Introduction
Peritoneal dialysis (PD), based on the peritoneum as
a semi-permeable membrane, aims to filter out solutes for blood
purification (1). PD is an
economic and simple dialysis method, which can remove toxic
molecules and improve the survival rate (2). However, long-term exposure to a
hypertonic and high glucose environment can induce inflammation,
epithelial cell differentiation and angiogenesis, resulting in
fibrosis (3). Peritoneal fibrosis
(PF) is associated with abnormal histological changes in peritoneal
tissues and parenchymal cells necrosis caused by fibrous connective
tissue matrix deposition, which is also the main cause of
ultrafiltration dialysis failure (4). The exact molecular mechansisms
underlying the disease pathogenesis remain unclear. However, it has
been reported that a combination of mechanisms are involved in PF
pathogenesis, including a high peritoneal dialysate glucose
concentration, high osmotic pressure, low pH value, advanced
glycation end products (AGEs) and bacterial peritonitis (5). It has been demonstrated that a
number of cytokines are associated with PF, including transforming
growth factor (TGF)-β1, connective-tissue growth factor (CTGF),
vascular endothelial growth factor (VEGF) and platelet-derived
growth factor (PDGF) (6). In
addition, a previous study revealed that the activation of the
TGF-β/Smad signaling pathway in patients with PF confirms the
opposite role of Smad2 and Smad3 in PF and their involvement in the
progression of PF (7). In
addition, Zheng et al reported that Astragalus
membranaceus inhibited PF via the monocyte chemoattractant
protein (MCP)-1 and TGF-β1 pathways, exerting anti-fibrotic and
therapeutic effects (8). These
studies provide insight into the treatment of PF via the targeting
of TGF-β/Smad signaling.
The gene coding for microRNA (miR)-21 is located on
17q23.2 within the TMEM49 coding region and plays an important role
in DNA-dependent transcriptional regulation, which is of great
significance in the development of fibrotic diseases (9). TGF-β is a pleiotropic growth factor
that regulates cell proliferation, differentiation and the immune
response as well as apoptosis, and mainly exerts its role in tissue
fibrosis, stimulation, mesenchymal cell growth and collagen and
fibronectin increase (10). Smad
protein (Drosophila mothers against decapentaplegic protein)
can be divided into 3 subfamilies: receptor-regulated Smads
(R-Smads), common-mediator Smads (Co-Smads) and inhibitory Smads
(I-Smads). There are 8 types of Smad proteins in mammals and they
are TGF-β-mediated signal transduction proteins, and the TGF-β/Smad
signaling pathway is a key pathway in the occurrence and
progression of fibrosis, involved with a variety of fibrotic
diseases (11).
Certain experimental studies have detected an
abnormal miR-21 expression various organs affected by fibrosis;
thus, the investigation of the function of miR-21 in fibrosis has
implications for the elucidation of the pathogenesis, diagnosis and
treatment of fibrosis (7-9). Thus, this study aimed to explore the
mechanisms of action of miR-21 in PF and its association with the
TGF-β/Smad pathway using dialysis effluents from patients with PF
and by establishing a mouse model of PF.
Materials and methods
Study subjects
Patients with PF receiving a stable treatment for PD
at the Huaihe Hospital of Henan university, Henan, China from
October, 2014 to June, 2015 were selected for the study. The 30
patients included were >18 years of age and had received
dialysis for >3 months. More specifically, the study subjects
consisted of 23 males and 7 females with an average age of 58.5
years, whose disease duration ranged from half a month to 3 years,
with an average of 15.2 months. The disease for each included
subject was diagnosed from 3 days to 5.1 months from admission to
the hospital with 2.1 months on average. The diagnosis was
conducted and confirmed through clinical manifestations and imaging
examination. Clinical manifestations were shown as follows: back
pain, leg edema, abdominal pain, lower abdominal mass, and the
remaining symptoms included bloating, headaches, nausea and
vomiting. The diagnostic method for the imaging examination
included a B-ultrasound examination firstly, followed by a CT scan
after hydronephrosis and ureterectasia were found. Ureteral
compression was also examined if the soft tissue density was found
to be below the abdominal aortic bifurcation (12). According to the length of the
dialysis treatment, there were a total of 3 groups with a 1-year
interval. For the dialysis treatment duration, there were 9 cases
with <1-year duration; 14 cases with a 1-2-year duration and 7
cases with a 2-3-year duration. During peritoneal dialysis, the
omentum was obtained for the collection of mesothelial cells. This
study was approved by the Ethics Committee of Huaihe Hospital of
Henan University. All enrolled patients had signed an informed
consent. Patients suffering from a mental illness or unable to
cooperate were excluded from this study.
Establishment of the mouse model of
PF
Male Kunming mice at the specific pathogen-free
(SPF) level and weighing approximately 30-40 g, were provided by
the Animal Research Department of Nanchang University, Nanchang,
China. All mice were reared in an SPF level environment, with a
temperature of 25±1°C and a constant humidity of 40-60%, and free
access to food and water. The feeding lasted for 7 days. A total of
20 mice were randomly divided into 2 groups with 10 mice in each
group, the experimental and control groups, respectively. The mice
in the experimental group were intraperitoneally injected with 0.1
mg/kg lipopolysaccharide on day 1, 3, 5 and 7 and with 4.25%
glucose dialysate (100 ml/kg) (13) each day (lasting 27 days). On day
28, the mice were sacrificed by cervical dislocation. All
procedures involving animals were performed according the Guide for
the Care and Use of Laboratory Animals. The animal experiments were
also approved by the Ethics Committee of Huaihe Hospital of Henan
University, while the peritoneal mesothelial cells from 3 mice in
each group were collected every 3 days during peritoneal dialysis.
For the control group, 0.9% saline was intraperitoneally injected
daily, lasting for 28 days.
Peritoneal equilibrium test (PET)
A PET assay was performed 28 days after modeling.
The mice were substituted with 20 ml of 2.5% glucose-based
dialysate. Two hours later, the mice were sacrificed by cervical
dislocation. The catheter drainage was measured, marked as S1, and
the rest of glucose-based dialysate was collected using a straw,
marked as S2. Ultrafiltration (ml) = S1 + S2 -20. The endpoint
method was used to measure the dialysate urea concentration (D),
plasma urea concentration (Purea), the initial glucose
concentration of peritoneal dialysis solutions (D0) and
the glucose concentration of the dialysate effluent
(D2), and the results of D/Purea and
D2/D0 were calculated. Finally, the parietal
peritoneum was fixed in 1% formaldehyde for pathological
examination.
Hematoxylin and eosin (H&E) staining
and Masson's staining
The visceral peritoneum specimens from the 2 groups
of mice were soaked in paraformaldehyde solution and then
dehydrated, embedded in paraffin, sectioned (4 µm
thickness), and then baked for 12 h in an oven at 56°C. Through
H&E and Masson's staining, 10 visual fields were selected
randomly (x200 magnification) and 5 positions were taken in each
field. Image-Pro Plus 6.0 software was used for the analysis.
Peritoneal tissue thickness, the inflammatory cell number, and the
number of blood vessels were also measured.
Peritoneal mesothelial cell culture and
transfection
HMrSV5 human peritoneal mesothelial cells were
obtained from the Institute of Nephrology in the First Affiliated
Hospital of Sun Yat-sen University. The HMrSV5 cells were cultured
in DMEM-F12 medium (Gibco, Grand Island, NY, USA) containing 10%
fetal bovine serum (FBS) in 37°C with 5% CO2. The groups
of peritoneal mesothelial cells were as follows: i) blank group
(untreated peritoneal mesothelial cells); ii) high glucose group
(4.5% high glucose-induced peritoneal mesothelial cells); iii) high
glucose + miR-21 inhibitor group (4.5% high glucose-induced
peritoneal mesothelial cells were to be treated with miR-21
inhibitors); iv) high glucose + miR-21 inhibitor control group
(4.5% high glucose-induced peritoneal mesothelial cells were to be
treated with miR-21 inhibitor negative controls). The miR-21
inhibitors and inhibitor negative controls were purchased from
Biomics Biotechnology Corp. (Nantong, China). Twenty-four hours
before treatment, high glucose-induced peritoneal mesothelial cells
were seeded in a 6-well plate. When cell confluence reached 50%,
they were incubated in a mixture of 250 µl serum-free
Opti-MEM diluted miR-21 inhibitors (4.0 µg) or miR-21
inhibitor NC (4.0 µg) and 250 µl serum-free Opti-MEM
diluted lipofectamin 2000 (10 µl; Invitrogen, Carlsbad, CA,
USA) for 6-8 h in an atmosphere of 5% CO2 at 37°C. When
the cells were incubated for 24 h, the total RNA was extracted; 72
h later, total protein was extracted. The expressio levels of
miR-21, Zonula occludens-1 (ZO-1), TGF-β, Smad-3, vimentin, and
CTGF were detected, and some of the cells were retained for
fluorescence microscopy.
