Introduction
Genome-wide association studies (GWAS) in large
cohorts with varying genetic backgrounds (1–3)
have revealed that common variants of melatonin receptor 1B
(MTNR1B), the gene encoding melatonin receptor 2 (MT2) have a high
and reproducible association with a higher risk of impaired insulin
secretion and increased fasting glucose levels. Particularly, when
compared with 43 other glycemia-associated genetic loci, the MTNR1B
variant appears to carry the strongest effect on diminished
glucose-stimulated insulin secretion (GSIS) in isolated human
islets (3). The present research
group demonstrated for the first time that the MTNR1B
rs10830963(G/C) variant is strongly associated with type 2 diabetes
mellitus (T2DM) in Han Chinese individuals (4). Therefore, the MTNR1B gene is widely
accepted as a diabetes risk gene.
Melatonin (5-methoxy-N-acetyltryptamine) is a
hormone synthesized by the circadian system (5) that provides timing cues to tissues
expressing melatonin receptors (6). The endocrine pancreatic islet is
known as an important melatonin target tissue that expresses
melatonin receptors MT1 and MT2 in rodents and humans (7,8).
It has been shown that melatonin and insulin secretion have an
inverse association (9). Rats and
patients with T2DM exhibit decreased melatonin levels and slightly
increased insulin levels (10,11)
It has been reported that growth factors, nutrients
and hormones, including insulin-like growth factor-1, incretins,
glucose, triiodothyronine, prolactin and insulin, require
activation of extracellular signal-regulated kinase (ERK)1/2 and
the phosphoinositide 3-kinase (PI3K)/Akt/mechanistic target of
rapamycin signaling pathways to fully induce rodent β-cell
replication (12,13). ERK1/2 are important for insulin
gene transcription in pancreatic β-cells, which produce insulin in
response to increases in the levels of circulating glucose in order
to enable efficient glucose utilization and storage (14). Various factors, including
Raf-1(15), Src and
sedoheptulokinase(16), have been
indicated to induce ERK1/2 to activate a downstream effect, among
which Raf-1 is one of the potential activators of ERK1/2.
The Raf/mitogen-activated protein kinase (MAPK)/ERK
arm of the insulin signaling pathway serves critical roles in
β-cell survival and proliferation (17). Alejandro et al (15) demonstrated that low doses of
insulin rapidly activated Raf-1 in human islets and MIN6 cells, and
revealed that the phosphorylation of ERK by insulin was eliminated
by exposure to a Raf inhibitor (GW5074) or transfection with a
dominant-negative Raf-1 mutant. A study conducted by Pardo et
al (18) revealed that adult
Raf-1 kinase inhibitor protein 1 knockout mice exhibit a rapid
reversal of streptozotocin-induced diabetes compared with control
mice. Taking the aforementioned findings into consideration, it is
unclear whether Raf-1/ERK pathway activation is affected by
melatonin and its receptor MTNR1B in pancreatic β-cells, and
consequently influences insulin synthesis and secretion.
Therefore, the present study explored the role of
melatonin and its receptor in the regulation of the Raf-1/ERK
pathway as well as insulin synthesis and secretion in MIN6 mouse
insulinoma cells.
Materials and methods
Cell culture
MIN6 cells, a mouse pancreatic β-cell line, were
generously provided by Dr Yang of Joslin Diabetes Center (Harvard
Medical School, Boston, MA, USA). The MIN6 cells were cultured in
Dulbecco's modified Eagle's medium (DMEM, containing 25 mM glucose,
4.0 mM L-glutamine and 1.0 mM sodium pyruvate; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) 15% fetal bovine serum
(FBS; Invitrogen; Thermo Fisher Scientific, Inc.), 2.5 mM
β-mercaptoethanol, 100 U/ml penicillin and 100 µg/ml
streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at 37°C in 5%
CO2. When the cells reached 65% attachment, they were
synchronized using serum-free medium for 8 h. Insulin
concentrations were measured in the supernatants following various
incubation experiments via ELISA (cat. no. 90080; Crystal Chem,
Inc., Downers Grove, IL, USA).
Chemicals and reagents
Antibodies targeting the following proteins were
obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA):
Phospho-Src (Tyr416; cat. no. 6943), total-Src (cat. no. 2108),
phospho-Raf-1 (Ser338; cat. no. 9427), phospho-Raf-1 (Ser259; cat.
no. 9421), total-Raf-1 (cat. no. 9422), phospho-ERK (cat. no. 4370)
and β-actin (cat. no. 4970). Antibodies targeting MTNR1B (cat. no.
ABT1890), ERK inhibitor (U0126) and GW5074 were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The MIN6 cells were exposed to melatonin at
concentrations of 0.01, 0.1, 1, 10 and 100 µM for 3 or 6 h.
In subsequent inhibition experiments, 10 µM U0126, 5
µM GW5074 or 0.1% dimethylsulfoxide control was added to the
medium 30 min prior to cell stimulation with melatonin. Following
the treatments, total cellular RNA was extracted from the cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
The samples were treated with reverse transcriptase kit (cat. no.
4374966; Thermo Fisher Scientific, Inc.) to synthesize cDNA. The
reverse transcription was performed sequentially as follows: 25°C
for 10 min, 37°C for 120 min, 85°C for 5 min. qPCR was then
performed with SYBR-Green Master mix on an ABI 7000 sequence
detection system (both Applied Biosystems; Thermo Fisher
Scientific, Inc.). The mRNA levels of the mice genes, insulin 1
(Ins1) and insulin 2 (Ins2), were normalized according to the
levels of GAPDH. Primer sequences were as follows: Ins1 forward,
5′-CGTGTAAATGCCACTGAAGC-3′ and reverse,
5′-CGGATGGACTGTTTGTAACCT-3′; Ins2 forward
5′-CTTCAGCCCCTCTGGCCATC-3′ and reverse,
5′-GAAACAATGACCTGCTTGCTGAT-3′; GAPDH forward,
5′-AACTTTGGCATTGTGGAAGG-3′ and reverse, 5′-ACACATTGGGGGTAGGAACA-3′.
PCR cycling conditions were as follows: Pre-incubation at 95°C for
10 min, followed by 40 cycles of denaturation for 15 sec at 95°C,
annealing for 60 sec at 60°C, and extension for 60 sec at 72°C; and
finally, termination for 8 min at 72°C. The expression levels of
target genes were quantified according to the 2−ΔΔCq
method as previously described (19).
Western blot analysis
Following the various treatments, cells were
harvested with radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology, Jiangsu, China) at 4°C and
lysates were centrifuged at 12,000 × g for 10 min. 5X sodium
dodecyl sulfate (SDS) loading buffer (Beyotime Institute of
Biotechnology) was added to the supernatant prior to denaturation
at 100°C for 10 min. A BCA kit (Beyotime Institute of
Biotechnology) was used to determine the protein concentration in
the supernatant.
