Introduction
The epithelial-to-mesenchymal transition (EMT) is a
complicated and critical process in the metastatic spread of cancer
cells. The transition of morphology of a cancer cell from an
epithelial to a mesenchymal morphology leads to increased migratory
and invasive cellular properties. Previous studies have
demonstrated that EMT in cancer cells involves the upregulation of
mesenchymal markers and the downregulation of epithelial cell
markers (1,2). For example, fibroblast specific
protein-1 (S100A4) is a prototypical fibroblast marker of EMT in
cancer; the expression of S100A4 is associated with cellular
motility and the regulation of cell polarization through its
effects on the localization of protrusions, which involves
interactions with myosin-IIA (1,3).
The α-smooth muscle actin (αSMA) protein is expressed by vascular
smooth muscle and myoepithelial cells, and is also considered a
marker of EMT. There is also evidence that type 3 EMT is associated
with αSMA in basal-like breast cancer (2,4).
Therefore, the function of these molecules expressed by stromal
fibroblasts and epithelium-derived cancer cells may be importance
in EMT.
Fibroblast activation protein-α (FAPα) is an
integral membrane serine peptidase. Previous studies have shown
that FAPα is expressed primarily in fetal mesenchymal tissues,
stromal fibroblasts, wounded tissues and stromal fibroblasts of
malignant epithelial tumors (5–7).
FAPα expressed by stromal cancer-associated fibroblasts has
dipeptidyl peptidase activity (8,9)
and collagenolytic activity (10,11). By degrading the extracellular
matrix, FAPα promotes the growth and metastasis of cancer cells
(12–14). The overexpression of FAPα is also
associated with distant metastasis, tumor recurrence and poor
survival rates (15,16). In addition, previous studies have
shown that FAPα is expressed in certain types of epithelium-derived
cancer, including breast (17),
gastric (18), esophageal
(19), ovarian (20) and colorectal cancer (21).
The functions of cancer cell-expressed FAPα have
been investigated in a number of studies and the results from these
suggest that FAPα has a non-enzymatic function. Wang et al
analyzed the effect of the overexpression of FAPα on the LX-2 human
hepatic stellate cell line (22);
it was found that the overexpression of FAPα increased the
adhesion, migration and invasion of LX-2 cells, and that the
proteolytic activity of FAPα was not necessary for these functions
(22). Huang et al used
two inhibitors, PT-630 and LAF-237, to inhibit the dipeptidyl
peptidase activity of FAPα (23),
and found that the inhibitors were unable to slow the growth of
tumors in severe combined immunodeficient (SCID) mice implanted
with FAPα-expressing breast cancer WTY-1/6 cells (MDA MB-231 cells
transfected with FAPα) and MDA-MB-435 cells (endogenously express
FAPα). In addition, breast cancer cells expressing a catalytically
inactive mutant of FAPα produced tumors, which grew rapidly
(23). Wang et al found
that the knockdown of FAPα in oral squamous cancer cells suppressed
cell proliferation in vitro and inhibited the growth of
tumor xenografts in mice in vivo. Notably, suppressing FAPα
in oral squamous cancer cells significantly decreased the
expression of phosphorylated phosphatidylinositol-3-kinase (PI3K),
protein kinase B (AKT), mitogen-activated protein kinase kinase 1/2
and extracellular signal-regulated kinase (ERK)1/2, and upregulated
the expression of phosphatase and tensin homolog (PTEN) (24). In our previous study (25), the overexpression of wild-type
FAPα and the enzymatic mutant FAPS624A, in which the serine
catalytic triad was disrupted, markedly increased cellular growth
and motility in MCF-7 breast cancer cells. These observations were
consistent with those for FAPα-silenced BT549 breast cancer cells.
Western blot analysis also revealed that the overexpression of
wild-type FAPα and the enzymatic mutant FAPS624A resulted in the
activation of PI3K/AKT and matrix metalloproteinase 2/9 (MMP2/9)
(25). These results suggested
that FAPα may serve as an oncogene and be involved in the
regulation of cell signaling pathways.
Therefore, the present study analyzed the function
of FAPα in lung cancer cells in order to examine its non-enzymatic
function. It was hypothesized that, as a membrane protein, FAPα may
be involved in the regulation of certain signaling pathways and,
through this mechanism, exert its effect on lung cancer cells.
Materials and methods
Materials and cell lines
The SK-MES-1 squamous cell carcinoma (SCC) and A549
adenocarcinoma (AC) human lung cancer cell lines were obtained from
the American Type Culture Collection (Manassas, VA, USA). Primary
antibodies against human FAPα (cat. no. AF3715) were from R&D
Systems, Inc. (Minneapolis, MN, USA). Primary antibodies against
glyceraldehyde 3-phosphate dehydrogenase (GAPDH; cat. no. sc-32233)
and an inhibitor (Y-27632; cat. no. sc-3536) of Rho-associated
protein kinase (ROCK) were from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). The focal adhesion kinase (FAK) inhibitor
(cat. no. 3414) was from Tocris Bioscience (Bristol, UK). The
ERK1/2 inhibitor (FR180204, cat. no. 328007), phospholipase C-γ
(PLCγ) inhibitor (U73122, cat. no. 662035), neural Wiskott-Aldrich
syndrome protein (NWASP) inhibitor (wiskostatin, cat. no. 681525),
PI3K inhibitor (wortmannin, cat. no. 681675), c-Jun N-terminal
kinase (JNK) inhibitor (SP600125, cat. no. 420119), sonic hedgehog
(SHH) inhibitor (cyclopamine, cat. no. 239803) and a negative
control of cyclopamine (tomatidine, cat. no. 614350) were from
Calbiochem; Merck Millipore (Darmstadt, Germany). Matrigel
(reconstituted basement membrane) was purchased from Collaborative
Biomedical Products, Inc. (Bedford, MA, USA). Transwell plates
equipped with a porous insert (8-μm pore size) were from BD
Biosciences (Franklin Lakes, NJ, USA). Reverse
transcription-polymerase chain reaction (RT-PCR) reagents and
plasmid extraction kits were from Sigma-Aldrich, Inc. (St. Louis,
MO, USA). Antibodies against human PI3K (cat. no. ab22653), AKT
(cat. no. ab8805), SHH (cat. no. ab53281), Patched (Ptch; cat. no.
ab53715), and GLI family zinc finger 1 (Gli1; cat. no. ab151796)
were from Abcam PLC (Cambridge, UK).
Construction of FAPα-expressing lung
cancer cell lines
The cDNA of FAPα (NM_004460) was inserted into a
pEF6/V5-His vector (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and maintained at −80°C in the Laboratory of
Carcinogenesis and Translational Research, Peking University Cancer
Hospital and Institute, as described in our previous study
(25). The SK-MES-1 and A549 lung
cancer cells were cultured in DMEM/F12 media at 37°C with 5%
CO2. The cells were transfected with the pEF6/V5-FAPα
plasmid by electroporation. Following the selection of transfected
cells with blasticidin S (5 μg/ml) and verification via
RT-PCR analysis (RT-reaction: annealing at 65°C for 5 min,
extension at 55°C for 20 min, inactivation at 75°C for 15 min. PCR
reaction: initial denaturation at 94°C for 5 min; then denaturation
at 94°C for 30 sec, annealing at 55°C for 30 sec, extension at 72°C
for 1 min in each cycle for totally 25 cycles, followed by a final
extension of 10 min at 72°C), the following stably transfected
cells were established: FAPα-overexpressing cells (SK-MES-1exp and
A549exp), plasmid vector-transfected control cells (SK-MES-1pef and
A549pef), and wild-type cells (SK-MES-1wt and A549wt). The
transfected cells were continually cultured in a maintenance
medium, which contained 0.5 μg/ml blasticidin S.
In vitro growth, adhesion and invasion
assays
For the growth assay, the cells were plated into a
96-well plate at a density of 2,000 cells/well (n=12) followed by a
period of incubation. The cells were either fixed in 10%
formaldehyde on the day of plating or 3 days later. Crystal violet
(0.5% w/v) was used to stain the cells. Subsequently, the crystal
violet was dissolved with 10% (v/v) acetic acid and the absorbance
of cells, which represented the cell number, was determined at a
wavelength of 540 nm.
For the adhesion assay, a 96-well plate was
pre-coated with 5 μg of Matrigel and left to dry overnight.