Immunofluorescence
The immunofluorescence method was applied to observe
he transfected cells, and fibronectin was used as a reference for
fibrosis (14). Fibronectin
antibody (ab2413) and biotin-labeled goat anti-rabbi IgG (1:1,000)
were purchased from Abcam Inc. (Cambridge, MA, USA). Primary
antibodies (20 µg/ml) were diluted with 1% FBS, and
secondary antibodies were diluted to 1:200. After the cells were
grown on coverslips, the primary antibody was added, followed by
incubation at 37°C for 2 h; the secondary antibody was then added,
followed by incubation at 37°C for 1 h; finally 5 µg/ml DAPI
were added the cells were stained for 5 min. A fluorescence
microscope (Nikon, Tokyo, Japan) was used for the observation.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The recovered peritoneal mesothelial cells from
patients and mice were collected every 3 days, and the extraction
of total RNA from the transfected cell lines was performed using an
RNeasy mini kit (Qiagen, Hilden, Germany) according to the
manufacturer's instructions. A UV Spectrophotometer (NanoDrop,
Walsingreen Co., Beijing, China) was used to measure the A260/A280
value and RNA concentration. RNA was reverse transcribed into cDNA
using a reverse transcription kit (Promega, Madison, WI, USA). A
fluorescence quantitative PCR kit was purchased from Shanghai
GenePharma Technology Co., Ltd., Shanghai, China). An ABI 7500
real-time PCR instrument (Applied Biosystem, Foster City, CA, USA)
was used for the PCR reaction. The reaction volume was 20
µl, and the reaction conditions were 40 cycles at 95°C for 3
min, 95°C for 12 sec and 62°C for 50 sec. miR-21 expression was
measured quantitatively using u6 as an internal control, and the
mRNA expression levels of ZO-1, TGF-β, Smad, vimentin and CTGF were
measured using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as
an internal reference gene. The primers used are shown in Table I. Relative expression was
calculated using the 2−ΔΔCq method.
 | Table IPrimers used for RT-qPCR. |
Table I
Primers used for RT-qPCR.
| Gene | Primer sequences |
|---|
| miR-21 | F:
5′-GCGGTAGCTTATCAGACTG-3′ |
| R:
5′-TGCGTGTCGTGGAGTC-3′ |
| U6 | F:
5′-CTCGCTTCGGCAGCACA-3′ |
| R:
5′-AACGCTTCACGAATTTGCGT-3′ |
| ZO-1 | F:
5′-AATGAATGATGGTTGGTATGG-3′ |
| R:
5′-TGACAGGTAGGACAGACG-3′ |
| TGF-β | F:
5′-GTTCTTCAATACGTCAGACATTCG-3′ |
| R:
5′-CATTATCTTTGCTGTCACAAGAGC-3′ |
| Smad-3 | F:
5′-AAATGACAGCAGCAGGGACACTA-3′ |
| R:
5′-TGAGGAGGTAGGACCCACAGTAGA-3′ |
| Vimentin | F:
5′-CGTTTCCAAGCCTGACCTCACC-3′ |
| R:
5′-GCCATCTTTACATTGAGCAGGT-3′ |
| CTGF | F:
5′-TGTGAAGACCTACCGGGCTA-3′ |
| R:
5′-TTCATGATCTCGCCATCGGG-3′ |
| GAPDH | F:
5′-CGCTGAGTACGTCGTGGAGT-3′ |
| R:
5′-GTCGCTGTTGAAGTCAGAGGAG-3′ |
Western blot analysis
The cells and tissues were washed with
phosphate-buffered saline (PBS), followed by the addition of 100
µl of cell lysates and incubation at 4°C for 30 min.
Centrifugation was performed at 12,000 × g for 10 min. The
supernatant was collected and the protein concentration was
measured using the Bradford method. The total protein (5 µg)
extracted was electrophoresed on a 12% sodium dodecyl sulfate
(SDS)-polyacrylamide gel. After being transferred onto
polyvinylidene fluoride (PVDF) membranes (Jiuding High Tech. Ltd.,
Beijing, China), the total protein was sealed with 5% dried skimmed
milk at room temperature for 1 h. The membranes were then incubated
with mouse anti human monoclonal antibodies to ZO-1 (ab61357; 5
µg/ml), TGF-β (ab64715, 1-2 µg/ml), Smad (ab75512,
diluted at 1:5,000), vimentin (ab8978, diluted at 1:500), CTGF
(ab94939, 10 µg/ml) and GAPDH (ab8245, diluted at 1:1,000)
(all from Abcam) overnight at 4°C. After washing, the membranes
were incubated with IRDye™ 800DX marked goat anti-mouse IgG
(ab6789, Abcam; diluted at 1:10,000) for 1 h. After being
cultivated in the dark at room temperature for 1 h, the membranes
were washed and placed into the Odyssey two-color infrared laser
imaging system (LICOR Biosciences, Lincoln, NE, USA) for scanning
and imaging. The integral optical density of each strip was
calculated, and the ratio of the integral optical density value of
the target band with the integrated optical density value of the
reference GAPDH, was the relative expression value of the target
protein. This experiment was repeated 3 times.
Statistical analysis
IBM SPSS statistical software (version 19.0; SPSS,
Inc., Chicago, IL, USA) was used for statistical analysis. Count
data was expressed in rate or percentage, and the Chi-square test
was used to make comparisons between 2 groups. Measurement data in
groups were compared using One-way ANOVA (prior to analysis,
detection of homogeneity of variance was performed), and the
LSD-t-test of mean value was applied between 2 groups. Correlation
analysis referred to Pearson's correlation analysis. A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
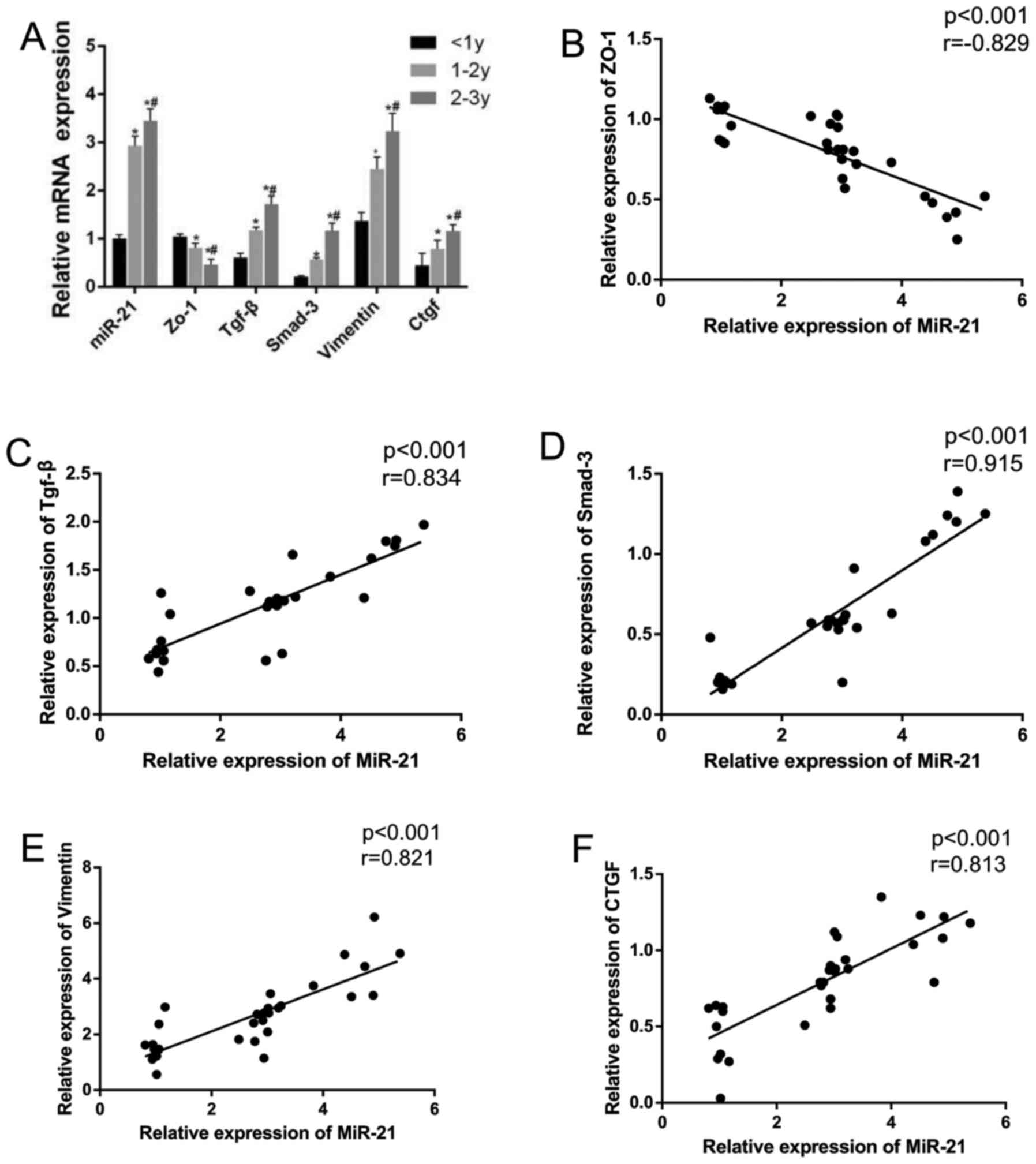
Expression of miR-21 and mRNA expression
levels of TGF-β, Smad-3, vimentin and CTGF in patients with PF
receiving dialysis
The expression of miR-21 and the mRNA expression
levels of TGF-β, Smad-3, vimentin and CTGF were increased with the
increased duration of the dialysis sessions, while the expression
of ZO-1 was decreased (all P<0.05). In the patients receiving
dialysis treatment for a course of 2–3 years, all expression of
indicators exhibited a significant difference compared to those of
patients receiving dialysis treatment for 1–2 years or <1 year
(all P<0.05) (Fig. 1).
