Introduction
Postmenopausal osteoporosis (PMO) is a worldwide
health problem (1,2). It is a systemic skeletal disease
characterized by a reduction of bone density and
micro-architectural deterioration, which increases the risk of
fractures in aged women (3,4).
Estrogen serves a principal role in the processes of skeletal
growth and bone homeostasis as well as bone metabolism (5). However, in postmenopausal women, the
deficiency of estrogen results in a reduction of bone mass
(6). Experimental evidence
suggests that in osteoporotic patients, osteoblast activity reduces
whereas osteoclast activity increases, and thus bone resorption
exceeds bone formation (4,7).
Therefore, the focus of cytological experiments on the origin of
PMO has been osteoblast activity and bone resorption processes,
followed by osteoblast genesis and the differentiation potential of
bone marrow mesenchymal stem cells (BMSCs) (8). BMSCs are pluripotent and capable of
self-renewal, supporting hematopoiesis, bone marrow formation and
differentiating into multiple tissues in vitro and in
vivo (9,10). Studies have demonstrated that
BMSCs derived from osteoporotic postmenopausal women exhibit
disturbed intrinsic properties and impaired differentiation ability
compared with normal BMSCs (11).
However, it is unclear whether the BMSCs from a rat model of PMO
are similarly affected.
Stromal cell-derived factor-1α (SDF-1α) and its
cognate receptor CXC chemokine receptor type 4 (CXCR4) have been
reported to be highly active in the reparation and regeneration of
various injured tissues by promoting the migration of stem cells
(12,13). In addition, SDF-1α is considered
by various researchers as a major homing factor for the targeting
of hematopoietic stem cells and mesenchymal stem cells (MSCs), both
of which express CXCR4, to the bone marrow (14–16). Recent studies suggest that the
CXCR4/SDF-1α axis may be important in maintaining the biological
and physiological functions of BMSCs (17,18). Furthermore, the axis may help to
increase the homing efficiency of BMSCs (19). Increasing the concentration of
SDF-1α has been demonstrated to cause a dose-dependent induction of
the chemotactic effects of MSCs (20,21). Other studies have identified an
association between decreased CXCR4 expression in BMSCs and
endothelial progenitor cells and their functions in tissue
regeneration (22,23).
On the basis of these observations, it is
hypothesized that: i) CXCR4 expression in BMSCs isolated from a PMO
rat model (OVX-BMSCs) is reduced; and ii) CXCR4-deficiency affects
the migration ability and osteogenic differentiation of OVX-BMSCs.
To evaluate these hypotheses, the general and CXCR4-related
biological characteristics of OVX-BMSCs and normal rat (Sham-BMSCs)
were examined and compared using in vitro experiments in the
present study. In addition, the chemotaxis of the BMSCs towards
SDF-1α and associated molecular mechanisms were investigated. To
the best of our knowledge, this is the first study to conduct a
comparison of Sham-BMSCs and OVX-BMSCs using a PMO rat model.
Materials and methods
Experimental animals and ethics
statement
A total of 60 female Sprague-Dawley (SD) rats aged 8
weeks and weighing 200±20 g were purchased from the Laboratory
Animal Center of the Fourth Military Medical University (Xi'an,
China). All protocols were approved by the Institutional Animal
Care and Use Committee at the Fourth Military Medical University
and were consistent with the Guidelines of Intramural Animal Use
and Care Committee of the Fourth Military Medical University. The
animals had free access to food and water.
Establishment of the animal models
According to the techniques established by Wronski
et al (24), the rats
underwent ovariectomy (OVX, n=30) or a sham surgery (Sham, n=30),
following the intraperitoneal injection of 2% pentobarbital sodium
(Nembutal; Abbott Laboratories, Lake Bluff, IL, USA) at a dose of
30 mg/kg. Following surgery, the rats were raised with a 12/12-h
light/dark cycle in a temperature-controlled (21–23°C) room with a
relative humidity of 50–60%. All the rats from each group were
sacrificed at 3 months following the surgery. To verify the animal
model, micro-computed tomography (micro CT; Siemens Inveon
Micro-CT; Siemens AG, Munich, Germany) was used to observe the
femoral bone mass changes in the OVX and Sham groups. All femoral
bone samples were totally scanned and 3D images were constructed.
The CT settings were as follows: Voltage, 40 kV; current, 250
µA; voxel size, 10×10 µm; slice thickness, 20
µm; exposure time, 3,000 msec; pixel matrix, 1,024×1,024.
The bone mineral content (BMC), bone mineral density (BMD) and
ratio of bone total volume/trabecular volume (BV/TV) were evaluated
using analytical software (IRW 3.0; Inveon Research Workplace;
Siemens AG). The Max load was tested using a universal mechanical
testing machine (AGS 10 kN; Shimadzu Corporation, Kyoto, Japan).
Osteoporosis in the rats was defined as a BMD of >2.5 standard
deviations (SDs) below the mean value acquired from the sham group
as previously described (25).
Isolation and culture of BMSCs
Rats BMSCs from the Sham and OVX groups were
isolated and cultured according to previously reported methods with
minor modifications (26).
Briefly, tibias and femurs were removed after the rats were
euthanized. After excision of the epiphysis, bone marrow from
bilateral rat tibias or femoral diaphyses was flushed with
α-minimum essential medium (α-MEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.,
Hangzhou, China) and 1% penicillin and streptomycin, and incubated
at 37°C in a humidified atmosphere of 5% CO2. The
culture medium was changed every 2–3 days. When the cells
approached ~80% confluence, adherent cells were treated with a
0.25% trypsin solution and passaged. Cells at passages (P) 2–6 were
used in subsequent experiments. The morphology of the cultured
cells was observed and documented by phase contrast microscopy
(CKX41; Olympus Corporation, Tokyo, Japan).
Colony formation assay
BMSCs (P3) were plated into 9-cm-diameter culture
dishes (Costar) at a density of 1×103 cells/dish and
cultured in α-minimum essential medium (α-MEM; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.) and
1% penicillin and streptomycin (Gibco) prior to the colony
formation assay. The medium was changed every 2 days, and after 14
days, the cells were fixed in 4% paraformaldehyde at 4°C for 30 min
prior to staining with 0.1% crystal violet 25°C (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 10–15 min, and observation
under a stereomicroscope. The number of colony-forming units (CFUs;
≥50 cells) was quantified for statistical analysis.
Cell proliferation assay
BMSCs (P3) were plated into 96-well dishes (Costar)
at a density of 2×103 cells/well and cultured in
complete medium for 1–10 days and the medium was refreshed every 3
days. According to the instructions of the Cell Counting kit-8
(CCK-8; Beyotime Institute of Biotechnology, Shanghai, China),
CCK-8 solution was added to each well at a fixed time of day.
Following incubation for 4 h at 37°C in the dark, the absorbance of
the colored solution was measured using a microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) at a wavelength of
450 nm.
Flow cytometric analysis
The cells were blocked for 1 h in PBS containing 5%
(w/v) non-fat dry milk and incubated at 4°C overnight. The
immunophenotype of the cultured BMSCs (P3) was analyzed using flow
cytometry as previously described (27).
Briefly, following trypsinization and washing twice
with phosphate-buffered saline (PBS) containing 3% FBS, the cells
were transferred into EP tubes (300 µl/tube). Each sample
was then incubated with antibodies (2 µl) against rat
cluster of differentiation (CD)29 (cat. no. 555004; 1:100), CD34
(cat. no. 560518; 1:100), CD43 (cat. no. 553271; 1:100), CD90 (cat.
no. 551401; 1:100) and CD105 (cat. no. 550546; 1:100) (all from BD
Biosciences, San Jose, CA, USA) at 4°C for 1 h in the dark. A tube
without antibodies served as a negative control. Finally, the
samples were washed twice with PBS containing 3% FBS and analyzed
using a Beckman Coulter Epics XL flow cytometer (Beckman Coulter,
Inc., Fullerton, CA, USA). Data were analyzed using Flowjo software
v8.1 (Tree Star, Ashland, OR, USA).
