Introduction
Rapidly growing tumors have large energy demands,
but the insufficient nascent vasculature fails to provide
sufficient nutrition and metabolic substrates for malignant
progression. Thus, cancer cells induce adjacent activated
fibroblasts to increase lactate production via the glycolysis
pathway. Subsequently, the cancer cells use metabolic intermediates
release from fibroblasts to 'fuel' tumor growth (1,2).
This reciprocal metabolic reprogramming between cancer cells and
the surrounding stroma has been widely recognized as a 'reverse
Warburg effect' (3). As the most
common metabolic intermediate of glycolysis, lactate is transported
via unidirectional mono-carboxylate transporters (MCTs). This
'lactate shuttle' was first reported between astroglial cells and
neurons through MCT1 and MCT2 transporters, respectively (4,5).
In the tumor microenvironment (TME), MCTs control the lactate
exchange between glycolytic and oxidative cancer cells, and between
stromal and epithelial cells (5).
In the past decade, research has revealed a one-way flow of lactate
shuttling from tumor mesenchyme to epithelium (6). This lactate fluxion has essential
roles not in adjusting intracellular acid-base balance (7,8),
and also providing metabolic fuel for cancer cells.
Oral squamous cell carcinoma (OSCC) is the most
common oral malignancy with poor 5-year survival rate (9). Components of the tumor stroma,
particularly cancer-associated fibroblasts (CAFs), may be partially
responsible for the unfavorable therapeutic results and poor
prognosis of OSCC. This raises the questions, is the 'reverse
Warburg effect' involved in OSCC progression? Does the
stromal-epithelial lactate shuttle 'fuel' oral cancer growth? In
our previous study, bioinformatics network construction identified
interleukin-1β (IL-1β) as one of the key node genes in the TME
during oral carcinogenesis (10).
Is this tumor derived-IL-1β a major contributing factor for the
lactate flux to cancer cells?
In this study, it was aimed to investigate the
function of cancer cell-derived IL-1β on reciprocal metabolic
reprogramming between the epithelium and mesenchyme. Thus, a
co-culture system was created using OSCC cells and separated
fibroblasts from patients or healthy volunteers, and the results
revealed that interactions between stromal and epithelial cells
upregulated IL-1β expression in tumor cells, which enhanced
glycolysis and lactate release in CAFs, which subsequently resulted
in accelerated malignant cell proliferation.
Materials and methods
Cell lines
UM1, SCC25 and CAL27 human OSCC cell lines and
normal oral keratinocyte (NOK) were used in this study. SCC25 and
CAL27 were purchased from American Type Culture Collection
(Manassas, VA, USA). OSCC cell line UM1 and NOK were provided by
Professor J. Silvio Gutkind (University of California, San Diego,
CA, USA). UM1, SCC25 and CAL27 were maintained in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) containing 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.). NOK cells were maintained
in Keratinocyte-SFM (Gibco; Thermo Fisher Scientific, Inc.). Cells
were incubated at 37°C in 5% CO2. This study was
approved by the Ethical Review Committee of Guanghua School of
Stomatology, Hospital of Stomatology, Institute of Stomatological
Research, Sun Yat-sen University (Guangzhou, China; approval no.
ERC-2012-15). All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. And all participants provided written informed consent
to take part in the study.
Sample selection and primary cell
culture
Fibroblasts separated from patients with OSCC,
patients with oral leukoplakia and healthy volunteers, were termed
CAFs (n=8), Dysplasia-fs (n=8) and normal fibroblasts (NFs; n=10),
respectively. All samples were obtained from Department of Oral and
Maxillofacial Surgery, Guanghua School of Stomatology, Sun Yat-sen
University between February 2014 and September 2015. All patients
provided written informed consent to participate in this study.
Samples were cautiously washed with phosphate-buffered saline and
then digested with 0.2% neutral protease Dispase II (Roche
Diagnostics GmbH, Mannheim, Germany) overnight at 4°C. The stromal
layer was carefully split from the epithelial layer and shredded
into tiny fragments. The fragments were digested with 0.25%
trypsin-EDTA (Gibco; Thermo Fisher Scientific, Inc.) at 37°C for 1
min. Following neutralization, centrifugation (at 120 × g for 5
min) and re-suspension at room temperature (RT), fragments were
seeded into flasks and incubated in primary culture medium (DMEM
containing penicillin, streptomycin and 20% FBS) at 37°C for ~14
days. Subsequently, the fibroblasts were sub-cultured and purified
according to the 'adherent time lag' between fibroblasts and
epithelial cells. Fibroblasts between passage 4 and 10 (P4-P10)
were used.
Co-culture system and fibroblasts
activation
OSCC cells and fibroblasts were seeded onto the
insert or the bottom well of co-culture Transwell inserts (Corning
Incorporated, Corning, NY, USA), respectively, and independently
cultured until the cells adhered. The co-culture durations are
slightly different between the experiments, and the specific time
has been noted in each figure. Once adhered, the insert was placed
back into the well for co-culture.
Immunofluorescence (IF) staining
Cells were fixed with pre-cooled 100% methanol for
20 min at −20°C, following permeabilizing [using 0.1% Tween-20 and
1% bovine serum albumin (BSA; Beyotime Biotechnology, Shanghai,
China)] for 10 min at RT and antigen blocking for 30 min ar RT,
cells were successively incubated with mouse monoclonal
anti-α-smooth muscle actin (α-SMA; cat. no. ab7817; Abcam,
Cambridge, MA, USA; diluted 1:200) at 4°C overnight, secondary
antibody H&L DyLight® 488 (cat. no. ab96875; Abcam;
diluted 1:200) at RT for 1 h, and 100 ng/ ml DAPI (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) at RT for 5 min. Images were
acquired with a confocal microscope (Carl Zeiss AG, Oberkochen,
Germany).
Glucose uptake and lactate assays
The conditioned media (CM) was harvested and stored
at −80°C. Glucose and lactate content were measured with a glucose
assay kit (Applygen Technologies, Inc., Beijing, China) and lactate
assay kit (Nanjing KeyGen Biotech, Co., Ltd., Nanjing, China),
respectively, according to the manufacturers' protocols. Absorbance
values of test groups were normalized against protein
concentrations (n=3).
IL-1β ELISA assay
CM from NOK and OSCC cell lines was harvested after
48 h incubation. IL-1β was measured with ELISA kit (cat. no.
SEA563Hu; Cloud-Clone Corp., Katy, TX, USA) according to the
manufacturer's protocol. The values were normalized to the cell
count (n=3).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific, Inc.) and then reverse transcribed using qScript
cDNA synthesis kit (Roche Diagnostics GmbH). All of the primers
were produced by Takara Biotechnology Co., Ltd. (Dalian, China;
Table I). qPCR was performed
using the the LightCycler® 480 SYBR-Green I Master and
the LightCycler® 480 instrument (Roche Diagnostics
GmbH). PCR was carried out at 95°C (10 min) and 45 cycles at 95°C
(10 sec), 65°C (15 sec) and 72°C (15 sec). Each step was performed
according to the manufacturers' protocols. The amounts of target
genes were normalized to GAPDH using 2−ΔΔCq calculations
(11).
 | Table IPrimer nucleotide sequences. |
Table I
Primer nucleotide sequences.