Following rehydration with serum-free media, 1,500 cells were
seeded into each well (n=6). Following incubation for 60 min, the
non-adherent cells were washed off using BSS buffer. The remaining
cells were fixed with 4% formalin and stained with 0.5% crystal
violet. The numbers of adherent cells were then counted under an
Olympus CX31 microscope.
For the invasion assay, Transwell inserts with a
8-μm pore size were coated with 50 μg of Matrigel and
air-dried. Following rehydration with serum-free media, the cells
were seeded at a density of 30,000 cells/insert (n=5). After 3 days
of incubation at 37°C with 5% CO2, the cells, which had
migrated through the matrix and adhered to the other side of the
insert were fixed in 4% formalin, stained with 0.5% (w/v) crystal
violet, and counted under a microscope.
Electric cell-substrate impedance sensing
(ECIS)-based cell attachment and migration assay
An ECIS instrument (Applied BioPhysics, Inc., Troy,
NY, USA) was used to record the adhesive and migratory abilities of
the cells, which were determined from the changes in the impedance
of cells. 96W1E arrays were incubated with complete medium for 1 h,
following which 50,000 lung cancer cells were seeded into each well
(n=16). Electrical changes were continuously monitored for up to 24
h, with electrical wounding performed at 6 h. Multiple frequencies
(1,000, 2,000, 4,000 and 8,000 Hz) were used to assess the nature
of changes in resistance.
Inhibition of signaling pathways with
inhibitors
In order to examine the potential crosstalk between
FAPα and adhesion and migration-associated signaling pathways,
inhibitors of FAK, ERK1/2, ROCK, PLCγ, JNK, NWASP, PI3K and SHH
were used in an ECIS-based cell function assay. A total of 50,000
cells were suspended in 200 μl DMEM with inhibitors of FAK,
ERK1/2, ROCK, PLCγ, NWASP, JNK, PI3K and SHH, respectively, to a
final concentration of 100 nM. Changes in electrical resistance
under multiple frequencies (1,000, 2,000, 4,000 and 8,000 Hz) were
continuously monitored for up to 24 h, with electrical wounding
performed at 6 h in the ECIS-based wounding assay (n=8).
Western blot analysis
Western blot analysis was performed to detect the
expression of FAPα and other downstream signaling pathway molecules
in the transfected SK-MES-1 lung cancer cells and control cells.
The confluent cells were pelleted and then lysed in lysis buffer
containing 2.4 mg/ml Tris, 4.4 mg/ml NaCl, 5 mg/ml sodium
deoxycholate, 20 μg/ml sodium azide, 1.5% Triton X-100, 100
μg/ml phenylmethylsulfonyl fluoride, 1 μg/ml
leupeptin and 1 μg/ml aprotinin, for 45 min at 4°C.
Following lysis and centrifugation at 10,000 × g for 15 min at 4°C,
the isolated proteins were diluted in sample buffer and the
concentration of each sample was measured using an improved Lowry
assay (DC protein assay kit; Bio-Rad Laboratories, Inc., Hercules,
CA, USA). The samples were adjusted to equal concentrations (2
mg/ml) with sample buffer and then boiled at 100°C for 5 min prior
to separation on a 10% polyacrylamide gel. Following
electrophoresis, the separated protein samples were transferred
onto polyvinylidenedifluoride membranes (EMD Millipore, Billerica,
MA, USA) and incubated at 4°C overnight with primary antibodies
against FAPα, PI3K, AKT, SHH, Ptch, Gli1 and GAPDH at a dilution of
1:200. The membranes were then incubated with specific
peroxidase-conjugated secondary antibodies (cat. nos. sc-2357 or
sc-2031; Santa Cruz Biotechnology, Inc.) at a dilution of 1:1,000
for 40 min at room temperature. The protein signals were detected
using an enhanced chemiluminescence system (Pierce; Thermo Fisher
Scientific, Inc.).
Statistical analysis
All results are expressed as the mean ± standard
error of the mean. One-way analysis of variance and an independent
samples t-test were used to evaluate the differences between groups
in all the assays performed, with the resultant P-values
representing two-sided tests of statistical significance. All
statistical analyses were performed using SPSS 17.0 software (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
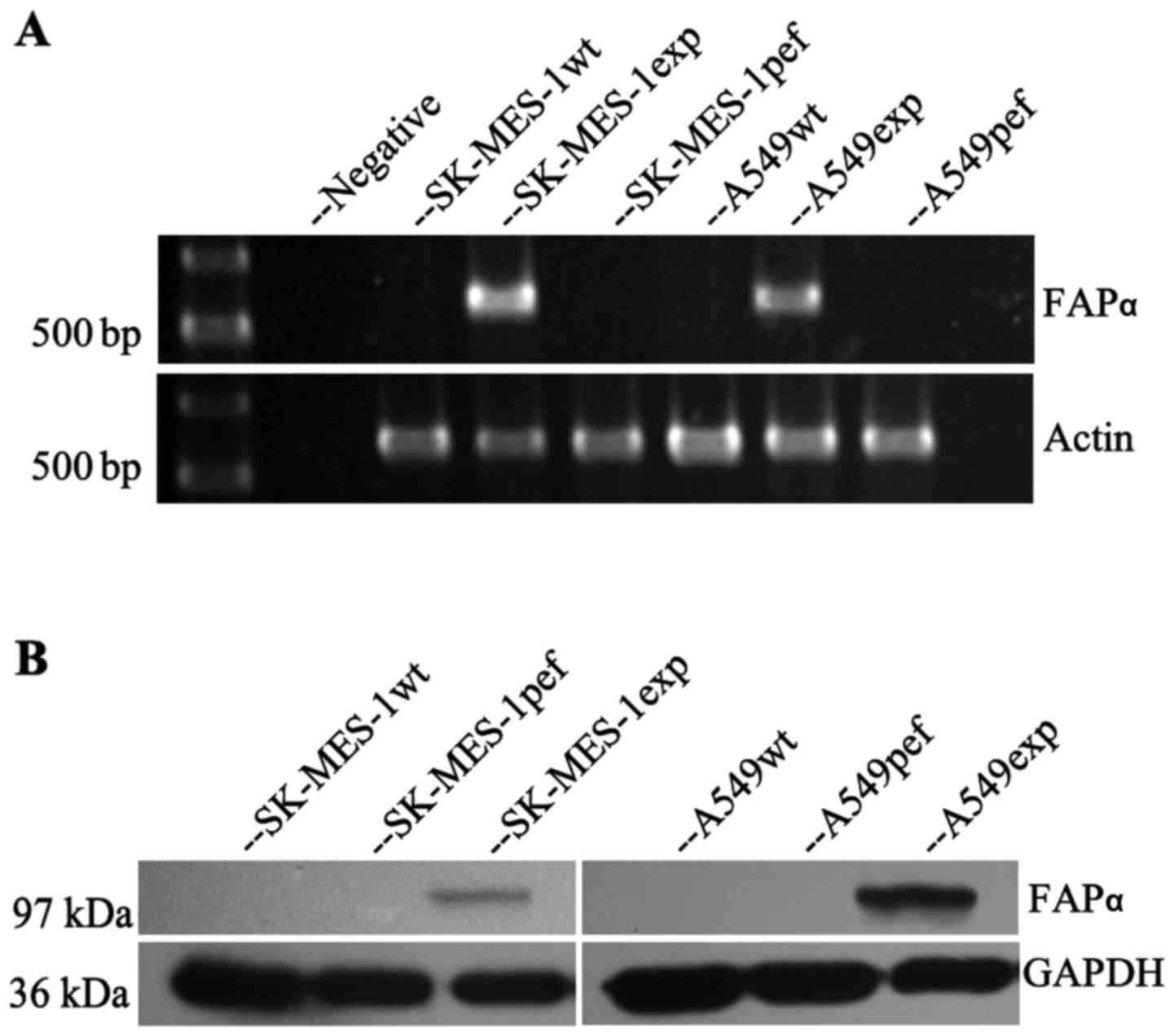
Expression of FAPα in lung cancer
cells
To identify the oncogenic function of FAPα in lung
cancer cells, FAPα was transfected into the FAPα-null SK-MES-1 and
A549 lung cancer cell lines. RT-PCR and western blot analyses were
used to confirm the expression of FAPα in the transfected cells.
The overexpression of FAPα in the SK-MES-1 and A549 cells was
successfully established (Fig.
1).