Multiple differentiation analysis
The two types of BMSCs (P3) were plated into 6-well
dishes at a density of 5×105 cells/well and cultured in
complete medium until the cells reached 80% confluence. To assess
the ability of the BMSCs to form mineral nodules, the medium was
changed to osteo-inductive medium [complete medium supplemented
with 50 µg/ml vitamin C, 10 nM dexamethasone and 10 mM
β-glycerophosphate (all Sigma-Aldrich; Merck KGaA)]; the medium was
refreshed every 3 days. Following osteogenic induction for 14 or 21
days, the cells were fixed in 4% paraformaldehyde for 30 min at
room temperature and stained with Alizarin Red S (Sigma-Aldrich;
Merck KGaA) for 12 h at room temperature. Mineralization nodules
were imaged using an inverted microscope. Furthermore, alkaline
phosphatase (ALP) staining was conducted using a BCIP/NBT Alkaline
Phosphatase Color Development kit (Beyotime Institute of
Biotechnology), and ALP enzymatic activity was performed using an
ALP detection kit (Nanjing Jiancheng Bioengineering Institute,
Nanjing, China) according to the manufacturer's protocol. For
adipogenic lineage differentiation, 80% confluent cells were
induced using adipo-inductive medium [complete medium supplemented
with 0.5 mM 3-isobutyl-1-methylxanthine, 1 µM dexamethasone,
0.1 mM indomethacin and 10 µg/ml insulin (all Sigma-Aldrich;
Merck KGaA)]. The medium was refreshed every 3 days. After 7 and 14
days, the cells were fixed in 4% paraformaldehyde for 30 min at
room temperature and stained with Oil Red O (Sigma-Aldrich; Merck
KGaA). Photographic images were captured and the area of staining
was measured using Image-Pro Plus 6.0 software (Media Cybernetics,
Inc., Rockville, MD, USA).
Transwell migration assay
The chemotactic effects of SDF-1α on cell migration
were observed using a Transwell membrane system comprising a
polycarbonate Transwell culture insert with an 8-µm pore
size (Corning Incorporated, Corning, NY, USA) in 24-well plates.
Prior to testing in the migration assay, the cells were starved and
incubated in α-MEM without FBS. To identify a desirable
concentration of SDF-1α (Peprotech, Inc., Rocky Hill, NJ, USA) for
cell migration, different SDF-1α concentrations were used in the
lower chambers of the system (total volume, 600 µl/well).
The medium used for this experiment was α-MEM (supplemented with 5%
FBS) containing 0, 30, 50, 100 or 150 ng/ml SDF-1α. A total of
1×105 BMSCs (P3) were placed in each upper chamber
following resuspension in 150 µl α-MEM containing 5% FBS.
The system was incubated for 6 h and fixed with 4% paraformaldehyde
at 4°C for 20 min. Each Transwell membrane was removed after
stripping to detach non-migrated cells. Finally, the cells that had
migrated to the lower surface of each membrane were stained with
0.2% crystal violet at 25°C (Sigma-Aldrich; Merck KGaA) for 1 h,
and cells in five random fields (×100 magnification) were counted
for statistical analysis. Based on the data obtained from the cell
migration assay using this Transwell membrane system, a
concentration of 100 ng/ ml SDF-1α was selected for all further
experiments. To examine the effect of the presence of SDF-1α on
cell migration, prior to the migration assay, the BMSCs were
incubated in α-MEM without FBS but with a CXCR4 antagonist
(AMD3100; Sigma-Aldrich; Merck KGaA) at a concentration of 10
µg/ml for 2 h.
Immunofluorescence staining
The CXCR4 expression profiles in the two types of
BMSCs at two passages (P3 and P6) were assessed by
immunofluorescence staining. Cells were collected and fixed in 4%
paraformaldehyde at 4°C for 20 min. Following treatment with 0.5%
Triton X-100 and 1% bovine serum albumin (Thermo Fisher Scientific,
Inc.) for 1 h at 25°C, cells were incubated with rabbit anti-CXCR4
antibody (ab124824; 1:100; Abcam, Cambridge, MA, USA) overnight at
4°C. After washing with PBS, cells were incubated with Alexa
Fluor® 488-conjugated anti-CXCR4 antibody (ab208128;
Abcam) for 30 min at 37°C followed by DAPI (Sigma-Aldrich; Merck
KGaA) staining for 2 min, to counterstain the cell nuclei. The
sections were mounted and then observed under a Zeiss LSM 710
confocal microscope (Carl Zeiss AG, Oberkochen, Germany) and the
intensity of the CXCR4-positive area were measured using Image-Pro
Plus 6.0 software. The surface expression of CXCR4 was also
analyzed using flow cytometric analysis as previously described
(22). The cells were incubated
with FITC-conjugated anti-CXCR4 antibody (Abcam).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
After 21 or 14 days of osteogenic/adipogenic
induction, the cells were harvested. The total RNA of the cells was
extracted using TRIzol reagent and complementary DNA (cDNA) was
obtained using SuperScript II reverse transcriptase (both
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Reverse transcription of the total RNA was
performed using a RevertAid First Strand cDNA Synthesis kit (Takara
Bio Inc., Otsu, Japan). qPCR was performed on 10-fold-diluted cDNA
samples in double-distilled water using SYBR-Green Master (Roche
Diagnostics GmbH, Mannheim, Germany) and a CFX Connect™ Real-Time
PCR Detection system (Bio-Rad Laboratories, Inc.). Amplification
was performed under the following conditions: Denaturation at 95̊C
for 3 min, prior to 39 cycles at 95̊C for 15 sec and 60̊C for 30
sec. The primers used for qPCR were: Runt-related transcription
factor 2 (Runx2) forward, 5′-GCC ACC TTC ACT TAC ACC CC-3′ and
reverse, 5′-CGC TGA CGA AGT ACC ATA GTA GAG-3′; osteocalcin (OCN)
forward, 5′-AGA CTC CGG CGC TAC CTC AAC AAT-3′ and reverse, 5′-CAG
CTG TGC CGT CCA TACT-3′; ALP forward, 5′-CTA GTT CCT GGG AGA TGG
TA-3′ and reverse, 5′-GTG TTG TAC GTC TTG GAG AGA-3′; peroxisome
proliferator-activated receptor (PPAR)γ forward, 5′-AGT GGA GAC CGC
CC-3′ and reverse, 5′-GCA GCA GGT TGT CTT-3′; GAPDH forward, 5′-GGA
AAC CCA TCA CCA TCT TC-3′ and reverse, 5′-GCC ATC CAC AGT CTT CTG
AGT-3′. The mRNA expression levels of the target genes were
compared following normalization with the internal control GAPDH
based on the 2−ΔΔCq method (28). In addition, the total RNA of the
two groups of BMSCs at different passages (P3 and P6) was extracted
for relative gene expression analysis by RT-qPCR based on the
2−ΔΔCq method. The target genes and primer sequences
were as follows: CXCR4 forward, 5′-CCT CCT CCT GAC TAT CCC TGA-3′
and reverse, 5′-CGA ACT CAC ATC CTT GCT TG-3′; GAPDH forward,
5′-GGA AAC CCA TCA CCA TCT TC-3′ and reverse, 5′-GCC ATC CAC AGT
CTT CTG AGT-3′. GAPDH served as an internal reference.
Protein isolation and western blot
analysis
Western blot analysis was performed as previously
described (22). Briefly, the two
types of BMSCs at P3 and P6 were lysed using lysis buffer
(Sigma-Aldrich; Merck KGaA) on ice following a standard procedure
and then centrifuged at 16096 × g for 10 min at 4̊C to collect the
total cell protein. The cell protein concentrations were determined
using a BCA Protein Assay kit (Beyotime Institute of Biotechnology)
according to the manufacturer's protocol. The proteins were
subsequently denatured by boiling and resolved using 12% SDS-PAGE
and transferred onto polyvinylidene difluoride membranes by
electrophoretic separation. The membranes were blocked with 5%
skimmed milk in TBS-T buffer (25 mm Tris-HCl, pH 8.00, 125 mm NaCl
and 0.1% Tween-20) for 1 h at room temperature. The membranes were
then incubated overnight with primary antibodies at 4̊C. The
primary antibodies included anti-CXCR4 (ab124824; 1:500; Abcam) and
anti-GAPDH (ab8245; 1:1000; Abcam), anti-AKT (cat. no. 9272; 1:500;
Cell Signaling Technology, Danvers, MA, USA) and anti-p-AKT (cat.
no. 9271; 1:500; Cell Signaling Technology). The membranes were
washed three times with TBS-T to remove the excess antibodies prior
to incubation with anti-rabbit secondary antibodies (A0192;
1:8,000; Beyotime Institute of Biotechnology) conjugated with
horseradish peroxidase. The proteins were detected by enhanced
chemiluminescence (Sigma-Aldrich; Merck KGaA). GAPDH was used as an
internal control to normalize the samples. To investigate the
signal transduction mechanisms involved in SDF-1α-induced
chemotactic activity, total AKT (t-AKT) and phosphorylated AKT
(p-AKT) protein levels were analyzed using western blot analysis in
the two sets of BMSCs. Prior to protein isolation and western blot
analysis, the two sets of BMSCs were treated with 100 ng/ml SDF-1α
for 5, 15, 30, 60 or 120 min. To further examine the effects of the
presence of SDF-1α/CXCR4 on p-AKT, prior to the treatment with
SDF-1α, the BMSCs were incubated in α-MEM without FBS but with
CXCR4 antagonist at a concentration of 10 µg/ml for 2 h. The
two sets of BMSCs were then treated with 100 ng/ml SDF-1α for 5,
15, 30 or 60 min.