Following this, the protein extracts were separated
(50 µg each sample) using 10% SDS-PAGE and transferred onto
a polyvinylidene difluoride membrane. The membranes were blocked in
Tris-buffered saline containing 0.05% Triton X-100 and 4% (w/v)
milk for 2 h at room temperature. The membrane was incubated for 2
h at room temperature with anti-β-actin (dilution, 1:5,000)
antibody or overnight at 4°C with the other primary antibodies. The
membranes were then incubated with secondary antibodies (cat. no.
sc2955; dilution, 1:5,000; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) for 1 h at room temperature. Visualization using
enhanced chemiluminescence (Bio-Rad laboratories, Inc., Hercules,
CA, USA) was performed. Protein expression levels were quantified
by scanning the immunostaining bands and analyzing using LabWork
4.5 image analysis software (UVP, LLC, Upland, CA, USA). Each
experiment was performed at least three times
Plasmid package and transfection
To overexpress MTNR1B in MIN6 cells, the plasmid
GV341-MTNR1B was constructed. The cDNA sequence of human MTNR1B was
amplified and sub-cloned into the AgeI and NheI sites
of the GV341 vector (Genomeditech, Shanghai, China) using the
following primers: Forward 5′-TCCGGAACGCAGGTAATTTGT-3′ and reverse
5′-GCCCAGCCGTCATAGAAGAT-3′. The primers were synthesized by
Shanghai GeneChem Co., Ltd. (Shanghai, China). For short hairpin
RNA (shRNA) vector construction, pairs of complementary
oligonucleotides interfering with MTNR1B (shMTNR1B-1, shMTNR1B-2,
and shMTNR1B-3) were synthesized by Shanghai GeneChem Co., Ltd.,
annealed, and ligated into vector GV248 (Genomeditech). The
targeting sequences of MTNR1B were as follows: shMTNR1B-1,
AGCTACTTACTGGCTTATT; shMTNR1B-2, AACCATGTTTGTGGTGTTT; and
shMTNR1B-3, GAGCTTTCTAACCATGTTT. A non-targeting scrambled siRNA
sequence (TTCTCCGAACGTGTCACGT) was used as a control.
For plasmid transfection, MIN6 cells were seeded
into a six-well plate with a density of 5×105
cells/well. After 24 h when the cells were 60-70% confluent, the
cells were trans-fected with shMTNR1B-1 (50 nM), shMTNR1B-2 (50
nM), shMTNR1B-3 (50 nM) or negative control in serum-free medium
using lipofectamine 3000™ (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Following incubation for
12 h at 37°C, the medium in each well was then replaced with DMEM,
containing 15% heat-inactivated FBS for another 48 h. Following
transfection, green fluorescence was used to observe the
transduction efficiency.
GSIS in MIN6 cells in vitro
The MIN6 cells were starved with DMEM (0.1% BSA and
3 mM glucose) overnight. Next, the cells were starved again with
Krebs-Ringer Bicarbonate buffer containing 125 mM NaCl, 4.74 mM
KCl, 1 mM CaCl2, 1.2 mM KH2PO4,
1.2 mM MgSO4, 5 mM NaHCO3, 25 mM HEPES (pH
7.4) and 3 mM glucose for 1 h. The medium was then changed to KRB
with either 3 or 25 mM glucose. The medium was collected after 1 h,
and the insulin concentration was measured using the aforementioned
ELISA kit.
Statistical analysis
For statistical evaluation and the significance
testing of differences, the results are expressed as the mean ±
standard error of the mean. The nonparametric Mann- Whitney U test
was performed with GraphPad software (version 6.0c; GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference. Each experiment
was performed for more than 3 times.
Results
Exogenous melatonin inhibits the
expression of insulin mRNA in MIN6 cells
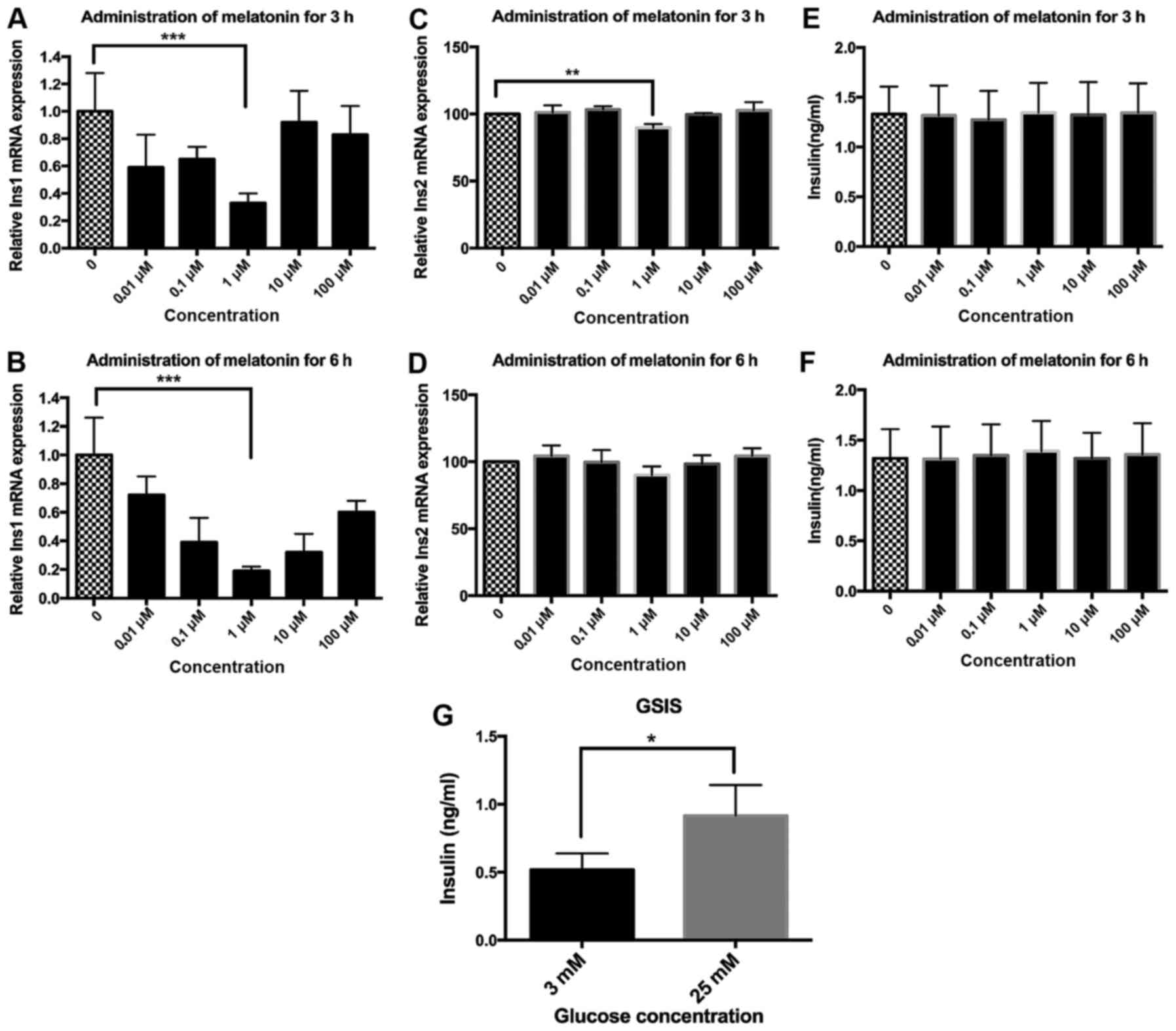
To characterize the influence of melatonin on
insulin gene expression, MIN6 cells were treated with 0.01, 0.1, 1,
10 or 100 µM melatonin for 3 or 6 h. Firstly, the mRNA
levels of Ins1 were evaluated. Melatonin inhibited Ins1 gene
expression potently and more strongly at the 6-h time point
compared with the 3-h time point (Fig. 1A and B). Melatonin decreased Ins1
mRNA expression with a nadir at 1 µM in dose-response
experiments where Ins1 mRNA was significantly reduced by 67±7 and
81±3% at 3 and 6 h, respectively, compared with that in the
untreated control cells (Fig. 1A and
B). Secondly, the impact of melatonin on Ins2 expression was
investigated. Notably, melatonin was observed to exert much weaker
effects on the expression of Ins2 than on the expression of Ins1
(Fig. 1C and D). Melatonin
significantly decreased Ins2 mRNA expression by 12±1.6% when
applied at a concentration of 1 µM for 3 h when compared
with the untreated control (Fig.
