Introduction
Mesenchymal stem cells (MSCs) are non-hematopoietic,
self-renewing cells that are capable of clone-forming and
multilineage differentiation (1).
MSCs have been detected in numerous adult tissues, including bone
marrow, skeletal muscles, adipose tissues, synovial fluid and
synovium (2–5). The presence and characteristics of
MSCs in synovium specimens were first reported by De Bari et
al (6) and have been studied
extensively in recent years (7–13).
Synovium specimen-derived MSCs (SSMSCs) have a higher proliferative
capacity and chondrogenic potential than MSCs derived from other
sources; therefore, these cells are regarded as a promising cell
source for MSC-based therapeutic strategies used to treat cartilage
damage (5,11,14–17).
Generally, synovium specimens are obtained through
surgery, including open surgery or arthroscopic surgical procedures
(18–21). Previous studies have demonstrated
that MSCs can be isolated from surgery-obtained synovial specimens
(SSSs) using the same protocol as that employed for synovial
fibroblast cultivation. These cells exhibit ultrastructural and
morphological features similar to those of type B synoviocytes
(6,22). However, SSS cells (SSSCs) exhibit
heterogeneity. For example, Harvanova et al (19) reported that 40–50% of SSSCs are
cluster of differentiation (CD)105+ subpopulation cells
prior to immunomagnetic separation. These data suggest that SSMSCs
correspond to a subset of adherent cells in SSSs.
SSSs generally consist of at least two anatomically
distinct layers: The synovium (intima) and the underlying layer
(subintima). However, since there is currently no effective method
for the separation of these two tissue layers, SSMSCs reported in
previous studies were not entirely derived from the intima
(5,6,9).
Furthermore, no specific marker of synovial MSCs (SMSCs), which are
derived from the intima only, has been identified to date.
Therefore, the characteristics of SMSCs remain poorly
understood.
The present study isolated and characterized
synovial fragments (SFs) present in synovial fluid dilutions
extracted from patients with temporomandibular joint (TMJ)
osteoarthrosis. These synovial fluid-derived SFs consisted of
several cell layers, indicating that they originated from the
intima. Subsequently, the histological characteristics of SFs were
compared with those of SSSs. Following isolation and expansion
in vitro, the characteristics of both cell types were
investigated, including in vitro proliferation and
morphology, surface marker expression, and multilineage
differentiation capabilities.
Materials and methods
Ethics statement
The present study was approved by the Institutional
Ethics Board of the Hospital of Stomatology, Sun Yat-sen University
(Guangzhou, China). Written informed consent was obtained from all
subjects.
Collection of SFs and SSSs
SFs were collected, between October 2014 and April
2016, during TMJ arthrocentesis from patients with TMJ
osteoarthrosis that showed no response to conservative treatment.
Briefly, a no. 8 needle was punctured into the upper joint
compartment. A total of 2.0 ml lidocaine was infused and then
withdrawn. Diluted synovial fluid samples were collected from ~800
patients (age, 16–68 years), and SFs were obtained from 17 of these
samples. These 17 patients (age, 18–61 years) had no other systemic
diseases; among these patients, 3 were male and 14 were female. In
addition, 8 SSSs (~0.3×0.5 cm) were obtained aseptically from
patients with TMJ osteoarthrosis at the time of surgical
debridement treatment for osteoarthrosis or joint disk perforation.
The 8 donors (age, 25–50 years) had no other systemic diseases;
among these patients, 1 was male and 7 were female.
Culture of human SFCs and SSSCs
SFs from the synovial fluid were washed three times
and were then digested with 4 mg/ml type I collagenase for 2.5 h at
37°C. The specimens were dispersed by pipetting and then filtered
through a 200-mesh screen. Cells were centrifuged at 300 × g at
room temperature for 5 min and cultured with complete culture
medium [α-minimum essential medium (α-MEM)] supplemented with 10%
fetal bovine serum (FBS) and 1X GlutaMAX (all Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in 5% CO2.
The SSSCs were isolated and cultured in the same manner as the
SFCs.
Surface antigen expression profile
A total of 3 SFs and 6 SSSs samples were employed
for surface antigen expression analysis. For surface marker
detection, ~300,000 dissociated cells were collected. Following
incubation with primary antibodies or isotype control antibodies
for 30 min, the cells were centrifuged at 300 × g at room
temperature for 8 min. The supernatant was discarded prior to
resuspension of the cells. Flow cytometric analysis was performed
using an FC 500 flow cytometer (Beckman Coulter, Miami, FL, USA),
and the results were analysed using MXP Software version 2.0
(Beckman Coulter). The antibodies used are listed in Table I. Cells obtained from 3 SFs and 3
SSSs (samples 4–6) were used for subsequent experiments.
 | Table IAntibodies used in flow
cytometry. |
Table I
Antibodies used in flow
cytometry.
| Antibody | Dilution | Supplier |
|---|
| Peridinin
chlorophyll protein-Cy5.5-conjugated anti-human CD105 | 1:20 | BD
Biosciencesa |
|
Allophycocyanin-conjugated anti-human
CD73 | 1:20 | BD Biosciences |
| Fluorescein
isothiocyanate-conjugated anti-human CD90 | 1:20 | BD Biosciences |
|
Phycoerythrin-conjugated anti-human
CD44 | 1:20 | BD Biosciences |
|
Phycoerythrin-conjugated anti-human
CD45/CD34 | 1:20 | BD Biosciences |
|
Phycoerythrin-conjugated anti-human
CD45/CD34 CD11b/CD19/HLA-DR isotype control | 1:20 | BD Biosciences |
Cell proliferation assay
SFCs and SSSCs derived from 3 samples were mixed and
seeded in 96-well plates at a density of 800 cells/well at passage
4.
Cell proliferation was evaluated using Cell Counting
kit-8 (Shanghai Yeasen Biotechnology Co., Ltd., Shanghai, China).
The reagent was employed with the concentration of 10% per well.
Culture medium with the reagent served as a blank control. The
optical density (OD) of the supernatant was measured using a
microplate reader (Infinite 200; Tecan Group, Ltd., Männedorf,
Switzerland) after a 2-h incubation at 37°C. Repeated measurements
(n=3) were conducted at each time point. The population doubling
(PD) and doubling time (DT) were evaluated using the following
formulas: PD = (lnN − lnN0)/ln2 and DT = T/PD. N,
ODcells − ODblank control at the end point;
N0, ODcells − ODblank control at the initial
time point; T, time interval; lnN=log(e,N).
Colony-forming assay for SFCs and
SSSCs
SFCs and SSSCs (passage 4) were plated at a density
of 5 cells/cm2. After culturing in complete medium for 2
weeks, the cells were fixed in 4% paraformaldehyde for 10 min at
room temperature and were then stained with 0.5% crystal violet for
counting.
Differentiation of isolated SFCs and
SSSCs
Osteogenic differentiation
Cells were seeded at a density of 5,000
cells/cm2 in 6-well plates and were induced to
differentiate the next day. The differentiation medium consisted of
high-glucose Dulbecco's modified Eagle's medium (H-DMEM) containing
10% FBS (both Gibco; Thermo Fisher Scientific, Inc.), 10 mM sodium
β-glycerophosphate (Santa Cruz Biotechnology, Inc., Dallas, TX,
USA), 100 nM dexamethasone (MP Biomedicals, LLC, Santa Ana, CA,
USA) and 50 μg/l ascorbic acid-2-phosphate (Wako Pure
Chemical Industries, Ltd., Osaka, Japan).
Adipogenic differentiation
Cells were seeded at a density of 5,000
cells/cm2 in 6-well plates and were induced to
differentiate the next day. The medium consisted of H-DMEM
containing 10% FBS (both Gibco; Thermo Fisher Scientific, Inc.),
200 mM indomethacin (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), 0.5 mM isobutyl methylxanthine, 1 mM dexamethasone and 10
mg/ml insulin (both MP Biomedicals, LLC). The medium was replaced
every 3 days.
Chondrogenic differentiation
Approximately 300,000 cells were collected in a
15-ml centrifuge tube and were centrifuged at 300 × g at room
temperature for 5 min. Subsequently, cells were resuspended in 450
μl medium consisting of H-DMEM, 1X
insulin-transferrin-selenium-sodium pyruvate (both Gibco; Thermo
Fisher Scientific, Inc.), 100 nM dexamethasone (MP Biomedicals,
LLC), 50 mM ascorbic acid, 40 mg/ml proline (both Sigma-Aldrich;
Merck KGaA) and 10 ng/ml transforming growth factor-β1 (PeproTech,
Inc., Rocky Hill, NJ, USA). After a 10 min centrifugation at 450 ×
g at room temperature, the cells were incubated at 37°C in an
atmosphere containing 5% CO2.