Statistical analysis
Results are presented as the mean ± SD and all
experiments were repeated three times. Data were analyzed by
one-way analysis of variance (ANOVA) followed by Tukey's post hoc
test using SPSS 18.0 software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference for all tests.
Results
Cell culture and general
characterization
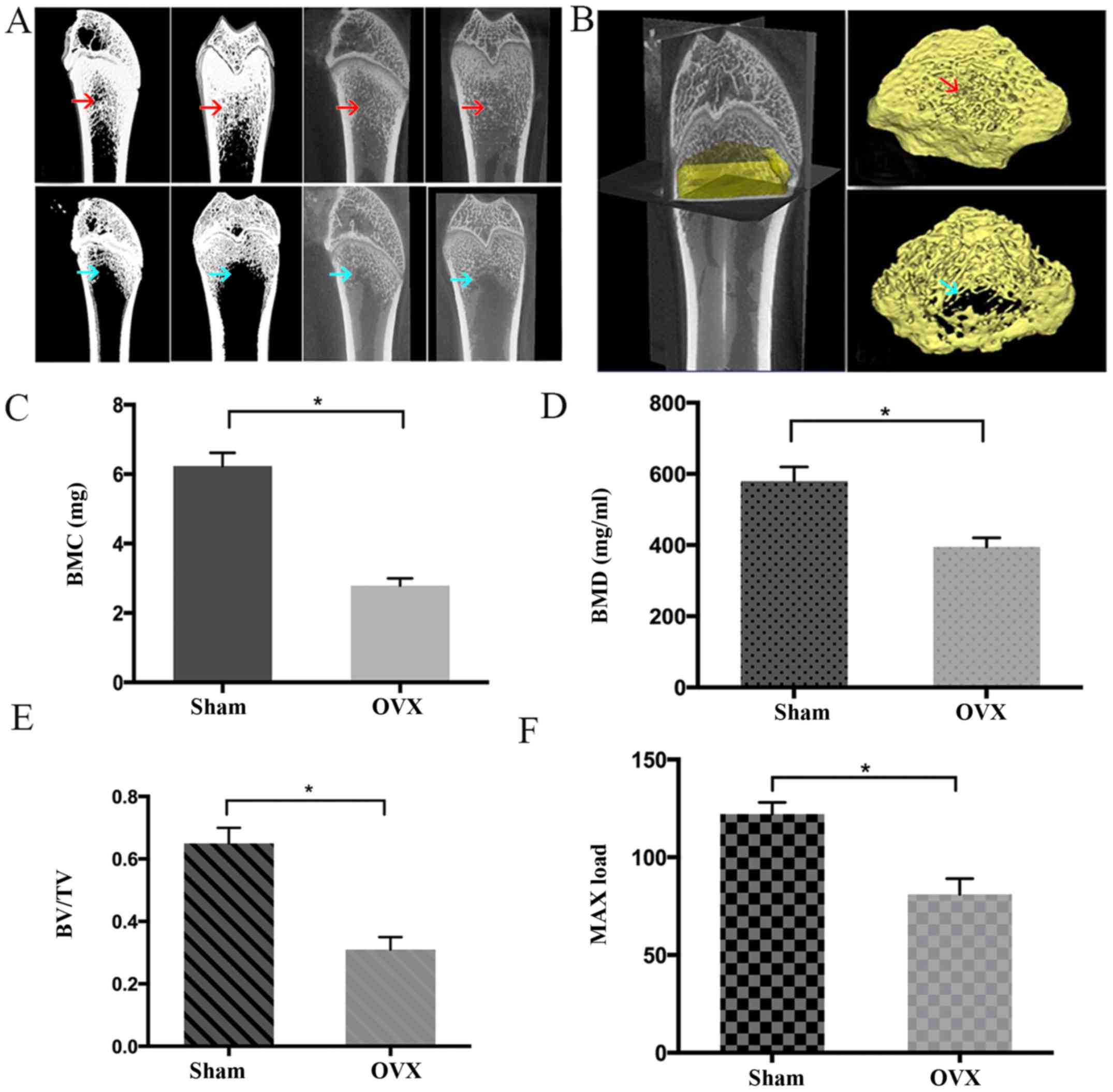
As shown in Fig.
1, the BMC, BMD, BV/TV and Max load in the OVX group were
significantly lower compared with those in the Sham group
(P<0.05), indicating that the rats in OVX-group had poorer bone
tissue-forming ability compared with those in the Sham group. In
addition, the rats in the OVX group had a BMD of >2.5 SDs below
the mean BMD of the Sham group, suggesting the PMO rat model was
successfully established (25).
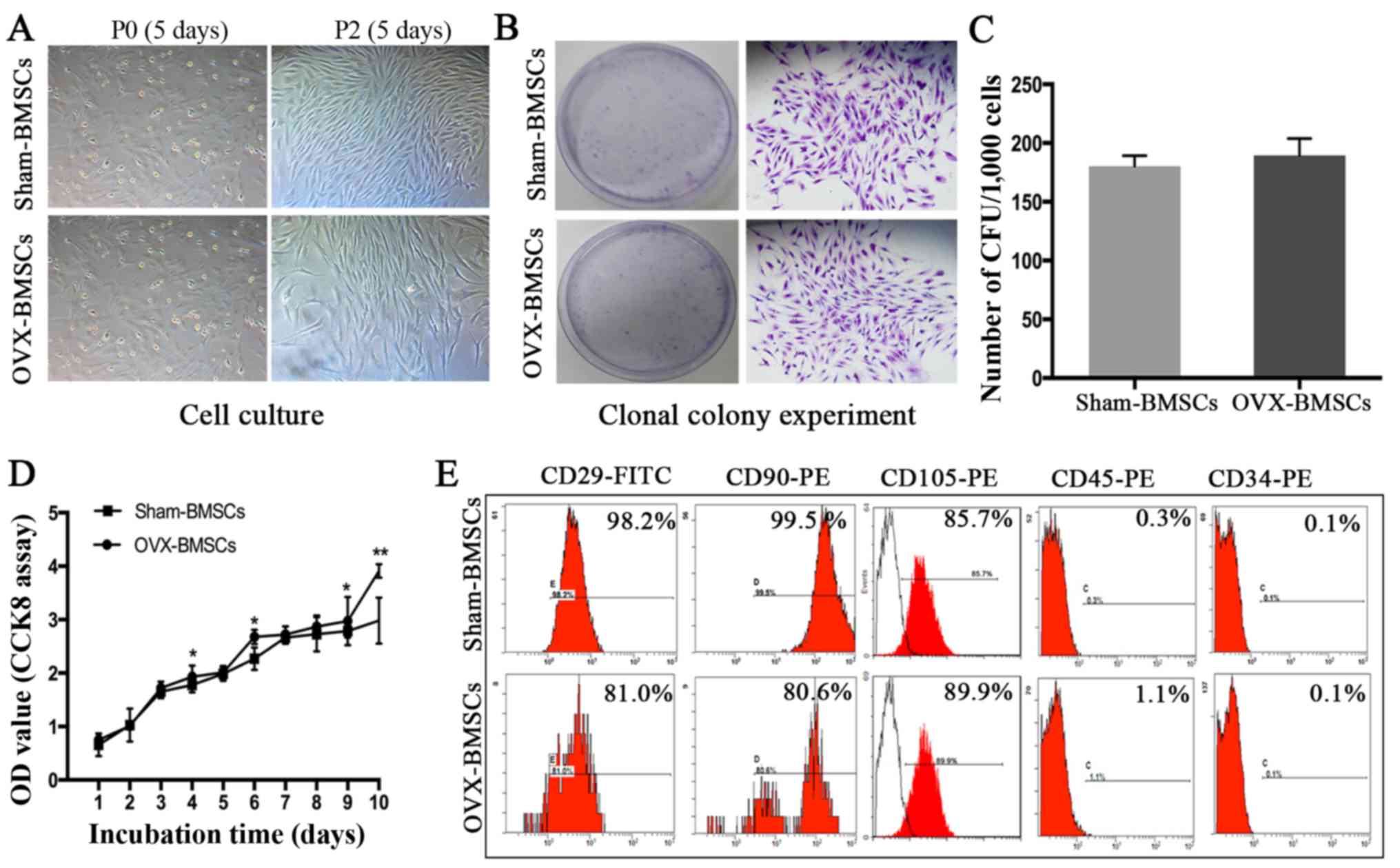
It was observed that BMSCs at P0 adhered to the culture plates and
had a mixture of round and spindle-like morphology, whereas cells
at P2 exhibited normal growth and a typical spindle-like appearance
(Fig. 2A). However, no marked
difference was observed between the BMSCs from the Sham and OVX
groups at each time point. In addition, no significant difference
in colony-forming ability was detected between the two sets of
cells (Fig. 2B and C). However,
OVX-BMSCs exhibited a stronger proliferative ability than
Sham-BMSCs on days 4, 6, 9 and 10- (P<0.05; Fig. 2D). The results of the
immunophenotypic analysis revealed that the two sets of BMSCs
exhibited similar patterns of surface molecule expression;
specifically, they were positive for MSC-specific surface markers
(CD29, CD90 and CD105) and negative for hematopoietic cell markers
(CD34 and CD45; Fig. 2E).