1C). There was also a 10±3.8% reduction of Ins2 mRNA expression
following the 6-h treatment with 1 µM melatonin, but this
reduction was not significant (P=0.057; Fig. 1D).
In order to investigate the effects of melatonin on
the secretion of insulin by the MIN6 cells, the insulin
concentrations in the culture medium were measured using ELISA. The
results demonstrated that treatment with various concentrations of
melatonin for either 3 or 6 h had no effect on the quantity of
insulin secreted by the MIN6 cells (Fig. 1E and F). In some cases, MIN6 cells
exhibit weak or no responsiveness to stimuli such as glucose
(20). To validate the
responsiveness of the MIN6 cells to stimuli in the present study,
in vitro GSIS experiments were performed. The results
demonstrated that 25 mM glucose significantly increased insulin
secretion by 1.78-fold compared with 3 mM glucose (Fig. 1G), indicating the satisfactory
responsiveness of the MIN6 cells used in the present study to
stimuli.
Melatonin inhibits the activation of
Raf-1 in MIN6 cells
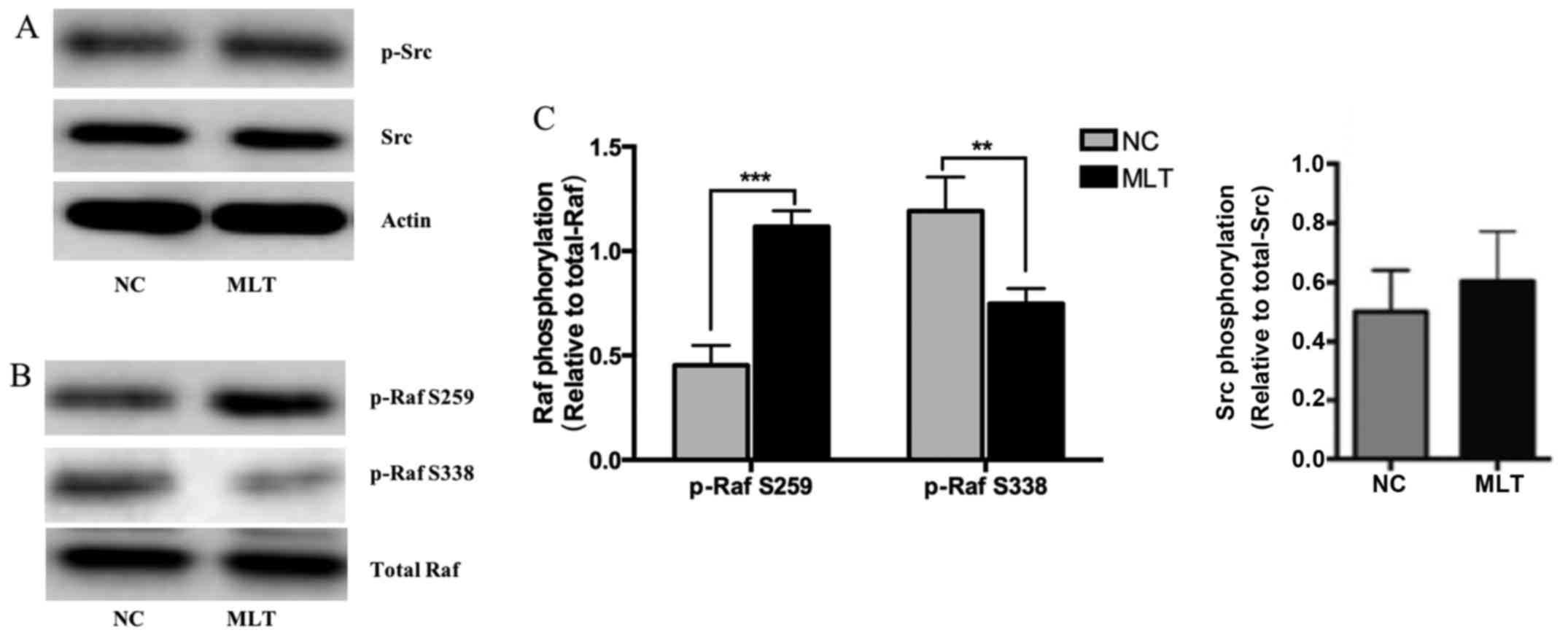
To investigate whether melatonin is capable of
regulating the MAPK signaling pathway in MIN6 cells, the activities
of two components of the pathway, namely Src and Raf-1 were
evaluated. Western blot analysis revealed that treatment with 1
µM melatonin for 3 h did not change the phosphorylation
levels of Src (Fig. 2A). A study
by Alejandro et al (15)
demonstrated that Raf-1 is activated by dephosphorylation at serine
259 and phosphorylation at serine 338 in human islets, mouse islets
and MIN6 cells. The phosphorylation levels at the Ser338 and Ser259
sites of Raf-1 were detected in the present study. The results
indicated that the phosphorylation of Raf-1 at the promotional site
Ser338 was significantly decreased by melatonin; the
phosphorylation of Raf-1 at the inhibitory site Ser259, however,
was significantly increased (Fig. 2B
and C), which suggests that the activation of Raf-1 was
repressed by the administration of melatonin.