| Gene | Primer nucleotide
sequences (5′ to 3′) | Product size
(bp) |
|---|
| GAPDH | F:
GCACCGTCAAGGCTGAGAAC | 138 |
| R:
TGGTGAAGACGCCAGTGGA |
| GLUT1 | F:
GCCTGAAGTCGCACAGTGAATAA | 145 |
| R:
GCTCATTGGGCCCATACAAAG |
| HK2 | F:
CTCAACCATGACCAAGTGCAGAA | 97 |
| R:
CCTTGCGGAACCGCTTAGAG |
| PFKM | F:
GCCAGTCTAATTGCCGTTCC | 212 |
| R:
TACCAACTCGAACCACAGCC |
| PKM2 | F:
CCACTTGCAATTATTTGAGGAA | 148 |
| R:
GTGAGCAGACCTGCCAGACT |
| LDHA | F:
ATCTTGACCTACGTGGCTTGGA | 180 |
| R:
CCATACAGGCACACTGGAATCTC | |
| MCT1 | F:
TTATCCTGCCACACCAGCAG | 352 |
| R:
TGCTGTCACACACAGACACA | |
| MCT4 | F:
ATTGGCCTGGTGCTGCTGATG | 243 |
| R:
CGAGTCTGCAGGAGGCTTGTG | |
| IL-1β | F:
TCGCCAGTGAAATGATGGCTTA | 197 |
| R:
GTCCATGGCCACAACAACTGA | |
Western blotting
Cells were harvested and lysed in
radioimmunoprecipitation assay buffer and measured with
bicinchoninic acid protein assay kit (both from Sigma-Aldrich:
Merck KGaA). Then samples (30 mg/lane) were separated by 10%
SDS-PAGE and transferred to a polyvinylidene difluoride membrane.
Following antigen blocking in 5% nonfat milk for 1 h at RT, the
membrane was incubated with primary antibodies overnight at 4°C.
The following primary antibodies were used: Rabbit monoclonal
anti-α-tubulin (cat. no. 2125; diluted 1:1,000), anti-GAPDH (cat.
no. 2118; diluted 1:1,000), anti-hexokinase 2 (HK2) (cat. no. 2867;
diluted 1:1,000), anti-lactate dehydrogenase (LDHA; cat. no. 3582;
diluted 1:1,000) and anti-IL-1β (cat. no. 12703; diluted 1:1,000)
all from Cell Signaling Technology, Inc. (Danvers, MA, USA); mouse
monoclonal anti-MCT1 (cat. no. sc-365501; diluted 1:200) and rabbit
polyclonal anti-MCT4 (cat. no. sc-50329; diluted 1:400) from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA); mouse monoclonal
anti-glucose transporter 1 (GLUT1) (cat. no. 202921; diluted 1:250)
from R&D Systems, Inc. (Minneapolis, MN, USA). The membrane was
then washed three times with TBS-Tween, and incubated with
secondary antibodies goat anti-rabbit IgG (cat. no. 7074; diluted
1:2,000) or horse anti-mouse IgG (cat. no. 7076; diluted 1:2,000)
both from Cell Signaling Technology, Inc. for 1 h at RT, according
to the source of primary antibodies. The immune-reactive bands were
visualized with an enhanced chemiluminescence system (EMD
Millipore, Billerica, MA, USA). Band intensities were
semi-quantitated using ImageJ software (National Institutes of
Health).
Small interfering RNA (siRNA)-mediated
downregulation of IL-1β
IL-1β siRNA (si-IL-1β) was designed by and purchased
from Invitrogen (Thermo Fisher Scientific, Inc.). Cells were
transfected with siRNA pool (including the following sequences:
IL-1β-siR NA1 for ward, 5′-GCUCGCCAGUGAAAUGAUGGCUUAU-3′ and
reverse, 5′-AUAAGCCAUCAUUUCACUGGCGAGC-3′; IL-1β-siRNA2 forward,
5′-GGAUGACUUGUUCUUUGAAGCUGAU-3′ and reverse
5′-AUCAGCUUCAAAGAACAAGUCAUCC-3′; IL-1β-siRNA3 forward,
5′-GGAUAUAACUGACUUCACCAUGCAA-3′ and reverse
5′-UUGCAUGGUGAAGUCAGUUAUAUCC-3′) or Stealth RNAi®
negative control (cat. no. 12935200) by Lipofectamine®
RNAiMAX reagent (Invitrogen; Thermo Fisher Scientific, Inc.). UM1
cells (1×106/ml) were cultured in 6-well plates with
DMEM containing 10% FBS for 24 h and maintained in a 37°C incubator
under a humidified atmosphere containing 5% CO2 (until
the density of the transfected cells reached 30–80%). The siRNA or
negative control were transfected into the cells using
Lipofectamine® RNAiMAX at 10 nM according to the
manufacturer's instructions. The transfection efficiency was
confirmed by RT-qPCR and western blot analysis (data not
shown).
Proliferation assay with
carboxy-fluorescein succinimidyl ester (CFSE)
On the basis that CFSE cell fluorescence
progressively decreases as cells divide, UM1 cell proliferation was
determined using CFSE. Cells were re-suspended in 1 µM CellTrace™
CFSE staining solution (Invitrogen; Thermo Fisher Scientific,
Inc.), and incubated at 37°C for 20 min, protected from light. Then
5 times the original staining volume of complete culture medium was
added to the cells and the mixture as incubated at 37°C for a
further 5 min to terminate the staining reaction. Following three
washes with culture media, UM1 cells were co-cultured with
fibroblasts for 72 h or cultured alone as the control. UM1 cells
were detached from the plates and fixed with 4% paraformaldehyde at
24 h intervals at RT. Subsequently, the CFSE fluorescence was
detected by flow cytometry (Beckman Coulter, Inc., Brea, CA, USA)
with an excitation light of 488 nm. Results were expressed as a
proliferation index (mean level of cell divisions frequency), using
ModFit V3.0 software (Verity Software House, Topsham, ME, USA).
Statistical analysis
Data are presented as the mean ± standard deviation
from at least three independent experiments. Statistical analysis
of the data was performed by Student's t-test, one-way ANOVA using
SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA). P<0.05
(two-tailed) was considered to indicate a statistically significant
difference.