Differentiation potential analysis
The multiple differentiation potentials of the two
groups of BMSCs were determined using osteogenic and adipogenic
differentiation assays (Fig. 3).
The osteogenic ability was indicated by the area of Alizarin Red S
staining (for mineralized nodules) following osteogenic induction
(Fig. 3A–C). Quantitative
analysis revealed that the Sham-BMSCs produced a larger area of
mineralized nodules than the OVX-BMSCs at 14 and 21 days
(P<0.05). Subsequently, ALP was detected using an ALP color
development kit. Consistent with the observed Alizarin Red S
staining results on days 7 and 14, the ALP staining results
indicated that OVX-BMSCs displayed less osteogenic differentiation
than Sham-BMSCs (Fig. 3D and E).
In order to observe the dynamic changes of ALP in the two cell
types, ALP activity was detected. In accordance with the above
osteogenic induction results, OVX-BMSCs exhibited reduced ALP
activity, indicative of impaired osteogenic ability, at all time
points, although there was only a statistically significant
difference on days 6, 8, 12 and 14 (P<0.05; Fig. 3F). Such a trend was further
confirmed by the results of RT-qPCR. Following osteogenic
induction, it was detected that the expression levels of osteogenic
marker genes (Runx2, OCN and ALP) in the Sham-BMSCs group were
significantly higher than those in the OVX-BMSCs group (P<0.05;
Fig. 3J). Following adipogenic
induction, Oil Red O-positive lipid droplets were observed in the
two cell lines, indicating that the two types of BMSCs possessed
adipogenic differentiation potential. However, the OVX-BMSCs
exhibited stronger adipogenic activity with a significantly larger
area of lipid droplets following Oil Red O staining on day 14
(P<0.05; Fig. 3G–I). Likewise,
OVX-BMSCs had a higher mRNA level of the adipogenic
differentiation-associated gene PPARγ compared with the Sham-BMSCs
(P<0.05; Fig. 3K).
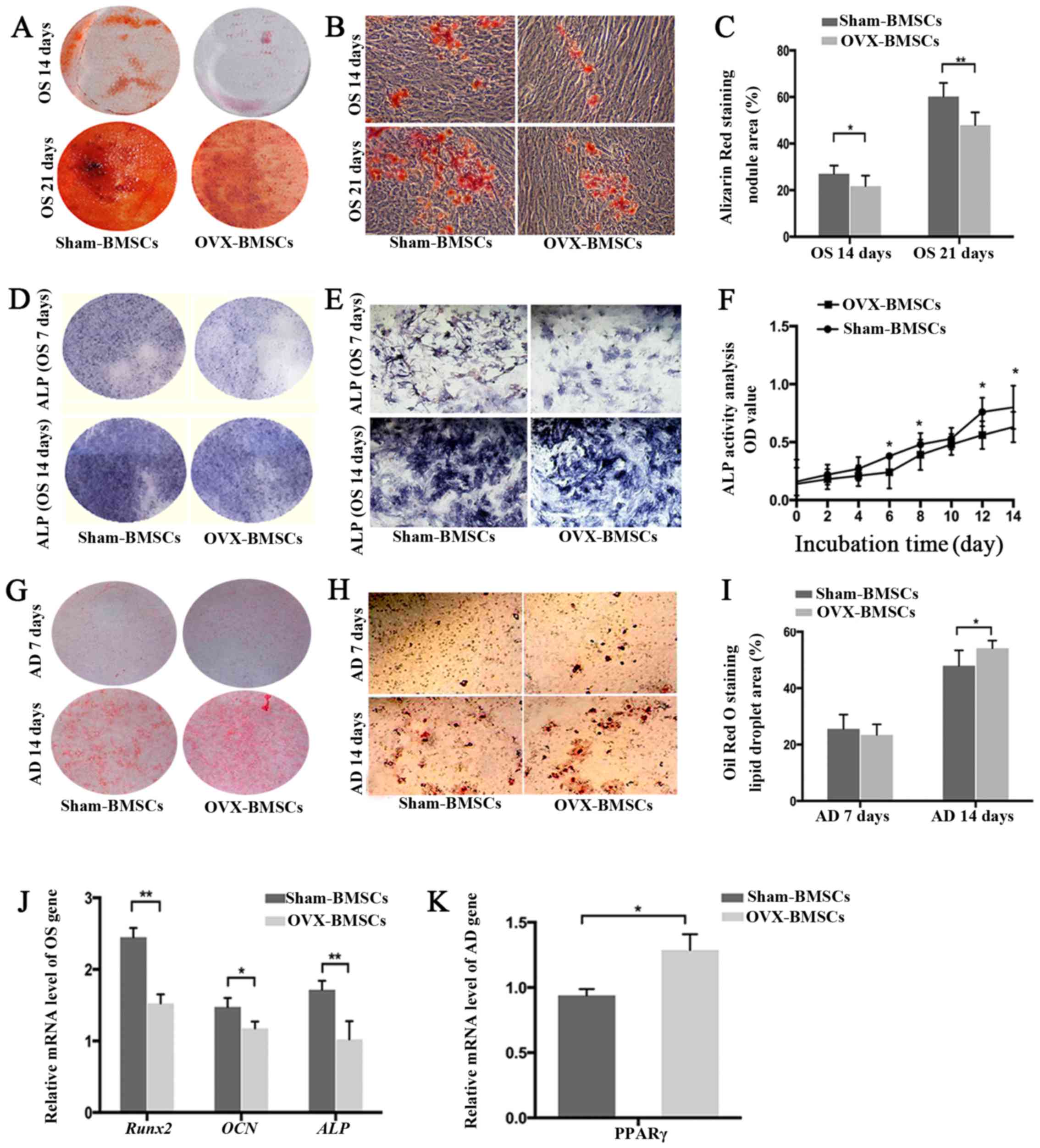 | Figure 3Differentiation potential of
Sham-BMSCs and OVX-BMSCs. (A and B) Representative images of
Alizarin Red S-stained mineral deposits following 14 or 21 days of
osteogenic induction (A, general view; B, ×40 magnification) and
(C) quantification of the area of dye absorption for Alizarin Red S
in the Sham-BMSCs and OVX-BMSCs. (D and E) Representative images of
ALP staining following 7 or 14 days of osteogenic induction (D,
general view; E, ×200 magnification) and (F) quantification of ALP
activity as different time points in the Sham-BMSCs and OVX-BMSCs.
(G and H) Representative images of Oil Red O-stained lipid
inclusions following 7 and 14 days of adipogenic induction (G,
general view; H, ×200 magnification) and (I) quantification of the
area of dye absorption for Oil Red O in Sham-BMSCs and OVX-BMSCs.