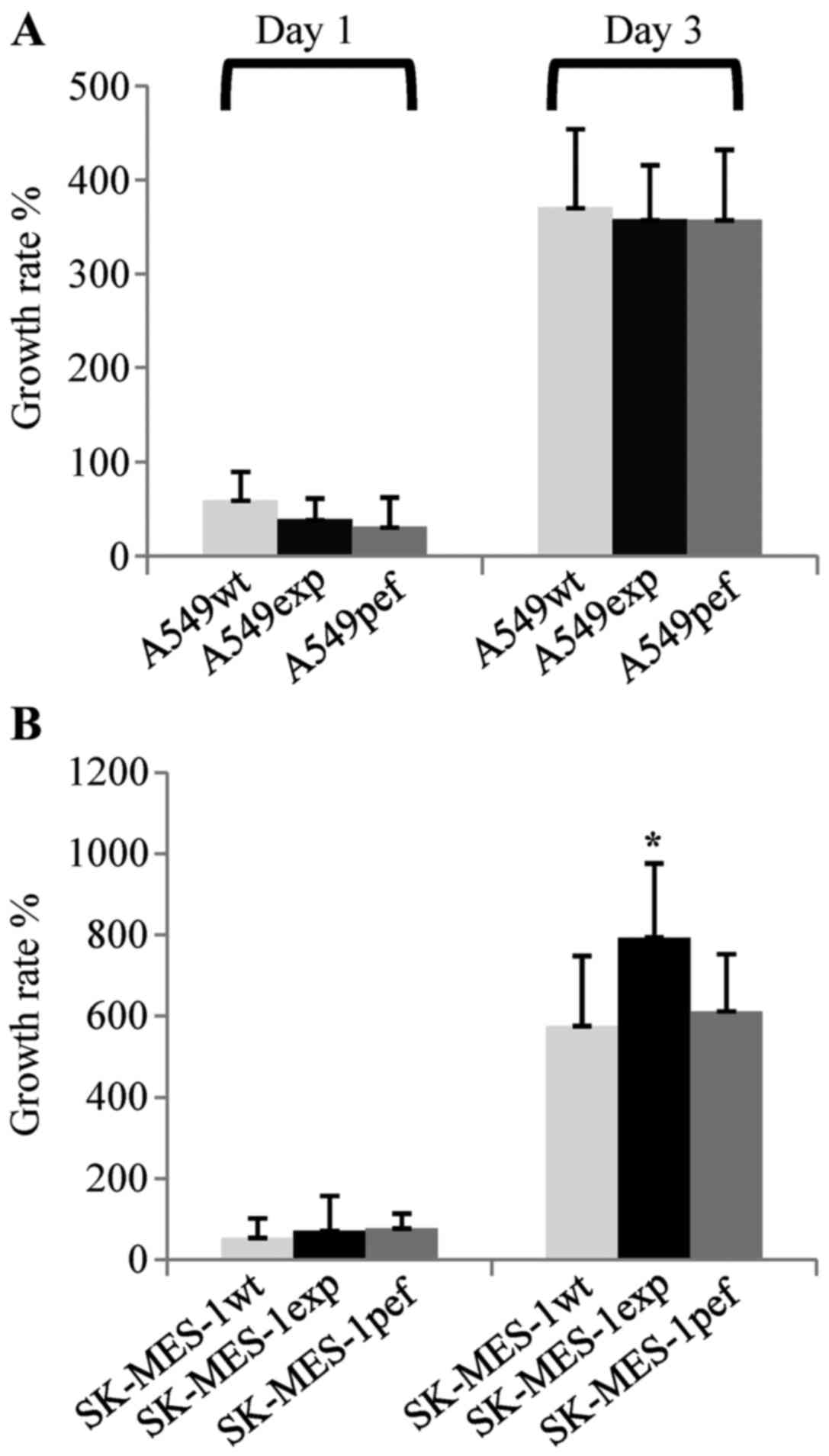
Overexpression of FAPα promotes the
growth of SK-MES-1 cells
The present study first examined the effect of the
overexpression of FAPα on cellular growth in vitro. Compared
with the SK-MES-1wt cells, the overexpression of FAPα significantly
increased the growth rate of the SK-MES-1exp cells at 3 days
post-seeding (n=12; 575.5±171.9 vs. 793.6±140.1%, respectively;
P=0.003), which was also significantly higher, compared with that
of the SK-MES-1pef cells (611.6±181.8; P=0.012). By contrast, the
growth rate of the A549exp cells (357.5±74.2%) was similar to the
growth rates of the A549wt cells (370.6±83.2; n=12; P=0.686) and
A549pef cells (357.8±57.4%, P=0.989) at 3 days post-seeding
(Fig. 2).
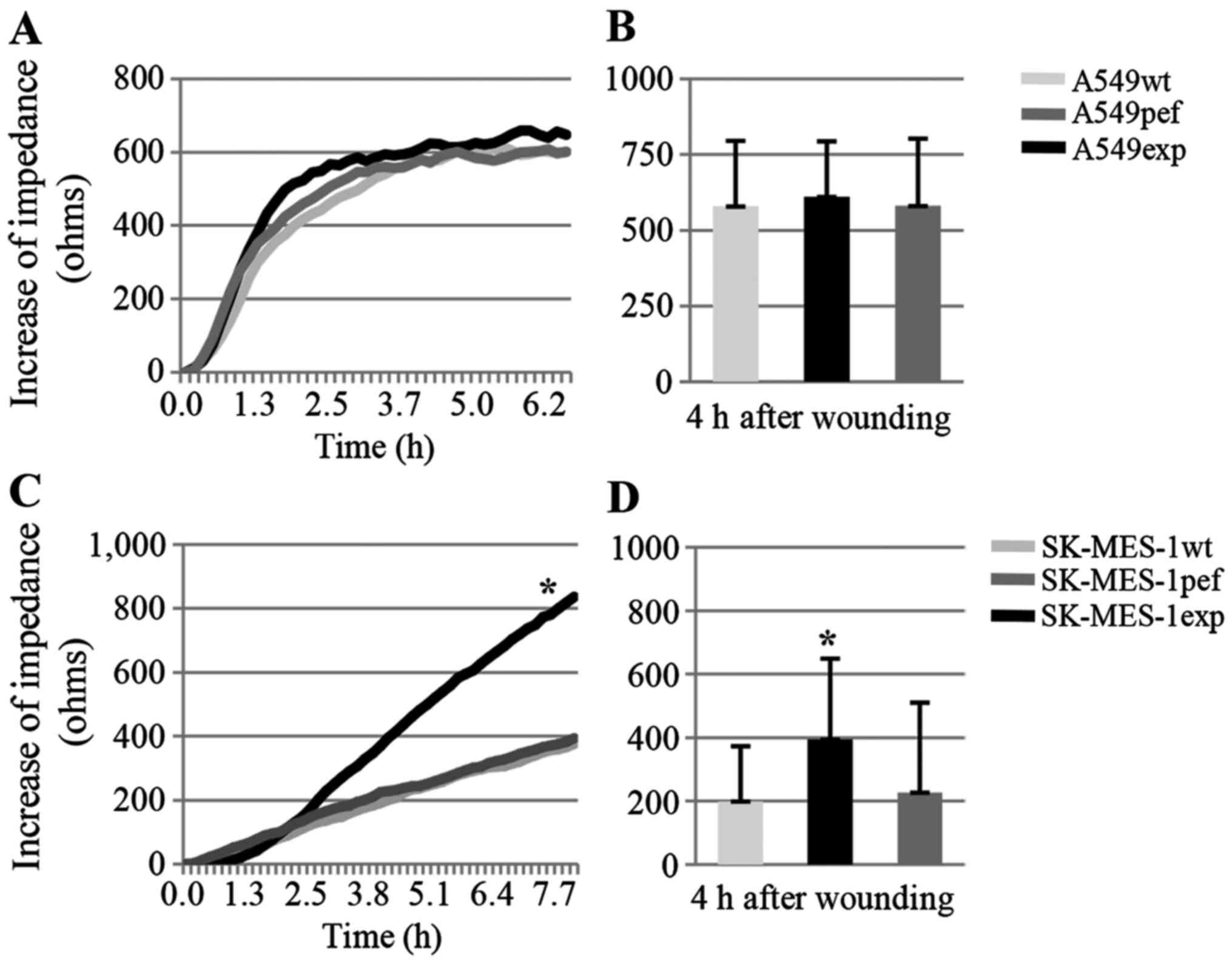
Overexpression of FAPα increases the
attachment of SK-MES-1 cells to basement membrane proteins
In a matrix gel-based adhesion assay, compared with
the control cells, FAPα-expressing SK-MES-1exp and A549exp cells
showed increased adhesion to the matrix gel 1 h following seeding;
however, no statistically significant differences were found
between the groups of SK-MES cells (n=6; SK-MES-1wt vs.