Melatonin acts via MTNR1B and the
Raf-1/ERK signaling pathway
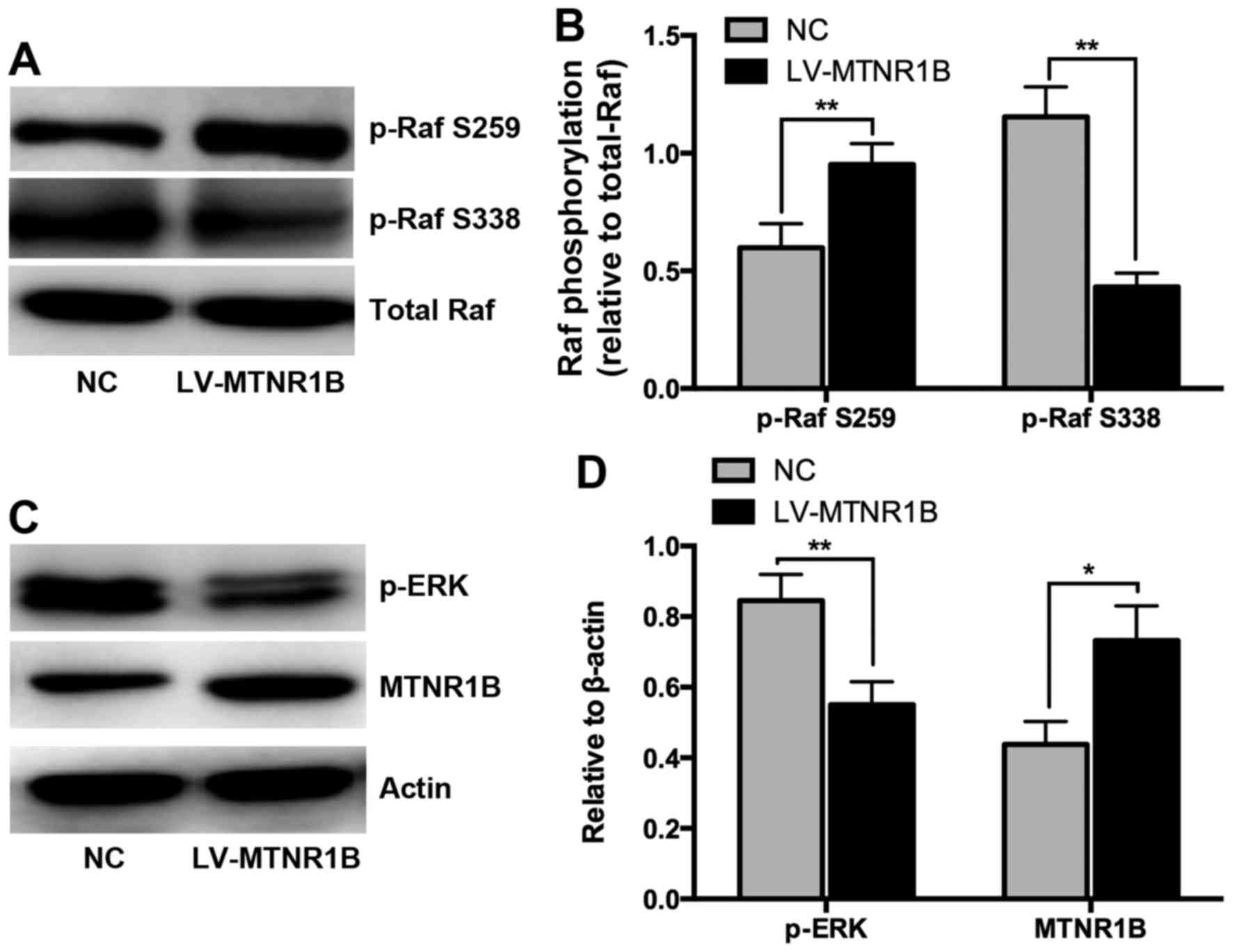
The aforementioned results show that melatonin
repressed the activation of Raf-1 (Fig. 2). In order to clarify whether
MTNR1B participates in the melatonin-induced regulation of Raf-1
activity, MTNR1B was overexpressed and silenced, respectively, in
MIN6 cells, and then the regulation of the activities of Raf-1 and
ERK by melatonin was investigated. MIN6 cells were transfected with
lentivirus overexpressing MTNR1B or shRNA targeting MTNR1B for 48
h, treated with 1 µM melatonin for 3 h, and then the levels
of phosphorylated Raf-1 and ERK were detected. The overexpression
of MTNR1B resulted in decreased phosphorylation of Raf-1 at Ser338
and increased phosphorylation at Ser259 (Fig. 3A and B), along with reduced
phosphorylation of ERK (Fig. 3C and
D), indicating that inactivation of the Raf-1/ERK signaling
pathway had occurred. By contrast, the knockdown of MTNR1B led to
the activation of Raf-1/ERK signaling due to increased
phosphorylation of Raf-1 at Ser338, decreased phosphorylation of
Raf-1 at Ser259 as well as increased phosphorylation of ERK
(Fig. 4A–D).
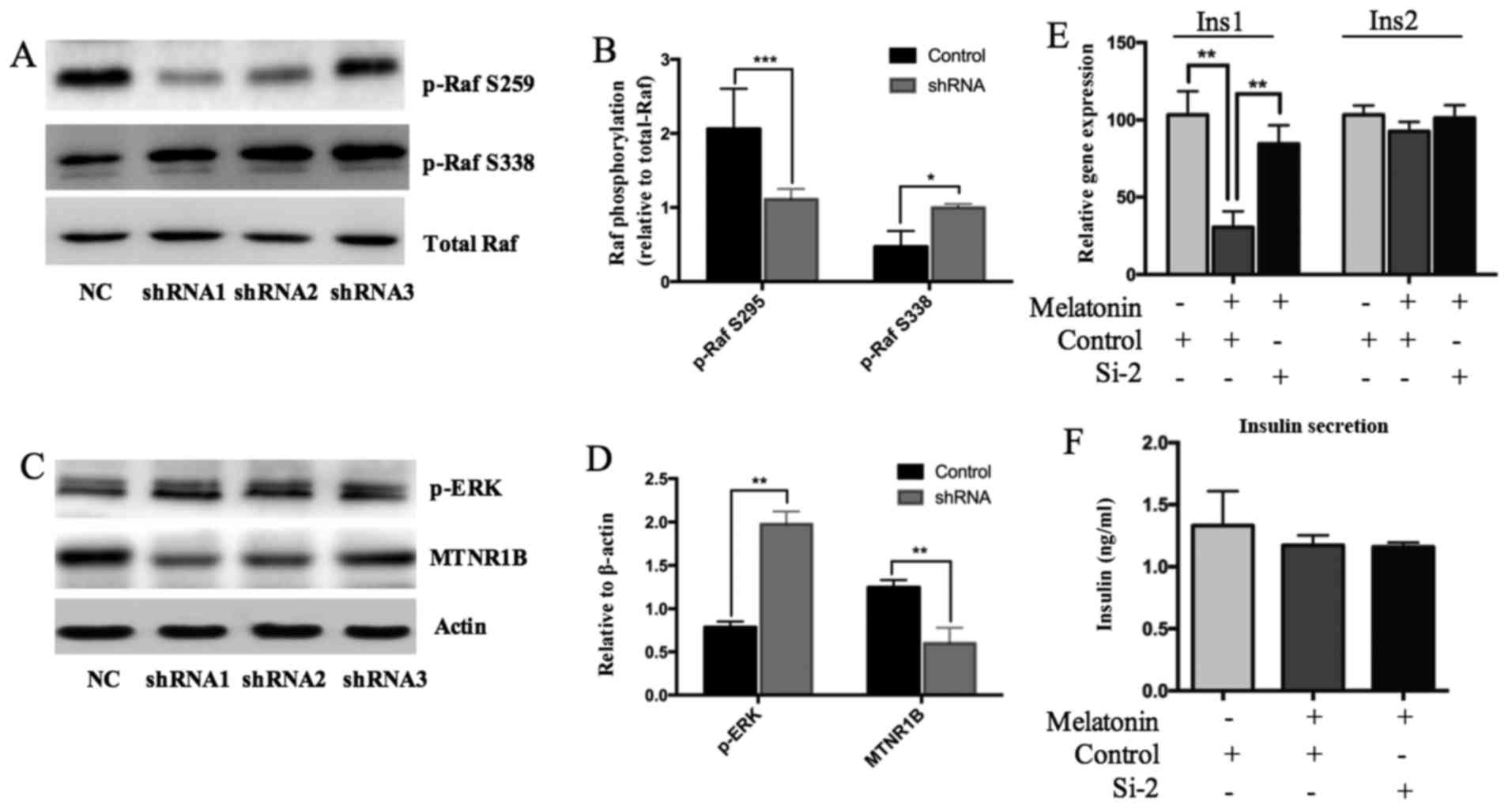 | Figure 4MIN6 cells were transfected with
MTNR1B shRNA and control shRNA for 48 h, and then the cells were
treated with 1 µM melatonin for an additional 3 h. (A and B)
Phosphorylation of Raf-1 and (C and D) ERK1/2 were investigated
using western blot analysis. (A) Representative western blots and
(B) quantification of Raf-1 phosphorylation; (C) representative
western blots and (D) quantification of ERK1/2 phosphorylation. (E)
Ins1 and Ins2 gene expression as well as (F) insulin secretion were
detected using reverse transcription-quantitative polymerase chain
