min)
followed by 120 min of reperfusion by gently removing the clip.
After 120 min of reperfusion, the mice were euthanized and the
intestinal tissues and blood were collected and processed for
further analysis.
Experimental protocol
After surgical preparation, the animals were
randomly allocated into 8 groups as follows (n=8 in each group): i)
S group: Sham surgical preparation with isolation of the SMA but
without occlusion; ii) IIR group: SMA occlusion for 45 min followed
by 120 min of reperfusion; iii) A+S group: Sham surgery plus ATRA
treatment; iv) A+IIR group: IIR procedure plus ATRA treatment; v)
B+S group: Sham surgery plus brusatol treatment; vi) B+IIR group:
IIR procedure plus brusatol treatment; vii) T+S group: Sham surgery
plus t-BHQ treatment; viii) T+IIR group: IIR procedure plus t-BHQ
treatment. For ATRA treatment, the animals received ATRA [2 mg/ml
dissolved in 1% dimethyl sulfoxide (DMSO); 10 ml/kg
intraperitoneally per day] for two weeks prior to the experiment
(13). Brusatol was diluted with
1% DMSO to 0.5 mg/ml and 4 ml/kg was injected intraperitoneally
once every 2 days for 10 days prior to the experiment (14). t-BHQ was diluted with 1% DMSO and
16.7 mg/kg was administered intraperitoneally 3 times/day (every 8
h) for 3 days prior to the experiment, as described previously
(12).
Histopathology of the intestinal
tissue
After reperfusion, 1 cm of small intestine without
adipose tissue was biopsied from the same site from each animal at
the distal end of the ileum and fixed in 4% formaldehyde. Sections
(4-µm) were prepared from the paraffin-embedded tissue and
assessed by hematoxylin and eosin (H&E) staining (hematoxylin
staining for 10–30 sec and eosin staining 1–3 min at 23±1°C) and
light microscopic examination (original magnification, ×200;
Olympus BX50; Olympus Optical, Tokyo, Japan). Intestinal mucosal
damage was evaluated in at least 2 different sections of each
specimen using the improved Chiu et al scoring method
(15), with blinding to the
experimental groups, using a 5-point grading scale according to the
changes in the villi and the glands of the intestinal mucosa: 0,
normal mucosa; 1, development of subepithelial Gruenhagen's space
at the tip of a villus; 2, extension of the space with moderate
epithelial lifting; 3, massive epithelial lifting with a few
denuded villi; 4, denuded villi with exposed capillaries; and 5,
disintegration of the lamina propria, ulceration and
hemorrhage.
Analysis of intestinal edema
Tissue edema was detected by the wet/dry weight
ratio of the biopsied gut segments. At the end of the experiments,
1 cm of small intestine without adipose tissue was taken from the
same site in each animal, weighed and then placed in a drying oven
at 80°C for 24 h. After this drying procedure the specimens were
reweighed, and the ratio of the weight prior to and after drying
was calculated.
ELISA
D-LA, I-FABP, IL-1β, IL-6, IL-10 and TNF-α levels in
the intestinal mucosa and in the serum were measured following the
standard procedures of the ELISA kits.
Terminal deoxynucleotidyl
transferase-mediated 2′-deoxyuridine 5′-triphosphate-biotin nick
end labeling (TUNEL) assay
Apoptosis in the intestinal sections was examined
after TUNEL staining with the Click-iT TUNEL Alexa Fluor 488
Imaging assay (cat. no. C10245; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The 4-µm
paraffin-embedded sections were deparaffinized in xylene and double
diluted water. The sections were then treated with proteinase K for
20 min at room temperature and subsequently incubated with a
mixture of fluorescent labeling solution and TdT enzyme for 1 h in
a humidified atmosphere. After washing with phosphate-buffered
saline (PBS) and drying, the sections were incubated with DNase I
for 10 min in a humidified atmosphere at room temperature. The
fluorescein isothiocyanate-labeled TUNEL-positive cells were imaged
using fluorescence microscopy. DAPI (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to stain the nuclei. The average number
of apoptotic cells was calculated from five random fields with
Image-Pro Plus software (version 6.0; Media Cybernetics, Rockville,
MD, USA).
Immunohistochemical analysis
The 5-µm paraffin-embedded sections were
stained using the streptavidin-biotin complex immunohistochemistry
technique for Nrf2, NF-κB and p-IκBα detection. A positive signal
was visualized by a 3,3′-diaminobenzidine color reaction.
The nuclei were stained with hematoxylin. Brown staining in the
cytoplasm and the nucleus was considered an indicator of protein
expression. Two different sections of each specimen were examined
(original magnification, ×400; Olympus BX50; Olympus Optical). The
results were semi-quantitatively evaluated with
Image-Pro® Plus version 6.0 according to the optical
density values of protein expression. For this purpose, five fields
per slide were randomly selected by the viewer for evaluation.
Western blot analysis
Endochylema and cellular nuclear proteins were
extracted from frozen intestinal tissues with a nuclear extract kit
(cat. no. P0028; Beyotime Institute of Biotechnology, Haimen,
China) according to the manufacturer's protocol. Protein
concentration was determined using a BCA assay. Equal amounts (100
µg per lane) of protein were subjected to 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) at 100 v for
3 h. After electrophoresis, the proteins were transferred onto
polyvinylidene difluoride membranes (cat. no. 88520; Thermo Fisher
Scientific, Inc.) at 200 mA for 2 h. The membranes were incubated
overnight at 4°C with rabbit anti-mouse polyclonal antibodies to
Nrf2 (1:200 dilution), NF-κB (1:1,000 dilution), p-IκB-α (1:1,000
dilution), β-actin (1:2,000 dilution) and lamin B1 (1:200
dilution). After washing for three times with Tris-buffered saline
containing Tween-20, the membranes were incubated with the
corresponding goat anti-rabbit horseradish peroxidase-conjugated
secondary antibody (1:10,000 dilution) for 1 h at room temperature.
The intensity of the bands was detected using an Odyssey two-color
infrared laser imaging system and densitometry was performed using
Odyssey 1.0 software (both LI-COR Biosciences).