SK-MES-1exp, 25±14.4 vs. 37.3±19.9; P=0.246; SK-MES-1pef vs.
SK-MES-1exp, 31.7±10.3 vs. 37.3±19.9; P=0.549) or A549 cells
(A549wt vs. A549exp, 17.7±15.7 vs. 24.5±12.3; P=0.422; A549pef vs.
A549exp, 14.5±14.4 vs. 24.5±12.3, P=0.225) (Fig. 3A and B). To confirm the results of
the matrix gel-based adhesion assay, an ECIS-based cell function
assay was performed as a more sensitive and accurate assessment. In
the first phase post-cell seeding, increased impedance reflects the
ability of cells to adhere to the substrate; the faster the
impedance increases, the higher the rate of cell attachment to the
substrate. In the present study, compared with the SK-MES-1wt and
SK-MES-1pef control cells, the FAPα-expressing SK-MES-1exp cells
showed increased attachment at 1 h post-seeding (n=12; SK-MES-1wt
vs. SK-MES-1exp, 343.1±135.3 vs. 948.3±442.7 ohms; P=0.001;
SK-MES-1pef vs. SK-MES-1exp, 566.3±321.0 vs. 948.3±442.7 ohms;
P=0.037). However, in the A549 cells, the vector-transfected
A549pef cells showed the highest attachment ability, whereas the
A549exp cells and A549wt cells exhibited similar attachment
abilities (n=12; A549wt vs. A549exp, 234.1±149.4 vs. 292.2±191.0
ohms; P=0.416; A549pef vs. A549exp, 497.7±184.5 vs. 292.2±191.0
ohms; P=0.014) (Fig. 3C–E).
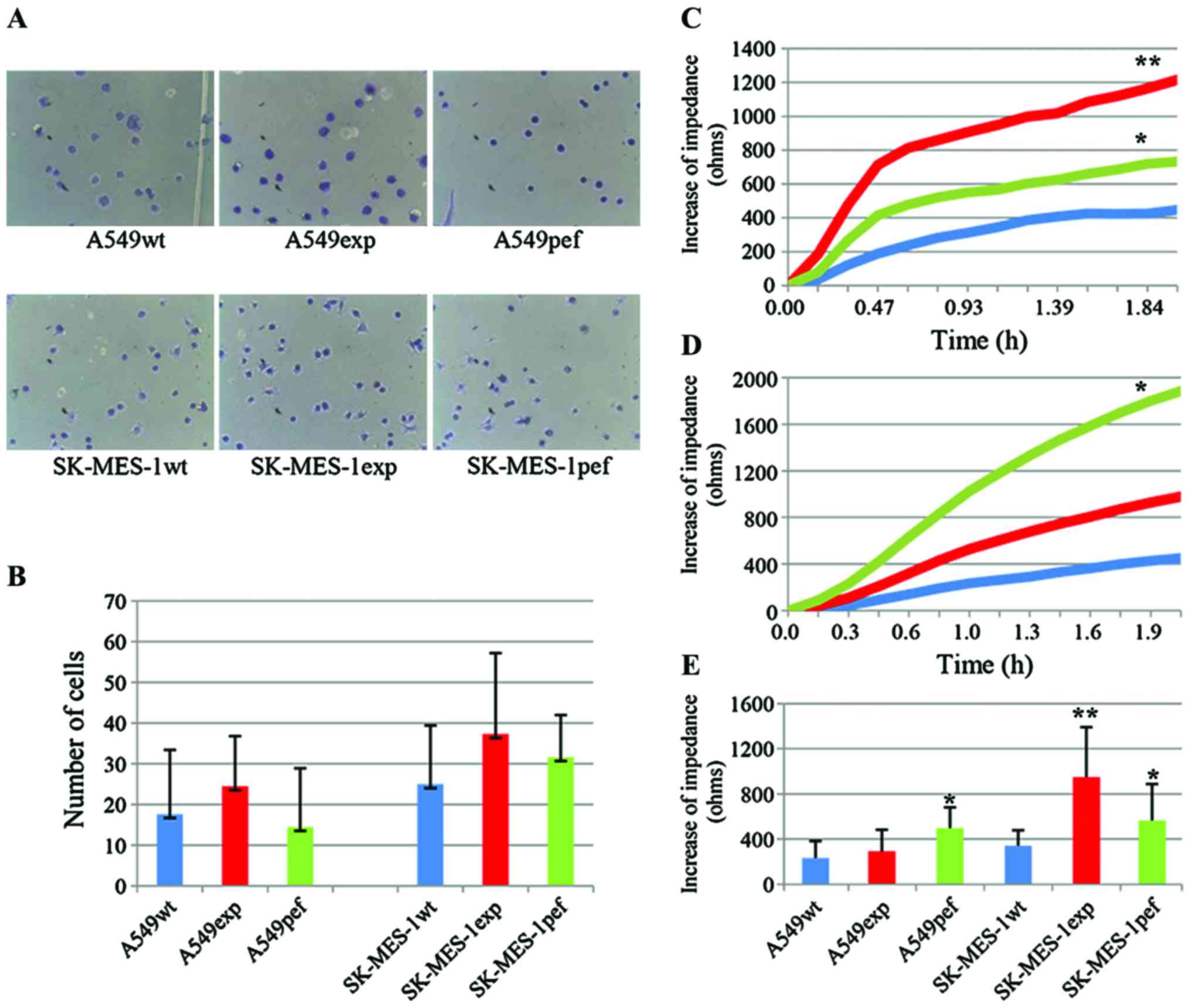 | Figure 3Overexpression of FAPα increases the
attachment of SK-MES-1 cells to the basement substrate. (A and B)
Results of the cell matrix adhesion assay showed that the
overexpression of FAPα enhanced the adhesive properties of SK-MES-1
and A549 cells, but not significantly (magnification, ×200). An
ECIS-based attachment assay confirmed the increased attachment
ability only in (C) SK-MES-1 cells, but not in (D) A549 cells over
time (n=12). (E) The increased impedance of lung cancer cells in
attachment phase of ECIS assay at 1 h after seeding.
*P<0.05, SK-MES-1pef vs. SK-MES-1exp and A549pef vs.