(J) Reverse transcription-quantitative polymerase chain reaction
measurements of OCN, Runx2 and ALP genes following osteogenic
induction. The values are ratios relative to GAPDH (n=4). (K)
Relative levels of expression of the PPARγ gene following 14 days
of osteogenic induction (n=4). Data are presented as the mean ±
standard deviation. *P<0.05 and
**P<0.01 for Sham-BMSCs vs. OVX-BMSCs. BMSCs, bone
marrow mesenchymal stem cells; OVX, ovariectomy; ALP, alkaline
phosphatase; OS, osteogenic; AD, adipogenic; RunX2, Runt-related
transcription factor 2; OCN, osteocalcin; ALP, alkaline
phosphatase;PPAR, peroxisome proliferator-activated receptor. |
Chemotactic activity assay
The results of the in vitro cell chemotaxis
and chemotaxis-blocking assays demonstrated that the migration of
BMSCs was dependent upon the concentration of SDF-1α, with a peak
migration rate at 100 ng/ml in the two sets of BMSCs. Therefore, a
concentration of 100 ng/ml SDF-1α was selected for all further
experiments. It was observed that the presence of the CXCR4
antagonist AMD3100 greatly decreased the migration activity of
Sham-BMSCs and OVX-BMSCs to nearly half of that for the respective
BMSCs treated with SDF-1α without antagonist. In addition, the
OVX-BMSCs exhibited significantly impaired chemotactic activity
compared with the Sham-BMSCs (P<0.05; Fig. 4A), with the exception of when 100
ng/ml SDF-1α plus AMD3100 was used.
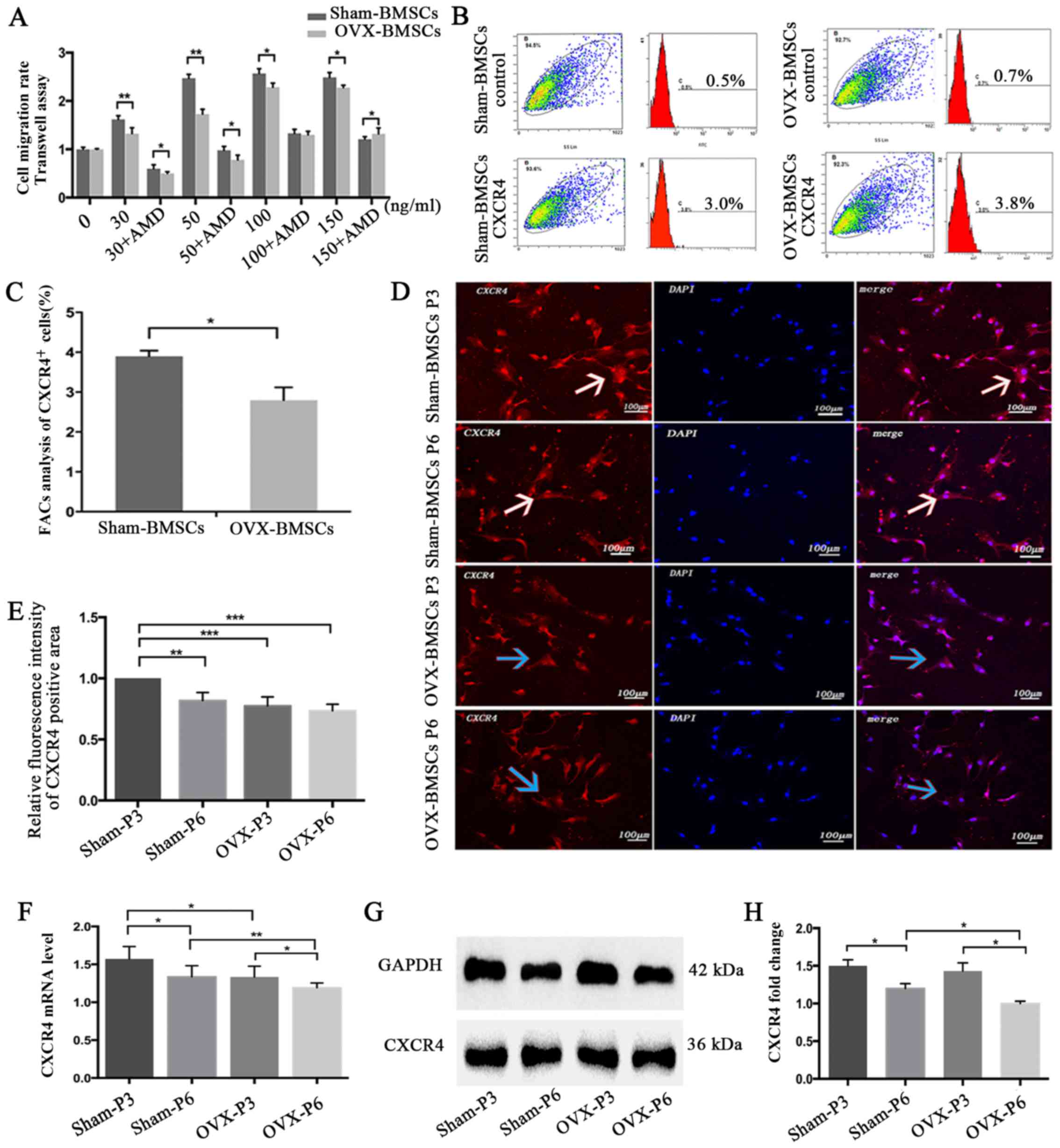 | Figure 4Chemotaxis assay and CXCR4 expression
analysis of Sham-BMSCs and OVX-BMSCs. (A) Quantification of the
chemotactic activity of Sham-BMSCs and OVX-BMSCs towards SDF-1α in
the absence or presence of AMD3100. Culture medium with different
concentrations of SDF-1α (30, 50, 100 or 150 ng/ml) was applied to
the lower chamber of the Transwell system. (B) Representative
images showing the detection of surface CXCR4 expression by flow
cytometry in Sham-BMSCs and OVX-BMSCs (n=5) and (C) data analysis
of the percentage of CXCR4-positive cells. (D) Representative
immunofluorescent images of CXCR4 staining on Sham-BMSC and
OVX-BMSC (P3 and P6) membranes (×200 magnification) and (E)
quantitative analysis of CXCR4 fluorescence intensity. White arrows
indicate higher expression of CXCR4, whereas blue arrows indicate
lower expression of CXCR4 with weaker red fluorescence. (F) CXCR4
expression was analyzed by reverse transcription-quantitative
polymerase chain reaction and (G and H) western blot analysis in
Sham-BMSCs and OVX-BMSCs (P3 and P6). (G) Representative blots and
(H) quantified data, presented as the mean ± standard deviation.
*P<0.05 between the groups as indicated. CXCR4, CXC
chemokine receptor type 4; BMSCs, bone marrow mesenchymal stem
cells; OVX, ovariectomy; SDF-1α, stromal cell-derived factor-1α;
P3, passage 3; P6, passage 6. |
Temporal changes in CXCR4 expression
The expression of CXCR4 on the cell surface was
determined using flow cytometric analysis. The results revealed
that the Sham-BMSCs exhibited significantly higher CXCR4 expression
compared with the OVX-BMSCs (P<0.05; Fig. 4B and C). To compare the expression
of CXCR4 in the two sets of BMSCs, the expression of CXCR4 was
detected using an immunofluorescence, the CXCR4 mRNA level was
analyzed by RT-qPCR and the CXCR4 protein level was analyzed using
western blot analysis. At the same cell passage, the CXCR4
immunofluorescence, mRNA and protein level in the Sham-BMSCs were
much higher than those in the OVX-BMSCs, though no statistical
significance for CXCR4 immunofluorescence was noticed at P6
(Fig. 4D–H). In addition, the
results indicated that cell passage was likely to adversely affect
the expression of CXCR4, since the Sham-BMSCs and OVX-BMSCs
exhibited higher CXCR4 immunofluorescence, mRNA and protein levels
at P3 than at P6 (Fig. 4D–H).