Statistical analysis
Values are expressed as the mean ± standard
deviation. GraphPad Prism 5.0 statistical software (GraphPad
Software Inc., La Jolla, CA, USA) was used to manage the data and
calculate the results. A statistical evaluation of the data was
performed by one-way or a two-way analysis of variance, followed by
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Nrf2 activation reduces IIR-induced
intestinal damage
To investigate the underlying mechanisms of the
effect of Nrf2 on IIR-induced injury, animals were pretreated with
Nrf2 antagonists and an Nrf2 activator prior to reperfusion-induced
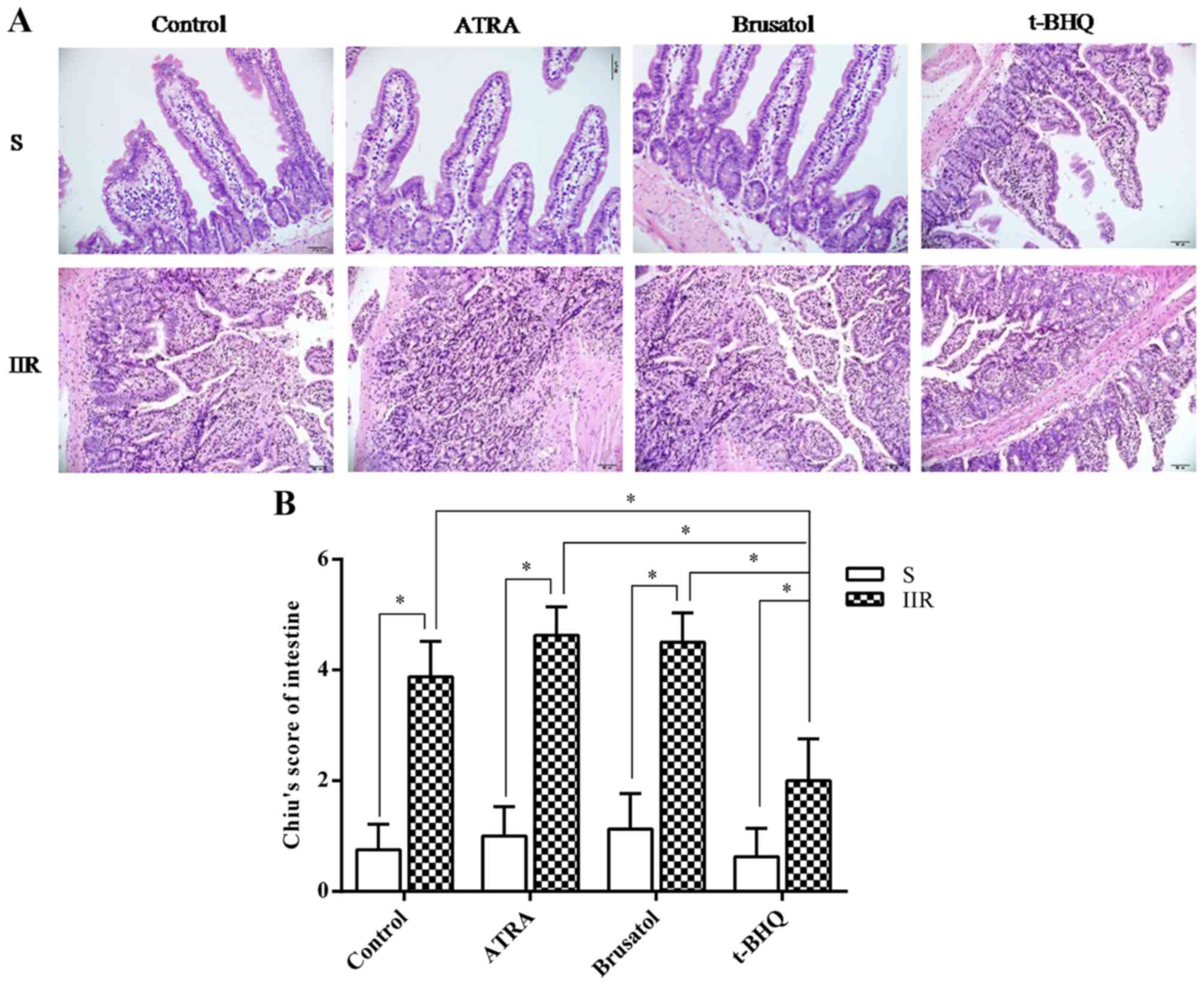
intestinal injury. H&E staining indicated that IIR induced
villous edema, inflammatory cell infiltration and capillary
congestion, and markedly increased the gap between epithelial cells
(Fig. 1A). These effects were
dramatically aggravated after administration of an Nrf2 antagonist
(ATRA or Brusatol) (Fig. 1). In
addition, IIR-induced intestinal injury was significantly
attenuated after treatment with the Nrf2 activator t-BHQ (Fig. 1). Chiu's scoring produced similar
results to those of H&E staining.
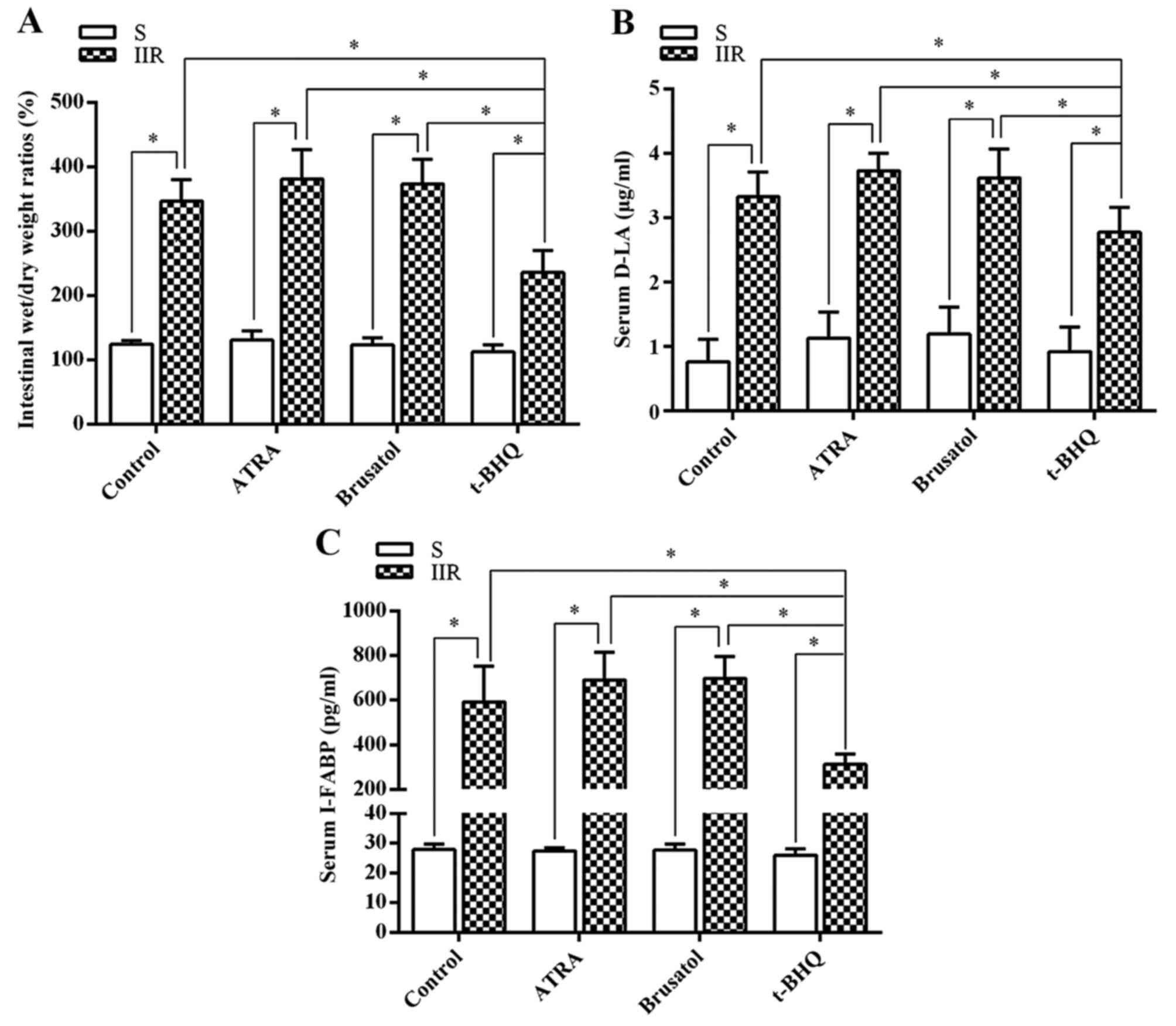
Next, intestinal permeability damage was assessed by
determining the intestinal wet/dry weight ratios (Fig. 2A). The intestinal wet/dry weight
ratios were significantly higher in the IIR group than in the S
group (P=0.0085). Compared with the IIR group, the intestinal
wet/dry weight ratio was significantly decreased in the group
pretreated with the Nrf2 activator t-BHQ (P=0.021). Furthermore,
the serum levels of D-LA (Fig.