A549exp; **P<0.01, SK-MES-1exp vs. SK-MES-1wt. FAPα,
fibroblast activation protein α; exp, FAPα-expressing cells; pef,
vector-transfected control cells; wt, wild-type cells; ohms, unit
of electrical impedance. |
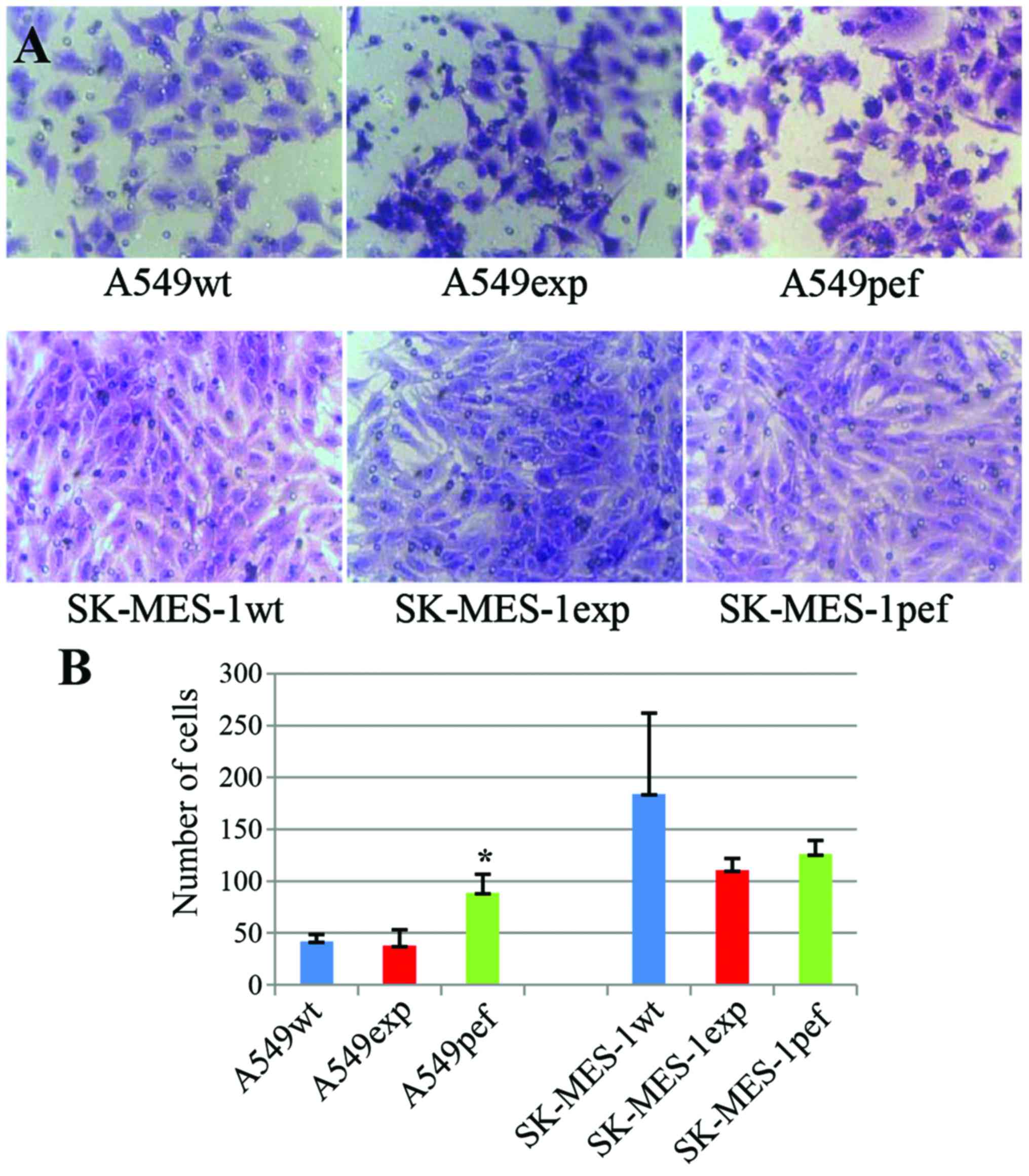
Overexpression of FAPα does not promote
the invasion of lung cancer cells
To analyze the effect of the overexpression of FAPα
on the invasion of lung cancer cells, the present study performed
an in vitro matrix gel-based invasion assay. Although FAPα
has dipeptidyl peptidase and collagenolytic activities, the results
showed that the overexpression of FAPα did not increase the
invasive ability of either SK-MES-1 or A549 cells. By contrast, the
number of invaded cells in the FAPα-expressing SK-MES-1 cell group
on day 3 was lower, compared with that in the wild-type and
vector-transfected control cell groups; however, no significant
differences were observed between the groups (n=5; SK-MES-1wt vs.
SK-MES-1exp, 184.2±77.8 vs. 110.4±11.4; P=0.138; SK-MES-1pef vs.
SK-MES-1exp, 126.0±13.2 vs. 110.4±11.4; P=0.081). In the A549
cells, the number of invaded cells in the FAPα-expressing A549exp
cell group on day 3 was similar to that in the A549wt cell group
(n=5; 37.8±15.4 vs. 42.0±6.5, respectively; P=0.59), but less than
that in the A549pef group (88.8±17.9 vs. 37.8±15.4; P=0.001)
(Fig. 4).
Overexpression of FAPα increases the
migration of SK-MES-1 cells
To investigate the effect of FAPα on the migration
of lung cancer cells, the more accurate ECIS-based wounding assay
was used rather than a physical scratch-wound assay. In the ECIS
method, the wound is created in the confluent cell monolayer using
a high voltage shock, and the faster the increase in impedance
following wounding, the higher the rate of cellular migration into
the wound. As an additional measure of accuracy, the change of
impedance is recorded automatically rather than using a manual
measurement. In the present study, the overexpression of FAPα
significantly elevated the migration ability of SK-MES-1 cells 4 h
post-wounding (n=16; SK-MES-1wt vs. SK-MES-1exp, 200.0±173.2 vs.
394.8±254.5 ohms; P=0.001; SK-MES-1pef vs. SK-MES-1exp, 228.0±282.6
vs. 394.8±254.5 ohms; P=0.017). However, the overexpression of FAPα
in A549 cells had no effect on cell migration rate when compared
with control cells 4 h post-wounding (n=16; A549wt vs. A549exp:
578.8±215.7 vs. 610.2±182.7 ohms; P=0.66; A549pef vs. A549exp,
580.2±221.8 vs. 610.2±182.7 ohms; P=0.68) (Fig. 5).
Inhibitors of SHH and PI3K inhibit the
increases in cell attachment and migration induced by the
overexpression of FAPα
To examine the potential interaction of FAPα with
signaling pathways potentially responsible for the increased
adhesive and migratory properties of SK-MES-1 lung cancer cells, a
panel of small-molecule inhibitors of a number of signaling
pathways were screened, including ROCK, FAK, NWASP, ERK1/2, PLCγ,
JNK, PI3K and SHH. Using an ECIS assay (n=8 for each experiment),
the effects of these inhibitors on cell attachment and migration
were assessed. Compared with the SK-MES-1pef cells, only the
inhibitors of SHH and PI3K significantly inhibited the increased
the cell attachment and motility (Fig. 6A–D) of the FAPα-expressing
SK-MES-1exp cells.
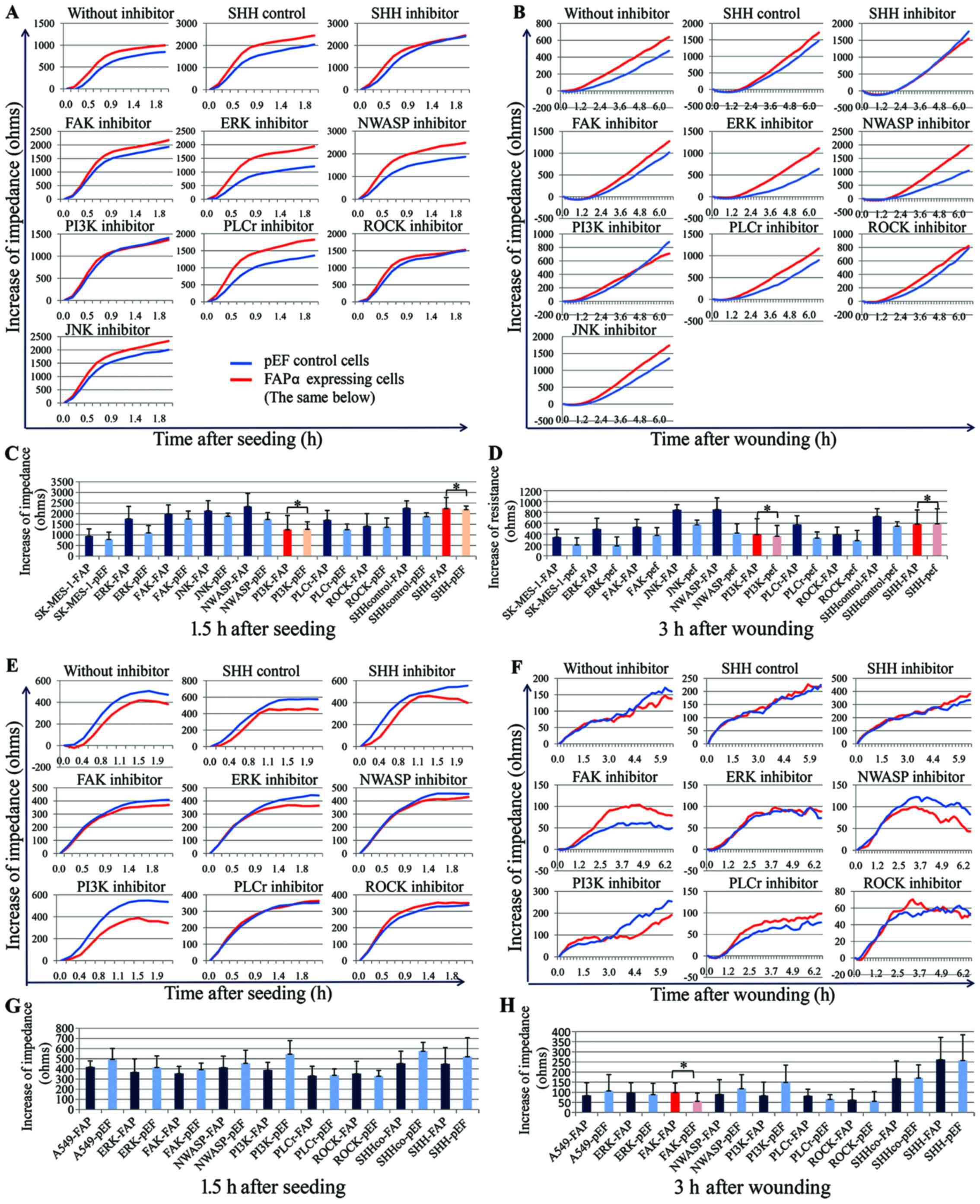 | Figure 6Effects of signaling pathway
inhibitors on the motility of SK-MES-1 lung cancer cells. A panel
of small inhibitors linked to cell motility was screened and only
the PI3K and SHH inhibitors had substantial effects. ECIS was used
to assess the effects of these inhibitors on the attachment and
migration abilities of (A–D) SK-MES-1 cells and (E–H) A549 cells.