Correlation analysis revealed that miR-21 expression positively
correlated with the mRNA expression of TGF-β, Smad-3, vimentin and
CTGF (r=0.997, P<0.001; r= 0.980, P<0.001; r= 0.911,
P<0.001; r= 0.876, P<0.001), while miR-21 expression was
negatively correlated with ZO-1 expression (r=-0.826, P<0.001)
(Fig. 1).
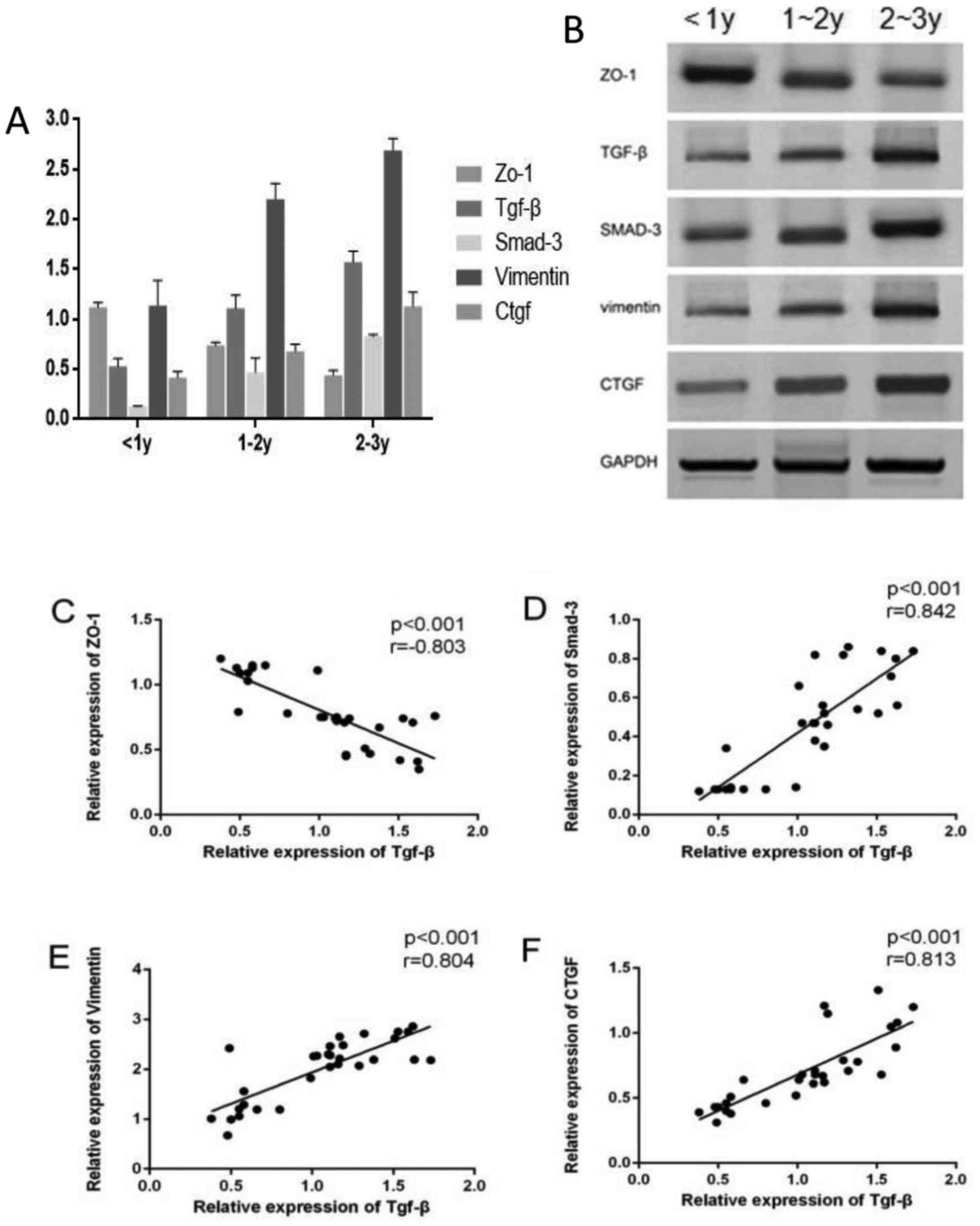
Protein expression levels of ZO-1, TGF-β,
Smad-3, vimentin and CTGF in patients with PF receiving
dialysis
The protein expression levels of TGF-β, Smad-3,
vimentin and CTGF were increased with the increased duration of the
treatment course, while ZO-1 protein expression decreased (all
P<0.05) (Fig. 2). The
expression of TGF-β was found to have a significant and positive
correlation with the expression levels of Smad-3, vimentin and CTGF
(r=0.978, P<0.001; r=0.981, P<0.001; r= 0.961, P<0.001,
while TGF-β expression was negatively correlated with ZO-1
(r=-0.913, P<0.001).