Evaluation of osteogenic, adipogenic, and
chondrogenic differentiation
After a 4-week osteogenic induction, the cells were
washed with PBS and were fixed with 4% paraformaldehyde at room
temperature for 10 min. Subsequently, cells were stained with fresh
0.1% Alizarin Red S solution for 30 min at 37°C and examined under
an inverted phase contrast microscope (Axiovert 40; Carl Zeiss AG,
Oberkochen, Germany).
Adipogenesis was assessed by Oil Red O staining
after 4 weeks of induction. The cells were washed and fixed as
aforementioned. Subsequently, 0.3% Oil Red O solution was used to
stain the cells for 150–180 sec. The cells were then examined under
an inverted phase contrast microscope (Axiovert 40; Carl Zeiss
AG).
Histological staining was employed to assess
chondrogenic differentiation after a 3-week induction. Cartilage
nodules formed by the cells were fixed with 4% paraformaldehyde at
4°C overnight and were then embedded in paraffin. The blocks were
cut into 5 μm sections. The expression levels of collagen
type II were then detected. Sections were incubated with rabbit
anti-human collagen type II antibodies (Sigma-Aldrich; dilution,
1:80; cat. no. SAB4500366) in blocking buffer (10% goat serum
sealant, cat. no. SL038; Solarbio, Beijing, China) for 16 h at 4°C.
Biotinylated goat anti-rabbit immunoglobulin G (cat. no. SA1022;
Wuhan Boster Biological Technology, Ltd., Wuhan, China) was used as
a secondary antibody incubating at 37°C for 30 min and was detected
using streptavidin-biotin complex reagent (cat. no. SA1022; Wuhan
Boster Biological Technology, Ltd.). The staining was visualized
with 3,3′-diaminobenzidine (cat. no. AR1022; Wuhan Boster
Biological Technology, Ltd.) and was observed under a light
microscope (Axioskop 40; Carl Zeiss AG). Sections incubated without
the primary antibody served as a control.
Sulfated glycosaminoglycan (GAG) assays were
performed to determine the levels of GAG. Chondrogenic nodules were
digested overnight at 56°C in 50 μg/ml proteinase K solution
Santa Cruz Biotechnology, Inc.) diluted in 100 mM
Na2HPO4 (pH 8.0), followed by inactivation
for 10 min at 90°C. After centrifugation, 500 μl working
1,9-dimethylmethylene blue (DMMB) solution (Santa Cruz
Biotechnology, Inc.; 100 ml 1M GuHCl, 1 g sodium formate, 1 ml 98%
formic acid, 25 ml 0.64‰ DMMB ethanol solution completed to 500 ml
with distilled water was defined as solution A; 100 ml 1M GuHCl, 1
g sodium formate, 1 ml 98% formic acid, 25 ml 100% ethanol
completed to 500 ml with distilled water was defined as solution B;
working DMMB solution was made by rapidly mixing solution A with
solution B) was added to 50 μl treated sample or standard
sample. Subsequently, the samples were mixed vigorously for 30 min
and centrifuged for 10 min at 12,000 × g at 4°C. Once the
supernatant was discarded, 1 ml DMMB decomplexation solution [50 mM
sodium acetate (pH 6.8) added with 10% methyl alcohol, 4M GuHCl]
was added. After a further 30 min of agitation, absorbance was
examined at 656 nm using a microplate reader (Infinite 200; Tecan
Group, Ltd.). Double-stranded DNA, which was obtained from the
chondrogenic nodules digestion fluid, as detected by Quant-iT
PicoGreen dsDNA reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), was used as an endogenous control.
Hematoxylin and eosin (HE) staining of SF
and SSS sections and histological immunostaining of SSS
sections
Sections were stained with Mayer's hematoxylin for
15 min and with eosin for 1 min. After gradient dehydration and
clearing, the sections were mounted and observed under a light
microscope (Axioskop 40; Carl Zeiss AG). Histological
immunostaining of SSS sections was performed in the same manner as
mentioned above. Mouse anti-human CD105 antibody (cat. no. ab11414;
dilution, 1:200; Abcam, Cambridge, UK) was used as the primary
antibody and biotinylated goat anti-mouae immunoglobulin G (cat.
no. A1001; Wuhan Boster Biological Technology, Ltd.) was used as
the secondary antibody.
Evaluation of gene expression by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen: Thermo Fisher Scientific, Inc.) and cDNA was
synthesized using a Transcriptor First Strand cDNA Synthesis kit
(Roche Diagnostics, Basel, Switzerland) according to the
manufacturers' protocols. qPCR analyses were performed using a
LightCycler® 480 Probes Master system (Roche
Diagnostics) with an initial denaturation at 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 60 sec. A
melting curve analysis was then performed. GAPDH was used as an
internal control. Relative mRNA expression levels were evaluated
using the following formula: (Cqtarget gene −
Cqgapdh) sample − (Cqtarget gene −
Cqgapdh)control (23).PCR primer sequences are listed in
Table II.
| Table IIOligonucleotide primers used in
quantitative polymerase chain reaction. |
Table II
Oligonucleotide primers used in
quantitative polymerase chain reaction.
| Gene | Primer sequence
(5′-3′) |
|---|
| GAPDH | F:
GACAGTCAGCCGCATCTTCT |
| R:
TTAAAAGCAGCCCTGGTGAC |
| RUNX-2 | F:
TCAACGATCTGAGATTTGTGGG |
| R:
GGGGAGGATTTGTGAAGACGG |
| OCN | F:
CCACCGAGACACCATGAGAG |
| R:
TCAGCCAACTCGTCACAGTC |
| ALP | F:
ACCATTCCCACGTCTTCACATTTG |
| R:
AGACATTCTCTCGTTCACCGCC |
| PPARG2 | F:
GCAAACCCCTATTCCATGCTG |
| R:
CACGGAGCTGATCCCAAAGT |
| LPL | F:
CAAGAGTGAGTGAACAAC |
| R:
AATTATGCTGAAGGACAAC |
| SOX-9 | F:
ACACACAGCTCACTCGACCTTG |
| R:
AGGGAATTCTGGTTGGTCCTCT |
| COL-2 | F:
GGCAATAGCAGGTTCACGTACA |
| R:
CGATAACAGTCTTGCCCCACTT |
Statistical analysis
Data were statistically analysed using SPSS 10.0
software (SPSS, Inc., Chicago, IL, USA). Experiments were repeated
at least 3 times and numerical data are presented as the mean ±
standard deviation. Student's t-test was used to analyze results
for 2 independent groups. Comparisons between multiple groups were
conducted using one-way analysis of variance (ANOVA) followed by
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Characterization of SFs, SSSs, SFCs and
SSSCs expanded in vitro
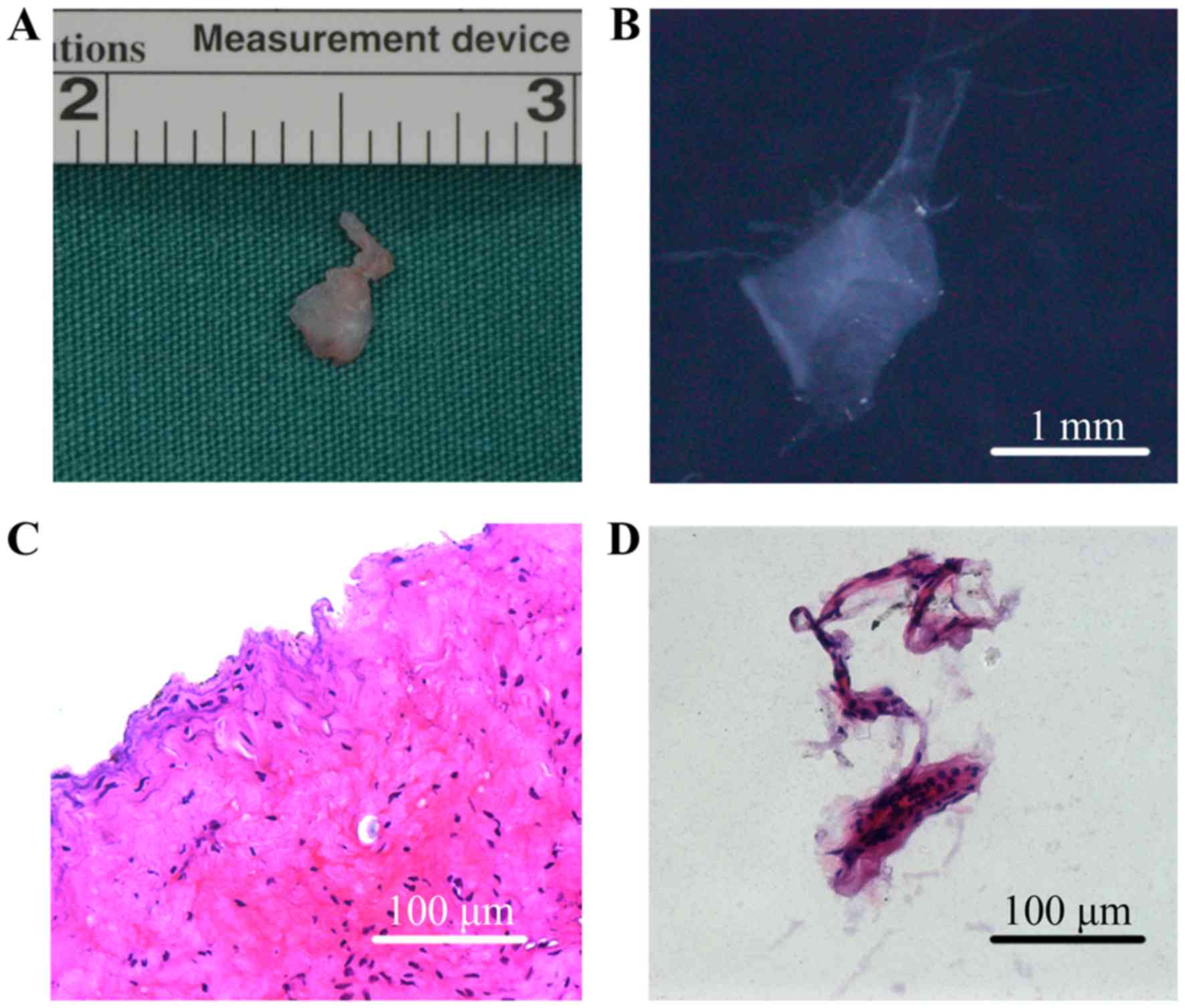
The SFs collected from patients with TMJ
osteoarthrosis were membranous and translucent, whereas SSSs were
masses of tissue (Fig. 1A and B).