2B) and I-FABP (Fig. 2C) were
assessed as biomarkers for the integrity of the intestinal
epithelium. Serum levels of D-LA and I-FABP were markedly increased
in the IIR group compared with those in the S group (P=0.0083 and
0.00009, respectively) and were significantly decreased in the
group pretreated with t-BHQ treatment (P=0.015 or 0.0003,
respectively, compared with the IIR group). However, the Nrf2
antagonists had no significant effect on serum D-LA or I-FABP.
Nrf2 regulates inflammatory cytokines in
the plasma and intestinal tissues after IIR
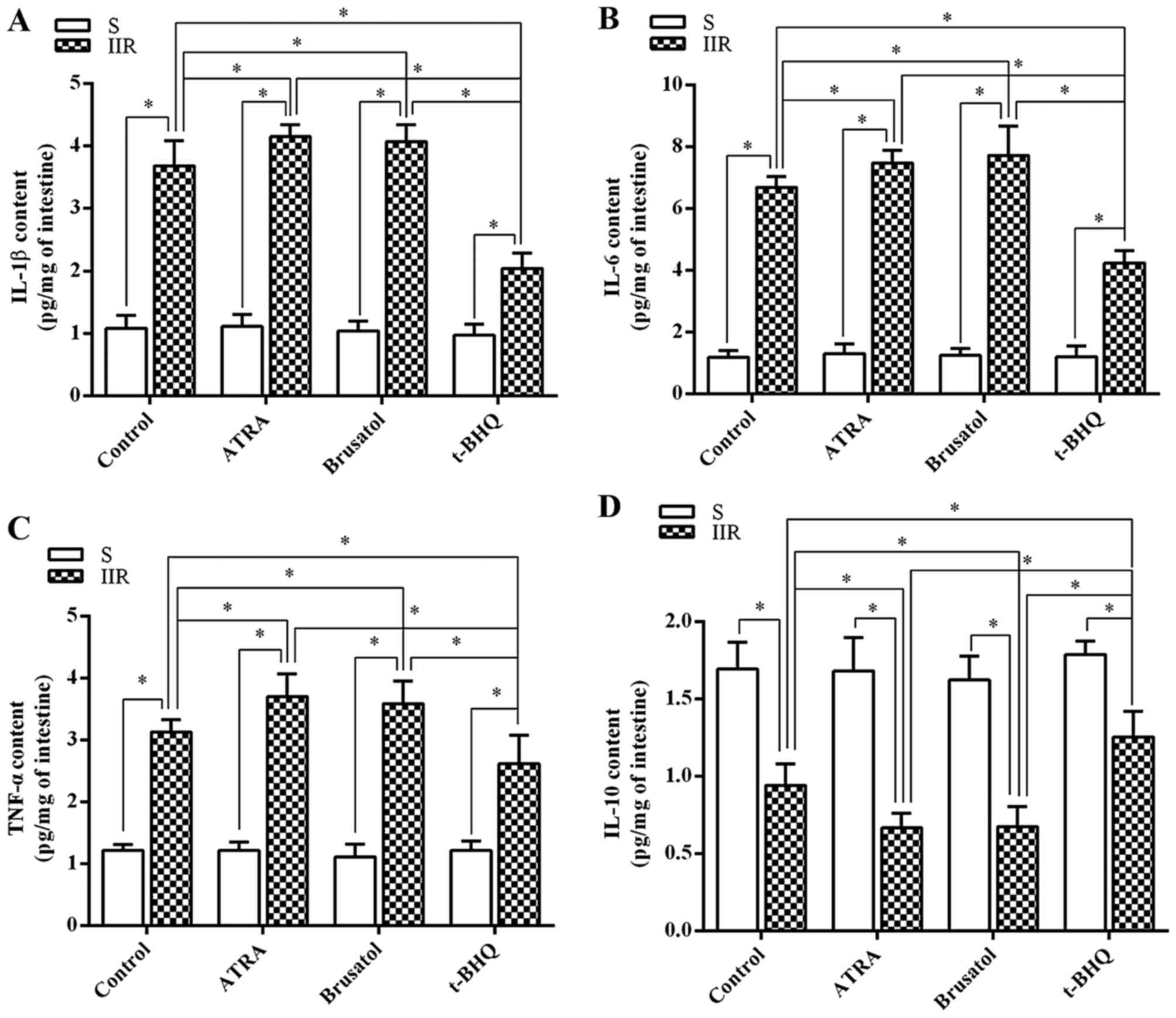
Next, the changes in inflammatory cytokine
expression in the intestine and serum were investigated. The levels
of tissue IL-1β, IL-6 and TNF-α in the IIR group were significantly
higher than those in the S group (P=0.021, 0.0076 and 0.033,
respectively) (Fig. 3A–C).
However, the tissue levels of IL-10 were markedly reduced in the
IIR group compared with those in the S group (P=0.044) (Fig. 3D). In addition, pretreatment with
ATRA or brusatol significantly aggravated the IIR-induced increases
in IL-1β, IL-6 and TNF-α levels, while further reducing IL-10
levels. The increases in the levels of IL-1β, IL-6 and TNF-α, and
the decrease of IL-10 induced by IIR were inhibited by pretreatment
with t-BHQ (P=0.032, 0.017, 0.026 and 0.023, respectively)
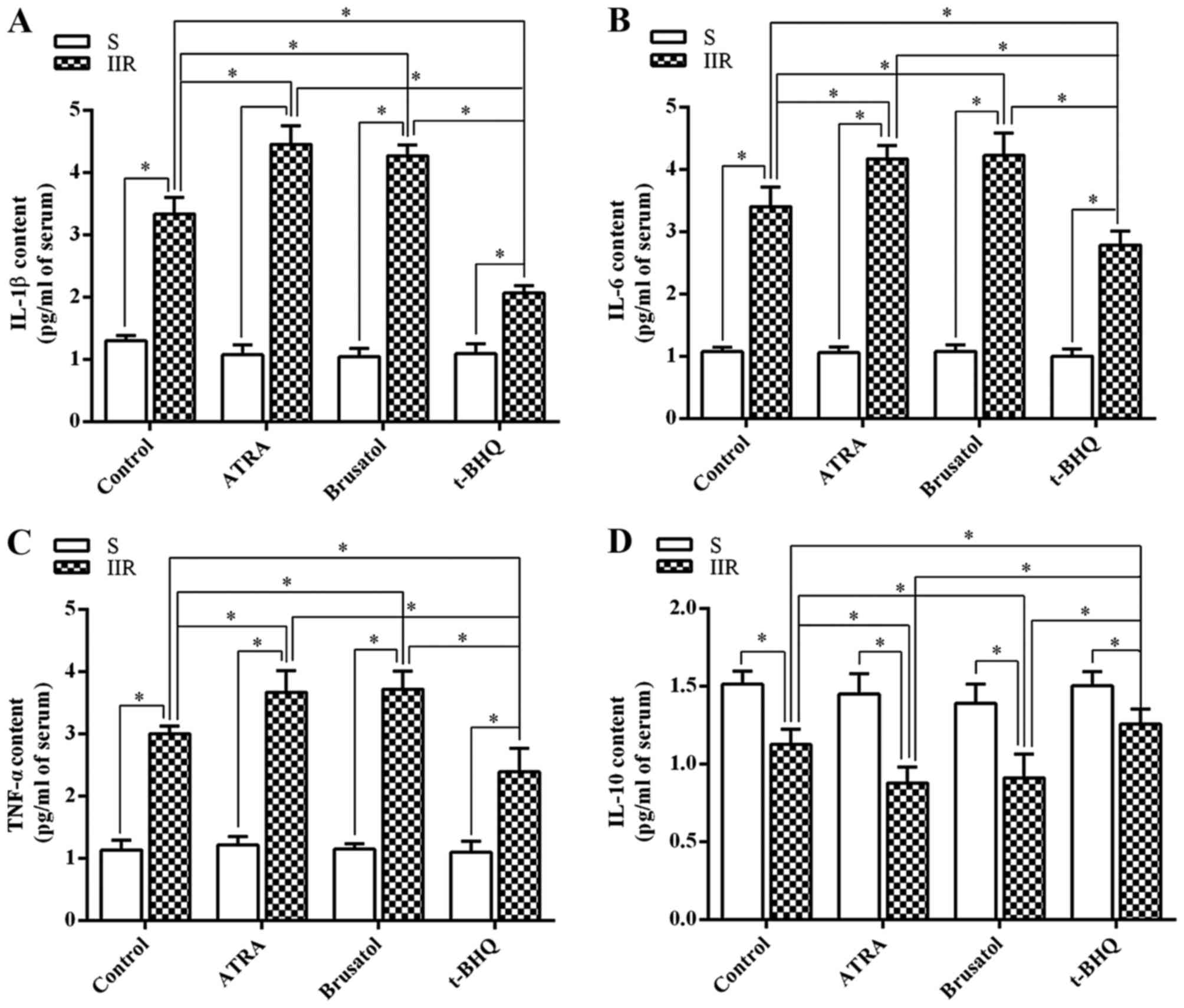
(Fig. 3). The changes in the
serum levels of inflammatory cytokines were consistent with those
in the intestinal tissue (Fig.