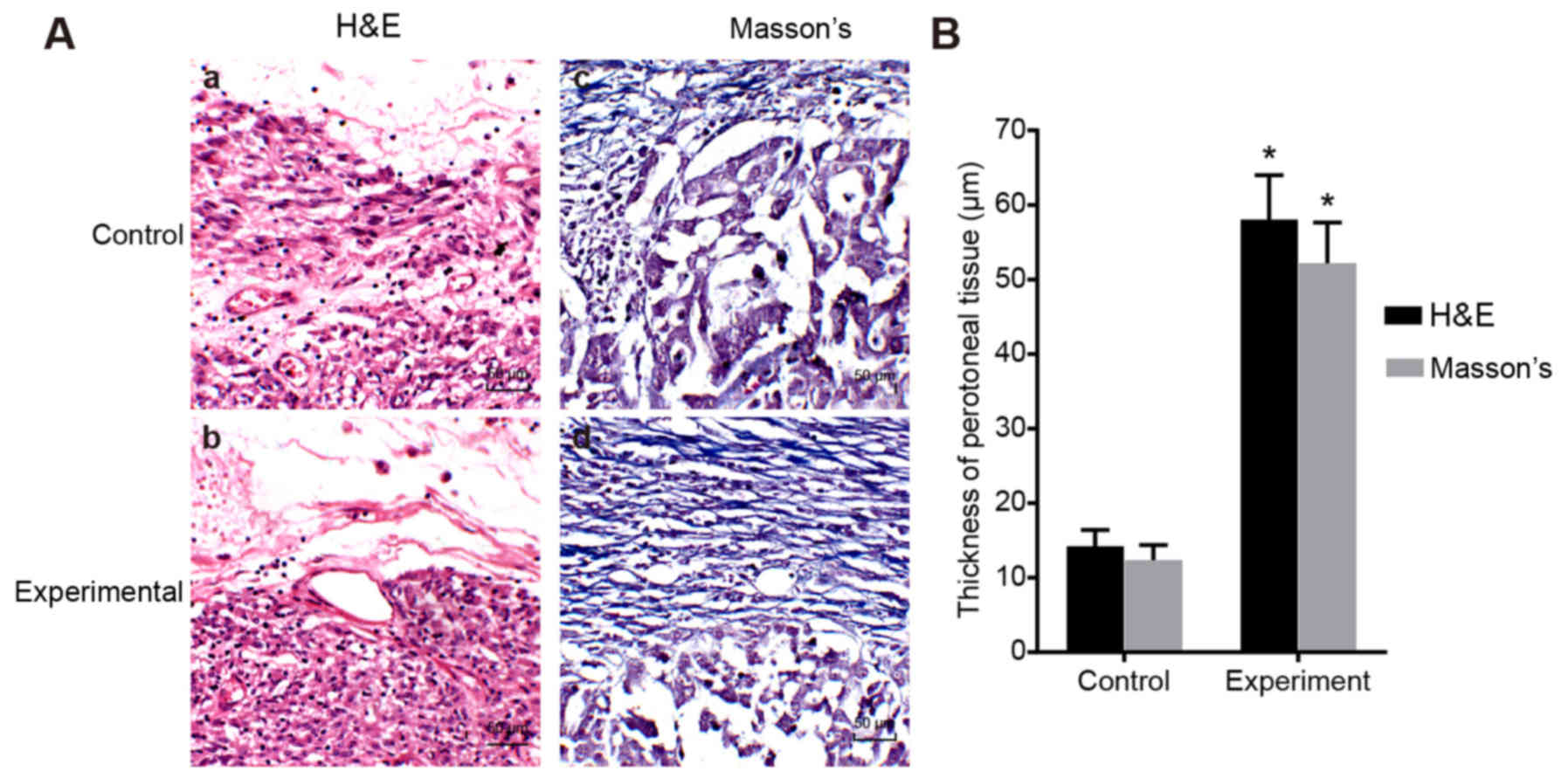
Peritoneal function and histological
analysis of mice in the experimental and control group
The PET results revealed that the ultrafiltration
volume and the D2/D0 were decreased (all
P<0.01), and that the D/Purea was increased in the
experimental group compared with the control group (P<0.05;
Table II). The H&E staining
results revealed that the wall of the peritoneal layer of the mice
in the experimental group had thickened significantly, and the
amount of fibrous tissue had significantly increased in the
experimental group. From the results of Masson's staining, the
cytoplasm in the experimental group had shrunk significantly and
the outer wall of the fibrous tissue was significantly thicker
compared with the control group (P<0.05; Fig. 3). As observed from the two
staining images, we found that the numbers of inflammatory cells
and vascular tissues in the experimental group were significantly
higher than those of the control group (P<0.05; Table II).
| Table IIChanges in peritoneal function in mice
in the experimental and control groups. |
Table II
Changes in peritoneal function in mice
in the experimental and control groups.
| Groups | Control group | Experimental
group | P-value |
|---|
| ultrafiltration
volume/ml | 9.05±1.59 | 1.02±0.32 | <0.001 |
|
D/Purea | 0.56±0.11 | 0.79±0.18 | 0.024 |
|
D2/D0 | 0.44±0.12 | 0.16±0.08 | <0.001 |
| Inflammatory
cells | 111.15±15.32 | 167.51±15.28 | <0.001 |
| Vascular
tissues | 23.40±5.93 | 61.26±14.77 | <0.001 |
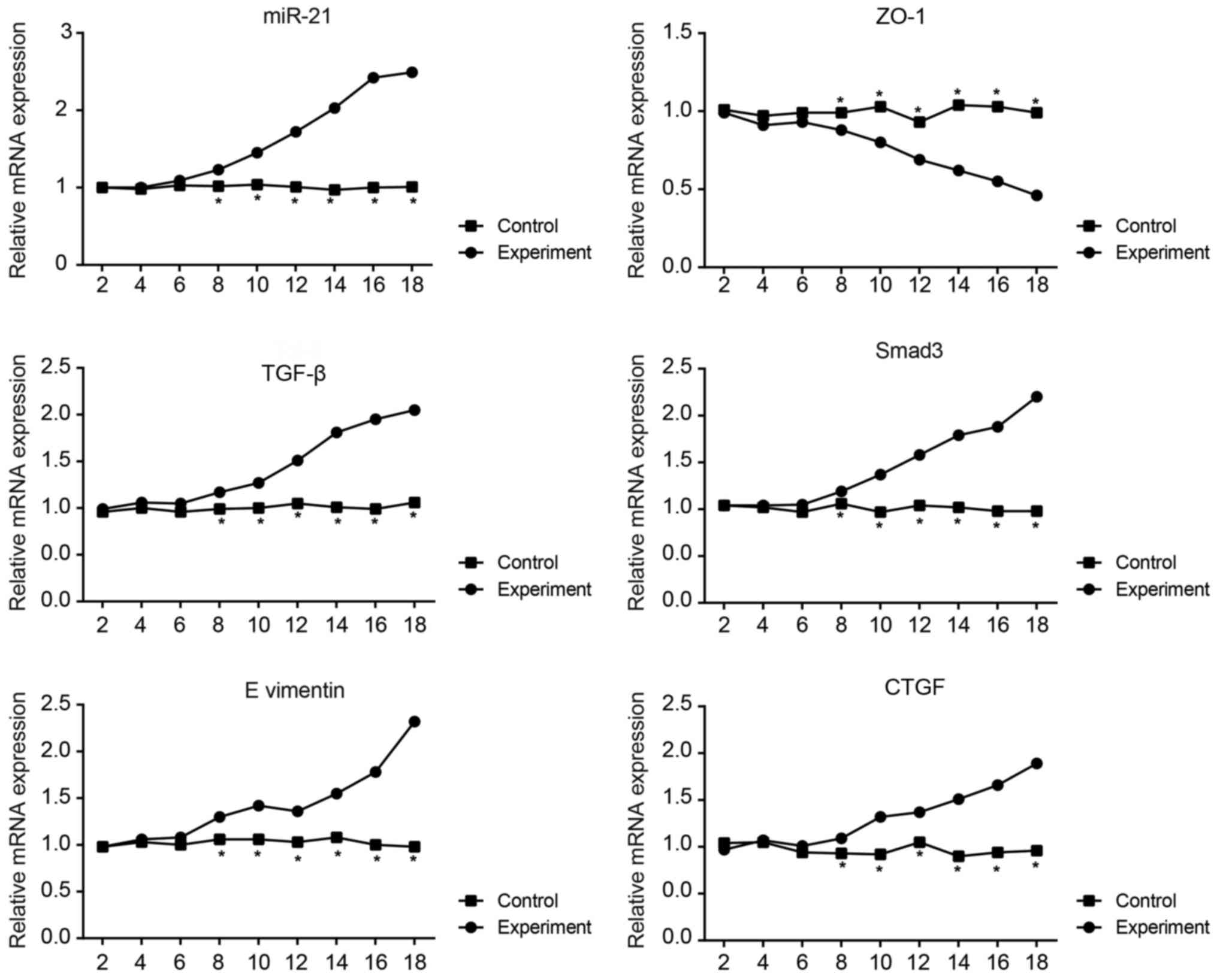
Expression of miR-21 and the mRNA
expression levels of ZO-1, TGF-β, Smad-3, vimentin and CTGF in the
mouse model of PF