HE staining revealed that SSSs exhibited a more complex
histological structure, containing intima and subintima (Fig. 1C), whereas the SFs were formed of
several layers of cells only, indicating that they were obtained
from the intima (Fig. 1D).
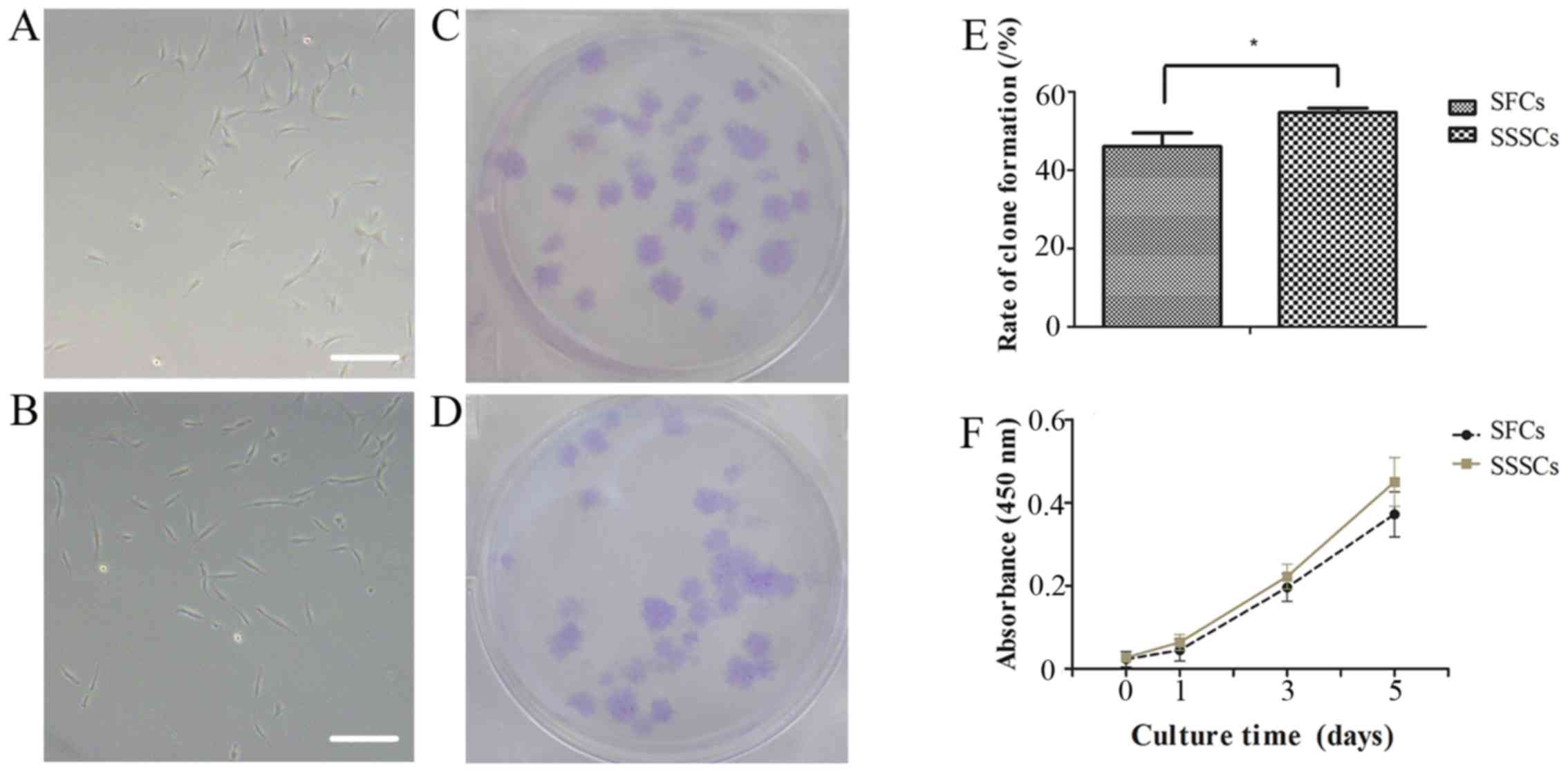
Adherent cells were obtained from SFs and SSSs. The
SFCs and SSSCs both exhibited a typical fibroblastic spindle shape
(Fig. 2A and B). In addition,
both cell types exhibited clone-forming potential (Fig. 2C and D). The clone-forming rate of
SFCs was slightly lower than that of SSSCs (Fig. 2E; P=0.014).
Cell proliferation curves demonstrated that SFCs
exhibited a growth pattern similar to that of SSSCs (Fig. 2F). The PD and DT of SFCs were
2.58±1.01 and 30.73±5.90 h, respectively, whereas the PD and DT of
SSSCs were 3.18±1.38 and 24.88±5.07 h, respectively, at passage
4.