4).
Nrf2 activation attenuates IIR-induced
apoptosis
To further investigate the effects of Nrf2 on
apoptosis after IIR, the intestine was examined by TUNEL staining
(Fig. 5). The amount of
TUNEL-positive intestinal cells increased significantly after IIR
(P=0.017) (Fig. 5A and B).
Pretreatment with ATRA and brusatol aggravated IIR-induced
apoptosis in epithelial cells. Conversely, the IIR-induced
apoptosis of intestinal epithelial cells was inhibited by
pretreatment with Nrf2 activator t-BHQ (P= 0.008).
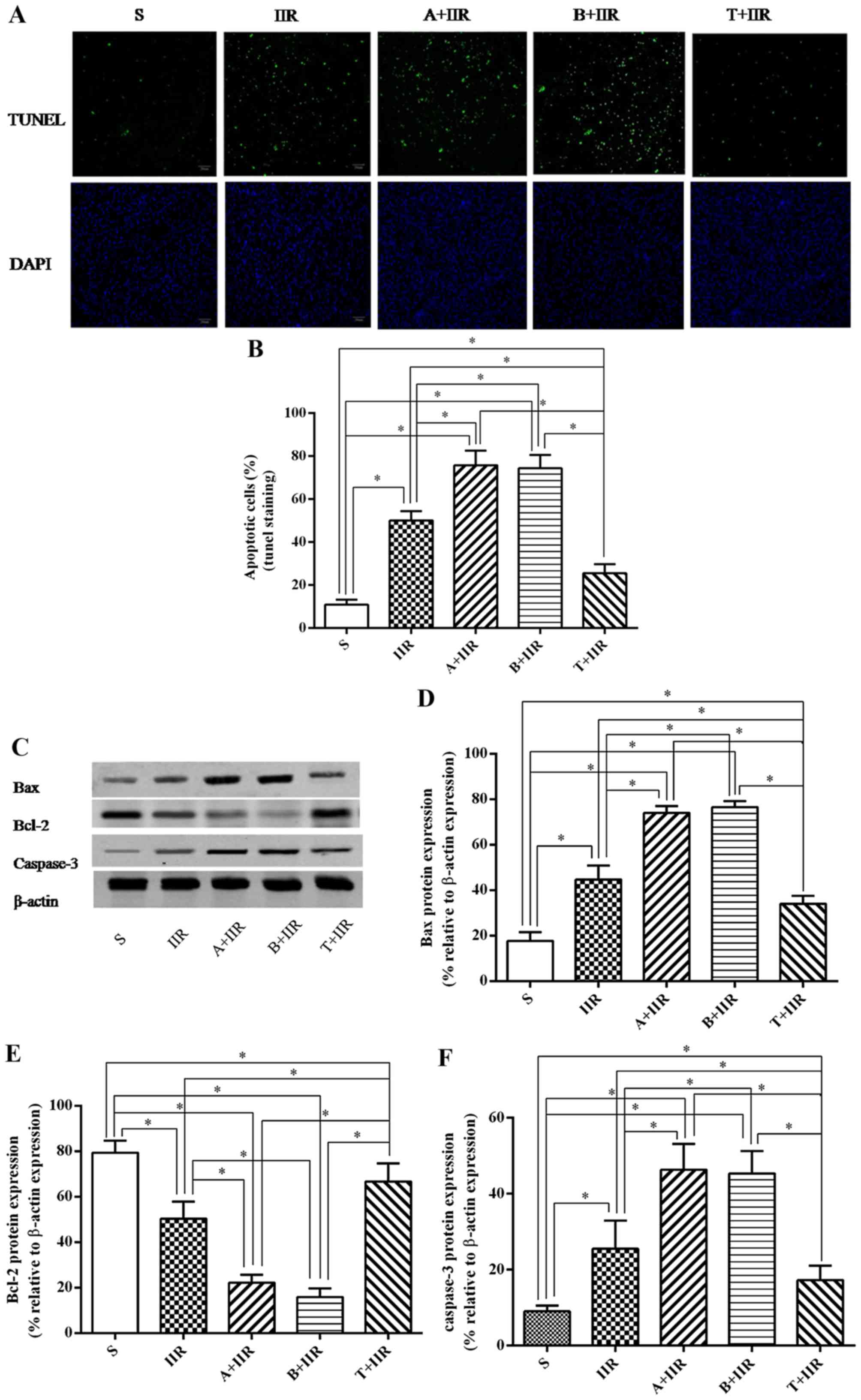 | Figure 5Effect of Nrf2 regulation on
IIR-induced apoptosis in intestinal epithelial tissue. (A and B)
Apoptosis in the intestine of animals from each group was detected
by a TUNEL assay. (A) Representative fluorescence microscopy images
(scale bar, 20 µm) and (B) quantified percentages of
TUNEL-stained cells in each group. (C–F) The expression of
apoptotic proteins was detected by western blot analysis. (C)
Representative western blot image and quantified expression levels
of (D) Bax, (E) Bcl-2 and (F) cleaved caspase-3. β-actin was used
as a loading control. Values are expressed as the mean ± standard
deviation (n=8). *P<0.05. S, sham surgery; IIR,
intestinal ischemia/reperfusion; A, all-trans retinoic acid;
B, brusatol; T, t-butylhydroquinone (Nrf2 activator); Nrf2, nuclear
factor erythroid 2-related factor 2; Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X protein; TUNEL, terminal deoxynucleotidyl
transferase deoxyuridinetriphosphate nick end labeling. |
The expression of apoptosis-associated proteins in
the intestine was then examined. It was observed that Bax and
cleaved caspase-3 were significantly increased after IIR treatment
(P=0.032 and 0.046, respectively) (Fig. 5C, D and F), which was markedly
exacerbated by pretreatment with Nrf2 antagonists ATRA (P=0.037 and
0.041, respectively) and brusatol (P=0.040 and 0.035, respectively)
(Fig. 5C, D and F). The
expression of Bcl-2 in the IIR group was markedly decreased
compared with that in the S group (P=0.028), and further
significant decreases were observed in the A+IIR and B+IIR groups
pretreated with the Nrf2 antagonists (P=0.029 and 0.033,
respectively, vs. IIR group) (Fig. 5C
and E). In addition, pretreatment with t-BHQ significantly
inhibited the IIR-induced increases in the expression of Bax
(P=0.041) (Fig. 5C and D) and the
levels of cleaved caspase-3 (P=0.037) (Fig. 5C and F), as well as the decrease
in the expression of Bcl-2 (P=0.022) (Fig. 5C and E).