The PI3K and SHH inhibitors inhibited the increased (A and C)
attachment and (B and D) migration of SK-MES-1 cells induced by the
overexpression of FAPα. In A549 cells, none of the screened
inhibitors affected (E and G) cell attachment, and only FAK
inhibitor appeared to stimulate (F and H) cell migration, with the
other inhibitors having no marked effect on cell migration.
*P>0.05. The difference between SK-MES-1exp and
SK-MES-1pef cells in the absence of inhibitors was significant
(P<0.05), but addition of inhibitors eliminated the difference.
FAPα, fibroblast activation protein α; FAP, FAPα-expressing cells;
pEF, vector-transfected control cells; ohms, unit of electrical
impedance; SHH, sonic hedgehog; SHHco, control of the SHH inhibitor
cyclopamine; FAK, focal adhesion kinase; ERK, extracellular
signal-regulated kinase; NWASP, neural Wiskott-Aldrich syndrome
protein; PI3K, phosphatidylinositol 3-kinase; PLCγ, phospholipase
C-γ; ROCK, Rho-associated protein kinase; JNK, c-Jun N-terminal
kinase. |
In the earlier cell function assay, the
overexpression of FAPα in A549 cells failed to increase cellular
adhesion and migration. The present study subsequently analyzed the
effects of the above inhibitors on the attachment and motility of
A549 cells. The results showed that there was minimal difference in
the attachment rate of the A549exp cells following treatment with
the inhibitors; only the inhibitor of FAK stimulated the migration
of A549exp cells, with the other inhibitors having no marked effect
on cell migration (Fig.
6E–H).
FAPα regulates cellular functions through
the PI3K/AKT and SHH/Gli1 pathways
To determine whether the effects of FAPα on the
cellular functions of SK-MES-1 cells were associated with the
PI3K/AKT and SHH pathways, western blot analysis was performed to
detect the protein expression levels of total PI3K, AKT, SHH, Ptch
and Gli1. As shown in Fig. 7,
compared with the control cells, the protein levels of AKT and PI3K
were markedly increased in the SK-MES-1exp cells. The protein
levels of SHH and Gli1 were also increased in the SK-MES-1exp
cells. By contrast, the levels of Ptch were similar in the
SK-MES-1exp and control cells.
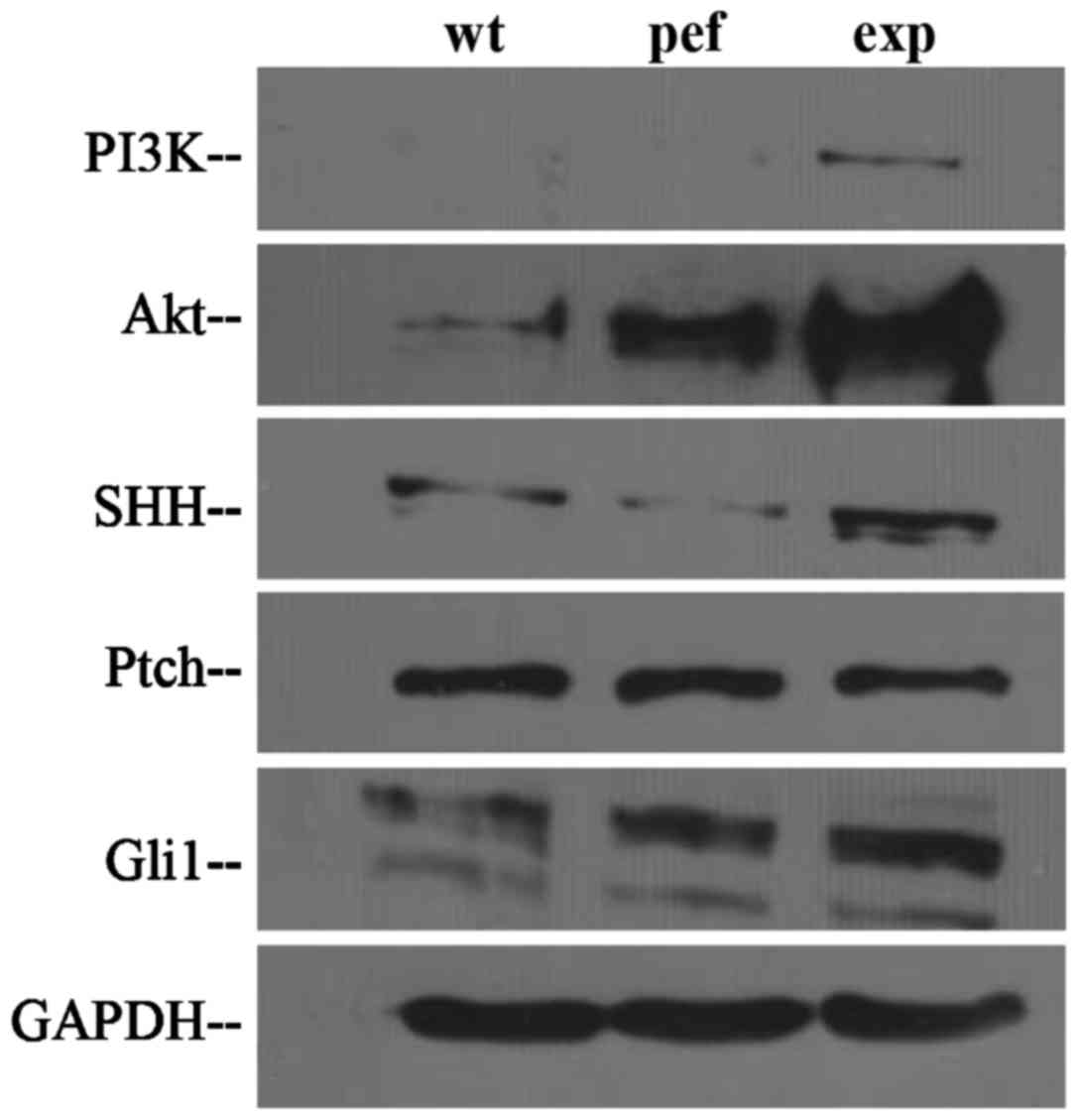 | Figure 7Detection of FAPα-mediated effects on
the PI3K/AKT and SHH pathways in SK-MES-1 lung cancer cells using
western blot analysis. Compared with control cells, the expression
levels of AKT, PI3K, SHH and Gli1 were markedly increased in the
SK-MES-1exp cells. No marked change in expression of Ptch was
observed in the SK-MES-1exp cells. FAPα, fibroblast activation
protein α; exp, FAPα-expressing cells; pef, vector-transfected
control cells; wt, wild-type cells; PI3K,
phosphatidylinositol-3-kinase; AKT, protein kinase B; SHH, sonic
hedgehog; Ptch, Patched; GAPDH, glyceraldehyde 3-phosphate
dehydrogenase. |
Discussion
FAPα is an integral membrane serine peptidase.
Preliminary studies have shown that FAPα is expressed primarily in
fetal mesenchymal tissues, stromal fibroblasts, wounded tissues,
and stromal fibroblasts of malignant epithelial tumors (5–7).