The results revealed that the miR-21, TGF-β, Smad-3,
vimentin and CTGF expression levels in the peritoneal mesothelial
cells from the experimental group tended to increase over time,
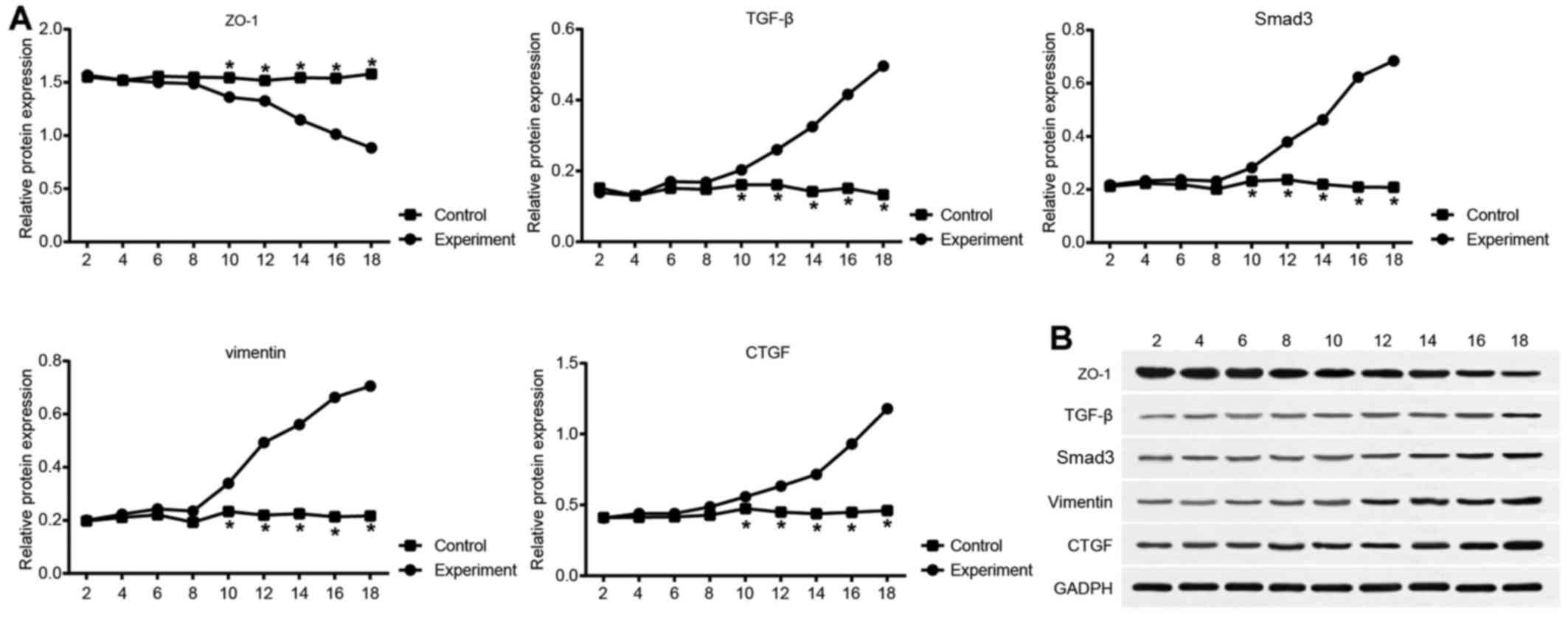
while ZO-1 expression tended to decrease (Fig. 4). The expression of each
biomolecule in the control group remained relatively stable. From
day 8, molecule expression in the experimental group differed
significantly compared with the control group (all P<0.05).
Protein expression levels of ZO-1, TGF-β,
Smad-3, vimentin and CTGF in the mouse model of PF
The results of western blot analysis for protein
expression are shown in Fig. 5.
Compared with the control group, differences in the expression
levels of molecules in the experimental group were not significant
on days 1-3. Each molecule expression level in the experimental
group changed over time, with the expression of TGF-β, Smad,
vimentin and CTGF increasing, and that of ZO-1 decreasing. However,
no significant changes in expression were observed in the control
group. From day 10, molecule protein expression in the experimental
group exhibited a statistically significant difference compared
with the control group (all P<0.05).
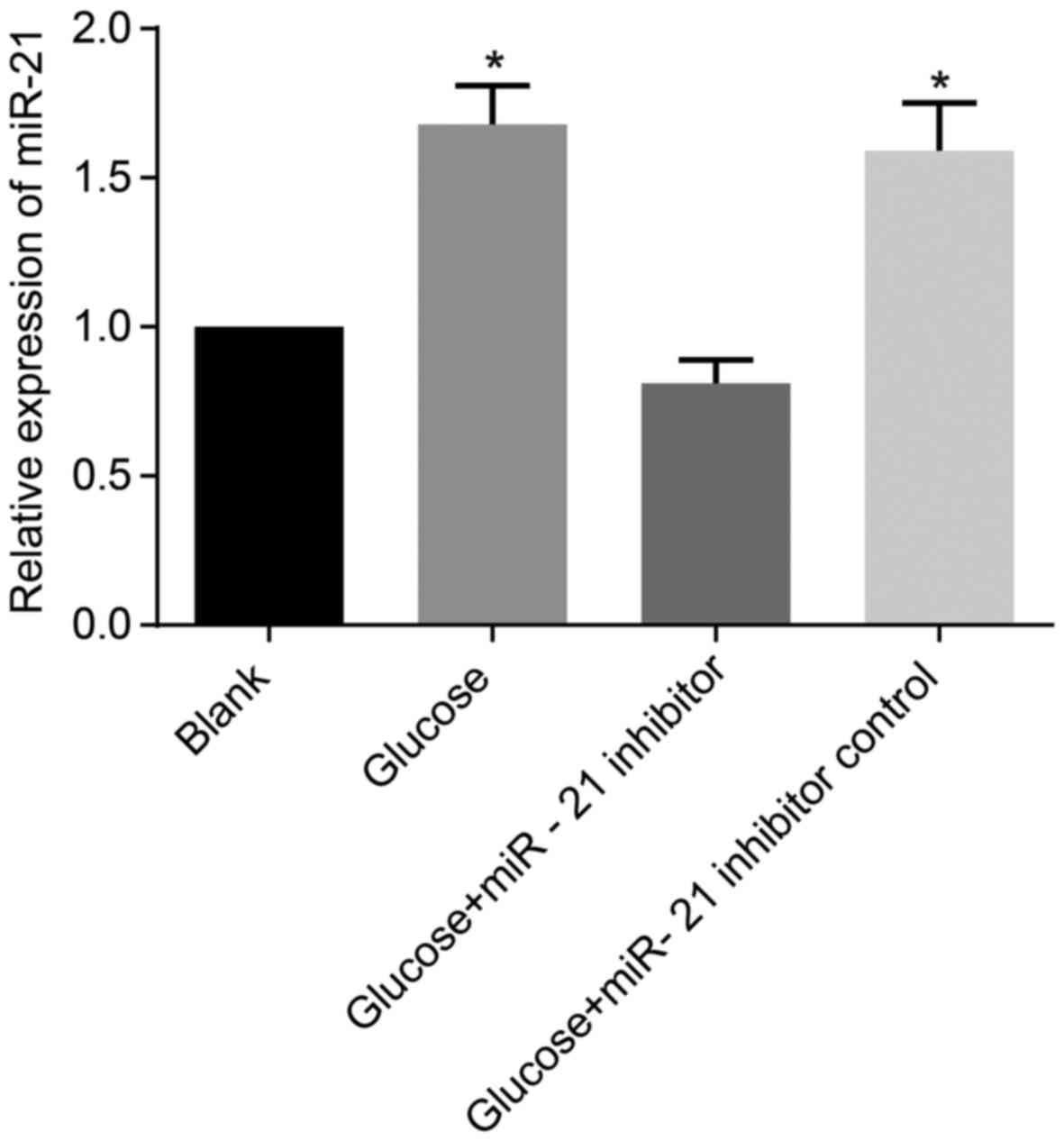
The miR-21 expression in HMrSV5 cells
following transfection with miR-21 inhibitor
The miR-21 expression levels in the cells in the
high glucose group and the high glucose + inhibitor control group
were significantly higher than those of the cells in the blank
group (all P<0.05). In the high glucose + miR-21 inhibitor
group, miR-21 expression exhibited no significant difference
compared with the blank group (P>0.05; Fig. 6).