Nrf2 activation inhibits the NF-κB
pathway
The Nrf2/ARE pathway is deeply involved in the
protection of organs from IIR injury. The present study evaluated
the expression of Nrf2 and inflammation-associated proteins in the
intestine by immunohistochemical staining and western blot
analysis. Positive immunohistochemical staining was indicated by
yellow-brown-stained granules. In the IIR group, protein expression
was mainly identified in the cytoplasm and the nuclei of intestinal
tissue cells. In the group pretreated with t-BHQ, cellular staining
for Nrf2 became lighter after IIR. In the IIR group, a large
proportion of the cytoplasm and nuclei of the intestinal tissue
cells appeared brownish-yellow or dark brown, and a large
proportion of cytoplasm and nuclei of the intestinal tissue cells
remained brownish-yellow or dark brown, indicating Nrf2 expression
in the group pretreated with t-BHQ prior to IIR (Fig. 6A). After IIR induction, the
cytoplasm and the nuclei of the tissues exhibited Nrf2 and NF-κB
expression in the epithelial lamina propria according to
immunohistochemical staining (Fig.
6A). Quantification of the staining revealed that the
expression of Nrf2 (P=0.034) (Fig.
6B) and NF-κB (P=0.029) (Fig.
6C) was significantly increased after IIR. Pretreatment with
ATRA did not affect the expression of Nrf2 in the cytoplasm and the
nuclei of intestinal cells compared with those in the IIR group,
while it remained significantly higher than that in the S group
(P=0.009) (Fig. 6A and B).
However, pretreatment with brusatol abolished the IIR-induced
expression with no significant difference compared with that in the
S group (P=0.098) (Fig. 6A and
B). The levels of NF-κB and p-IKBα in the IIR group were much
higher than those in the group pretreated with the Nrf2 antagonist
ATRA prior to IIR (P=0.025 and 0.022, respectively) (Fig. 6A, C and D). In the group
pretreated with t-BHQ, the accumulation of Nrf2 in the nuclei was
significantly increased compared with that in the IIR group
(P=0.0043) (Fig. 6A and B), while
the accumulation of NF-κB in the nuclei and the levels of p-IκBα in
the cytoplasm decreased significantly compared with those in the
IIR group (P=0.014 and 0.028, respectively) (Fig. 6A, C and D).
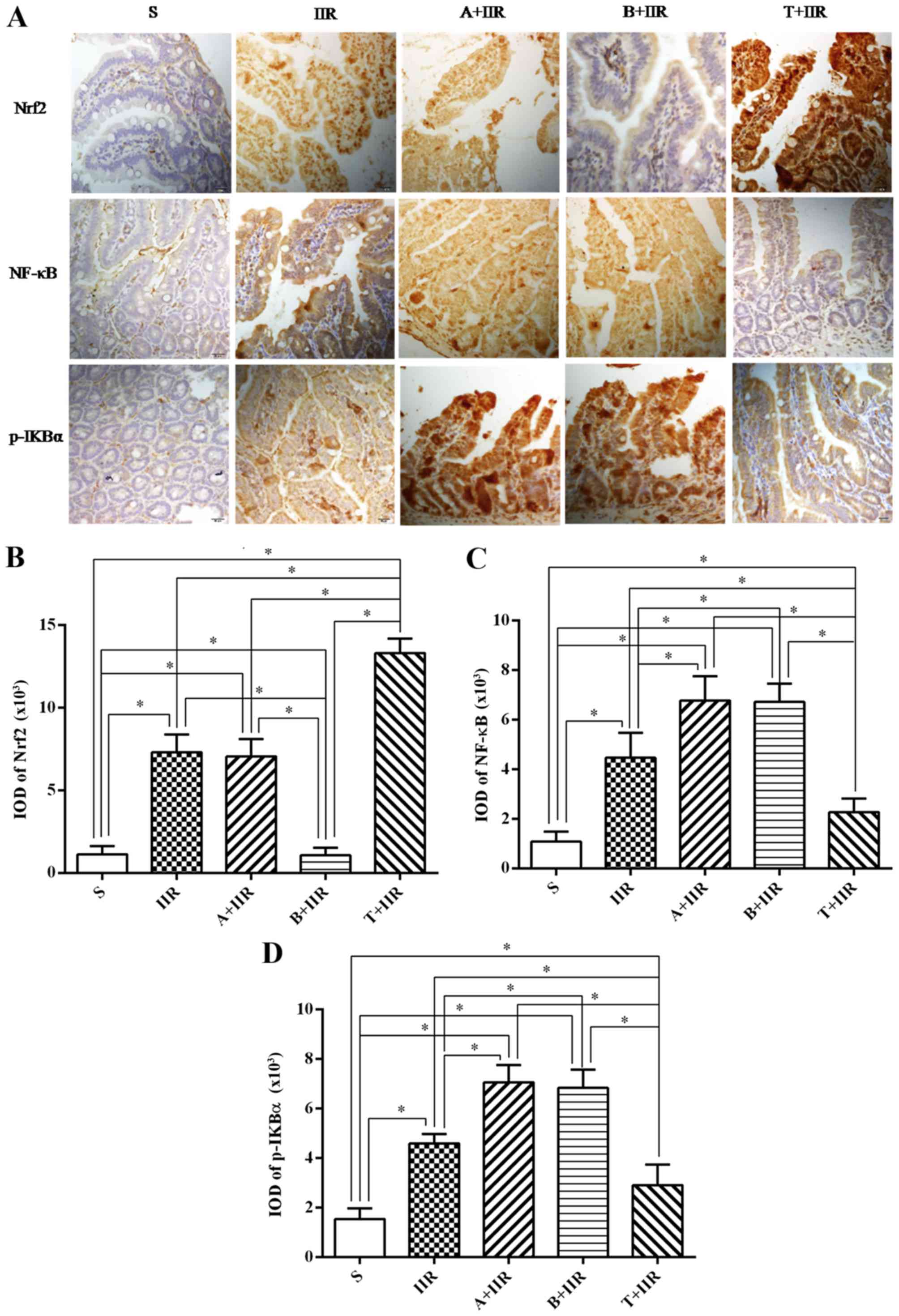 | Figure 6Nrf2 activation is involved in the
protection against IIR-induced apoptosis by inhibiting the NF-κB
pathway. (A) Representative immunohistochemical images with
staining performed using the streptavidin-biotin complex
immunohistochemistry technique. Positive staining was indicated by
brownish yellow or dark brown cytoplasm or nuclei. A large
proportion of the cytoplasm and nuclei of intestinal tissue cells
were stained for Nrf2 in the IIR group as well as in the T+IIR
group (scale bar, 20 µm). Quantified expression of (B) Nrf2,
(C) NF-κB and (D) p-IKBα. Values are expressed as the mean ±
standard deviation (n=8). *P<0.05. S, sham surgery;
IIR, intestinal ischemia/reperfusion; A, all-trans retinoic
acid; B, brusatol; T, t-butylhydroquinone (Nrf2 activator); Nrf2,
nuclear factor erythroid 2-related factor 2; NF-κB, nuclear
factor-κB; p-IKBα, phosphorylated inhibitor of NF-κB. |
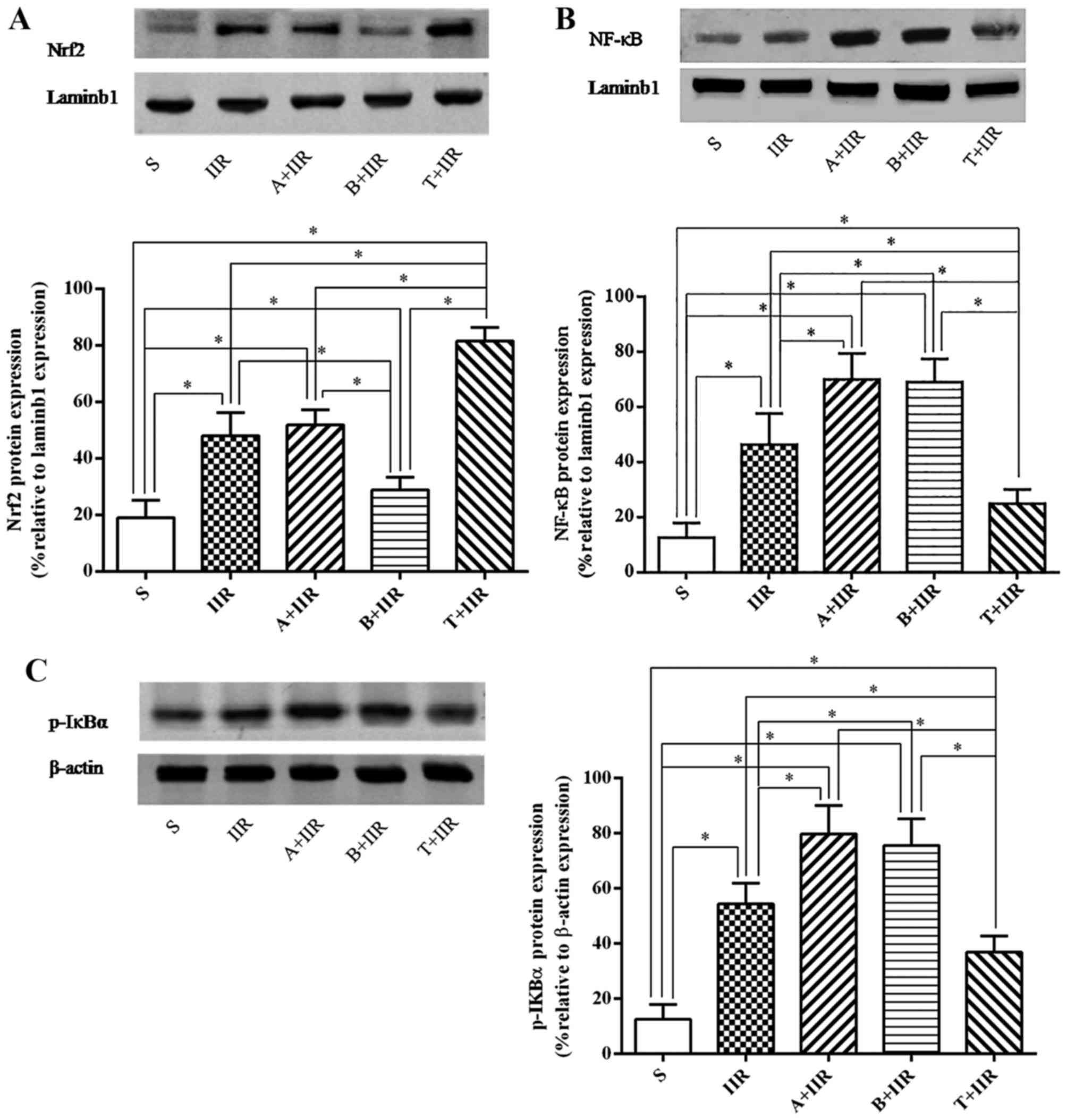
Furthermore, western blot analysis of the protein