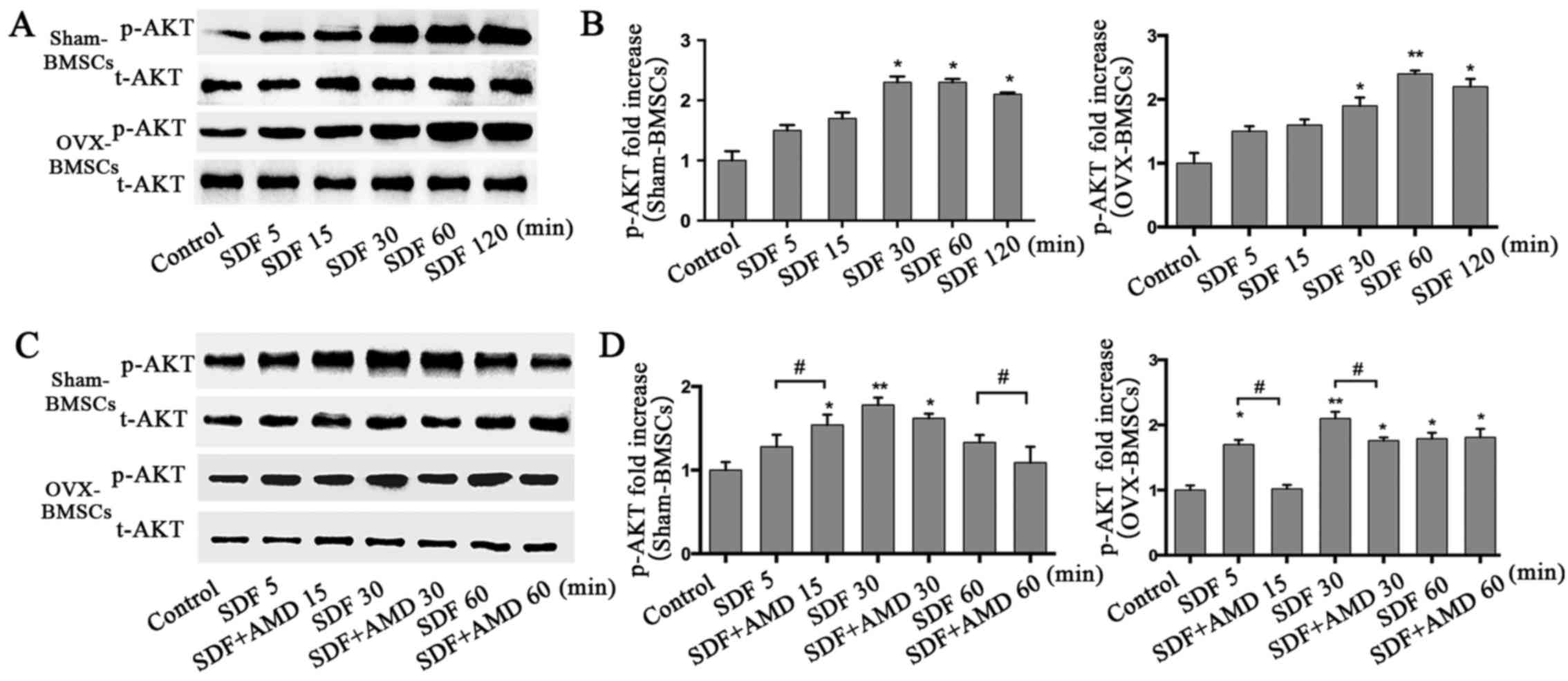
p-AKT analysis
To further investigate the signal transduction
mechanisms involved in the SDF-1α-induced chemotactic activity of
BMSCs from the PMO rat model, t-AKT and p-AKT protein levels were
analyzed at different time points as the duration of stimulation
increased using western blot analysis in the two sets of BMSCs
(Fig. 5). SDF-1α significantly
increased the protein levels of p-AKT at 30 min, which then started
to decline at 120 min in the Sham-BMSCs. However, for OVX-BMSCs,
the level of p-AKT peaked at 60 min and then declined at 120 min
(Fig. 5A and B). As shown in
Fig. 5C and D, BMSCs pretreated
with AMD3100 had reduced levels of p-AKT compared with those that
were not pretreated, but exhibited a similar trend.
Discussion
Osteoporosis is a common condition in postmenopausal
women, which predisposes individuals to an increased risk of bone
fragility and bone fractures. It is considered that in individuals
with osteoporosis, bone loss may be attributed to changes in the
osteogenic differentiation and adipocyte ability of BMSCs (11,29). In the present study, the general
biological characteristics of Sham-BMSCs and OVX-BMSCs were
compared. No marked difference between the two groups of BMSCs was
detected in terms of cell culture and the expression of stem cell
surface markers; however, OVX-BMSCs exhibited a greater ability to
proliferate. Furthermore, osteoporosis was indicated to impair
osteogenic ability since a smaller area of mineralized nodules and
lower ALP activity, as well as lower expression levels of
osteogenic marker genes (Runx2, OCN and ALP) were observed for
OVX-BMSCs compared with Sham-BMSCs. These results are in accordance
with those of Wu et al (29) and Yang et al (30) who reported that ALP activity was
more active in Sham-BMSCs than in OVX-BMSCs (29,30). However, the OVX-BMSCs exhibited
stronger adipogenic activity than Sham-BMSCs in the present study,
since a larger lipid droplet area with Oil Red O staining as well
as a higher expression level of the adipogenic
differentiation-associated gene PPARγ was observed. These results
are consistent with those in the studies described by Pino et
al (11). In the pathogenesis
of osteoporosis, the impairment of the functionality and
microenvironment of MSCs results in the dysregulation of bone turn
over, such that bone resorption exceeds bone formation (11). The OVX-BMSCs exhibited intrinsic
functional alterations, leading to poor osteogenic capability and
increased adipogenesis. The osteoporotic bone marrow
microenvironment differs from the normal microenvironment in that
it has increased concentrations of pro-adipogenic and
pro-inflammatory regulatory factors (31). The content and/or quality of
adipocytes in the bone marrow appears critical to the impairment of
BMSCs; thus, osteoporosis may be comparable with other
age-associated diseases, such as obesity, atherogenesis and
diabetes, which are characterized by unbalanced extramedullary
adipocyte formation and signaling (11,32,33).
The SDF-1α/CXCR4 axis is considered to be the key
pathway influencing the homing capacity of BMSCs. It has been
demonstrated that the augmentation of SDF-1α at the injury site or
transplantation of MSCs that overexpress CXCR4 may improve outcomes
(7,34). However, to the best of our
knowledge, whether osteoporosis affects the chemotaxis of BMSCs
towards SDF-1α has not been elucidated. In the present study, it
was observed that SDF-1α concentration-dependently increased the
migration of OVX-BMSCs and Sham-BMSCs within a limited
concentration range, whereas such migration was inhibited by
AMD3100, an antagonist of CXCR4. Notably, Sham-BMSCs exhibited
greater migration than OVX-BMSCs at nearly all tested
concentrations of SDF-1α, regardless of AMD3100, suggesting that
osteoporosis negatively affected SDF-1α-directed BMSC migration.
The surface expression level of CXCR4 on BMSCs has been indicated
to determine their efficiency of homing and the regeneration
response to target tissues (35,36). Wynn et al (37) found that the proportion human MSCs
with expression of the CXCR4 receptor on the cell membrane ranged
from 1 to 3.9%. Similarly, the flow cytometric analysis in the
present study demonstrated that the proportion of BMSCs positive
for the expression of CXCR4 in the Sham- and OVX-BMSCs was no more
than 4%. However, the percentage of CXCR4-positive cells was higher
in the Sham-BMSCs group compared with the OVX-BMSCs group. To
further investigate the possible mechanism underlying the reduced
migration of OVX-BMSCs towards SDF-1α, CXCR4 expression, mRNA
levels and protein levels were analyzed. The CXCR4
immunofluorescence intensity, and mRNA and protein expression
levels in the Sham-BMSCs were higher than those in the OVX-BMSCs.
The lower expression of CXCR4 in the OVX-BMSCs may partly explain
their lower chemotactic migration ability. By contrast, previous
studies have found that in certain acute disease models, such as
rat abdominal aortic aneurysm model (38) and severe acute pancreatitis model
(39), the mRNA and protein
levels of CXCR4 in BMSCs were much higher than those in the BMSCs
from the normal animals. These phenomena may result from an
increase in the number of BMSCs expressing CXCR4 as well as
promotion of the autocrine and paracrine activity of SDF-1α in
wounds or tissues affected by acute inflammation. Signaling by
SDF-1α stimulates the expression of CXCR4 on the cell surface and
promotes the homing of BMSCs towards defects; however, as soon as
the cells reach their destination, the expression of CXCR4 is
reduced to a low level (40). In
order to further investigate the expression of CXCR4 in BMSCs, cell
passage was investigated as an influential factor in the present
study, since BMSCs have highly multi-directional differentiation
potential during culture and cell passage. In the present study, a
decline in CXCR4 mRNA and protein expression was detected when the
Sham-BMSCs and OVX-BMSCs were cultured from P3 to P6. It has been
demonstrated that the expression of CXCR4 in normal human BMSCs can
be greatly attenuated during cell passage (41,42). However, to the best of our
knowledge, the present study is the first to demonstrate this in
rat Sham-BMSCs and OVX-BMSCs. Therefore, this observation combined
with the recommendations of previous researchers who identified
that cells at an early passage exhibit better stability, growth
rates and more potent anti-inflammatory activity, indicates that
BMSCs of passage 2–4 are likely to be the most effective for the
repair of tissue defects (41,43,44).