Surface marker assays for SFCs and
SSSCs
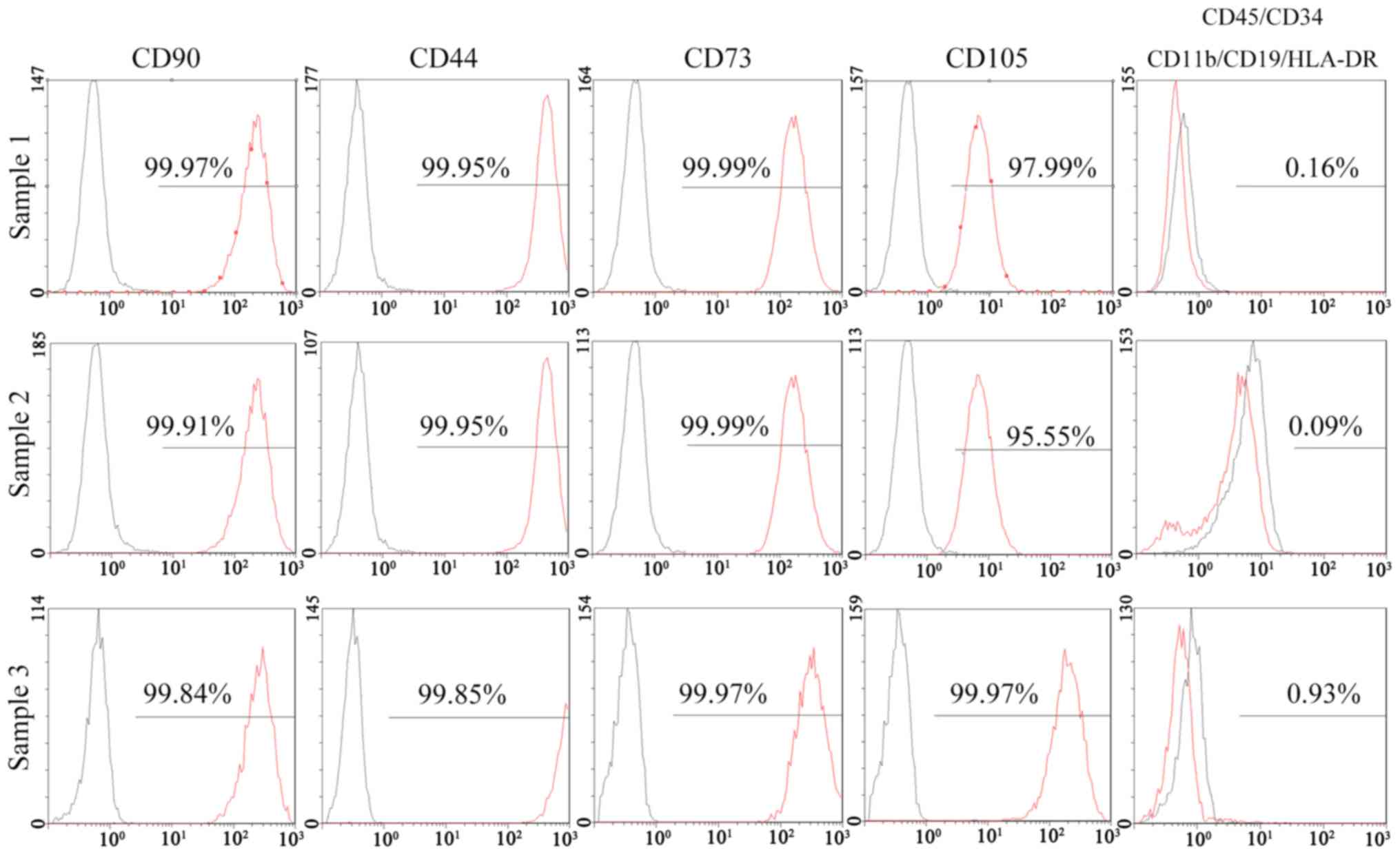
The results of a flow cytometric analysis indicated
that >95% of SFCs derived from all 3 SFs expressed positive
markers of MSCs, including CD90, CD44, CD73 and CD105. Negative
markers of MSCs: CD11b, CD19, CD34, CD45 and human leukocyte
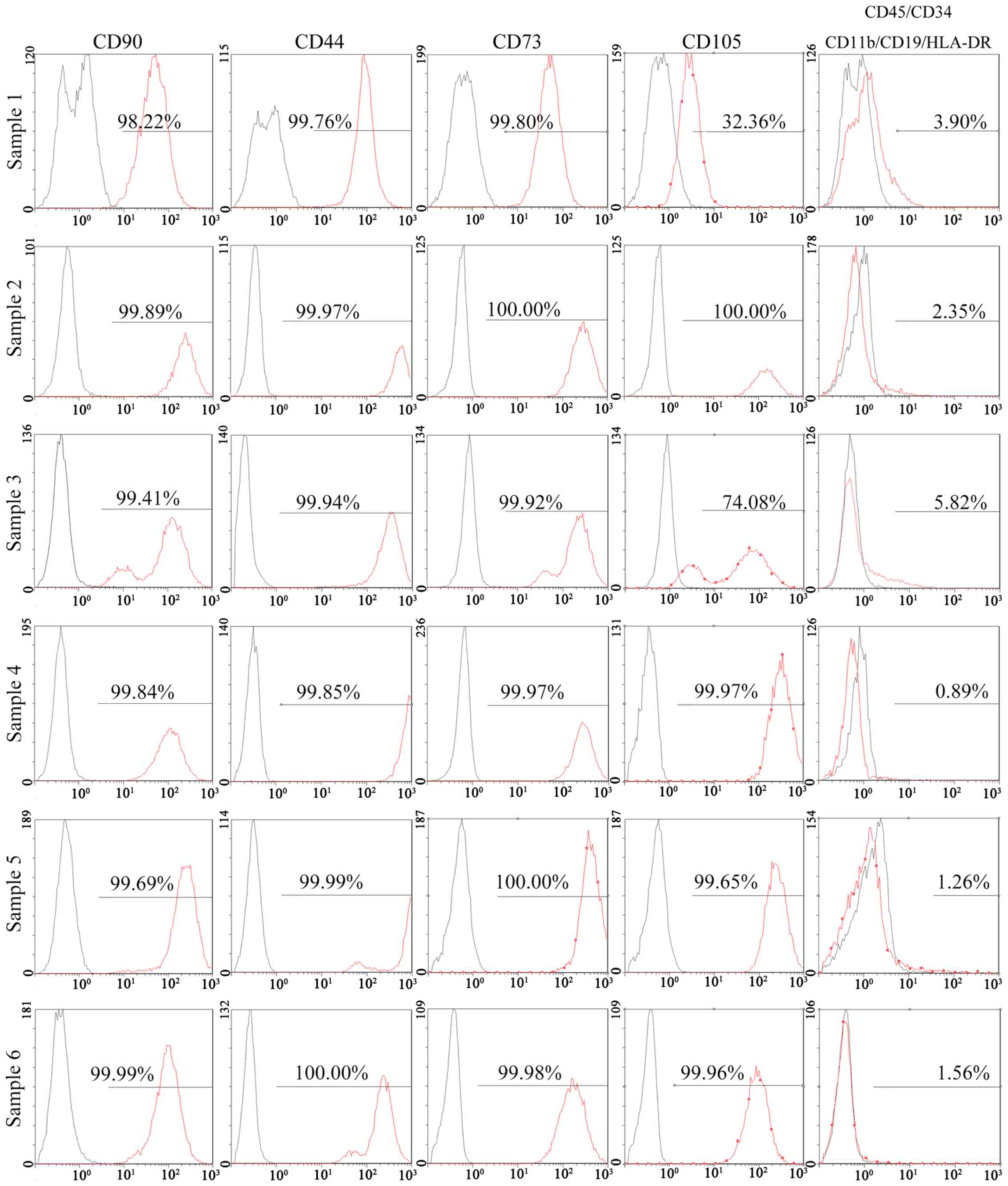
antigen (HLA)-DR, were positive in <2% of cells (Fig. 3). In addition, 95% of SSSCs
derived from all 6 SSSs expressed CD90, CD44 and CD73. However, the
percentage of CD105+ cells in two of the SSSs was much
lower (32.36 and 74.08%), whereas that in the other four samples
was >95%. Negative markers of MSCs were expressed in >2% of
cells for 3 SSSs (Fig. 4). The
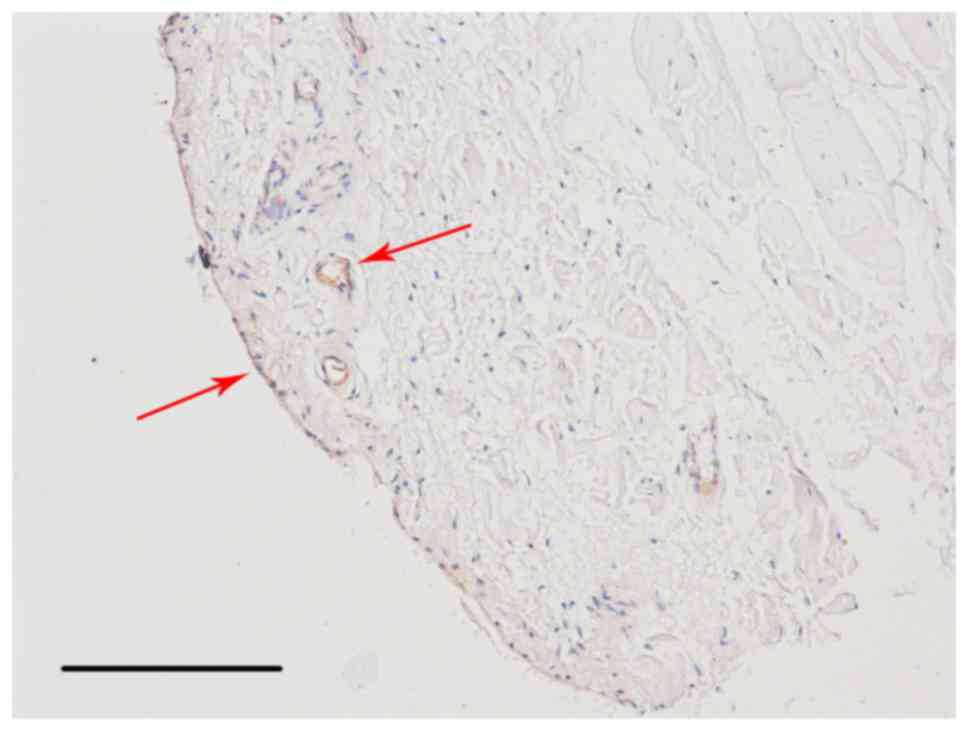
results of immunohistochemical staining demonstrated that
CD105+ cells were located in the intima and subintima,
indicating that CD105 was not a specific marker for SMSCs (Fig. 5).