reaction and ELISA, respectively. *P<0.05,
**P<0.01 and ***P<0.001, as indicated.
MTNR1B, melatonin receptor 1B; shRNA, short hairpin RNA; ERK,
extracellular signal-regulated kinase; p, phosphorylated; Ins1,
insulin 1; Ins2, insulin 2; NC, negative control. |
To determine whether MTNR1B was involved in the
melatonin-induced regulation of insulin expression, Ins1 and Ins2
gene expression was detected following the knockdown of MTNR1B
expression, and the insulin levels in the culture medium were also
measured. The data indicated that following the knockdown of
MTNR1B, the effects of melatonin on Ins1 and Ins2 expression were
totally attenuated (Fig. 4E),
suggesting that melatonin regulated insulin gene expression via
MTNR1B. Insulin secretion by the cells was not affected by the
manipulation of MTNR1B (Fig. 4F),
consistent with the observation that melatonin exerted no effects
on insulin secretion (Fig. 1E and
F).
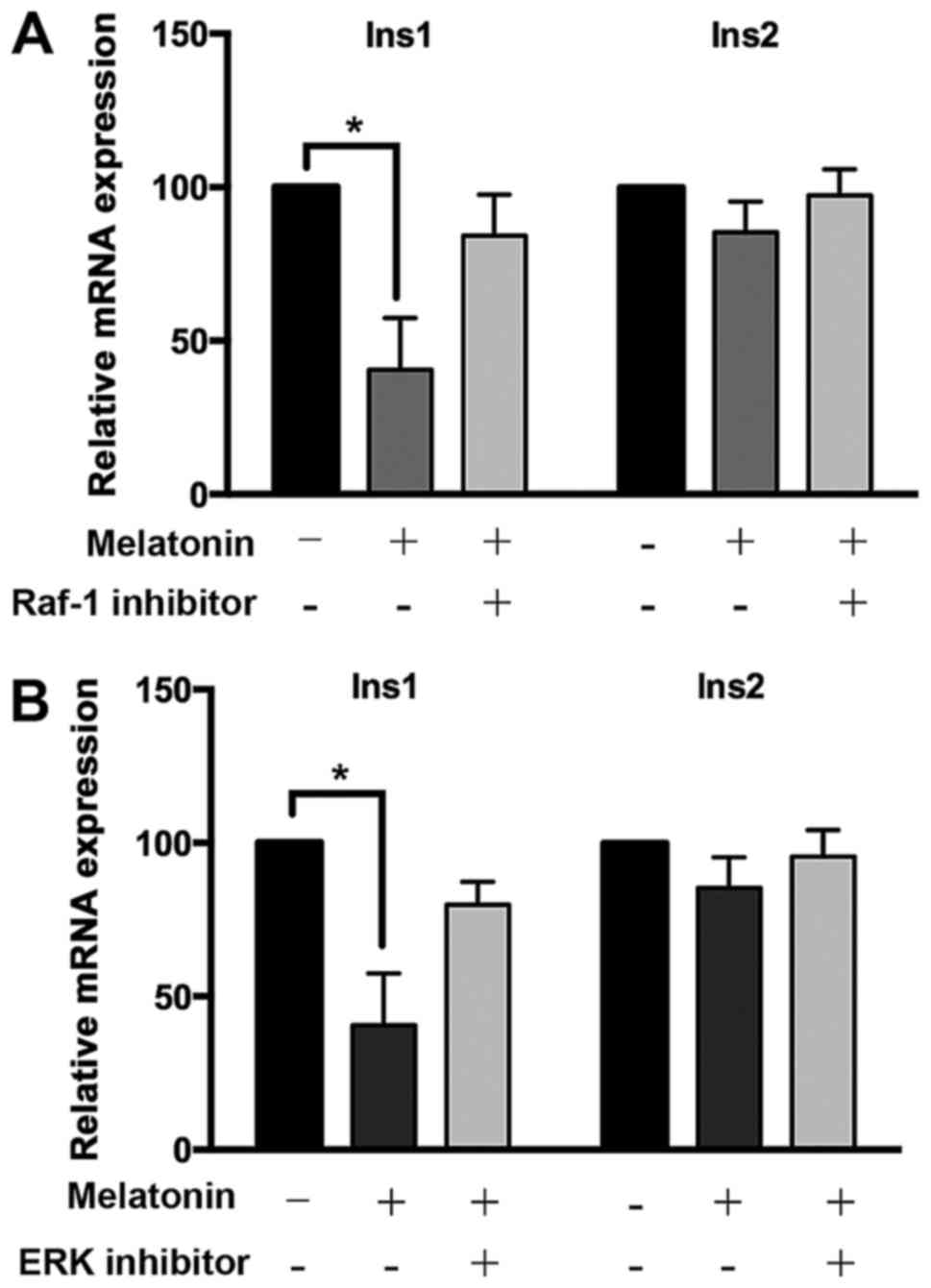
To identify whether melatonin exerted its effect on
insulin gene expression through the Raf-1/ERK signaling pathway,
the impact of melatonin on Ins1 and Ins2 expression was
investigated after blocking the activity of Raf-1 and ERK using the
chemical inhibitors GW5074 and U0126, respectively. Compared with
the blank group, melatonin significantly suppressed the expression
of Ins1 mRNA; however, when the activity of Raf-1 or ERK was
blocked, the effect of melatonin on Ins1 mRNA levels was markedly
attenuated (Fig. 5). The mRNA
levels of Ins2 were decreased weakly by melatonin administration,
and the two inhibitors slightly attenuated this effect of melatonin
(Fig. 5). These data indicate
that melatonin regulates the mRNA levels of Ins1 and Ins2 via the
Raf-1/ERK signaling pathway.