The AKT signaling pathway is known to mediate the
migration of cells induced by cytokines such as SDF-1α in various
types of cells (45–47). Therefore, the protein levels of
t-AKT and p-AKT were analyzed by western blot analysis in the
present study. SDF-1α (100 ng/ml) induced time-dependent increases
of p-AKT protein levels in Sham-BMSCs and OVX-BMSCs (46,48). These results concur with those of
other studies, which indicated that the AKT signaling pathway is
involved in the induction of BMSC migration by SDF-1α. The
activation of AKT has been suggested to serve multiple functions in
the process of BMSC migration. For example, it may increase the
secretion of actin and myosin, which are considered to be
associated with BMSC movement (49). In addition, it may contribute to
the secretion of matrix metalloproteinase to promote BMSC migration
via degradation of the extracellular matrix and basilar membrane
(50). The results of the present
study revealed that the time point at which the p-AKT levels peaked
in the two types of BMSCs was significantly different, and was
later in the OVX-BMSCs. These results are in accordance with those
of Shahzad et al (51).
The weaker sensitivity of p-AKT in OVX-BMSCs may be attributed to
their lower expression of CXCR4 (52). The cells pretreated with AMD3100
exhibited lower protein levels of p-AKT than the cells that were
not pretreated. These results are in accordance with dose-dependent
increase in the migration of the BMSCs induced by SDF-1α and the
inhibition of the migration by AMD3100, suggesting that the reduced
chemotaxis towards SDF-1α of OVX-BMSCs may be associated with
reduced sensitivity of p-AKT.
In conclusion, the general and SDF-1α/CXCR4-related
biological characteristics of OVX-BMSCs and Sham-BMSCs were
investigated and compared, and their molecular mechanisms were
evaluated. Compared with Sham-BMSCs, the OVX-BMSCs exhibited
increased proliferation ability and impaired osteogenic ability as
well as lower chemotactic activity towards SDF-1α, which may be
partially associated with lower expression of CXCR4 and restricted
AKT phosphorylation.
Abbreviations:
|
BMSCs
|
bone marrow mesenchymal stem cells
|
|
PMO
|
postmenopausal osteoporosis
|
|
SDF-1
|
stromal cell-derived factor-1
|
|
CXCR4
|
CXC chemokine receptor type 4
|
|
PBS
|
phosphate-buffered saline
|
|
α-MEM
|
α-minimum essential medium
|
|
FBS
|
fetal bovine serum
|
|
micro-CT
|
micro-computed tomography
|
|
CFUs
|
colony-forming units
|
References
|
1
|
Baccaro LF, Conde DM, Costa-Paiva L and
Pinto-Neto AM: The epidemiology and management of postmenopausal
osteoporosis: A viewpoint from Brazil. Clin Interv Aging.
10:583–591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Diab DL and Watts NB: Postmenopausal
osteoporosis. Curr Opin Endocrinol Diabetes Obes. 20:501–509. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shang F, Ming L, Zhou Z, Yu Y, Sun J, Ding
Y and Jin Y: The effect of licochalcone A on cell-aggregates ECM
secretion and osteogenic differentiation during bone formation in
metaphyseal defects in ovariectomized rats. Biomaterials.
35:2789–2797. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
No authors listed: NIH Consensus
Development Panel on Osteoporosis Prevention, Diagnosis, and
Therapy, March 7–29, 2000: highlights of the conference. South Med
J. 94:569–573. 2001.
|
|
5
|
Riggs BL, Khosla S and Melton LJ III: Sex
steroids and the construction and conservation of the adult
skeleton. Endocr Rev. 23:279–302. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pacifici R: Cytokines, estrogen, and
postmenopausal osteoporosis - the second decade. Endocrinology.
139:2659–2661. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Anbinder AL, Moraes RM, Lima GM, Oliveira
FE, Campos DR, Rossoni RD, Oliveira LD, Junqueira JC, Ma Y and
Elefteriou F: Periodontal disease exacerbates systemic
ovariectomy-induced bone loss in mice. Bone. 83:241–247. 2016.
View Article : Google Scholar
|
|
8
|
Shoback D: Update in osteoporosis and
metabolic bone disorders. J Clin Endocrinol Metab. 92:747–753.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiao Y, Wang Y, Li L, Li YH, Pang Y, Song
JY and Jiang ZJ: Homing of chloromethylbenzoyl ammonia-labeled bone
marrow mesenchymal stem cells in an immune-mediated bone marrow
failure mouse model in vivo. Genet Mol Res. 13:11–21. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yagi H, Soto-Gutierrez A, Parekkadan B,
Kitagawa Y, Tompkins RG, Kobayashi N and Yarmush ML: Mesenchymal
stem cells: mechanisms of immunomodulation and homing. Cell
Transplant. 19:667–679. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pino AM, Rosen CJ and Rodríguez JP: In
osteoporosis, differentiation of mesenchymal stem cells (MSCs)
improves bone marrow adipogenesis. Biol Res. 45:279–287. 2012.
View Article : Google Scholar
|
|
12
|
Lau TT and Wang DA: Stromal cell-derived
factor-1 (SDF-1): homing factor for engineered regenerative
medicine. Expert Opin Biol Ther. 11:189–197. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Orimo A, Gupta PB, Sgroi DC,
Arenzana-Seisdedos F, Delaunay T, Naeem R, Carey VJ, Richardson AL
and Weinberg RA: Stromal fibroblasts present in invasive human
breast carcinomas promote tumor growth and angiogenesis through
elevated SDF-1/CXCL12 secretion. Cell. 121:335–348. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bleul CC, Fuhlbrigge RC, Casasnovas JM,
Aiuti A and Springer TA: A highly efficacious lymphocyte
chemoattractant, stromal cell-derived factor 1 (SDF-1). J Exp Med.
184:1101–1109. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gupta SK, Lysko PG, Pillarisetti K,
Ohlstein E and Stadel JM: Chemokine receptors in human endothelial
cells. Functional expression of CXCR4 and its transcriptional
regulation by inflammatory cytokines. J Biol Chem. 273:4282–4287.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ji JF, He BP, Dheen ST and Tay SS:
Interactions of chemokines and chemokine receptors mediate the
migration of mesenchymal stem cells to the impaired site in the
brain after hypoglossal nerve injury. Stem Cells. 22:415–427. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Herberg S, Fulzele S, Yang N, Shi X, Hess
M, Periyasamy-Thandavan S, Hamrick MW, Isales CM and Hill WD:
Stromal cell-derived factor-1β potentiates bone morphogenetic
protein-2-stimulated osteoinduction of genetically engineered bone
marrow-derived mesenchymal stem cells in vitro. Tissue Eng Part.
19:1–13. 2013. View Article : Google Scholar
|
|
18
|
Herberg S, Kondrikova G,
Periyasamy-Thandavan S, Howie RN, Elsalanty ME, Weiss L, Campbell
P, Hill WD and Cray JJ: Inkjet-based biopatterning of SDF-1β
augments BMP2-induced repair of critical size calvarial bone
defects in mice. Bone. 67:95–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang D, Sun S, Wang Z, Zhu P, Yang Z and
Zhang B: Stromal cell-derived factor-1 receptor
CXCR4-overexpressing bone marrow mesenchymal stem cells accelerate
wound healing by migrating into skin injury areas. Cell Reprogram.