Differentiation potential of SFCs and
SSSCs
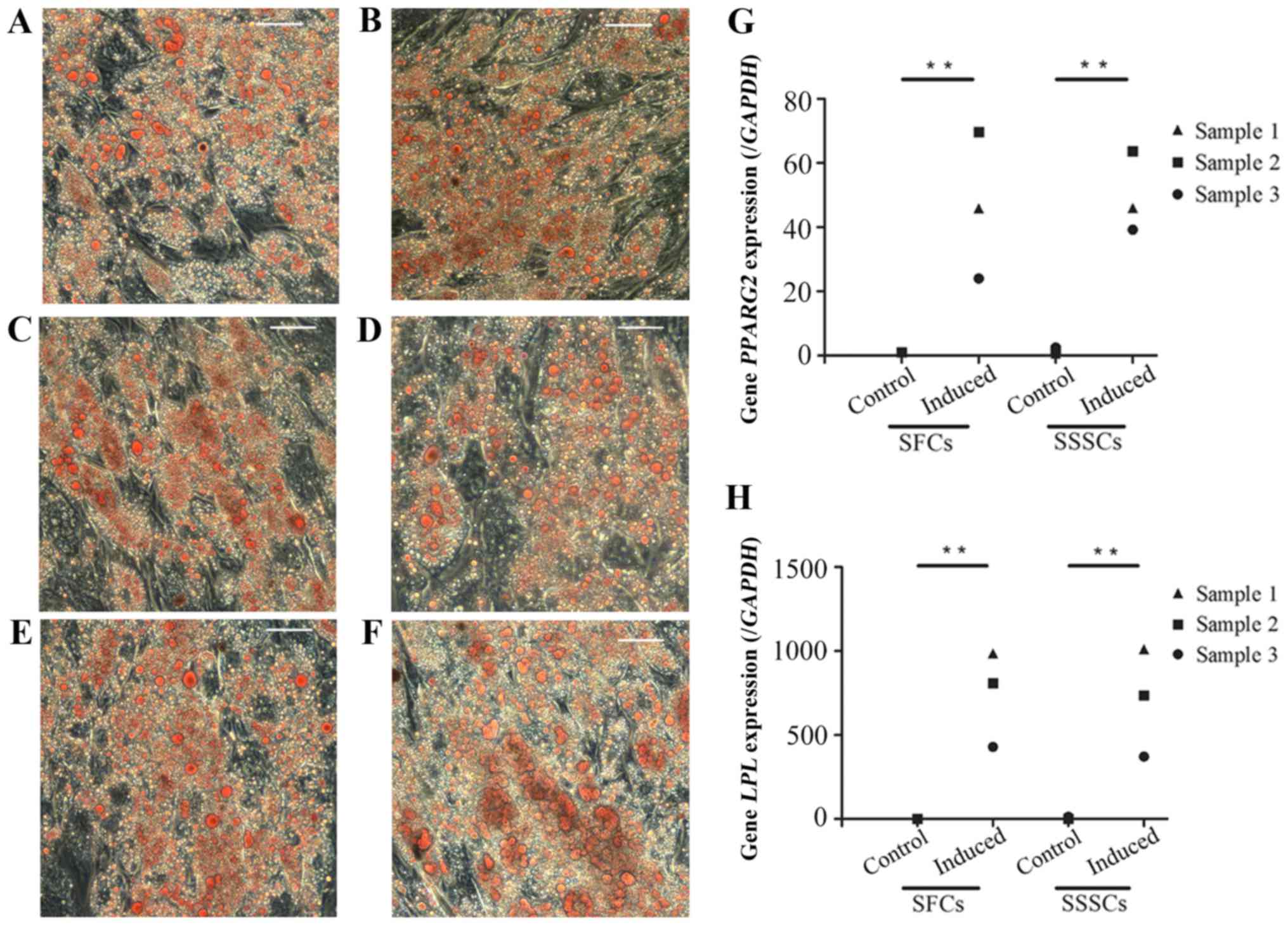
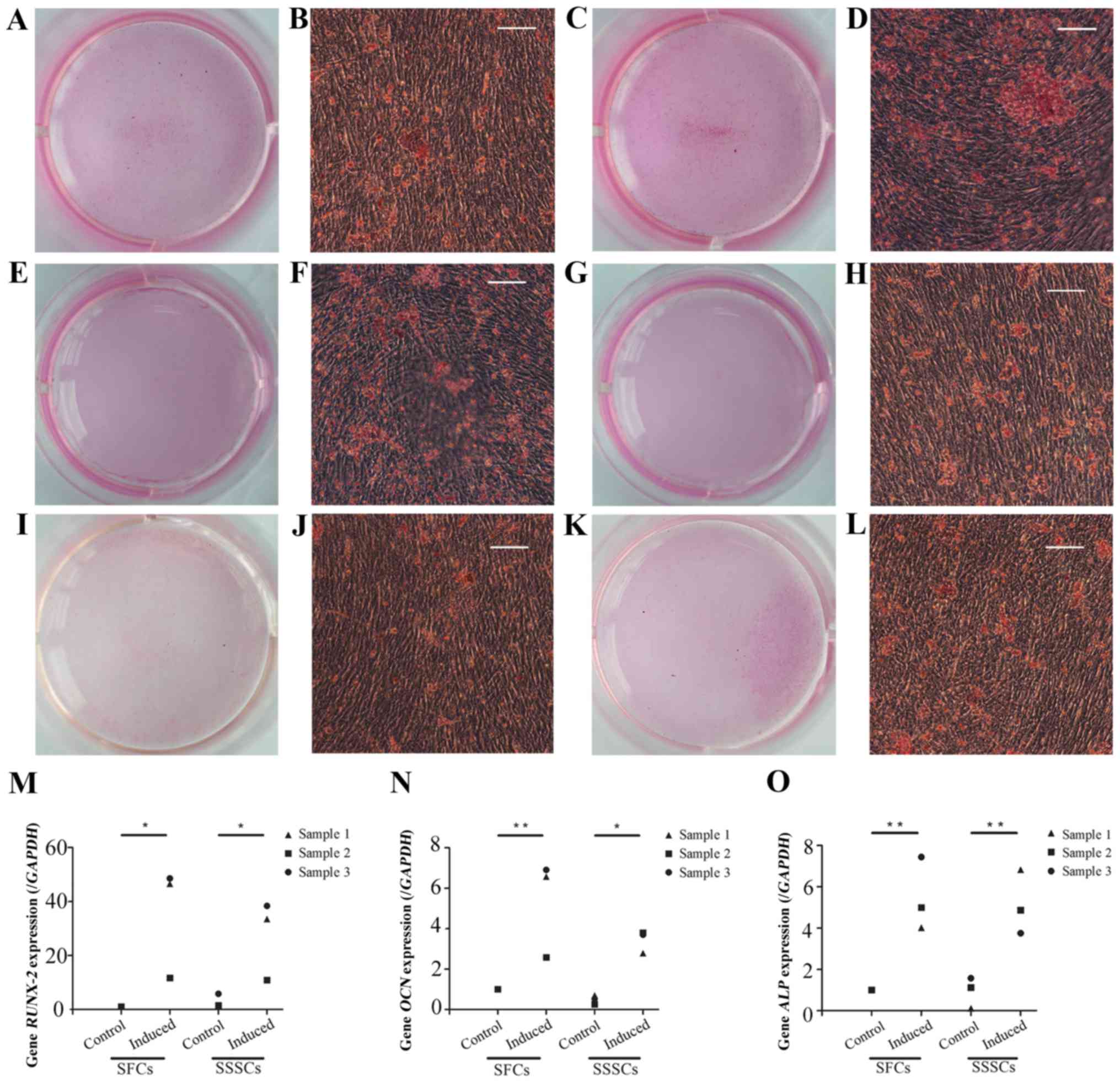
Osteogenic differentiation
Calcium deposits were detected in both groups after
a 4-week osteogenic induction, as confirmed by Alizarin Red S
staining (Fig. 6A–L). The
expression levels of osteogenesis-associated genes, including
runt-related transcription factor 2 (Fig. 6M), osteocalcin (Fig. 6N) and alkaline phosphatase
(Fig. 6O), were significantly
upregulated in induced SFCs and SSSCs compared with those in the
corresponding control groups. However, there were no differences
between the two induced groups. Results are presented in Table III.