Results
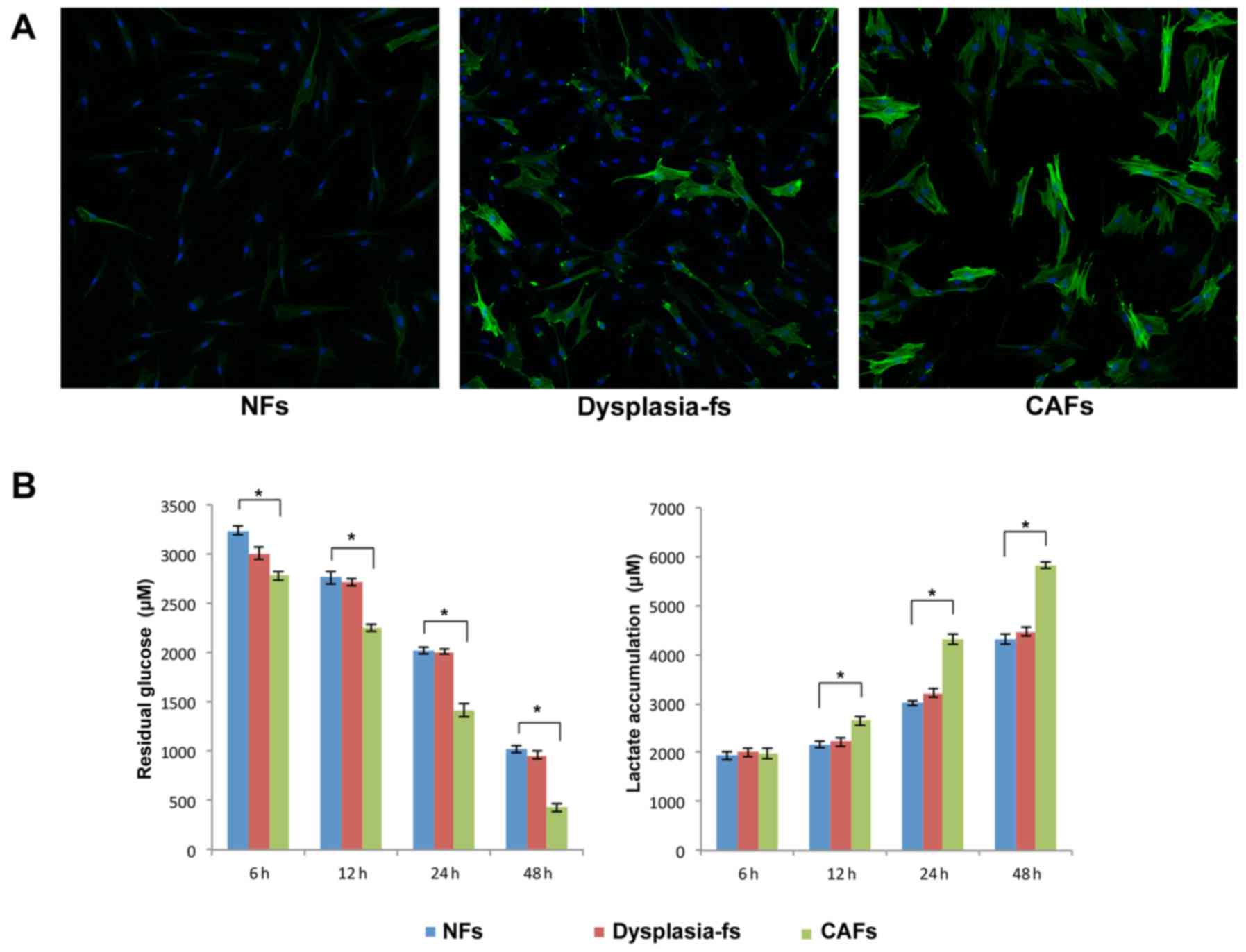
Glycolysis is increased in activated
fibroblasts from patients with oral carcinogenesis
To investigate the biological characteristic of
fibroblasts isolated from oral premalignant and malignant lesions,
IF staining for α-SMA was performed. A marked increase in α-SMA
expression was observed in Dysplasia-fs and CAFs, compared with
negative/low expression in NFs (Fig.
1A). These results indicated that fibroblasts were activated
during oral carcinogenesis. Additionally, glucose and lactate assay
kits were used to measure the levels of fibroblasts glycolysis.
Glucose utilization and lactate production were increased
significantly in CAFs compared with NFs (Fig. 1B).
Enhanced stromal glycolysis is cancer
cell-dependent
To evaluate the association between stromal
glycolysis and oral cancer cells, NFs and UM1 cells were
co-cultured using a Transwell insert for 48 h, and then α-SMA
staining was performed on the fibroblasts. As shown in Fig. 2A, fibroblasts were activated
following cultivation with oral cancer cells and α-SMA expression
of co-cultured fibroblasts was increased compared with fibroblasts
without co-cultivation.
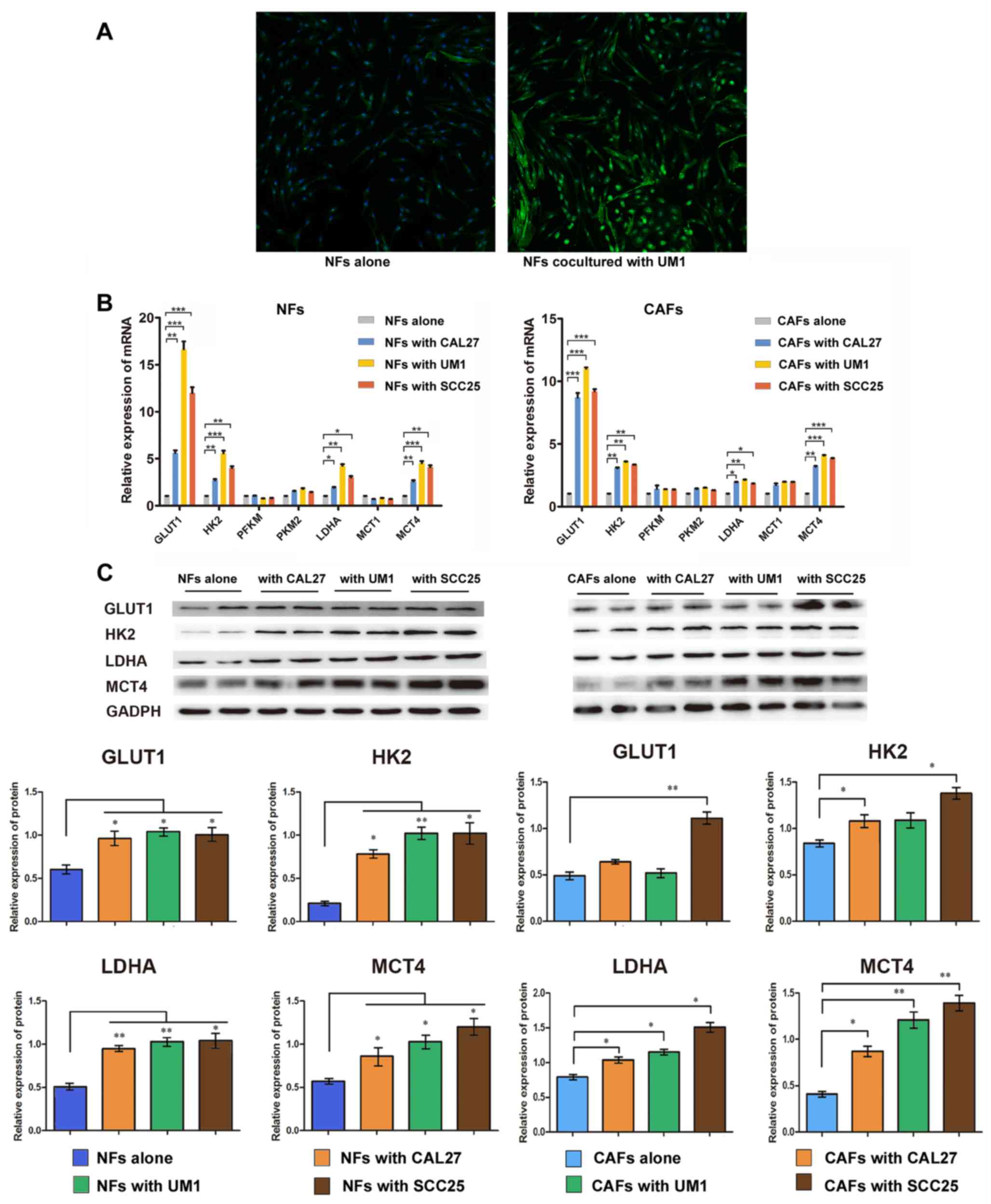 | Figure 2Oral cancer cells enhanced glycolysis
of fibroblasts. (A) α-smooth muscle actin (green) staining in NFs
cultured alone and NFs co-cultured with UM1 cells. Magnification,
×50. Fibroblasts (NFs and CAFs) were co-cultured with oral squamous
cell carcinoma cell lines (CAL27, UM1 and SCC25) respectively, for
48 h before harvest. (B) Reverse transcription-quantitative
polymerase chain reaction analysis for the expression of a series
of glycolytic genes in fibroblasts. Columns present the mean ± SD
of triplicate determinations. (C) Western blot analysis of GLUT1,
HK2, LDHA and MCT4 in NFs and CAFs. Densitometry was used to
determine target gene/GAPDH ratios. Data are presented as the mean
± SD of three independent experiments. *P<0.05 and
**P<0.01 vs. cells cultured alone. SD, standard deviation; NF,
normal fibroblasts; CAFs, cancer-associated fibroblasts; GLUT1,
glucose transporter 1; HK2, hexokinase 2; PFK, phosphofructokinase;
PKM2, pyruvate kinase M2; LDHA, lactate dehydrogenase; MCT,
mono-carboxylate transporter. |
Activated fibroblasts were harvested after 48 h
co-culture with CAL27, UM1 and SCC25 for subsequent analysis of
glycolysis-associated gene expression. RT-qPCR and western blot
results (Fig. 2B and C) showed
that GLUT1, HK2, LDHA and MCT4 expression was increased in
activated fibroblasts compared with fibroblasts cultured alone.