There is increasing evidence that the expression of FAPα is not
confined to stromal fibroblasts but is also present in various
types of epithelium-derived cancer cells (17–20). Therefore, the role of FAPα in
cancer cells may differ from that in fibroblasts. Goodman et
al found that the suppression of FAPα in MDA-MB-435 and
MDA-MB-436 human breast cancer cell lines, which normally express
FAPα, rendered these cells sensitive to serum starvation (26). Cheng et al reported that
mice inoculated with FAPα-transfected HEK293 cells were two to four
times more likely to develop tumors, compared with those inoculated
with FAPα-null control cells (27). Additionally, in an in vivo
mouse model, FAPα-expressing MDA-MB-231 breast cancer cells grew
more rapidly than control cells (28). Previous studies have confirmed
these early findings and suggested that FAPα expressed by cancer
cells may serve as an oncogene. For example, it was found that FAPα
promoted ovarian cancer cell proliferation, drug resistance,
invasiveness and migration in vitro, and that the silencing
of FAPα in SKOV3 cells significantly reduced tumor growth in a
xenograft mouse model (29). Our
previous study (25) showed that
the overexpression of FAPα markedly increased the growth, adhesion,
invasion and migration abilities of MCF-7 breast cancer cells,
whereas the knockdown of FAPα in BT549 breast cancer cells
decreased these abilities. In the present study, the overexpression
of FAPα markedly increased the growth, adhesion and migration of
SK-MES-1 lung cancer cells. Therefore, these results suggested that
the FAPα expressed by epithelial cancer cells had an oncogenic
function. However, the overexpression of FAPα failed to promote the
growth and motility of A549 cells in the present study. One reason
for this may be the difference in cell type between SK-MES-1 (SCC)
and A549 (AC) cells. However, Du et al reported that FAPα
was expressed in AC specimens from patients with lung cancer
(30). Therefore, the role of
FAPα in lung AC cells requires clarification in further
investigations.
The detailed mechanisms by which FAPα in epithelial
cancer cells promote cell growth and motility remains to be fully
elucidated. However, studies have revealed that FAPα has important
non-enzymatic functions, which enable it to regulate the
proliferation and spread of cancer cells (22,23,25). Huang et al found that
inhibitors of FAPα did not decrease the growth of FAPα-expressing
breast cancer cells, and that breast cancer cells expressing a
catalytically inactive mutant of FAPα produced tumors rapidly in a
SCID mouse model (23). Our
previous study also showed that an enzymatic mutant of FAP, in
which the serine catalytic triad was disrupted, markedly increased
cellular growth and motility in MCF-7 breast cancer cells (25). Taken together with previous
results, it appears that the tumorigenicity of FAPα expressed by
cancer cells is not dependent on its enzymatic activity. There is
also increasing evidence that FAPα is involved in the regulation of
signaling pathways. In oral SCC cells, it has been reported that
the knockdown of FAPα inactivated the PTEN/PI3K/AKT and Ras-ERK
signaling pathways, resulting in the suppression of oral SCC cell
proliferation, migration and invasion (24). In ovarian cancer cells, FAPα in
combination with integrin α3β1 and the uPAR signaling complex
mediate cellular migration via the small GTPase Rac1 pathway
(31). Our previous study showed
that the FAPα-mediated promotion of cell growth and motility in
breast cancer cells was accompanied by the upregulation of MMP2/9,
phosphorylated-PI3K and phosphorylated-AKT (25). In addition, FAPα expressed by
stromal cells has been reported to be involved in the regulation of
signaling pathways, including RhoA (32), β-catenin (33) and fibroblast growth factor 1
(FGF1)/FGF receptor 3 (34). In
the present study, the potential interactions of FAPα with a number
of signaling pathways were analyzed, including FAK, ERK1/2, ROCK,
PLCγ, JNK, NWASP, PI3K and SHH. The results showed that, for
SK-MES-1 cells, treatment with PI3K or SHH inhibitors significantly
inhibited the increase in cellular motility induced by the
overexpression of FAPα. The results of the western blot analysis
showed that the overexpression of FAPα was accompanied by increases
in the protein expression levels of PI3K, AKT, SHH and Gli1, which
indicated activation of the PI3K-AKT and SHH-Gli1 pathways.
There remains no direct evidence to show that FAPα
is involved in the EMT process of cancer cells. However, Du et
al analyzed the expression of transforming growth factor-β
(TGF-β), Twist and FAPα in lung cancer tissues, and found that
these three molecules were expressed at various levels in different
lung cancer tissues in situ, including in AC tissue
(30). TGF-β is a multifunctional
protein and has been demonstrated to be involved in promoting EMT
(35). Twist is a basic
helix-loop-helix transcription factor and may promote EMT by
downregulating E-cadherin, either directly or by interacting with
other transcription factors (36). It has also been demonstrated that
the SHH pathway is important in the development and metastasis of
lung cancer. In a previous study, the expression of Gli1 was
associated with the expression of EMT markers E-cadherin and
β-catenin in lung SCC specimens. Inhibition of the SHH/Gli pathway
suppressed the migration and upregulation of E-cadherin in lung SCC
cells, whereas subsequent stimulation of the SHH pathway increased
migration and downregulated the expression of E-cadherin in the
lung SCC cells (37). In
pancreatic cancer, Xu et al found that five members of the
S100 gene family, namely S100A2, S100A4, S100A6, S100A11 and
S100A14, were significantly downregulated upon Gli1 knockdown. The
migration of pancreatic cancer cells was also significantly
increased in a Gli1 expression-dependent manner (38). These results suggest that FAPα may
be indirectly involved in the EMT process by regulating certain
signaling pathways, including SHH and PI3K. Further investigations
are required to investigate the mechanisms involved.
In conclusion, the present study provided evidence
that FAPα promoted cellular growth and migration in lung SCC cells,
and that the PI3K-AKT and SHH-Gli1 signaling pathways may be
involved in the effects of FAPα on lung SCC cells.
Acknowledgments
This study was funded by the Beijing Natural Science
Foundation of Beijing Municipality, China (grant no. 7132048; Dr
Jun Jia) and the Cancer Research Wales and the Albert Hung
Foundation (Professor Wen G. Jiang). Dr Jun Jia is a recipient of
Cardiff's China Medical Scholarship.
Glossary
Abbreviations
Abbreviations:
|
FAPα
|
fibroblast activation protein α
|
|
SHH
|
sonic hedgehog
|
|
PI3K
|
phosphatidylinositol-3-kinase
|
References
|
1
|
Natarajan J, Chandrashekar C and
Radhakrishnan R: Critical biomarkers of epithelial-mesenchymal
transition in the head and neck cancers. J Cancer Res Ther.
10:512–518. 2014.PubMed/NCBI
|
|
2
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li ZH and Bresnick AR: The S100A4
metastasis factor regulates cellular motility via a direct
interaction with myosin-IIA. Cancer Res. 66:5173–5180. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sarrió D, Rodriguez-Pinilla SM, Hardisson
D, Cano A, Moreno-Bueno G and Palacios J: Epithelial-mesenchymal
transition in breast cancer relates to the basal-like phenotype.
Cancer Res. 68:989–997. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Scanlan MJ, Raj BK, Calvo B, Garin-Chesa
P, Sanz-Moncasi MP, Healey JH, Old LJ and Rettig WJ: Molecular
cloning of fibroblast activation protein alpha, a member of the
serine protease family selectively expressed in stromal fibroblasts
of epithelial cancers. Proc Natl Acad Sci USA. 91:5657–5661. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huber MA, Kraut N, Park JE, Schubert RD,
Rettig WJ, Peter RU and Garin-Chesa P: Fibroblast activation
protein: Differential expression and serine protease activity in
reactive stromal fibroblasts of melanocytic skin tumors. J Invest
Dermatol. 120:182–188. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kraman M, Bambrough PJ, Arnold JN, Roberts
EW, Magiera L, Jones JO, Gopinathan A, Tuveson DA and Fearon DT:
Suppression of antitumor immunity by stromal cells expressing
fibroblast activation protein-alpha. Science. 330:827–830. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aertgeerts K, Levin I, Shi L, Snell GP,
Jennings A, Prasad GS, Zhang Y, Kraus ML, Salakian S, Sridhar V, et
al: Structural and kinetic analysis of the substrate specificity of
human fibroblast activation protein alpha. J Biol Chem.