 | Figure 6Osteogenic differentiation of SFCs
and SSSCs. Calcium deposits were confirmed by Alizarin Red S
staining in (A, B, E, F, I and J) SFCs and (C, D, G, H, K and L)
SSSCs. Scale bars, 100 μm. qPCR results for (M)
RUNX-2, (N) OCN, and (O) ALP expression in
SFCs and SSSCs compared with those in the corresponding control
groups. *P<0.05, **P<0.01. ALP,
alkaline phosphatase; OCN, osteocalcin; RUNX-2,
runt-related transcription factor 2; SFCs, synovial fragment cells;
SSSCs, surgery-obtained synovium specimen cells. |
| Table IIIExpression after multipotent
differentiation. |
Table III
Expression after multipotent
differentiation.
| Genes and
proteins | SFCs-control | SFCs-induced | SSSCs-control | SSSCs-induced | P-values
|
|---|
| SFCs-control vs.
SFCs-induced | SSSCs-control vs.
SSSCs-induced | SFCs-induced vs.
SSSCs-induced |
|---|
| RUNX-2 | 1.00±0.00 | 35.61±20.77 | 2.92±2.50 | 27.58±14.68 | 0.01 | 0.05 | 0.46 |
| OCN | 1.00±0.00 | 5.35±2.41 | 0.50±0.23 | 3.43±0.56 | 0.003 | 0.02 | 0.10 |
| ALP | 1.00±0.00 | 5.49±1.76 | 0.94±0.75 | 5.15±1.56 | 0.002 | 0.003 | 0.75 |
| PPARG2 | 1.00±0.00 | 46.56±22.84 | 1.59±0.93 | 49.65±12.59 | 0.003 | 0.002 | 0.78 |
| LPL | 1.00±0.00 | 741.14±284.52 | 5.42±7.80 | 705.91±320.99 | 0.003 | 0.004 | 0.85 |
| SOX-9 | 1.00±0.00 | 1.53±0.31 | 0.76±0.42 | 1.84±0.04 | 0.04 | 0.001 | 0.18 |
| COL-2 | 1.00±0.00 | 6.00±0.82 | 0.88±0.15 | 6.58±0.31 | <0.001 | <0.001 | 0.15 |
| GAG | 1.36±0.06 | 7.61±0.55 | 1.36±0.06 | 8.07±1.81 | <0.001 | <0.001 | 0.57 |
Adipogenic differentiation
Oil red O-positive, lipid-laden fat cells were
detected in SFCs and SSSCs after a 4-week adipogenic induction
(Fig. 7A–F). In addition, the
expression levels of peroxisome proliferator-activated receptor γ,
transcript variant 2 (Fig. 7G)
and lipoprotein lipase (Fig. 7H)
were significantly upregulated in both induced cell types compared
with in the control groups. No differences were detected between
the two induced groups. Results are presented in Table III.
Chondrogenic differentiation
Chondrogenic differentiation was assessed by
detection of cartilage nodules and collagen type II expression
(Fig. 8). Cartilage nodules were
formed by SFCs (Fig. 8A, E and I)
and SSSCs (Fig. 8C, G and K)
after a 3-week chondrogenic induction. In addition, collagen type
II expression was detected in cartilage nodule sections formed by
SFCs (Fig. 8B, F and J) and SSSCs
(Fig. 8D, H and L), as confirmed
by immunohistochemical staining. Immunofluorescence staining was
also conducted on cells without induction, which served as the
control group; however, no immunostaining was detected (data not
shown). In addition, the mRNA expression levels of sex-determining
region Y-box 9 (Fig. 8M) and type
II collagen (Fig. 8N), and the
production of GAG (Fig. 8O) were
significantly upregulated in both cell types compared with in the
control groups. No differences were detected between the two
induced groups. Results are presented in Table III.
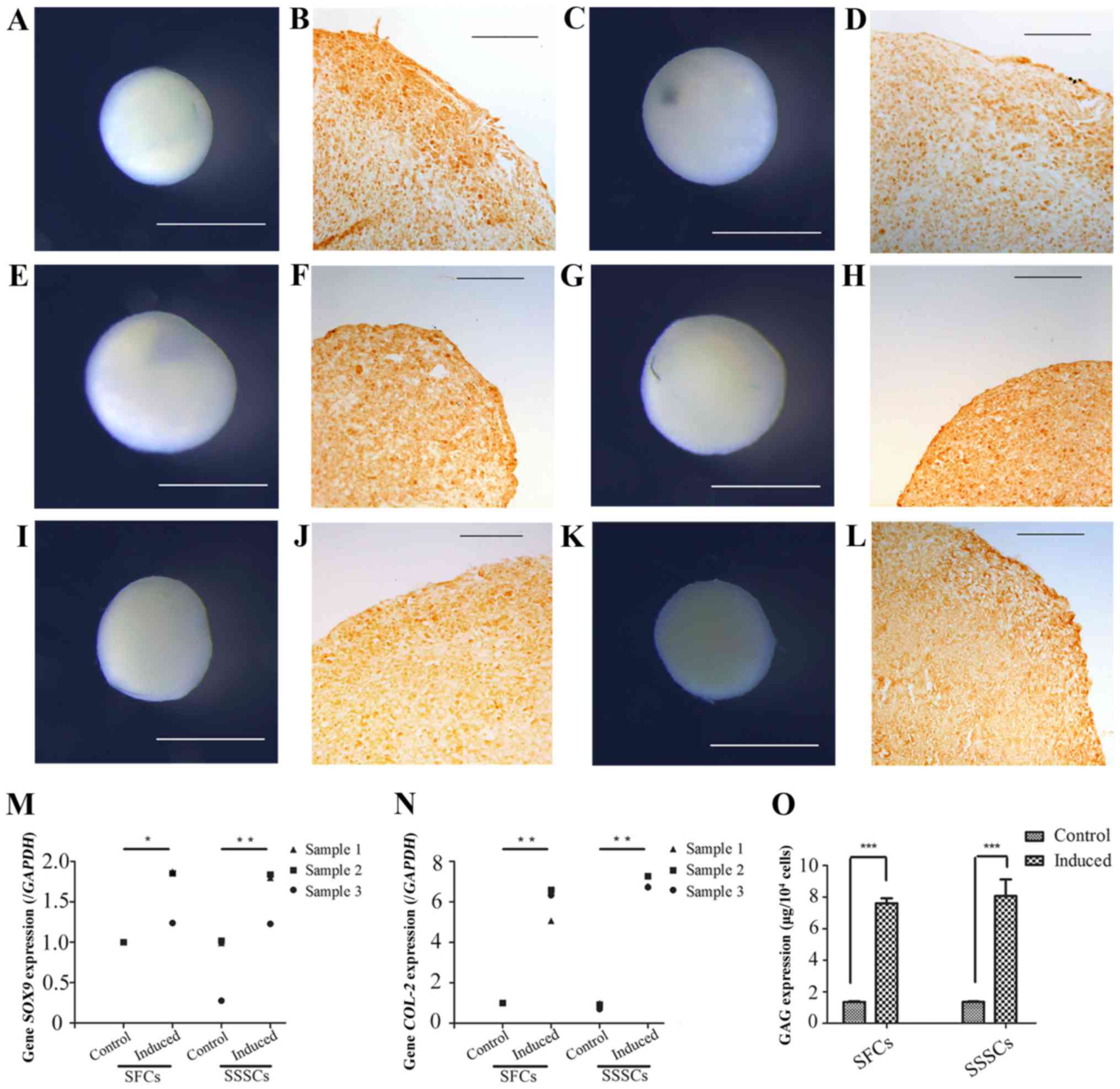 | Figure 8Chondrogenic differentiation of SFCs
and SSSCs. Gross morphology of cartilage nodules formed by (A, E
and I) SFCs and (C, G and K) SSSCs. Immunohistochemical staining of
type II collagen in the cartilage nodules formed by (B, F and J)
SFCs and (D, H and L) SSSCs. Scale bars, (B, D, F, H, J and L) 100
μm and (A, C, E, G, I and K) 1 mm. qPCR results for (M)
SOX-9 and (N) COL-2 expression in SFCs and SSSCs
compared with in the corresponding control groups. (O) GAG levels
in cartilage nodules formed by SFCs and SSSCs.