Discussion
The study of melatonin-insulin interactions has
revealed an inverse association between these two hormones
(21). As previously mentioned,
type 2 diabetic rats and humans exhibit increased plasma levels of
insulin, and decreased melatonin levels (10,11). Type 1 diabetic rats exhibit
extremely reduced levels or the absence of insulin, but
statistically significant increases in melatonin levels (22). These results are in agreement with
observations that the pinealectomy of rodents caused
hyperinsulinemia (23), and that
following perfusion with melatonin, stimulated insulin secretion
was inhibited in the pancreatic islets of normoglycemic Wistar and
type 2 diabetic Goto-Kakizaki rats (24). Ramracheya et al (25) observed that melatonin inhibits
glucose-stimulated insulin release from MIN6 cells. In the present
study, the effects of melatonin on the expression of Ins1 and Ins2
genes in MIN6 cells were investigated and the results revealed that
melatonin exerted a potent inhibitory effect on Ins1 mRNA, but a
much weaker one on Ins2 mRNA expression. The treatment of MIN6
cells with 1 µM melatonin for 6 h reduced Ins1 mRNA
expression by 81±3%. The maximum effect of melatonin on Ins2 mRNA
expression was observed following incubation with 1 µM
melatonin for 3 h, with a reduction of 12±1.6%. The two mouse
preproinsulin genes are located at different chromosomes, at
chromosome 19 for Ins1 and chromosome 7 for Ins2, respectively
(26). Regarding the disparity of
the response to melatonin between Ins1 and Ins2, it may be
speculated that melatonin preferentially regulated Ins1 expression.
In addition to the expression of insulin at the genetic level, the
regulation of the secretion of insulin by melatonin was also
investigated in the present study. Notably, the results
demonstrated that the treatment of MIN6 cells with different
concentrations of melatonin for 3 or 6 h had no effect on the
amount of insulin secreted by the cells. However, when the MIN6
cells were cultured in DMEM with 25 mM glucose, the high
concentration of glucose potently stimulated insulin secretion. It
may be assumed that the high concentration of glucose came up with
a high threshold for insulin secretion to other stimuli, which may
have resulted in the MIN6 cells being unresponsive to melatonin.
Previous studies concerning the impact of melatonin on insulin
secretion are conflicting. Although the inhibitory effect of
melatonin is predominant (27,28), there are studies indicating that
melatonin has neutral or stimulatory effects on insulin secretion
(29,30).
GWAS have revealed that the gene for the MT2
receptor is a locus with a high and reproducible association with a
higher risk of impaired insulin secretion and increased fasting
glucose levels (2,3). The transcriptional and protein
levels of MT1 and MT2 have been demonstrated to be significantly
higher in the pancreatic tissues from patients with T2DM compared
with those from metabolically healthy controls (31). These results are in accordance
with the detection of increased MT2 receptor mRNA expression in the
islets of individuals carrying the T2DM risk allele (3). A European cohort study (32) identified that rare MTNR1B variants
causing impairment of the function of MTNR1B contributed to type 2
diabetes. It may be hypothesized that melatonin, when combined with
the functional impairment of MTNR1B, would exert a more pronounced
effect on the increased risk of T2DM.
The mechanism by which melatonin affects the
stimulation of insulin secretion through MT2 is unclear. According
to Picinato et al (33),
melatonin regulates the growth and differentiation of pancreatic
cells by activating two intracellular signaling pathways: PI3K/Akt
and MEK/ERK. Therefore, the impact of melatonin on the MAPK
signaling pathway was investigated in the present study.
MT receptors have been reported to couple with
Giα-proteins, which inhibit the production of cyclic adenosine
monophosphate (cAMP) (34).
Melatonin has been shown to reduce the production cAMP in INS1
cells, resulting in diminished insulin release (30). It is well established that cAMP
and its principal target, the cAMP-dependent protein kinase (PKA),
are extensively involved in the impact of hormones on metabolic
pathways, as well as cell growth and proliferation (35). MAPK, also known as ERK, is a
target of cAMP that is activated or inhibited by cAMP in a cell-
specific manner (35). ERKs are
intracellular signaling molecules involved in the regulation of
cell proliferation and other cellular functions (36). It has been reported that ERK1/2
are required for the stimulatory effect of glucose on insulin gene
transcription (14). To elucidate
the mechanism underlying the impact of melatonin on insulin
transcription and secretion, the regulation of the MAPK signaling
pathway by melatonin and its receptor MTNR1B was investigated. The
Src/Ras-related protein 1/B-Raf and Ras/Raf-1 pathways are two
upstream cascades converging on ERKs (35). Insulin secretion has been shown to
be significantly reduced in mouse β-cell Raf-1-knockout islets
(37). An association of Src
kinases with insulin secretion has also been reported (16). Therefore, whether the activity of
Src and/or Raf-1 is regulated by melatonin was investigated in the
present study. The data demonstrated that the phosphorylation of
Src was not affected by melatonin, whereas Raf-1 was strongly
inactivated by the administration of melatonin to MIN6 cells,
implying that the effect of melatonin on the MAPK pathway was
mediated by Raf-1, not via Src. In a study by Kowluru et al
(38), the overexpression of
Raf-1 led to a significant glucose-mediated activation of ERK1/2 in
INS 832/13 cells, and the pharmacological inhibition of Raf-1
kinase markedly reduced the stimulatory effects of glucose on
ERK1/2 phosphorylation and insulin secretion. In the present study,
it was observed that following the inhibition of the activity of
Raf-1 or ERK using chemical inhibitors, the effects of melatonin on
the mRNA expression of Ins1 and Ins2 in MIN6 cells were attenuated,
suggesting the involvement of the Raf-1/ERK pathway in
melatonin-regulated insulin gene transcription. Since the majority
of melatonin's functions are fulfilled through its receptors
(39), it was next determined
whether the MT2 receptor participated in the melatonin-induced
regulation of Raf-1/ERK signaling. It was observed that the
overexpression of MTNR1B significantly decreased Raf-1 and ERK
activities in the melatonin-treated MIN6 cells, whereas the
knockdown of MTNR1B resulted in significant activation of Raf-1/ERK
signaling. Notably, following MTNR1B silencing, the effect of
melatonin on insulin gene transcription was totally attenuated.