Among these genes, GLUT1 was significantly upregulated with the
highest fold change of mRNA level. Taken together, these data
indicated that OSCC-activated fibroblasts displayed enhanced
glycolysis, accompanied by an increase the expression of glycolysis
associated genes. In our previous study, IL-1β was identified as a
key node gene in OSCC progression, and elevated IL-1β expression
was parallel with oral carcinogenesis (10). Thus, does IL-1β participate in
stromal glycolysis induced by oral cancer cells?
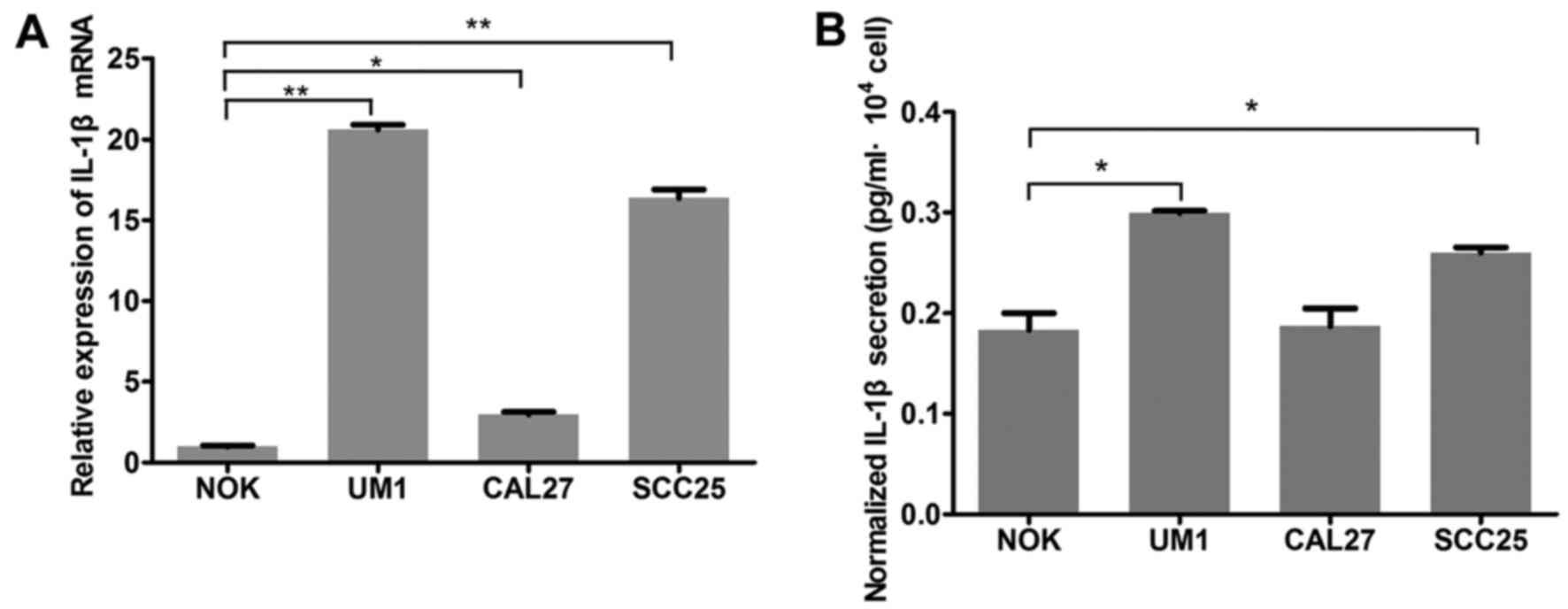
IL-1β secretion from oral cancer cells is
increased by activated fibroblasts
IL-1β mRNA expression was signifi-cantly increased
in OSCC cell lines, compared with NOK cells (Fig. 3A). ELISA results demonstrated that
UM1 and SCC25 cells secreted more IL-1β than NOK cells, and
particularly that IL-1β secretion of UM1 was nearly twice that of
NOK (Fig. 3B). However, CAL27
cells exhibited elevated IL-1β expression at the mRNA level only,
with no significant alteration in IL-1β secretion detected by
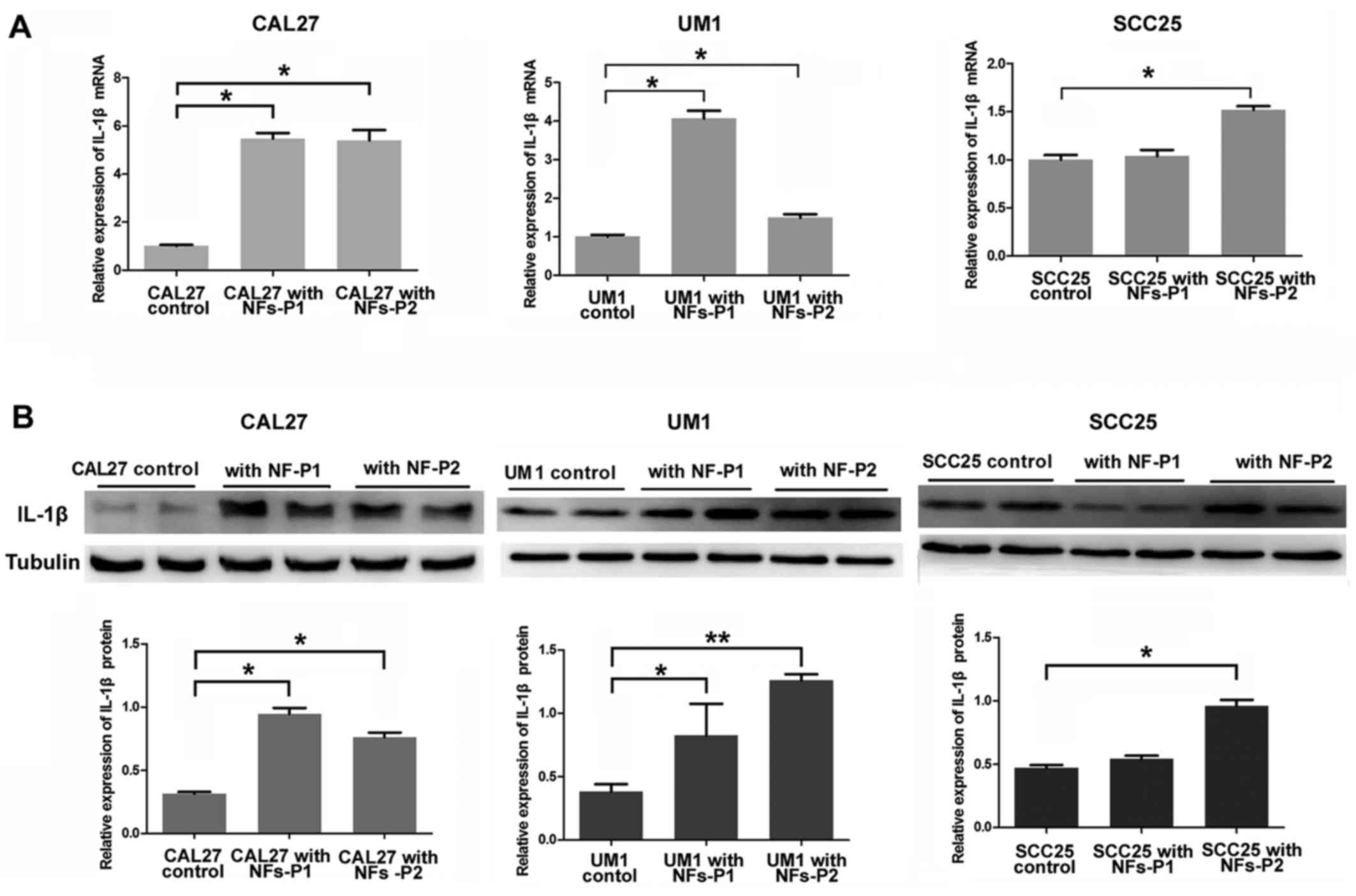
ELISA. Furthermore, when OSCC cells were co-cultured with
fibroblasts (NFs), all the OSCC cell lines in our experiment
expressed increasing IL-1β in a time-dependent manner, moreover,
and UM1 cell was the most sensitive to the stimulation of
fibroblasts (Fig. 4).