*P<0.05, **P<0.01 and
***P<0.001. COL-2, type II collagen; GAG,
glycosaminoglycan; SFCs, synovial fragment cells; SOX-9,
sex-determining region Y-box 9; SSSCs, surgery-obtained synovium
specimen cells. |
Discussion
The present study investigated the morphological
characteristics of cells derived from SFs and SSSs. The main
differences between SFCs and SSSCs are summarized in Table IV. The main similarities between
the cell types were as follows: i) Positive expression of typical
markers of MSCs, including CD44, CD73, CD90 and CD105, and negative
expression of CD34, CD45, CD79α/CD19, CD14/CD11b and HLA-DR; ii)
fibroblast-like and spindle-shaped morphology; iii) multi-lineage
differentiation potential; and iv) clone-forming potential. These
cells exhibited similar morphological characteristics, and no
significant differences in osteogenic, adipogenic or chondrogenic
differentiation potential.
| Table IVMain differences between SFCs and
SSSCs. |
Table IV
Main differences between SFCs and
SSSCs.
| Properties | SFCs | SSSCs |
|---|
| Source of separated
MSCs | Only from the
intima (synovium) | Intima (synovium)
and subintima, including MSCs from pericytes |
| Markers of MSC
expression in primary cells | Higher
uniformity | Less
uniformity |
| Ethical
controversy | Less | More |
After the initial characterization of bone marrow
stromal cells in the late 1960s, the concept of MSCs was
established, and progenitor cells with similar properties have been
isolated from various sources (24). In 2006, the International Society
for Cellular Therapy (ISCT) set criteria for the definition of
MSCs, defining MSCs as cells with the ability to adhere to plastic
in standard culture conditions and to differentiate into
osteogenic, adipogenic and chondrogenic lineages. Furthermore,
according to the ISCT criteria, >95% of cells must express CD90,
CD105 and CD73, whereas <2% of cells should express CD34, CD45,
CD79α/CD19, CD14/CD11b and HLA-DR (14). In the present study, all 3 SFC
samples met these criteria. Conversely, in 3 out of 6 SSS samples,
the percentage of SSSCs that expressed negative markers was >2%.
Furthermore, in 2 out of 6 SSS samples, the percentage of SSSCs
that expressed CD105 was <95%.
Synovium samples obtained through arthroscopic or
surgical procedures consist of two layers: The intima (synovial
membrane) and subintima. The intima consists of several cell
layers, whereas the subintima consists of various cells lying over
loose connective tissue, alongside matrix proteins, fibroblasts,
macrophages, lymphocytes, blood vessels and MSCs (25–29). MSCs are present in various areas
of the joint, including the intima and subintima (30–32). Since there is no effective method
for the separation of the intima and subintima, the SSMSCs reported
previously were not only from the synovium (intima), but also from
the subintima. Therefore, MSCs specifically from the intima (SMSCs)
remain poorly understood.
The present study demonstrated that SFs were present
in the synovial fluid obtained from patients with TMJ
osteoarthrosis. Shearing forces, natural remodelling and relative
tissue weakening from lack of nutrients may contribute to the
avulsion of the superficial lining (32,33). SFs consist of several layers of
cells only, indicating that this tissue is shed from the intima and
may serve as a better source of SMSCs. Furthermore, SFs can be
obtained by arthrocentesis, which is a common more acceptable
treatment strategy for patients with TMJ osteoarthrosis, as it is
less invasive than open surgery. Accordingly, SFs are a
patient-friendly source for SMSCs, which may be used in studies
regarding TMJ osteoarthrosis.
MSCs can be isolated from synovium samples and are
thought to be a promising cell type for cartilage repair due to
anatomical position (34,35). Compared with other MSC sources,
the synovium is the closest tissue to articular cartilage, and
SSSCs exhibit a higher chondrogenic capacity, and can be harvested
through routine arthroscopic or surgical procedures (9,18,36,37). The sources of these MSCs are
complex and include the intima, subintima and peripheral
circulation. Due to vascular recruitment, MSCs located around the
blood vessels in the subintima may have misled studies regarding
the origin of synovial membrane-derived MSCs (SMMSCs) (20,38,39). Li and Makarov generated animal
models of rheumatoid arthritis and demonstrated that arthritic
fibroblast-like synoviocytes contain a substantial (>30%)
fraction of bone marrow-derived precursors that can differentiate
into various mesenchymal cell types in vitro (40). Furthermore, the higher
chondrogenic capacity of SMMSCs remains poorly understood. The
present study indicated that the differentiation potentials of
SFCs, including chondrogenic differentiation potential, were not
significantly different from those of SSSCs, thus suggesting that
the higher chondrogenic capacity of SMMSCs was not determined by
MSCs residing in the intima.
No specific markers for SMSCs have been identified
to date. Harvanova et al reported that 40–50% of cells
isolated from synovial samples are positive for CD105 (19). CD105 serves as an immunomagnetic
separation marker for isolating SMMSC populations (19). The present study also confirmed
that CD105− cells were present in the cell population
isolated from SSSs, but not that isolated from SFs. However,
CD105+ cells were present in the intima and subintima,
indicating that CD105 was not a specific marker for SMSCs. Since
SFs consist of only intima, this tissue may allow for the
exploration of markers specific to SMSCs.
In conclusion, the present study isolated MSCs from
SFs and demonstrated that these SFCs were similar in morphology,
growth and trilineage differentiation potential to SSSCs. However,
SFCs exhibited more homogeneous characteristics than SSSCs. Not
only were SFCs more uniform than SSSCs in terms of MSC surface
marker expression in primary cells, but SFCs derived from SFs also
consisted only of intima (synovium), thus exhibiting more
homogeneity with regards to cell source. Conversely, SSSCs were
derived from SSSs, which consisted of intima and subintima. In
addition, the source of separated MSCs from SSSs exhibited
heterogeneity, since SSSCs may contain MSCs from pericytes. Unlike
other joints, including knee joints, the size of SSSs are limited;
therefore, removing excessive synovial specimens from the TMJ
raises ethical concerns. However, the process for obtaining SFs is
simpler and less invasive, thereby making it more acceptable to
patients and more in compliance with ethical and moral standards.
Therefore, SFs may serve as an improved cell source for the study
of SMSCs. Notably, obtaining MSCs from the intima is a key step in
the exploration of specific SMSC markers. In the present study,
although SFCs were confirmed as MSCs derived from the intima,
specific markers for SMSCs remain to be identified; therefore, our
future studies aim to investigate these markers.
Acknowledgments
The present study was supported by a grant from the
National Science Foundation of China (grant no. 81271115).
Abbreviations:
|
MSCs
|
mesenchymal stem cells
|
|
SSMSCs
|
synovium specimen-derived mesenchymal
stem cells
|
|
SSSs
|
surgery-obtained synovium
specimens
|
|
SSSCs
|
surgery-obtained synovium specimen
cells
|
|
SFs
|
synovial fragments
|
|
SFCs
|
synovial fragment cells
|
|
SMSCs
|
synovial mesenchymal stem cells
|
|
ISCT
|
International Society for Cellular
Therapy
|
References
|
1
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jackson WM, Nesti LJ and Tuan RS:
Potential therapeutic applications of muscle-derived mesenchymal
stem and progenitor cells. Expert Opin Biol Ther. 10:505–517. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Labusca LS, Botez P, Zugun Eloae F and
Mashayekhi K: Stem cells derived from osteoarthritic knee
mesenchymal tissues: A pilot study. Eur J Orthop Surg Traumatol.