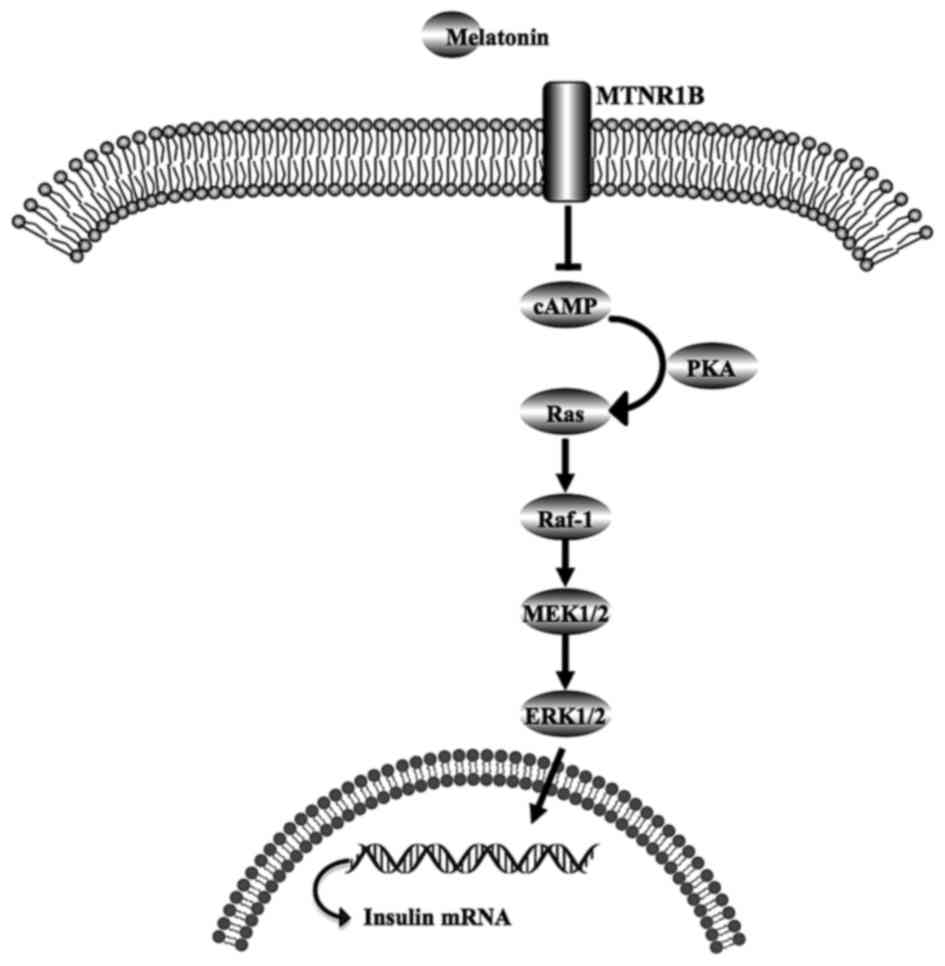
Based on all the data in the present study, a melatonin/MT2
receptor/Raf-1/ERK signaling pathway for the regulation of insulin
gene expression in MIN6 cells is proposed, which is illustrated in
Fig. 6. In this, melatonin binds
to the MT2 receptor, which reduces cAMP production and PKA
inhibition, in turn inactivating the Ras/Raf-1/ERK pathway, and
eventually decreasing insulin gene transcription.
In conclusion, the present study provides evidence
that melatonin inhibits insulin gene expression in MIN6 mouse
pancreatic β-cells, but has no effect on insulin secretion. The
present data also indicate for the first time that melatonin, via
its receptor MT2, exerts an inhibitory effect on the Raf-1/ERK
pathway in MIN6 cells.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81270903
to Jie Wen and 81370936 to Xuanchun Wang) and the Natural Science
Foundation of Shanghai City (grant no. 10ZR1405500 to Jie Wen). The
study was also sponsored by grants from Shanghai Pujiang Program
(grant no. 16PJ1401700 to Xuanchun Wang) and the Science and
Technology Commission of Shanghai Municipality (grant no.
16140901200 to Xuanchun Wang).
References
|
1
|
Peschke E, Bähr I and Mühlbauer E:
Experimental and clinical aspects of melatonin and clock genes in
diabetes. J Pineal Res. 59:1–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Simonis-Bik AM, Nijpels G, van Haeften TW,
Houwing-Duistermaat JJ, Boomsma DI, Reiling E, van Hove EC, Diamant
M, Kramer MH, Heine RJ, et al: Gene variants in the novel type 2
diabetes loci CDC123/CAMK1D, THADA, ADAMTS9, BCL11A, and MTNR1B
affect different aspects of pancreatic beta-cell function.
Diabetes. 59:293–301. 2010. View Article : Google Scholar
|
|
3
|
Lyssenko V, Nagorny CL, Erdos MR, Wierup
N, Jonsson A, Spégel P, Bugliani M, Saxena R, Fex M, Pulizzi N, et
al: Common variant in MTNR1B associated with increased risk of type
2 diabetes and impaired early insulin secretion. Nat Genet.
41:82–88. 2009. View
Article : Google Scholar
|
|
4
|
Rönn T, Wen J, Yang Z, Lu B, Du Y, Groop
L, Hu R and Ling C: A common variant in MTNR1B, encoding melatonin
receptor 1B, is associated with type 2 diabetes and fasting plasma
glucose in Han Chinese individuals. Diabetologia. 52:830–833. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stehle JH, Saade A, Rawashdeh O, Ackermann
K, Jilg A, Sebestény T and Maronde E: A survey of molecular details
in the human pineal gland in the light of phylogeny, structure,
function and chronobiological diseases. J Pineal Res. 51:17–43.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sharma S, Singh H, Ahmad N, Mishra P and
Tiwari A: The role of melatonin in diabetes: Therapeutic
implications. Arch Endocrinol Metab. 59:391–399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nagorny CL, Sathanoori R, Voss U, Mulder H
and Wierup N: Distribution of melatonin receptors in murine
pancreatic islets. J Pineal Res. 50:412–417. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mühlbauer E, Gross E, Labucay K, Wolgast S
and Peschke E: Loss of melatonin signalling and its impact on
circadian rhythms in mouse organs regulating blood glucose. Eur J
Pharmacol. 606:61–71. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bazwinsky-Wutschke I, Wolgast S, Mühlbauer
E, Albrecht E and Peschke E: Phosphorylation of cyclic AMP-response
element-binding protein (CREB) is influenced by melatonin treatment
in pancreatic rat insulinoma β-cells (INS-1). J Pineal Res.