expression of nuclear Nrf2 and NF-κB as well as the cytoplasmic
levels of p-IKBα provided similar results to those obtained by
immunohistochemistry (Fig.
7).
Discussion
The present study confirmed that IIR causes severe
intestinal tissue damage and cell apoptosis, in accordance with the
increased intestinal permeability and reduced integrity of the
intestinal epithelia. Furthermore, via Nrf2 antagonist or Nrf2
activator treatment, it was demonstrated that Nrf2 attenuated
IIR-induced apoptosis by regulating the systemic inflammatory
response. The results of the present study suggested that an
activator of Nrf2 had a beneficial effect against IIR-induced
intestinal apoptosis through exerting anti-inflammatory effects via
inhibiting the NF-κB pathway.
IIR injury is associated with a wide range of
pathological conditions in experimental models and clinical
conditions. The original tissue ischemia causes endothelial barrier
dysfunction and increased endothelial permeability (16). Subsequent reperfusion has various
consequences, including the activation of apoptosis (17) and the stimulation of the innate
and adaptive immune responses (18). A self-perpetuating signaling
cascade escalates to a vicious cycle of continuously increasing
intestinal permeability and bacterial translocation, a stronger
inflammatory response, apoptosis and eventual multiple organ
failure. The present study verified that occlusion of the SMA for
45 min followed by reperfusion for 2 h caused significant
intestinal injury in mice. This injury resulted in mucosal edema,
decreased epithelial cells, villi destruction, inflammatory cell
infiltration and a sharp increase in Chiu's score. All of these
observations are consistent with those of previous studies
(19).
The normal structure and function of the intestinal
mucosa is important in the prevention of the translocation of
bacteria and other noxious substances (20). Furthermore, the intestinal mucosa
is hypersensitive to IIR, which results in intestinal barrier
dysfunction. D-LA (21) is a
product that is released by numerous microfloras, while I-FABP
(22) is only released by damaged
intestinal epithelial cells. These factors are known as useful
biomarkers for intestinal barrier dysfunction. The molecules that
are released upon the disruption of epithelial integrity may be
measured and quantified. Upon IIR, significant increases in the
serum concentrations of D-LA and I-FABP were detected in the
present study, which strongly indicated mucosal integrity
impairment and intestinal barrier dysfunction.
Inflammation is an essential part of the innate
immune response that prevents tissue damage and helps tissue
healing in various ways. However, uncontrollable inflammation gives
rise to advanced tissue damage. An IIR challenge leads to the
translocation of bacteria and toxins (23), amplifying systemic inflammation
and apoptosis and resulting in SIRS and MODS. Ischemia induces the
rapid recruitment of inflammatory cells, and the cytokines produced
by these inflammatory cells further facilitate an inflammatory
status. Intensified inflammation also promotes apoptosis (24). Targeting the inflammatory response
is a crucial therapeutic strategy for the treatment of IIR injury.