23:169–176. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Murray IR, West CC, Hardy WR, James AW,
Park TS, Nguyen A, Tawonsawatruk T, Lazzari L, Soo C and Péault B:
Natural history of mesenchymal stem cells, from vessel walls to
culture vessels. Cell Mol Life Sci. 71:1353–1374. 2014. View Article : Google Scholar
|
|
5
|
Ogata Y, Mabuchi Y, Yoshida M, Suto EG,
Suzuki N, Muneta T, Sekiya I and Akazawa C: Purified human synovium
mesenchymal stem cells as a good resource for cartilage
regeneration. PloS One. 10:e01290962015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Bari C, Dell'Accio F, Tylzanowski P and
Luyten FP: Multipotent mesenchymal stem cells from adult human
synovial membrane. Arthritis Rheum. 44:1928–1942. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Imanishi Y, Miyagawa S, Kitagawa-Sakakida
S, Taketani S, Sekiya N and Sawa Y: Impact of synovial
membrane-derived stem cell transplantation in a rat model of
myocardial infarction. J Artif Organs. 12:187–193. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bernardo ME, Pagliara D and Locatelli F:
Mesenchymal stromal cell therapy: A revolution in Regenerative
Medicine. Bone Marrow Transplant. 47:164–171. 2012. View Article : Google Scholar
|
|
9
|
Arufe MC, De la Fuente A, Fuentes I, de
Toro FJ and Blanco FJ: Chondrogenic potential of subpopulations of
cells expressing mesenchymal stem cell markers derived from human
synovial membranes. J Cell Biochem. 111:834–845. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fan J, Varshney RR, Ren L, Cai D and Wang
DA: Synovium-derived mesenchymal stem cells: A new cell source for
musculoskeletal regeneration. Tissue Eng Part B Rev. 15:75–86.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
de Sousa EB, Casado PL, Moura Neto V,
Duarte ME and Aguiar DP: Synovial fluid and synovial membrane
mesenchymal stem cells: Latest discoveries and therapeutic
perspectives. Stem Cell Res Ther. 5:1122014. View Article : Google Scholar
|
|
12
|
Burkandt A, Katzer A, Thaler K, Von Baehr
V, Friedrich RE, Rüther W, Amling M and Zustin J: Proliferation of
the synovial lining cell layer in suggested metal hypersensitivity.
In vivo. 25:679–686. 2011.PubMed/NCBI
|
|
13
|
Chen K, Man C, Zhang B, Hu J and Zhu SS:
Effect of in vitro chondrogenic differentiation of autologous
mesenchymal stem cells on cartilage and subchondral cancellous bone
repair in osteoarthritis of temporomandibular joint. Int J Oral
Maxillofac Surg. 42:240–248. 2013. View Article : Google Scholar
|
|
14
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop Dj and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The International Society for Cellular
Therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakagawa Y, Muneta T, Kondo S, Mizuno M,
Takakuda K, Ichinose S, Tabuchi T, Koga H, Tsuji K and Sekiya I:
Synovial mesenchymal stem cells promote healing after meniscal
repair in microminipigs. Osteoarthritis Cartilage. 23:1007–1017.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Katagiri H, Muneta T, Tsuji K, Horie M,
Koga H, Ozeki N, Kobayashi E and Sekiya I: Transplantation of
aggregates of synovial mesenchymal stem cells regenerates meniscus
more effectively in a rat massive meniscal defect. Biochem Biophys
Res Commun. 435:603–609. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hatsushika D, Muneta T, Nakamura T, Horie
M, Koga H, Nakagawa Y, Tsuji K, Hishikawa S, Kobayashi E and Sekiya
I: Repetitive allogeneic intraarticular injections of synovial
mesenchymal stem cells promote meniscus regeneration in a porcine
massive meniscus defect model. Osteoarthritis Cartilage.
22:941–950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sakaguchi Y, Sekiya I, Yagishita K and
Muneta T: Comparison of human stem cells derived from various
mesenchymal tissues: Superiority of synovium as a cell source.
Arthritis Rheum. 52:2521–2529. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harvanova D, Tothova T, Sarissky M,
Amrichova J and Rosocha J: Isolation and characterization of
synovial mesenchymal stem cells. Folia Biol. 57:119–124. 2011.
|
|
20
|
Nagase T, Muneta T, Ju YJ, Hara K, Morito
T, Koga H, Nimura A, Mochizuki T and Sekiya I: Analysis of the
chondrogenic potential of human synovial stem cells according to
harvest site and culture parameters in knees with medial
compartment osteoarthritis. Arthritis Rheum. 58:1389–1398. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Cai H, Fang W, Meng Q, Wu Y, Li J,
Deng M and Long X: Triple-layered cell sheet for tissue-engineering
the synovial membrane of the temporomandibular joint. Cells Tissues
Organs. 199:150–158. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vandenabeele F, De Bari C, Moreels M,
Lambrichts I, Dell'Accio F, Lippens PL and Luyten FP: Morphological
and immunocytochemical characterization of cultured fibroblast-like
cells derived from adult human synovial membrane. Arch Histol
Cytol. 66:145–153. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Friedenstein AJ, Piatetzky S II and
Petrakova KV: Osteogenesis in transplants of bone marrow cells. J
Embryol Exp Morph. 16:381–390. 1966.PubMed/NCBI
|
|
25
|
Smith MD: The normal synovium. Open
Rheumatol J. 5:100–106. 2011. View Article : Google Scholar
|
|
26
|
O'Connell JX: Pathology of the synovium.
Am J Clin Pathol. 114:773–784. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iwanaga T, Shikichi M, Kitamura H, Yanase
H and Nozawa-Inoue K: Morphology and functional roles of
synoviocytes in the joint. Arch Histol Cytol. 63:17–31. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kung M, Markantonis J, Nelson S and
Campbell P: The synovial lining and synovial fluid properties after
joint arthroplasty. Lubricants. 3:394–412. 2015. View Article : Google Scholar
|
|
29
|
Carvalho de Moraes LO, Tedesco RC,
Arraez-Aybar LA, Klein O, Merida-Velasco JR and Alonso LG:
Development of synovial membrane in the temporomandibular joint of
the human fetus. Eur J Histochemistry. 59:25692015. View Article : Google Scholar
|
|
30
|
El-Jawhari JJ, El-Sherbiny YM, Jones EA
and McGonagle D: Mesenchymal stem cells, autoimmunity and
rheumatoid arthritis. QJM. 107:505–514. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Khan IM, Bishop JC, Gilbert S and Archer
CW: Clonal chondroprogenitors maintain telomerase activity and Sox9
expression during extended monolayer culture and retain
chondrogenic potential. Osteoarthritis Cartilage. 17:518–528. 2009.
View Article : Google Scholar
|
|
32
|
Moskalewski S, Osiecka-Iwan A,
Jankowska-Steifer E and Hyc A: Synovial membrane asks for
independence. Folia Morphol. 73:395–398. 2014. View Article : Google Scholar
|
|
33
|
Dai L, Pessler F, Chen LX, Clayburne G and
Schumacher HR: Detection and initial characterization of synovial
lining fragments in synovial fluid. Rheumatology. 45:533–537. 2006.
View Article : Google Scholar
|
|
34
|
Fickert S, Fiedler J and Brenner RE:
Identification, quantification and isolation of mesenchymal
progenitor cells from osteoarthritic synovium by fluorescence
automated cell sorting. Osteoarthritis Cartilage. 1:790–800. 2003.
View Article : Google Scholar
|
|
35
|
Gullo F and De Bari C: Prospective
purification of a subpopulation of human synovial mesenchymal stem
cells with enhanced chondro-osteogenic potency. Rheumatology.
52:1758–1768. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mochizuki T, Muneta T, Sakaguchi Y, Nimura
A, Yokoyama A, Koga H and Sekiya I: Higher chondrogenic potential
of fibrous synovium-and adipose synovium-derived cells compared
with subcutaneous fat-derived cells: Distinguishing properties of
mesenchymal stem cells in humans. Arthritis Rheumatism. 54:843–853.
2006. View Article : Google Scholar
|
|
37
|
Dry H, Jorgenson K, Ando W, Hart DA, Frank
CB and Sen A: Effect of calcium on the proliferation kinetics of
synovium-derived mesenchymal stromal cells. Cytotherapy.
15:805–819. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
da Silva Meirelles L, Sand TT, Harman RJ,
Lennon DP and Caplan AI: MSC frequency correlates with blood vessel
density in equine adipose tissue. Tissue Eng Part A. 15:221–229.
2009. View Article : Google Scholar
|
|
39
|
Watt SM, Gullo F, van der Garde M,
Markeson D, Camicia R, Khoo CP and Zwaginga JJ: The angiogenic
properties of mesenchymal stem/stromal cells and their therapeutic
potential. Br Med Bull. 108:25–53. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li X and Makarov SS: An essential role of
NF-κB in the 'tumor-like' phenotype of arthritic synoviocytes. Proc
Nat Acad Sci USA. 103:17432–17437. 2006. View Article : Google Scholar
|