53:344–357. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Peschke E, Hofmann K, Pönicke K, Wedekind
D and Mühlbauer E: Catecholamines are the key for explaining the
biological relevance of insulin-melatonin antagonisms in type 1 and
type 2 diabetes. J Pineal Res. 52:389–396. 2012. View Article : Google Scholar
|
|
11
|
Bach AG, Mühlbauer E and Peschke E:
Adrenoceptor expression and diurnal rhythms of melatonin and its
precursors in the pineal gland of type 2 diabetic goto-kakizaki
rats. Endocrinology. 151:2483–2493. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim TK, Lee JS, Jung HS, Ha TK, Kim SM,
Han N, Lee EJ, Kim TN, Kwon MJ, Lee SH, et al: Triiodothyronine
induces proliferation of pancreatic beta-cells through the MAPK/ERK
pathway. Exp Clin Endocrinol Diabetes. 122:240–245. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Beith JL, Alejandro EU and Johnson JD:
Insulin stimulates primary beta-cell proliferation via Raf-1
kinase. Endocrinology. 149:2251–2260. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lawrence MC, Jivan A, Shao C, Duan L, Goad
D, Zaganjor E, Osborne J, McGlynn K, Stippec S, Earnest S, et al:
The roles of MAPKs in disease. Cell Res. 18:436–442. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Alejandro EU, Kalynyak TB, Taghizadeh F,
Gwiazda KS, Rawstron EK, Jacob KJ and Johnson JD: Acute insulin
signaling in pancreatic beta-cells is mediated by multiple Raf-1
dependent pathways. Endocrinology. 151:502–512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Q, Wang R, Zhou H, Zhang L, Cao Y,
Wang X and Hao Y: SHIP2 on pI3K/Akt pathway in palmitic acid
stimulated islet β cell. Int J Clin Exp Med. 8:3210–3218. 2015.
|
|
17
|
Stewart AF, Hussain MA, García-Ocaña A,
Vasavada RC, Bhushan A, Bernal-Mizrachi E and Kulkarni RN: Human
β-cell proliferation and intracellular signaling: Part 3. Diabetes.
64:1872–1885. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pardo FN, Altirriba J, Pradas-Juni M,
García A, Ahlgren U, Barberà A, Slebe JC, Yáñez AJ, Gomis R and
Gasa R: The role of Raf-1 kinase inhibitor protein in the
regulation of pancreatic beta cell proliferation in mice.
Diabetologia. 55:3331–3340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Cheng K, Delghingaro-Augusto V, Nolan CJ,
Turner N, Hallahan N, Andrikopoulos S and Gunton JE: High passage
MIN6 cells have impaired insulin secretion with impaired glucose
and lipid oxidation. PloS One. 7:e408682012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mühlbauer E, Albrecht E,
Bazwinsky-Wutschke I and Peschke E: Melatonin influences insulin
secretion primarily via MT(1) receptors in rat insulinoma cells
(INS-1) and mouse pancreatic islets. J Pineal Res. 52:446–459.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peschke E, Wolgast S, Bazwinsky I, Pönicke
K and Muhlbauer E: Increased melatonin synthesis in pineal glands
of rats in streptozotocin induced type 1 diabetes. J Pineal Res.
45:439–448. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nishida S, Segawa T, Murai I and Nakagawa
S: long-term melatonin administration reduces hyperinsulinemia and
improves the altered fatty-acid compositions in type 2 diabetic
rats via the restoration of Delta-5 desaturase activity. J Pineal
Res. 32:26–33. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peschke E, Schucht H and Mühlbauer E:
long-term enteral administration of melatonin reduces plasma
insulin and increases expression of pineal insulin receptors in
both Wistar and type 2-diabetic Goto-Kakizaki rats. J Pineal Res.
49:373–381. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ramracheya RD, Muller DS, Squires PE,
Brereton H, Sugden D, Huang GC, Amiel SA, Jones PM and Persaud SJ:
Function and expression of melatonin receptors on human pancreatic
islets. J Pineal Res. 44:273–279. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wentworth BM, Schaefer IM, Villa-Komaroff
L and Chirgwin JM: Characterization of the two nonallelic genes
encoding mouse preproinsulin. J Mol Evol. 23:305–312. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Peschke E, Peschke D, Hammer T and Csernus
V: Influence of melatonin and serotonin on glucose-stimulated
insulin release from perifused rat pancreatic islets in vitro. J
Pineal Res. 23:156–163. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Picinato MC, Haber EP, Cipolla-Neto J,
Curi R, de Oliveira Carvalho CR and Carpinelli AR: Melatonin
inhibits insulin secretion and decreases PKA levels without
interfering with glucose metabolism in rat pancreatic islets. J
Pineal Res. 33:156–160. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Frankel BJ and Strandberg MJ: Insulin
release from isolated mouse islets in vitro: No effect of
physiological levels of melatonin or arginine vasotocin. J Pineal
Res. 11:145–148. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Peschke E, Bach AG and Mühlbauer E:
Parallel signaling pathways of melatonin in the pancreatic
beta-cell. J Pineal Res. 40:184–191. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peschke E, Stumpf I, Bazwinsky I, Litvak
L, Dralle H and Mühlbauer E: Melatonin and type 2 diabetes - a
possible link? J Pineal Res. 42:350–358. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bonnefond A, Clément N, Fawcett K, Yengo
L, Vaillant E, Guillaume JL, Dechaume A, Payne F, Roussel R,
Czernichow S, et al: Rare MTNR1B variants impairing melatonin
receptor 1B function contribute to type 2 diabetes. Nat Genet.
44:297–301. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Picinato MC, Hirata AE, Cipolla-Neto J,
Curi R, Carvalho CR, Anhê GF and Carpinelli AR: Activation of
insulin and IGF-1 signaling pathways by melatonin through MT1
receptor in isolated rat pancreatic islets. J Pineal Res. 44:88–94.
2008.
|
|
34
|
Vanecek J: Cellular mechanisms of
melatonin action. Physiol Rev. 78:687–721. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stork PJ and Schmitt JM: Crosstalk between
cAMP and MAP kinase signaling in the regulation of cell
proliferation. Trends Cell Biol. 12:258–266. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pearson G, Robinson F, Beers Gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: Regulation and physiological functions.
Endocr Rev. 22:153–183. 2001.PubMed/NCBI
|
|
37
|
Alejandro EU, Lim GE, Mehran AE, Hu X,
Taghizadeh F, Pelipeychenko D, Baccarini M and Johnson JD:
Pancreatic β-cell Raf-1 is required for glucose tolerance, insulin
secretion, and insulin 2 transcription. FASEB J. 25:3884–3895.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kowluru A, Veluthakal R, Rhodes CJ, Kamath
V, Syed I and Koch BJ: Protein farnesylation-dependent
Raf/extracellular signal-related kinase signaling links to
cytoskeletal remodeling to facilitate glucose-induced insulin
secretion in pancreatic beta-cells. Diabetes. 59:967–977. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Boutin JA, Audinot V, Ferry G and
Delagrange P: Molecular tools to study melatonin pathways and
actions. Trends Pharmacol Sci. 26:412–419. 2005. View Article : Google Scholar : PubMed/NCBI
|