Accordingly, the UM1 cell line was selected for subsequent IL-1β
siRNA transfection experiments.
Enhanced stromal glycolysis in OSCC may
be IL-1β-dependent
To determine the effect of IL-1β on tumor
metabolism, IL-1β antagonists (IL-1Ra) was used to neutralize
secreted IL-1β in the supernatant, and IL-1β siRNA was used to
knock down endogenous IL-1β in OSCC cells, whereas exogenous
recombinant human IL-1β was used to promote IL-1β-induced effects.
OSCC cells and fibroblasts were treated with human recombination
IL-1β (20 ng/ml) alone or both IL-1β (20 ng/ml) and IL-1Ra (20
mg/ml). CM were harvested at 6, 12, 24 and 48 h. No significant
change in glucose consumption and lactate production was identified
in OSCC cell lines following treatment. However, NFs and CAFs
exhibited significantly increased glucose consumption after 24 h of
IL-1β treatment, and lactate accumulation increased was after IL-1β
treatment for 24 h in NFs and 48 h in CAFs. However, IL-1Ra
significantly attenuated the IL-1β-induced effects on glucose
consumption and lactate production in fibroblasts (Fig. 5A). Fibroblasts were harvested
after IL-1β treatment for 48 h, and RT-qPCR assay was performed to
investigate the expression of glycolysis-associated genes. As shown
in Fig. 5B, GLUT1 and HK2
expression was significantly increased in the IL-1β treated group
of NFs and CAFs. MCTs changed synergistically, with decreased
expression of MCT1 and increased expression of MCT4, compared with
untreated controls, respectively (Fig. 5B). However, when UM1 cells were
transfected with si-IL-1β, GLUT1, LDHA and MCT4 expression
significantly decreased, compared with the negative control siRNA
group, (Fig. 5C). Collectively,
these observations indicated that IL-1β stimulates glycolysis of
activated oral fibroblasts.
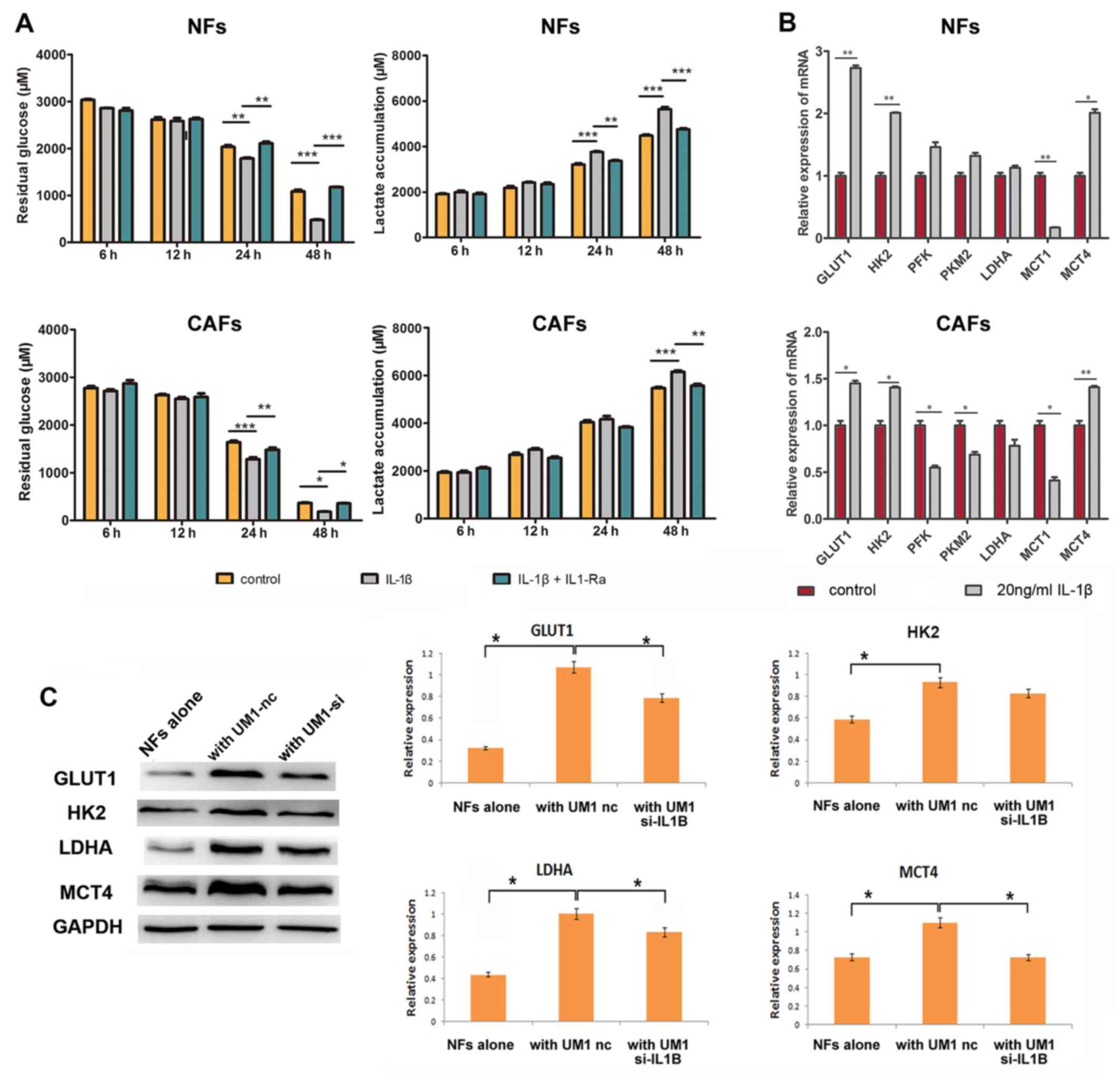 | Figure 5Tumor-derived IL-1β induces stromal
glycolysis. (A) NFs and CAFs were treated with 20 ng/ml IL-1β alone
or 20 ng/ml IL-1β + 20 mg/ml IL-1Ra for 48 h. CM were harvested at
6, 12, 24 and 48 h for residual glucose and lactate consumption
analysis using glucose and lactate assay kits, respectively. (B)
NFs and CAFs treated with 20 ng/ml IL-1β were collected for reverse
transcription-quantitative polymerase chain reaction assays.