The present results indicated that the proinflammatory cytokines
IL-1β, IL-6 and TNF-α sharply increased in the intestinal tissue
and serum after IIR injury, while the anti-inflammatory cytokine
IL-10 decreased. Furthermore, IIR markedly increased the number of
apoptotic cells, enhanced the expression of Bax and the levels of
cleaved caspase-3, and led to downregulation of the expression of
Bcl-2 in tissue.
Several signaling pathways are involved in
IIR-induced inflammation and apoptosis, reflecting the complexity
of the underlying mechanisms. The present study focused on two
important transcription factors, Nrf2 and NF-κB. As a prototypical
component of a proinflammatory signaling pathway, NF-κB induces the
transcription of numerous proinflammatory cytokines (TNF-α, IL-1β
and IL-6) and regulates anti-inflammatory cytokines (IL-10)
(25). Furthermore, the
auto-regulatory loop between NF-κB and proinflammatory cytokines
extensively aggravates the damaging effect of inflammation.
Activation of NF-κB leads to the production of several
proinflammatory cytokines (26),
including TNF-α and IL-1β, which in turn further induce the
activation of NF-κB. The present study indicated that IIR induced
NF-κB activation and significantly increased the tissue and serum
levels of TNF-α, IL-6 and IL-1β.
The Nrf2/ARE signaling pathway has a crucial role in
antioxidant and anti-inflammatory cellular responses (27). Under physiological conditions,
Nrf2 is retained in the cytoplasm by Keap1 and is degraded through
ubiquitination. Upon the infliction of oxidative stress, Nrf2
dissociates from Keap1, translocates to the nucleus and binds to
ARE. Therefore, Nrf2 initiates the transcription of genes that code
for phase II detoxifying enzymes, such as heme oxygenase-1 and
NAD(P)H:quinone oxidoreductase-1 (28). A recent study indicated that the
Nrf2/ARE pathway may also serve as an anti-inflammatory modulator
(29). Nrf2 activation has also
been reported to reduce organ damage and prevent inflammation from
hemolysis induction in sickle cell disease (30). Studies have confirmed that CO
released by tricarbonyldichlororuthenium (II) dimer mitigates
lipopolysaccharide-induced inflammation through the activation of
Nrf2 (31). The activation of
Nrf2 significantly reduces immune cell infiltration and decreases
the expression of the proinflammatory cytokines TNF-α, IL-1β and
IL-6 by inhibiting the NF-κB signaling pathway. In addition, Nrf2
depletion has been reported to enhance the inflammatory process
through the activation of NF-κB in the brain after traumatic brain
injury (32). Distinct patterns
of crosstalk between NF-κB and Nrf2 have been explored in different
cell types (11). The present
study indicated that the expression of intestinal Nrf2 was markedly
increased after IIR treatment. IIR also induced tissue damage, and
increased the expression of NF-κB and the levels of p-IκBα. These
observations suggested that IIR induces overexpression of Nrf2
through the innate immune response to counteract tissue injury, but
the protective effects of these factors are not sufficient to fully
protect the tissue. In the present study, two different potent
inhibitors, ATRA and brusatol, were applied, in order to provide
further evidence for the importance of Nrf2 in the protection from
IIR-induced injury. Administration of Nrf2 antagonists markedly
exacerbated intestinal injury, augmented the inflammatory response
and apoptosis, and increased the levels of NF-κB and p-IκBα. ATRA
inhibits the function of Nrf2 by stimulating the formation of
Nrf2:retinoic acid receptor α-containing complexes that do not bind
to ARE, while brusatol enhances Nrf2 degradation. Furthermore,
treatment with t-BHQ, an activator of Nrf2, was observed to
attenuate IIR-induced tissue injury, mitigate intestinal barrier
dysfunction, reduce proinflammatory cytokines and apoptotic
factors, and inhibit IκB kinase/IκB phosphorylation and NF-κB
nuclear translocation. NF-κB was also reported to inhibit Nrf2 at
the transcriptional level (11).
The present study indicated that activation of Nrf2 dramatically
mitigated pathological changes within the intestine and reduced the
inflammatory response and apoptosis by inhibiting the NF-κB
pathway, while Nrf2 antagonists have the opposite effect. All of
these results highlighted the importance of Nrf2 in the regulation
of the inflammatory response and apoptosis during IIR injury and
demonstrated the possibility of complex crosstalk between NF-κB and
Nrf2.
In conclusion, the present study indicated that Nrf2
has a critical role in the regulation of inflammation and apoptosis
induced by IIR. The beneficial effects of Nrf2 confer
anti-inflammatory properties against IIR-induced apoptosis,
potentially through the inhibition of the NF-κB pathway.
Acknowledgments
This study was supported by the Chinese Natural
Science Foundation (grant nos. 81671948, 81671891, 81401574 and
81400698).
Glossary
Abbreviations
Abbreviations:
|
IIR
|
intestinal ischemia reperfusion
|
|
SIRS
|
systemic inflammatory response
syndrome
|
|
MODS
|
multiple organ dysfunction
syndrome
|
|
IL
|
interleukin
|
|
TNF
|
tumor necrosis factor
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
ARE
|
antioxidant responsive elements
|
|
NF-κB
|
nuclear factor-κB
|
|
TUNEL
|
terminal deoxynucleotidyl transferase
deoxyuridinetriphosphate nick end labeling
|
|
ATRA
|
all-trans retinoic acid
|
|
SMA
|
superior mesenteric artery
|
|
D-LA
|
D-lactic acid
|
|
I-FABP
|
intestinal-type fatty acid-binding
protein
|
References
|
1
|
Huang CY, Hsiao JK, Lu YZ, Lee TC and Yu
LC: Anti-apoptotic PI3K/Akt signaling by sodium/glucose transporter
1 reduces epithelial barrier damage and bacterial translocation in
intestinal ischemia. Lab Invest. 91:294–309. 2011. View Article : Google Scholar
|
|
2
|
Gerlach UA, Atanasov G, Wallenta L, Polenz
D, Reutzel-Selke A, Kloepfel M, Jurisch A, Marksteiner M,
Loddenkemper C, Neuhaus P, et al: Short-term TNF-alpha inhibition
reduces short-term and long-term inflammatory changes
post-ischemia/reperfusion in rat intestinal transplantation.
Transplantation. 97:732–739. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim M, Park SW, Kim M, D'Agati VD and Lee
HT: Isoflurane post-conditioning protects against intestinal
ischemia-reperfusion injury and multiorgan dysfunction via
transforming growth factor-β1 generation. Ann Surg. 255:492–503.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crafts TD, Hunsberger EB, Jensen AR,
Rescorla FJ, Yoder MC and Markel TA: Direct peritoneal
resuscitation improves survival and decreases inflammation after
intestinal ischemia and reperfusion injury. J Surg Res.
199:428–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tian S, Guo R, Wei S, Kong Y, Wei X, Wang
W, Shi X and Jiang H: Curcumin protects against the intestinal
ischemia-reperfusion injury: Involvement of the tight junction
protein ZO-1 and TNF-α related mechanism. Korean J Physiol
Pharmacol. 20:147–152. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Komaravelli N, Tian B, Ivanciuc T,
Mautemps N, Brasier AR, Garofalo RP and Casola A: Respiratory
syncytial virus infection down-regulates antioxidant enzyme
expression by triggering deacetylation-proteasomal degradation of
Nrf2. Free Radic Biol Med. 88:391–403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gallorini M, Petzel C, Bolay C, Hiller KA,
Cataldi A, Buchalla W, Krifka S and Schweikl H: Activation of the
Nrf2-regulated antioxidant cell response inhibits HEMA-induced
oxidative stress and supports cell viability. Biomaterials.