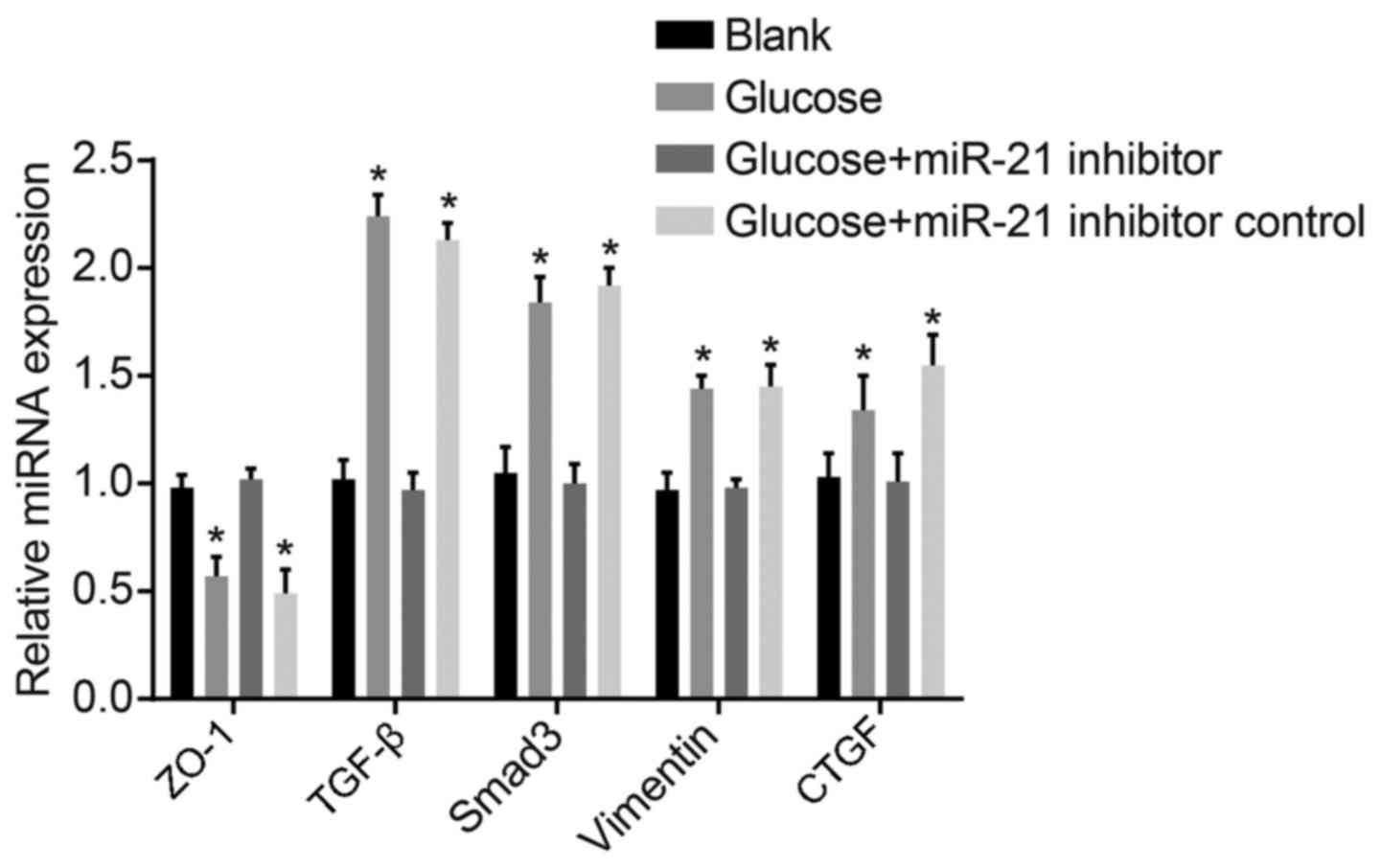
mRNA expression levels of ZO-1, TGF-β,
Smad-3, vimentin and CTGF in HMrSV5 cells following transfection
with miR-21 inhibitor
Compared with the blank group, the expression levels
of TGF-β, Smad-3, vimentin and CTGF in the high glucose group and
high glucose + inhibitor control group were higher, while those of
ZO-1 were decreased. Compared with the high glucose group and high
glucose + inhibitor control group, the TGF-β, Smad-3, vimentin and
CTGF expression levels in the high glucose + miR-21 inhibitor group
were significantly decreased, while those of ZO-1 were increased
(both P<0.05; Fig. 7). Hence,
miR-21 was proven to enhance TGF-β, Smad-3, vimentin and CTGF
expression, and to decrease ZO-1 expression, thus playing an
important regulatory role in PF.
Protein expression levels of ZO-1, TGF-β,
Smad-3, vimentin and CTGF in HMrSV5 cells following transfection
with miR-21 inhibitor
The protein expression levels of TGF-β, Smad,
vimentin and CTGF in both the high glucose group and high glucose +
inhibitor control group were higher than those in the blank group,
with the level of ZO-1 significantly decreased (P<0.05).
However, compared with the high glucose group and high glucose +
inhibitor control group, TGF-β, Smad, vimentin and CTGF expression
levels in the high glucose + miR-21 inhibitor group were
significantly decreased, while those of ZO-1 were significantly
increased (P<0.05; Fig.
8).
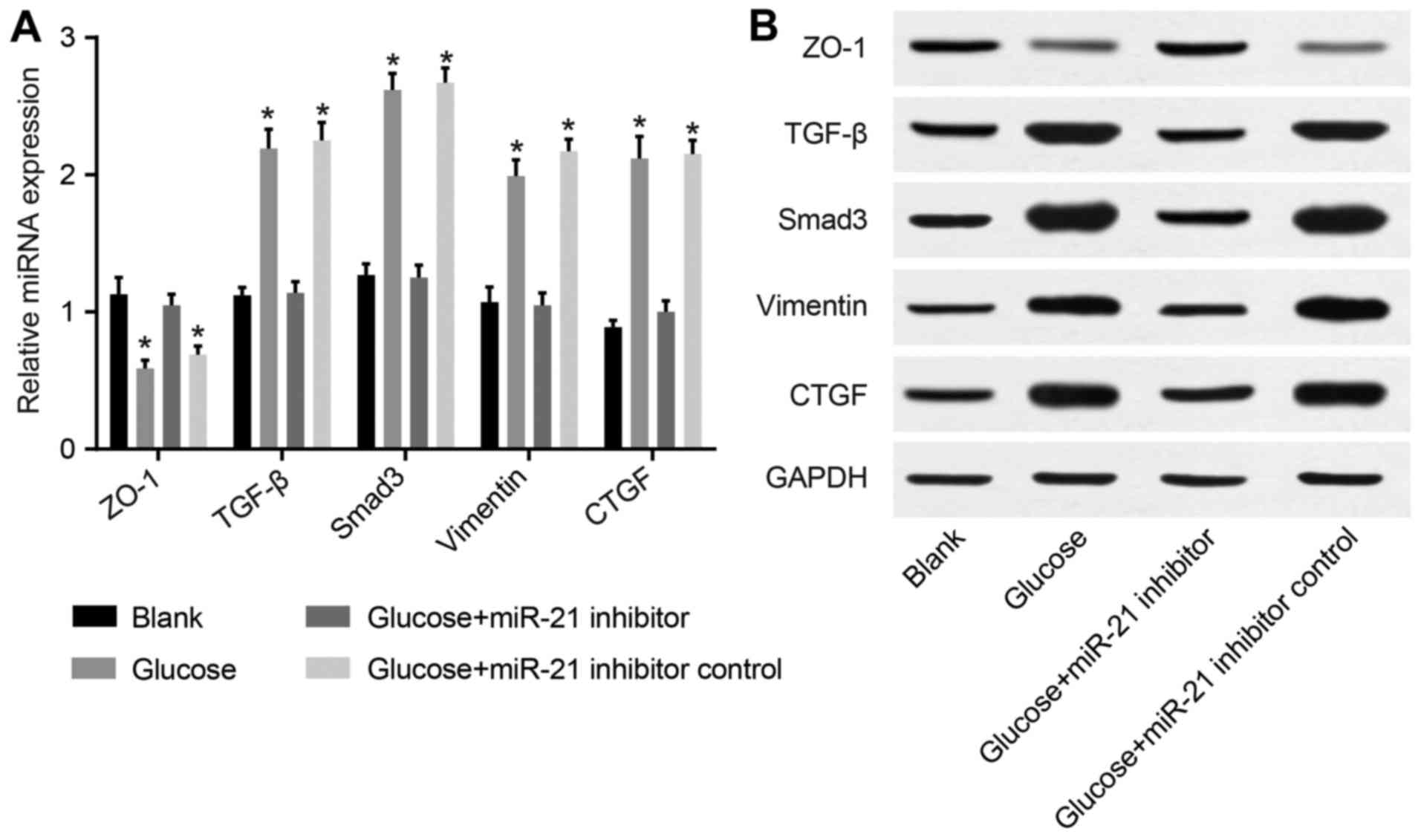 | Figure 8Comparison of the protein expression
of Zonula occludens-1 (ZO-1), transforming growth factor-β (TGF-β),
Smad, vimentin, and connective-tissue growth factor (CTGF) in
HMrSV5 cells following transfection with miR-21 inhibitor [(A)
comparison of the protein expression of ZO-1, TGF-β, Smad, vimentin
and CTGF in each group; (B) comparison of the protein expression of
ZO-1, TGF-β, Smad, vimentin, and CTGF detected by western blot
analysis]. *P<0.05 compared with the blank group. |
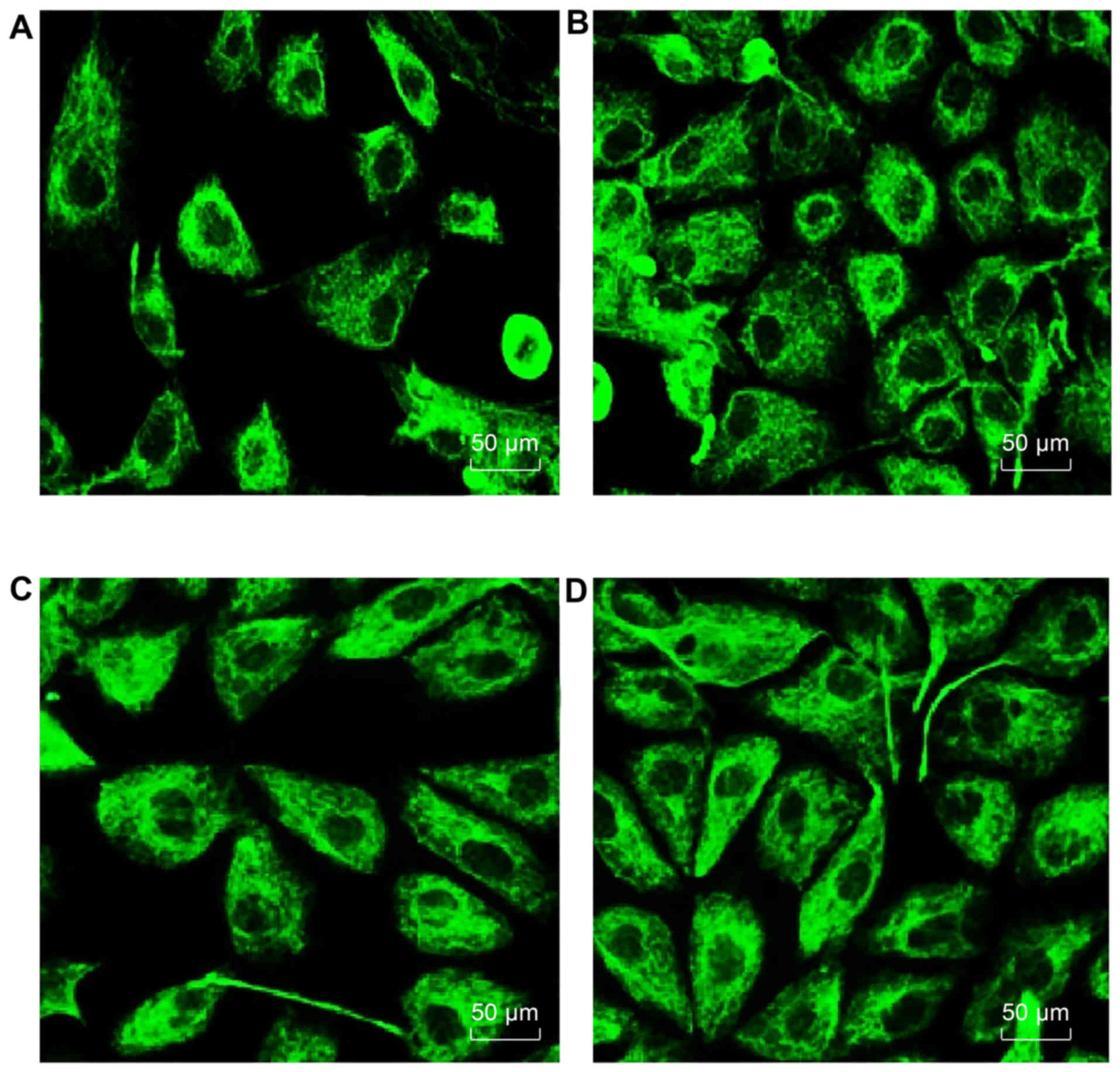
Degree of PF in HMrSV5 cells in each
group
As shown in Fig.
9, with high glucose as an induction source for retroperitoneal
fibrosis, compared with the blank group, the amount of
fibronectin-positive cells increased significantly, with a small
intercellular distance and high density, which was evidence of an
increased degree of fibrosis. By introducing miR-21 inhibitor, in
the high glucose + miR-21 inhibitor group, the number of
fibronectin-positive cells significantly decreased compared to the
high glucose group (P<0.05). However, there was no significant
difference between the high glucose + inhibitor control group and
the high glucose group. Thus, these data suggested that the degree
of PF correlated with miR-21 expression. When miR-21 expression was
increased, the degree of fibrosis was also increased; however, when
miR-21 was inhibited, the degree of fibrosis decreased.
Discussion
In this study, peritoneal mesothelial cells from
patients with PF receiving dialysis and from mice with PF were used
to explore the correlation of miR-21 expression with the TGF-β/Smad
signaling pathway and the role of miR-21 in the progression of PF.
We found that miR-21 enhanced the pathological processes of PF by
regulating the TGF-β/Smad signaling pathway.
PF is the main cause for patients abandoning PD
treatment. A previous study demonstrated that innate activating
fibroblast peritoneal mesothelial cytokines, inflammatory cells and