Columns represent the mean ± standard deviation of three
independent experiments. *P<0.05,
**P<0.01 and ***P<0.001. (C) NFs and
UM1 cells transfected with 10 nM nc or 10 nM si-IL1B were
co-cultured for 48 h. NFs were then harvested for western blot
analysis of GLUT1, HK2, LDHA and MCT4. Densitometry was used to
determine target gene/GAPDH ratios. *P<0.05 vs.
UM1-nc group. IL-1β, interleukin-1β; NFs, normal fibroblasts; CAFs,
cancer-associated fibroblasts; IL-1Ra, interleukin-1 receptor
antagonist; GLUT1, glucose transporter 1; HK2, hexokinase 2; LDHA,
lactate dehydrogenase; MCT, mono-carboxylate transporter; siRNA,
small interfering RNA; nc, negative control siRNA; si-IL1B, IL-1β
siRNA. |
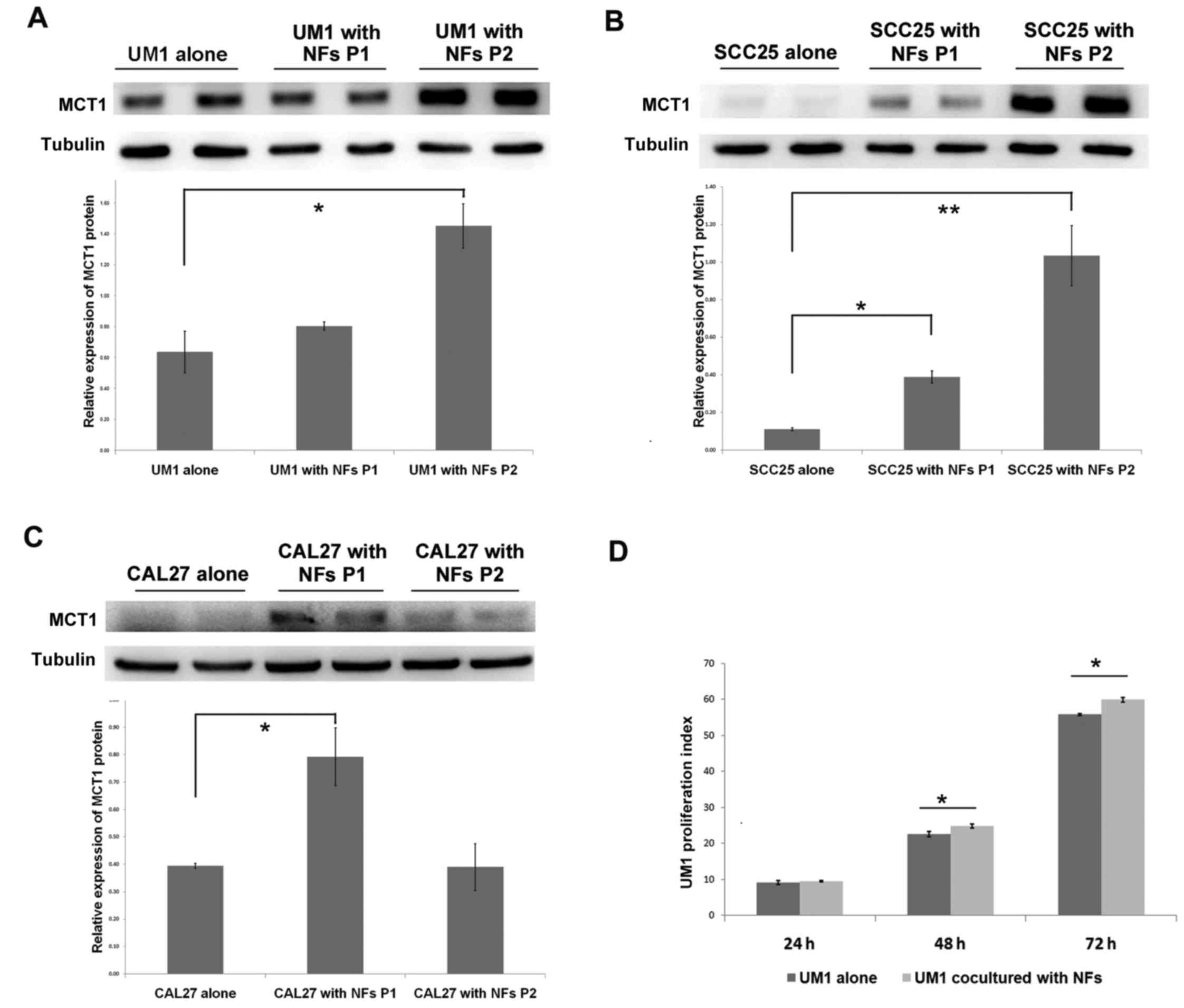
Fibroblasts promote lactate uptake and
proliferation of OSCC
The aforementioned findings demonstrated that oral
cancer cells enhanced stromal glycolysis in the activated
fibroblasts by secreting increased IL-1β, which resulted in MCT4 (a
lactate excretion transporter) upregulation in fibroblasts. To
further investigate the lactate destination and OSCC response to
this, the lactate uptake transporter MCT1 expression and cell
proliferation of OSCC cells in the co-culture system was examined.
In UM1 and SCC25 cells, MCT1 protein expression increased in a
time-dependent manner, and in CAL27 cells there was significant
upregulation of MCT1 in the first passage of cultivation (Fig. 6A–C). UM1 cells were pre-stained
with CFSE dye (which is equally distributed to two daughter cells
during mitosis), and then co-cultured with NFs to detect the effect
of stromal-epithelial lactate shuttle on OSCC cell proliferation.
As shown in Fig. 6D, the
proliferation index of UM1 cells in the co-culture group was
significantly higher than that in UM1 cells cultured alone after 48
and 72 h (Fig. 6D). The above
data suggest that lactate shuttle from activated oral fibroblasts
to oral cancer cells promotes cancer cell proliferation.
Discussion
Increasing evidence has highlighted the importance
of reciprocal metabolic reprogramming and lactate shuttling in
epithelial malignancies. However, the stromal-epithelial lactate
shuttle and the underlying mechanisms during oral carcinogenesis
still remain elusive. CAFs, also known as activated fibroblasts,
are the biggest population of cancer stromal cells and differ from
NFs in many respects, particularly epigenetics (12). For example, CAFs usually present
with a caveolin-1 (cav-1) loss (13) and express active markers such as
α-SMA, stromal cell derived factor-1 (14) and fibroblast activation protein
(15). Gene expression analysis
demonstrated that the activated fibroblasts, CAFs, secrete a large
amount of proteins and enzymes that disrupt the extracellular
matrix, recruit immunocytes, and promote angiogenesis and
metastasis (16). The findings of
the current study demonstrated that fibroblasts from different
stages of OSCC progression, NFs, Dysplasia-fs and CAFs, displayed
increased α-SMA expression, which suggested that the fibroblasts
were activated during oral carcinogenesis. Additionally, these
fibroblasts also exhibited increased glucose uptake and lactate
accumulation in their supernatant, which indicated that glycolysis
was enhanced fibroblasts from the TME. The increase in glycolysis
in fibroblasts had been demonstrated in various solid human tumors
(17). Pavlides et al
(17), Bonuccelli et al
(18) and Pavlides et al
(19) reported that glycolytic
enzymes were unregulated in CAFs with cav-1 loss, and they proposed
that hydrogen peroxide secreted by cancer cells induced oxidative
stress, mitophagy and aerobic glycolysis in CAFs.