56:114–128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Boyanapalli SS, Paredes-Gonzalez X,
Fuentes F, Zhang C, Guo Y, Pung D, Saw CL and Kong AN: Nrf2
knockout attenuates the anti-inflammatory effects of phenethyl
isothiocyanate and curcumin. Chem Res Toxicol. 27:2036–2043. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park JH, Choi JW, Ju EJ, Pae AN and Park
KD: Antioxidant and anti-inflammatory activities of a natural
compound, shizukahenriol, through Nrf2 activation. Molecules.
20:15989–16003. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao W, Sun Z, Wang S, Li Z and Zheng L:
Wnt1 participates in inflammation induced by lipopolysaccharide
through upregulating scavenger receptor A and NF-κB. Inflammation.
38:1700–1706. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wardyn JD, Ponsford AH and Sanderson CM:
Dissecting molecular cross-talk between Nrf2 and NF-κB response
pathways. Biochem Soc Trans. 43:621–626. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meng QT, Cao C, Wu Y, Liu HM, Li W, Sun Q,
Chen R, Xiao YG, Tang LH, Jiang Y, et al: Ischemic
post-conditioning attenuates acute lung injury induced by
intestinal ischemia-reperfusion in mice: Role of Nrf2. Lab Invest.
96:1087–1104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang XJ, Hayes JD, Henderson CJ and Wolf
CR: Identification of retinoic acid as an inhibitor of
transcription factor Nrf2 through activation of retinoic acid
receptor alpha. Proc Natl Acad Sci USA. 104:19589–19594. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ren D, Villeneuve NF, Jiang T, Wu T, Lau
A, Toppin HA and Zhang DD: Brusatol enhances the efficacy of
chemotherapy by inhibiting the Nrf2-mediated defense mechanism.
Proc Natl Acad Sci USA. 108:1433–1438. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiu CJ, McArdle AH, Brown R, Scott HJ and
Gurd FN: Intestinal mucosal lesion in low-flow states. I. A
morphological, hemodynamic, and metabolic reappraisal. Arch Surg.
101:478–483. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun Z, Wang X, Deng X, Lasson A, Wallén R,
Hallberg E and Andersson R: The influence of intestinal ischemia
and reperfusion on bidirectional intestinal barrier permeability,
cellular membrane integrity, proteinase inhibitors, and cell death
in rats. Shock. 10:203–212. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marques GMN, Rasslan R, Belon AR, Carvalho
JG, Felice Neto R, Rasslan S, Utiyama EM and Montero EF:
Pentoxifylline associated to hypertonic saline solution attenuates
inflammatory process and apoptosis after intestinal
ischemia/reperfusion in rats. Acta Cir Bras. 29:735–741. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang X, Bai H, Wang Y, Li J, Zhou Q, Cai
W, Han J, Zhu X, Dong M and Hu D: Deletion of regulatory T cells
supports the development of intestinal ischemia-reperfusion
injuries. J Surg Res. 184:832–837. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang Y, Zhou Z, Meng QT, Sun Q, Su W, Lei
S, Xia Z and Xia ZY: Ginsenoside Rb1 treatment attenuates pulmonary
inflammatory cytokine release and tissue injury following
intestinal ischemia reperfusion injury in mice. Oxid Med Cell
Longev. 2015:8437212015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schneider KM, Bieghs V, Heymann F, Hu W,
Dreymueller D, Liao L, Frissen M, Ludwig A, Gassler N, Pabst O, et
al: CX3CR1 is a gatekeeper for intestinal barrier integrity in
mice: Limiting steatohepatitis by maintaining intestinal
homeostasis. Hepatology. 62:1405–1416. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sheedy JR, Wettenhall RE, Scanlon D,
Gooley PR, Lewis DP, McGregor N, Stapleton DI, Butt HL and DE
Meirleir KL: Increased d-lactic Acid intestinal bacteria in
patients with chronic fatigue syndrome. In Vivo. 23:621–628.
2009.PubMed/NCBI
|
|
22
|
Khadaroo RG, Fortis S, Salim SY, Streutker
C, Churchill TA and Zhang H: I-FABP as biomarker for the early
diagnosis of acute mesenteric ischemia and resultant lung injury.
PLoS One. 9:e1152422014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Diebel ME, Diebel LN, Manke CW, Liberati
DM and Whittaker JR: Early tranexamic acid administration: A
protective effect on gut barrier function following
ischemia/reperfusion injury. J Trauma Acute Care Surg.
79:1015–1022. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Santamaría B, Ucero AC, Benito-Martin A,
Vicent MJ, Orzáez M, Celdrán A, Selgas R, Ruíz-Ortega M and Ortiz
A: Biocompatibility reduces inflammation-induced apoptosis in
mesothelial cells exposed to peritoneal dialysis fluid. Blood
Purif. 39:200–209. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ahmad SF, Attia SM, Bakheet SA, Zoheir KM,
Ansari MA, Korashy HM, Abdel-Hamied HE, Ashour AE and Abd-Allah AR:
Naringin attenuates the development of carrageenan-induced acute
lung inflammation through inhibition of NF-κB, STAT3 and
pro-inflammatory mediators and enhancement of IκBα and
anti-inflammatory cytokines. Inflammation. 38:846–857. 2015.
View Article : Google Scholar
|
|
26
|
Fan B, Dun SH, Gu JQ, Guo Y and Ikuyama S:
Pycnogenol attenuates the release of proinflammatory cytokines and
expression of perilipin 2 in lipopolysaccharide-stimulated
microglia in part via inhibition of NF-κB and AP-1 activation. PLoS
One. 10:e01378372015. View Article : Google Scholar
|
|
27
|
Wang Y, Wang B, Du F, Su X, Sun G, Zhou G,
Bian X and Liu N: Epigallocatechin-3-gallate attenuates oxidative
stress and inflammation in obstructive nephropathy via NF-κB and
Nrf2/HO-1 signalling pathway regulation. Basic Clin Pharmacol
Toxicol. 117:164–172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li L, Dong H, Song E, Xu X, Liu L and Song
Y: Nrf2/ARE pathway activation, HO-1 and NQO1 induction by
polychlorinated biphenyl quinone is associated with reactive oxygen
species and PI3K/AKT signaling. Chem Biol Interact. 209:56–67.
2014. View Article : Google Scholar
|
|
29
|
Park SY, Kim YH and Park G: Cucurbitacins
attenuate microglial activation and protect from neuroinflammatory
injury through Nrf2/ARE activation and STAT/NF-κB inhibition.
Neurosci Lett. 609:129–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Keleku-Lukwete N, Suzuki M, Otsuki A,
Tsuchida K, Katayama S, Hayashi M, Naganuma E, Moriguchi T, Tanabe
O, Engel JD, et al: Amelioration of inflammation and tissue damage
in sickle cell model mice by Nrf2 activation. Proc Natl Acad Sci
USA. 112:12169–12174. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qin S, Du R, Yin S, Liu X, Xu G and Cao W:
Nrf2 is essential for the anti-inflammatory effect of carbon
monoxide in LPS-induced inflammation. Inflamm Res. 64:537–548.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pan H, Wang H, Wang X, Zhu L and Mao L:
The absence of Nrf2 enhances NF-κB-dependent inflammation following
scratch injury in mouse primary cultured astrocytes. Mediators
Inflamm. 2012:2175802012. View Article : Google Scholar
|