280:19441–19444. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Levy MT, McCaughan GW, Abbott CA, Park JE,
Cunningham AM, Müller E, Rettig WJ and Gorrell MD: Fibroblast
activation protein: A cell surface dipeptidyl peptidase and
gelatinase expressed by stellate cells at the tissue remodelling
interface in human cirrhosis. Hepatology. 29:1768–1778. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Christiansen VJ, Jackson KW, Lee KN and
McKee PA: Effect of fibroblast activation protein and
alpha2-antiplasmin cleaving enzyme on collagen types I, III and IV.
Arch Biochem Biophys. 457:177–186. 2007. View Article : Google Scholar
|
|
11
|
Aggarwal S, Brennen WN, Kole TP, Schneider
E, Topaloglu O, Yates M, Cotter RJ and Denmeade SR: Fibroblast
activation protein peptide substrates identified from human
collagen I derived gelatin cleavage sites. Biochemistry.
47:1076–1086. 2008. View Article : Google Scholar
|
|
12
|
O'Brien P and O'Connor BF: Seprase: An
overview of an important matrix serine protease. Biochim Biophys
Acta. 1784:1130–1145. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lai D, Ma L and Wang F: Fibroblast
activation protein regulates tumor-associated fibroblasts and
epithelial ovarian cancer cells. Int J Oncol. 41:541–550. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wikberg ML, Edin S, Lundberg IV, Van
Guelpen B, Dahlin AM, Rutegård J, Stenling R, Oberg A and Palmqvist
R: High intratumoral expression of fibroblast activation protein
(FAP) in colon cancer is associated with poorer patient prognosis.
Tumour Biol. 34:1013–1020. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saigusa S, Toiyama Y, Tanaka K, Yokoe T,
Okugawa Y, Fujikawa H, Matsusita K, Kawamura M, Inoue Y, Miki C, et
al: Cancer-associated fibroblasts correlate with poor prognosis in
rectal cancer after chemoradiotherapy. Int J Oncol. 38:655–663.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cohen SJ, Alpaugh RK, Palazzo I, Meropol
NJ, Rogatko A, Xu Z, Hoffman JP, Weiner LM and Cheng JD: Fibroblast
activation protein and its relationship to clinical outcome in
pancreatic adenocarcinoma. Pancreas. 37:154–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia J, Martin TA, Ye L and Jiang WG: FAP-α
(Fibroblast activation protein-α) is involved in the control of
human breast cancer cell line growth and motility via the FAK
pathway. BMC Cell Biol. 15:162014. View Article : Google Scholar
|
|
18
|
Okada K, Chen WT, Iwasa S, Jin X, Yamane
T, Ooi A and Mitsumata M: Seprase, a membrane-type serine protease,
has different expression patterns in intestinal- and diffuse-type
gastric cancer. Oncology. 65:363–370. 2003. View Article : Google Scholar
|
|
19
|
Kashyap MK, Marimuthu A, Kishore CJ, Peri
S, Keerthikumar S, Prasad TS, Mahmood R, Rao S, Ranganathan P,
Sanjeeviah RC, et al: Genomewide mRNA profiling of esophageal
squamous cell carcinoma for identification of cancer biomarkers.
Cancer Biol Ther. 8:36–46. 2009. View Article : Google Scholar
|
|
20
|
Zhang MZ, Qiao YH, Nesland JM, Trope C,
Kennedy A, Chen WT and Suo ZH: Expression of seprase in effusions
from patients with epithelial ovarian carcinoma. Chin Med J (Engl).
120:663–668. 2007.
|
|
21
|
Iwasa S, Okada K, Chen WT, Jin X, Yamane
T, Ooi A and Mitsumata M: 'Increased expression of seprase, a
membrane-type serine protease, is associated with lymph node
metastasis in human colorectal cancer'. Cancer Lett. 227:229–236.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang XM, Yu DM, McCaughan GW and Gorrell
MD: Fibroblast activation protein increases apoptosis, cell
adhesion, and migration by the LX-2 human stellate cell line.
Hepatology. 42:935–945. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang Y, Simms AE, Mazur A, Wang S, León
NR, Jones B, Aziz N and Kelly T: Fibroblast activation protein-α
promotes tumor growth and invasion of breast cancer cells through
non-enzymatic functions. Clin Exp Metastasis. 28:567–579. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang H, Wu Q, Liu Z, Luo X, Fan Y, Liu Y,
Zhang Y, Hua S, Fu Q, Zhao M, et al: Downregulation of FAP
suppresses cell proliferation and metastasis through PTEN/PI3K/AKT
and Ras-ERK signaling in oral squamous cell carcinoma. Cell Death
Dis. 5:e11552014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lv B, Xie F, Zhao P, Ma X, Jiang WG, Yu J,
Zhang X and Jia J: Promotion of cellular growth and motility is
independent of enzymatic activity of fibroblast activation
protein-α. Cancer Genomics Proteomics. 13:201–208. 2016.PubMed/NCBI
|
|
26
|
Goodman JD, Rozypal TL and Kelly T:
Seprase, a membrane-bound protease, alleviates the serum growth
requirement of human breast cancer cells. Clin Exp Metastasis.
20:459–470. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cheng JD, Dunbrack RL Jr, Valianou M,
Rogatko A, Alpaugh RK and Weiner LM: Promotion of tumor growth by
murine fibroblast activation protein, a serine protease, in an
animal model. Cancer Res. 62:4767–4772. 2002.PubMed/NCBI
|
|
28
|
Huang Y, Wang S and Kelly T: Seprase
promotes rapid tumor growth and increased microvessel density in a
mouse model of human breast cancer. Cancer Res. 64:2712–2716. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang L, Ma L and Lai D: Over-expression of
fibroblast activation protein alpha increases tumor growth in
xenografts of ovarian cancer cells. Acta Biochim Biophys Sin
(Shanghai). 45:928–937. 2013. View Article : Google Scholar
|
|
30
|
Du H, Chen D, Zhou Y, Han Z and Che G:
Fibroblast phenotypes in different lung diseases. J Cardiothorac
Surg. 9:1472014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang W, Han W, Ye S, Liu D, Wu J, Liu H,
Li C and Chen H: Fibroblast activation protein-α promotes ovarian
cancer cell proliferation and invasion via extracellular and
intracellular signaling mechanisms. Exp Mol Pathol. 95:105–110.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chung KM, Hsu SC, Chu YR, Lin MY, Jiaang
WT, Chen RH and Chen X: Fibroblast activation protein (FAP) is
essential for the migration of bone marrow mesenchymal stem cells
through RhoA activation. PLoS One. 9:e887722014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zi FM, He JS, Li Y, Wu C, Wu WJ, Yang Y,
Wang LJ, He DH, Yang L, Zhao Y, et al: Fibroblast activation
protein protects bortezomib-induced apoptosis in multiple myeloma
cells through β-catenin signaling pathway. Cancer Biol Ther.
15:1413–1422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Henriksson ML, Edin S, Dahlin AM,
Oldenborg PA, Åberg Ö, Van Guelpen B, Rutegård J, Stenling R and
Palmqvist R: Colorectal cancer cells activate adjacent fibroblasts
resulting in FGF1/FGFR3 signaling and increased invasion. Am J
Pathol. 178:1387–1394. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Katsuno Y, Lamouille S and Derynck R:
TGF-β signaling and epithelial-mesenchymal transition in cancer
progression. Curr Opin Oncol. 25:76–84. 2013. View Article : Google Scholar
|
|
36
|
Sánchez-Tilló E, Liu Y, de Barrios O,
Siles L, Fanlo L, Cuatrecasas M, Darling DS, Dean DC, Castells A
and Postigo A: EMT-activating transcription factors in cancer:
Beyond EMT and tumor invasiveness. Cell Mol Life Sci. 69:3429–3456.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yue D, Li H, Che J, Zhang Y, Tseng HH, Jin
JQ, Luh TM, Giroux-Leprieur E, Mo M, Zheng Q, et al: Hedgehog/Gli
promotes epithelial-mesenchymal transition in lung squamous cell
carcinomas. J Exp Clin Cancer Res. 33:342014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xu X, Su B, Xie C, Wei S, Zhou Y, Liu H,
Dai W, Cheng P, Wang F, Xu X, et al: Sonic hedgehog-Gli1 signaling
pathway regulates the epithelial mesenchymal transition (EMT) by
mediating a new target gene, S100A4, in pancreatic cancer cells.
PLoS One. 9:e964412014. View Article : Google Scholar : PubMed/NCBI
|