It has been previously reported that cancer cells
utilize fibroblast-produced intermediate products, such as lactate,
ketone and glutamine, as nutrient substances (20). In human prostate cancer, tracing
experiments explicitly demonstrated the lactate flows from
fibroblasts to cancer cells in a common co-culture system. This
metabolic symbiosis phenomenon was an important supplement to the
Warburg effect, termed the 'reverse Warburg effect', which
indicates that there must be a reciprocal metabolic influence
between CAFs and cancer cells. In the current study, co-culture
with OSCC cells significantly enhanced glycolytic gene expression
in fibroblasts.
Our recent study identified IL-1β as a key node gene
during oral carcinogenesis, both in epithelium and sub-mucosal
fibroblasts. Notably, the IL-1β expression in epithelium increased
gradually as oral malignancy progressed (10). Inversely, saliva exosome analysis
of patients with OSCC revealed that secretory IL-1β significantly
decreased following surgical treatment (21). OSCC cells were further confirmed
as the source of intercellular IL-1β. In the current study, it was
confirmed that IL-1β expression and secretion were upregulated in
OSCC cell lines (UM1, SCC25 and CAL27). In the co-culture system,
activated fibroblasts stimulated IL-1β expression in OSCC cells.
Since the co-culture system imitated the epithelial-stromal
interaction in vivo, it is deduced that activated
fibroblasts participate in promoting IL-1β production in
premalignant and malignant oral epithelium, and this substantial
tumor-derived IL-1β may be a vital signal mediating
epithelium-mesenchyme dialogue.
The results of the present study demonstrated that
IL-1β expression in OSCC, activation of fibroblasts and aerobic
glycolysis in fibroblasts were simultaneously enhanced during
malignant progression of oral cancer cells. Does tumor-derived
IL-1β enhance stromal glycolysis and induce lactate shuttle to the
cancer cells? Further experiments demonstrated that exogenous
IL-1β-treated NFs consumed more glucose and release more lactate
into the supernatant. At the same time, the expression of
glycolysis and lactate emission-associated genes, such as GLUT1,
HK2, LDHA and MCT4, was increased in the IL-1β-treated NFs.
Conversely, IL-Ra significantly abrogated the IL-1β-induced
glycolysis. Knockdown using si-IL-1β in UM1 cells effectively
blocked the regulatory effect of OSCC on stromal glycolysis. For
OSCC cells in the co-culture system, the lactate uptake
transporter, MCT1 was correspondingly overexpressed. These
differentially expressed genes pointed to the reverse Warburg
effect, because GLUT1 is the predominant facilitative glucose
transporter in human cells, and overexpression of GLUT1 usually
predicts increased glucose uptake (22,23). HK2 is a key rate-limiting enzyme
in glycolysis, and LDHA catalyzes the conversion of pyruvate, a
glycolysis mediator-production, into lactate (24,25). MCT1/4 are vital lactate
transporters involved in acid-base balance, and promoting glucose
metabolism in tumor tissues (26). MCT4, usually expressed in
glycolytic cells, is especially enriched in cells of high oxygen
consumption, while MCT1 is rather ubiquitous (5).
MCT4 overexpression in CAFs and MCT1 overexpression
in transformed epithelium are regarded as independent indicators of
poor outcomes in human carcinoma (27). The intercellular coordination of
MCT1/4 expression in fibroblasts and cancer cells strongly indicate
a unidirectional lactate shuttle, from stromal cells to the cancer
cells (6,28,29). Thus, fibroblasts directly feed
neighboring cancer cells via lactate. In this study, MCT4 in
fibroblasts and MCT1 in OSCC cells were upregulated by the
stromal-epithelial co-culture, suggesting that lactate may be
transported to the oxidative cancer cells. Additionally, the
proliferation of OSCC cells was accelerated following
co-cultivation with the fibroblasts. All these findings support the
regulatory function of IL-1β in the reverse Warburg effect and
lactate reuse by cancer cells.
However, the underlying mechanism of IL-1β-modulated
metabolic reprogramming remains ambiguous. Preliminarily results
demonstrated that hypoxia inducible factor-1α (HIF-1α) may be
partially responsible (data not shown), as IL-1β directly promoted
HIF-1α expression in fibroblasts, which is a regulatory
transcription factor of various metabolic genes (30). Other potential mechanism have also
been reported, including that IL-1β induces accumulation of
reactive oxygen species (12,31), and that IL-1β directly activates
caspases in mitochondria (32),
resulting in TCA cycle inhibition and promotes fibroblasts to
utilize glucose through an inefficient method, anaerobic
glycolysis.
In conclusion, the results of the present study
identified a regulatory loop between fibroblasts and oral cancer
cells by which cancer cells activate fibroblasts and fibroblasts
promote IL-1β expression in OSCC. In turn, this tumor-derived IL-1β
induces lactate emission from fibroblasts to feed cancer cells as
'fuel'. The findings provide novel insight into the role of IL-1β
and lactate shuttle in oral cancer, and provide new therapeutic
targets for future OSCC treatment.
Acknowledgments
This study was supported by grants of the National
Natural Science Foundation of China (nos. 81272948, 81200787 and
81371148).
Abbreviations:
|
OSCC
|
oral squamous cell carcinoma
|
|
CAFs
|
cancer-associated fibroblasts
|
|
NFs
|
normal fibroblasts
|
|
IL-1β
|
interleukin-1β
|
|
α-SMA
|
α-smooth muscle actin
|
|
GLUT1
|
glucose transporter 1
|
|
HK2
|
hexokinase 2
|
|
LDHA
|
lactic dehydrogenase
|
|
MCT1
|
mono-carboxylate transporter 1
|
|
MCT4
|
mono-carboxylate transporter 4
|
|
CFSE
|
carboxyfluorescein succinimidyl
ester
|
References
|
1
|
Martinez-Outschoorn UE, Lisanti MP and
Sotgia F: Catabolic cancer-associated fibroblasts transfer energy
and biomass to anabolic cancer cells, fueling tumor growth. Semin
Cancer Biol. 25:47–60. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Paolicchi E, Gemignani F, Krstic-Demonacos
M, Dedhar S, Mutti L and Landi S: Targeting hypoxic response for
cancer therapy. Oncotarget. 7:13464–13478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Witkiewicz AK, Whitaker-Menezes D,
Dasgupta A, Philp NJ, Lin Z, Gandara R, Sneddon S,
Martinez-Outschoorn UE, Sotgia F and Lisanti MP: Using the 'reverse
Warburg effect' to identify high-risk breast cancer patients:
Stromal MCT4 predicts poor clinical outcome in triple-negative
breast cancers. Cell Cycle. 11:1108–1117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bröer S, Rahman B, Pellegri G, Pellerin L,
Martin JL, Verleysdonk S, Hamprecht B and Magistretti PJ:
Comparison of lactate transport in astroglial cells and
monocarboxylate transporter 1 (MCT 1) expressing Xenopus laevis
oocytes. Expression of two different monocarboxylate transporters
in astroglial cells and neurons. J Biol Chem. 272:30096–30102.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pérez-Escuredo J, Van Hée VF, Sboarina M,
Falces J, Payen VL, Pellerin L and Sonveaux P: Monocarboxylate
transporters in the brain and in cancer. Biochim Biophys Acta.
1863:2481–2497. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Whitaker-Menezes D, Martinez-Outschoorn
UE, Lin Z, Ertel A, Flomenberg N, Witkiewicz AK, Birbe RC, Howell
A, Pavlides S, Gandara R, et al: Evidence for a stromal-epithelial
'lactate shuttle' in human tumors: MCT4 is a marker of oxidative
stress in cancer-associated fibroblasts. Cell Cycle. 10:1772–1783.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang G, Qian G and Cheng D: The effect of
monocarboxylate transporter gene on the regulation of pHi and
growth character in cancer cells. Zhonghua Jie He He Hu Xi Za Zhi.