growth factors play an important role in PF (15). This study found that in
mesothelial cells, miR-21 expression and the corresponding
retroperitoneal fibrosis index, the expression of TGF-β, Smad-3,
vimentin and CTGF were increased with the increasing treatment
duration of dialysis; however, ZO-1 expression was decreased. TGF-β
is an important factor for the promotion of PF and its gene
expression is mainly regulated though the signaling protein
Smad2/3, with the key molecule Smad3 mediated by TGF-β in fibrosis
(16,17). A previous study demonstrated that
TGF-β significantly stimulated Smad2/3 activation in peritoneal
mesothelial cells and upregulated inflammatory cytokines, such as
interleukin (IL)-1, tumor necrosis factor (TNF)-α and intercellular
adhesion molecule 1 (ICAM-1), causing the trans-differentiation of
peritoneal mesothelial cells and the increase of extracellular
matrix (18). Furthermore, high
glucose dialysate can significantly upregulate CTGF and vimentin in
expression peritoneal mesothelial cells, and TGF-β may be used as a
downstream target protein to promote the synthesis and degradation
of the extracellular matrix, resulting in the deposition of
extracellular matrix and mediating the occurrence of PF (19). An in vitro study also found
that miR-21 expression was downregulated by extracellular regulated
protein kinase (ERK) inhibitor, Spry1, and that the expression of
miR-21 had an effect on the promotion of the survival of
fibroblasts and the secretion of fibroblast growth factor via the
ERK signaling pathway; these results indicated that miR-21
influenced the proliferation of fibroblasts, the deposition of
collagen in the extracellular matrix and fibrosis (20).
In this study, in order to further clarify the
association between miR-21 and PF, we constructed a mouse model of
PF, and the expression of miR-21, as well as that of
fibrosis-related proteins was measured. The mRNA and protein
expression of miR-21, TGF-β, Smad-3, vimentin and CTGF exhibited an
upward trend over time, while ZO-1 expression exhibited a downward
trend, and it was confirmed that the degree of fibrosis increased
with the unregulated expression of miR-21. According to previous
reports, TGF-β gene superfamily bound with serine/threonine kinase
receptors can activate R-Smad (Smad-2 and Smad-3) protein for
phosphorylation and cause Smad-7 to competitively bind with TGF-β
receptor through the TGF-β/Smad signaling pathway, which in turn
inhibits Smad-2/3 phosphorylation, and regulates TGF-β signal gene
transcription and expression, thereby regulating cell
proliferation, differentiation, migration and apoptosis (21,22). Studies have confirmed that TGF-β
is a target gene of miR-21 and TGF-β-induced endothelial cell
transition to mesenchymal has been shown to partly occur through
miR-21-mediated signaling pathways (23,24). In studies on renal fibrosis using
humans and animals, TGF-β binding to Smad3 was enhanced, which
increased the transcription of a variety of collagen genes and
miR-21 upregulation was involved in the fibrosis process. miR-21
can regulate the expression of renal extracellular matrix and
smooth muscle actin via TGF-β (25,26). miR-21 may play an important role
in PF by regulating the Smad7 gene; it is an important negative
regulator of proteins in the TGF-β signaling pathway (26). Based on the prediction from
bioinformatics, miR-21 can bind with the Smad7 3′uTR and inhibit
Smad7 expression (27,28).