15:206–215. 2013.PubMed/NCBI
|
|
20
|
Kitaori T, Ito H, Schwarz EM, Tsutsumi R,
Yoshitomi H, Oishi S, Nakano M, Fujii N, Nagasawa T and Nakamura T:
Stromal cell-derived factor 1/CXCR4 signaling is critical for the
recruitment of mesenchymal stem cells to the fracture site during
skeletal repair in a mouse model. Arthritis Rheum. 60:813–823.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lapidot T, Dar A and Kollet O: How do stem
cells find their way home. Blood. 106:1901–1910. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shao H, Xu Q, Wu Q, Ma Q, Salgueiro L,
Wang J, Eton D, Webster KA and Yu H: Defective CXCR4 expression in
aged bone marrow cells impairs vascular regeneration. J Cell Mol
Med. 15:2046–2056. 2011. View Article : Google Scholar :
|
|
23
|
Guang LG, Boskey AL and Zhu W: Age-related
CXC chemokine receptor-4-deficiency impairs osteogenic
differentiation potency of mouse bone marrow mesenchymal stromal
stem cells. Int J Biochem Cell Biol. 45:1813–1820. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wronski TJ, Lowry PL, Walsh CC and
Ignaszewski LA: Skeletal alterations in ovariectomized rats. Calcif
Tissue Int. 37:324–328. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Namkung-Matthai H, Appleyard R, Jansen J,
Hao Lin J, Maastricht S, Swain M, Mason RS, Murrell GA, Diwan AD
and Diamond T: Osteoporosis influences the early period of fracture
healing in a rat osteoporotic model. Bone. 28:80–86. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lennon DP and Caplan AI: Isolation of rat
marrow-derived mesenchymal stem cells. Exp Hematol. 34:1606–1607.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wen L, Wang Y, Wen N, Yuan G, Wen M, Zhang
L, Liu Q, Liang Y, Cai C, Chen X, et al: Role of endothelial
progenitor cells in maintaining stemness and enhancing
differentiation of mesenchymal stem cells by indirect cell-cell
interaction. Stem Cells Dev. 25:123–138. 2016. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
29
|
Wu Y, Zhang P, Dai Q, Yang X, Fu R, Jiang
L and Fang B: Effect of mechanical stretch on the proliferation and
differentiation of BMSCs from ovariectomized rats. Mol Cell
Biochem. 382:273–282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang Z, Huang JH, Liu SF, Zhao YJ, Shen
ZY, Wang YJ and Bian Q: The osteoprotective effect of psoralen in
ovariectomy-induced osteoporotic rats via stimulating the
osteoblastic differentiation from bone mesenchymal stem cells.
Menopause. 19:1156–1164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li CW, Liang B, Shi XL and Wang H:
Opg/Rankl mRNA dynamic expression in the bone tissue of
ovariectomized rats with osteoporosis. Genet Mol Res. 14:9215–9224.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Faienza MF, Ventura A, Marzano F and
Cavallo L: Postmenopausal osteoporosis: the role of immune system
cells. Clin Dev Immunol. 575936:2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kararigas G, Nguyen BT and Jarry H:
Estrogen modulates cardiac growth through an estrogen receptor
α-dependent mechanism in healthy ovariectomized mice. Mol Cell
Endocrinol. 382:909–914. 2014. View Article : Google Scholar
|
|
34
|
Gong J, Meng HB, Hua J, Song ZS, He ZG,
Zhou B and Qian MP: The SDF-1/CXCR4 axis regulates migration of
transplanted bone marrow mesenchymal stem cells towards the
pancreas in rats with acute pancreatitis. Mol Med Rep. 9:1575–1582.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu Q, Shao H, Darwin ED, Li J, Li J, Yang
B, Webster KA and Yu H: Extracellular calcium increases CXCR4
expression on bone marrow-derived cells and enhances
pro-angiogenesis therapy. J Cell Mol Med. 13:3764–3773. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bhakta S, Hong P and Koc O: The surface
adhesion molecule CXCR4 stimulates mesenchymal stem cell migration
to stromal cell-derived factor-1 in vitro but does not decrease
apoptosis under serum deprivation. Cardiovasc Revasc Med. 7:19–24.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wynn RF, Hart CA, Corradi-Perini C,
O'Neill L, Evans CA, Wraith JE, Fairbairn LJ and Bellantuono I: A
small proportion of mesenchymal stem cells strongly expresses
functionally active CXCR4 receptor capable of promoting migration
to bone marrow. Blood. 104:2643–2645. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Long MY, Li HH, Pen XZ, Huang MQ, Luo DY
and Wang PS: Expression of chemokine receptor-4 in bone marrow
mesenchymal stem cells on experimental rat abdominal aortic
aneurysms and the migration of bone marrow mesenchymal stem cells
with stromal-derived factor-1. Kaohsiung J Med Sci. 30:224–228.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qian D, Gong J, He Z, Hua J, Lin S, Xu C,
Meng H and Song Z: Bone marrow-derived mesenchymal stem cells
repair necrotic pancreatic tissue and promote angiogenesis by
secreting cellular growth factors involved in the SDF-1α/CXCR4 axis
in rats. Stem Cells Int. 302015:2015. View Article : Google Scholar
|
|
40
|
Zou Z, Zhang Y, Hao L, Wang F, Liu D, Su Y
and Sun H: More insight into mesenchymal stem cells and their
effects inside the body. Expert Opin Biol Ther. 10:215–230. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Karp JM and Leng Teo GS: Mesenchymal stem
cell homing: the devil is in the details. Cell Stem Cell.
4:206–216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Honczarenko M, Le Y, Swierkowski M, Ghiran
I, Glodek AM and Silberstein LE: Human bone marrow stromal cells
express a distinct set of biologically functional chemokine
receptors. Stem Cells. 24:1030–1041. 2006. View Article : Google Scholar
|
|
43
|
Safwani WK, Makpol S, Sathapan S and Chua
KH: Alteration of gene expression levels during osteogenic
induction of human adipose derived stem cells in long-term culture.
Cell Tissue Bank. 14:289–301. 2013. View Article : Google Scholar
|
|
44
|
Choi MR, Kim HY, Park JY, Lee TY, Baik CS,
Chai YG, Jung KH, Park KS, Roh W, Kim KS, et al: Selection of
optimal passage of bone marrow-derived mesenchymal stem cells for
stem cell therapy in patients with amyotrophic lateral sclerosis.
Neurosci Lett. 472:94–98. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhao D, Li XP, Gao M, Zhao C, Wang JL and
Wei LH: Stromal cell-derived factor 1α stimulates human endometrial
carcinoma cell growth through the activation of both extracellular
signal-regulated kinase 1/2 and Akt. Gynecol Oncol. 103:932–937.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bobis-Wozowicz S, Miekus K, Wybieralska E,
Jarocha D, Zawisz A, Madeja Z and Majka M: Genetically modified
adipose tissue-derived mesenchymal stem cells overexpressing CXCR4
display increased motility, invasiveness, and homing to bone marrow
of NOD/SCID mice. Exp Hematol. 39:686–696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Brennecke P, Arlt MJ, Campanile C, Husmann
K, Gvozdenovic A, Apuzzo T, Thelen M, Born W and Fuchs B: CXCR4
antibody treatment suppresses metastatic spread to the lung of
intratibial human osteosarcoma xenografts in mice. Clin Exp
Metastasis. 31:339–349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yu J, Li M, Qu Z, Yan D, Li D and Ruan Q:
SDF-1/CXCR4-mediated migration of transplanted bone marrow stromal
cells toward areas of heart myocardial infarction through
activation of I3K/Akt. J Cardiovasc Pharmacol. 55:496–505.
2010.PubMed/NCBI
|
|
49
|
Yu ZH, Wang YX, Song Y, Lu HZ, Hou LN, Cui
YY and Chen HZ: Up-regulation of KCa3.1 promotes human airway
smooth muscle cell phenotypic modulation. Pharmacol Res. 77:30–38.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kollet O, Shivtiel S, Chen YQ, Suriawinata
J, Thung SN, Dabeva MD, Kahn J, Spiegel A, Dar A, Samira S, et al:
HGF, SDF-1, and MMP-9 are involved in stress-induced human
CD34+ stem cell recruitment to the liver. J Clin Invest.
112:160–169. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shahzad U, Li G, Zhang Y, Li RK, Rao V and
Yau TM: Trans-myocardial revascularization enhances bone marrow
stem cell engraftment in infarcted hearts through SCF-C-kit and
SDF-1CXCR4 signaling axes. Stem Cell Rev. 11:332–346. 2015.
View Article : Google Scholar
|
|
52
|
Liu N, Tian J, Cheng J and Zhang J:
Migration of CXCR4 gene-modified bone marrow-derived mesenchymal
stem cells to the acute injured kidney. J Cell Biochem.
114:2677–2689. 2013. View Article : Google Scholar : PubMed/NCBI
|