24:666–670. 2001.In Chinese.
|
|
8
|
Baba M, Inoue M, Itoh K and Nishizawa Y:
Blocking CD147 induces cell death in cancer cells through
impairment of glycolytic energy metabolism. Biochem Biophys Res
Commun. 374:111–116. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
O'Callaghan K, Palagano E, Butini S,
Campiani G, Williams DC, Zisterer DM and O'Sullivan J: Induction of
apoptosis in oral squamous carcinoma cells by
pyrrolo-1,5-benzoxazepines. Mol Med Rep. 12:3748–3754. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu T, Hong Y, Jia L, Wu J, Xia J, Wang J,
Hu Q and Cheng B: Modulation of IL-1β reprogrammes the tumor
microenvironment to interrupt oral carcinogenesis. Sci Rep.
6:202082016. View Article : Google Scholar
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
12
|
Wang SN, Xie GP, Qin CH, Chen YR, Zhang
KR, Li X, Wu Q, Dong WQ, Yang J and Yu B: Aucubin prevents
interleukin-1 beta induced inflammation and cartilage matrix
degradation via inhibition of NF-κB signaling pathway in rat
articular chondrocytes. Int Immunopharmacol. 24:408–415. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shen XJ, Zhang H, Tang GS, Wang XD, Zheng
R, Wang Y, Zhu Y, Xue XC and Bi JW: Caveolin-1 is a modulator of
fibroblast activation and a potential biomarker for gastric cancer.
Int J Biol Sci. 11:370–379. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Al-Rakan MA, Colak D, Hendrayani SF,
Al-Bakheet A, Al-Mohanna FH, Kaya N, Al-Malik O and Aboussekhra A:
Breast stromal fibroblasts from histologically normal surgical
margins are pro-carcinogenic. J Pathol. 231:457–465. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mundim FG, Pasini FS, Nonogaki S, Rocha
RM, Soares FA, Brentani MM and Logullo AF: Breast
carcinoma-associated fibroblasts share similar biomarker profiles
in matched lymph node metastasis. Appl Immunohistochem Mol Morphol.
24:712–720. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang D, Gao J, Wang S, Ye N, Chong Y,
Huang Y, Wang J, Li B, Yin W and Wang D: Cancer-associated
fibroblasts promote angiogenesis in gastric cancer through
galectin-1 expression. Tumour Biol. 37:1889–1899. 2016. View Article : Google Scholar
|
|
17
|
Pavlides S, Whitaker-Menezes D,
Castello-Cros R, Flomenberg N, Witkiewicz AK, Frank PG, Casimiro
MC, Wang C, Fortina P, Addya S, et al: The reverse Warburg effect:
Aerobic glycolysis in cancer associated fibroblasts and the tumor
stroma. Cell Cycle. 8:3984–4001. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bonuccelli G, Whitaker-Menezes D,
Castello-Cros R, Pavlides S, Pestell RG, Fatatis A, Witkiewicz AK,
Vander Heiden MG, Migneco G, Chiavarina B, et al: The reverse
Warburg effect: Glycolysis inhibitors prevent the tumor promoting
effects of caveolin-1 deficient cancer associated fibroblasts. Cell
Cycle. 9:1960–1971. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pavlides S, Vera I, Gandara R, Sneddon S,
Pestell RG, Mercier I, Martinez-Outschoorn UE, Whitaker-Menezes D,
Howell A, Sotgia F, et al: Warburg meets autophagy:
Cancer-associated fibroblasts accelerate tumor growth and
metastasis via oxidative stress, mitophagy, and aerobic glycolysis.
Antioxid Redox Signal. 16:1264–1284. 2012. View Article : Google Scholar :
|
|
20
|
Capparelli C, Guido C, Whitaker-Menezes D,
Bonuccelli G, Balliet R, Pestell TG, Goldberg AF, Pestell RG,
Howell A, Sneddon S, et al: Autophagy and senescence in
cancer-associated fibroblasts metabolically supports tumor growth
and metastasis via glycolysis and ketone production. Cell Cycle.
11:2285–2302. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kamatani T, Shiogama S, Yoshihama Y, Kondo
S, Shirota T and Shintani S: Interleukin-1 beta in unstimulated
whole saliva is a potential biomarker for oral squamous cell
carcinoma. Cytokine. 64:497–502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Venturelli L, Nappini S, Bulfoni M,
Gianfranceschi G, Dal Zilio S, Coceano G, Del Ben F, Turetta M,
Scoles G, Vaccari L, et al: Glucose is a key driver for
GLUT1-mediated nanoparticles internalization in breast cancer
cells. Sci Rep. 6:216292016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brito AF, Abrantes AM, Ribeiro M, Oliveira
R, Casalta-Lopes J, Gonçalves AC, Sarmento-Ribeiro AB, Tralhão JG
and Botelho MF: Fluorine-18 fluorodeoxyglucose uptake in
hepatocellular carcinoma: Correlation with glucose transporters and
53 expression. J Clin Exp Hepatol. 5:183–189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma Y, Yu C, Mohamed EM, Shao H, Wang L,
Sundaresan G, Zweit J, Idowu M and Fang X: A causal link from ALK
to hexokinase II overexpression and hyperactive glycolysis in
EML4-ALK-positive lung cancer. Oncogene. 35:6132–6142. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Masoud GN and Li W: HIF-1α pathway: Role,
regulation and intervention for cancer therapy. Acta Pharm Sin B.
5:378–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi SY, Xue H, Wu R, Fazli L, Lin D,
Collins CC, Gleave ME, Gout PW and Wang Y: The MCT4 gene: A novel,
potential target for therapy of advanced prostate cancer. Clin
Cancer Res. 22:2721–2733. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lisanti MP, Sotgia F, Pestell RG, Howell A
and Martinez-Outschoorn UE: Stromal glycolysis and MCT4 are
hallmarks of DCIS progression to invasive breast cancer. Cell
Cycle. 12:2935–2936. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Andersen S, Solstad Ø, Moi L, Donnem T,
Eilertsen M, Nordby Y, Ness N, Richardsen E, Busund LT and Bremnes
RM: Organized metabolic crime in prostate cancer: The coexpression
of MCT1 in tumor and MCT4 in stroma is an independent
prognosticator for biochemical failure. Urol Onco. 33:338.e339–317.
2015.
|
|
29
|
Pértega-Gomes N, Vizcaíno JR, Attig J,
Jurmeister S, Lopes C and Baltazar F: A lactate shuttle system
between tumour and stromal cells is associated with poor prognosis
in prostate cancer. BMC Cancer. 14:3522014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Martinez-Outschoorn UE, Trimmer C, Lin Z,
Whitaker-Menezes D, Chiavarina B, Zhou J, Wang C, Pavlides S,
Martinez-Cantarin MP, Capozza F, et al: Autophagy in cancer
associated fibroblasts promotes tumor cell survival: Role of
hypoxia, HIF1 induction and NFκB activation in the tumor stromal
microenvironment. Cell Cycle. 9:3515–3533. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vallejo S, Palacios E, Romacho T,
Villalobos L, Peiró C and Sánchez-Ferrer CF: The interleukin-1
receptor antagonist anakinra improves endothelial dysfunction in
streptozotocin-induced diabetic rats. Cardiovasc Diabetol.
13:1582014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vecile E, Dobrina A, Salloum FN, Van
Tassell BW, Falcione A, Gustini E, Secchiero S, Crovella S, Sinagra
G, Finato N, et al: Intracellular function of interleukin-1
receptor antagonist in ischemic cardiomyocytes. PLoS One.
8:e532652013. View Article : Google Scholar : PubMed/NCBI
|