In conclusion, this study demonstrated that miR-21
may be involved in the progression of PF via regulating the
TGF-β/Smad signaling pathway. Thus, targeting miR-21 may prove to
be an effective therapy for PF. However, as we only investigated
the involvement of miR-21 expression in PF, further studies are
required to explore the potential target gene of miR-21 in PF.
Acknowledgments
We would like to acknowledge the helpful comments
for this study received from our reviewers.
References
|
1
|
Chou CY, Wang SM, Liang CC, Chang CT, Liu
JH, Wang IK, Hsiao LC, Muo CH, Chung CJ and Huang CC: Peritoneal
Dialysis is associated with a netter survival in cirrhotic patients
with chronic kidney disease. Medicine (Baltimore). 95:e24652016.
View Article : Google Scholar
|
|
2
|
Bowen T: A role for fibrocytes in
peritoneal fibrosis? Perit Dial Int. 32:4–6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stavenuiter AW, Farhat K, Vila Cuenca M,
Schilte MN, Keuning ED, Paauw NJ, ter Wee PM, Beelen RH and
Vervloet MG: Protective effects of paricalcitol on peritoneal
remodeling during peritoneal dialysis. BioMed Res Int.
2015:4685742015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
de Lima SM, Otoni A, Sabino AP, Dusse LM,
Gomes KB, Pinto SW, Marinho MA and Rios DR: Inflammation,
neoangiogenesis and fibrosis in peritoneal dialysis. Clin Chim
Acta. 421:46–50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kawanishi K, Honda K, Tsukada M, Oda H and
Nitta K: Neutral solution low in glucose degradation products is
associated with less peritoneal fibrosis and vascular sclerosis in
patients receiving peritoneal dialysis. Perit Dial Int. 33:242–251.
2013. View Article : Google Scholar :
|
|
6
|
Hung KY, Wu KD and Tsai TJ: In vitro study
of peritoneal fibrosis. Perit Dial Int. 27(Suppl 2): S72–S75.
2007.PubMed/NCBI
|
|
7
|
Duan WJ, Yu X, Huang XR, Yu JW and Lan HY:
Opposing roles for Smad2 and Smad3 in peritoneal fibrosis in vivo
and in vitro. Am J Pathol. 184:2275–2284. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Z, Zhang L, He W, Zhu C, Yang J and
Sheng M: Astragalus membranaceus inhibits peritoneal fibrosis via
monocyte chemoattractant protein (MCP)-1 and the transforming
growth factor-β1 (TGF-β1) pathway in rats submitted to peritoneal
dialysis. Int J Mol Sci. 15:12959–12971. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghosh AK, Nagpal V, Covington JW, Michaels
MA and Vaughan DE: Molecular basis of cardiac
endothelial-to-mesenchymal transition (EndMT): Differential
expression of microRNAs during EndMT. Cell Signal. 24:1031–1036.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lan HY: Diverse roles of TGF-β/Smads in
renal fibrosis and inflammation. Int J Biol Sci. 7:1056–1067. 2011.
View Article : Google Scholar :
|
|
11
|
Lan HY: Smads as therapeutic targets for
chronic kidney disease. Kidney Res Clin Pract. 31:4–11. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
van Bommel EF, van Spengler J, van der
Hoven B and Kramer P: Retroperitoneal fibrosis: report of 12 cases
and a review of the literature. Neth J Med. 39:338–345.
1991.PubMed/NCBI
|
|
13
|
Dou XR, Yu XQ, Li XY, Chen WF, Hao WK, Jia
ZJ, Peng WX, Wang X, Yin PD, Wang WJ and Zheng ZH: The role of
TGF-beta1/Smads in the development of peritoneal fibrosis induced
by high glucose peritoneal dialysate and LPS. Zhonghua Yi Xue Za
Zhi. 85:2613–2618. 2005.PubMed/NCBI
|
|
14
|
Lü ZD, Xu HM, Wang HB, Kong B, Li JG, Li
FN and Song YM: Effect of peritoneal fibrosis induced by
transforming growth factor-beta 1 on the adhesion of gastric cancer
cell. Zhonghua Yi Xue Za Zhi. 92:1698–1701. 2012.In Chinese.
|
|
15
|
Vassiliadis E, Oliveira CP,
Alvares-da-Silva MR, Zhang C, Carrilho FJ, Stefano JT, Rabelo F,
Pereira L, Kappel CR, Henriksen K, et al: Circulating levels of
citrullinated and MMP-degraded vimentin (VICM) in liver fibrosis
related pathology. Am J Transl Res. 4:403–414. 2012.PubMed/NCBI
|
|
16
|
Lan HY and Chung AC: TGF-β/Smad signaling
in kidney disease. Semin Nephrol. 32:236–243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhong X, Chung AC, Chen HY, Meng XM and
Lan HY: Smad3-mediated upregulation of miR-21 promotes renal
fibrosis. J Am Soc Nephrol. 22:1668–1681. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Meng XM, Huang XR, Xiao J, Chung AC, Qin
W, Chen HY and Lan HY: Disruption of Smad4 impairs TGF-β/Smad3 and
Smad7 transcriptional regulation during renal inflammation and
fibrosis in vivo and in vitro. Kidney Int. 81:266–279. 2012.
View Article : Google Scholar
|
|
19
|
Kok HM, Falke LL, Goldschmeding R and
Nguyen TQ: Targeting CTGF, EGF and PDGF pathways to prevent
progression of kidney disease. Nat Rev Nephrol. 10:700–711. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thum T, Gross C, Fiedler J, Fischer T,
Kissler S, Bussen M, Galuppo P, Just S, Rottbauer W, Frantz S, et
al: MicroRNA-21 contributes to myocardial disease by stimulating
MAP kinase signalling in fibroblasts. Nature. 456:980–984. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee UE and Friedman SL: Mechanisms of
hepatic fibrogenesis. Best Pract Res Clin Gastroenterol.
25:195–206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mormone E, George J and Nieto N: Molecular
pathogenesis of hepatic fibrosis and current therapeutic
approaches. Chem Biol Interact. 193:225–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang H, Zhang C, Ban T, Liu Y, Mei L,
Piao X, Zhao D, Lu Y, Chu W and Yang B: A novel reciprocal loop
between microRNA-21 and TGFβRIII is involved in cardiac fibrosis.
Int J Biochem Cell Biol. 44:2152–2160. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang T, Zhang L, Shi C, Sun H, Wang J, Li
R, Zou Z, Ran X and Su Y: TGF-β-induced miR-21 negatively regulates
the antiproliferative activity but has no effect on EMT of TGF-β in
HaCaT cells. Int J Biochem Cell Biol. 44:366–376. 2012. View Article : Google Scholar
|
|
25
|
Zhu H, Luo H, Li Y, Zhou Y, Jiang Y, Chai
J, Xiao X, You Y and Zuo X: MicroRNA-21 in scleroderma fibrosis and
its function in TGF-β-regulated fibrosis-related genes expression.
J Clin Immunol. 33:1100–1109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yao Q, Cao S, Li C, Mengesha A, Kong B and
Wei M: Micro-RNA-21 regulates TGF-β-induced myofibroblast
differentiation by targeting PDCD4 in tumor-stroma interaction. Int
J Cancer. 128:1783–1792. 2011. View Article : Google Scholar
|
|
27
|
Butz H, Rácz K, Hunyady L and Patócs A:
Crosstalk between TGF-β signaling and the microRNA machinery.
Trends Pharmacol Sci. 33:382–393. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fu RQ, Hu DP, Hu YB, Hong L, Sun QF and
Ding JG: miR-21 promotes α-SMA and collagen I expression in hepatic
stellate cells via the Smad7 signaling pathway. Mol Med Rep.
16:4327–4333. 2017.PubMed/NCBI
|