Introduction
Ovarian cancer comprises a large proportion of all
gynecological cancers, is associated with a high mortality rate,
and represents a major concern for women's health worldwide
(1–5). The currently applied treatments for
ovarian cancer mainly include radiotherapy, chemotherapy and/or
chemoradiotherapy combined with surgery. However, the lack of early
symptoms and effective biomarker screening make diagnosis
difficult, with >70% of the diagnosed patients presenting with
advanced epithelial ovarian carcinoma with distant metastases
(6–8). At present, the molecular
pathogenesis of ovarian cancer remains poorly understood. In
addition, the lack of diagnostic biomarkers and effective
therapeutic methods represents a major challenge in the management
of this disease. Therefore, the molecular mechanism underlying the
development of ovarian cancer must be urgently elucidated.
MicroRNAs (miRNAs), a type of endogenous, short and
non-coding RNAs consisting of ~20 nucleotides, are involved in
post-transcriptional regulation by targeting the 3′ untranslated
region (UTR) of target genes and affect biological processes
(6,9–13).
miRNAs have been found to act as a class of regulatory factors, and
are closely associated with the development and progression of
diverse diseases, such as cancer (7,11,14–17). Various miRNAs have been reported
to be involved in the development of ovarian cancer (18–25). The mechanism and function of
miR-152 have been investigated in gastrointestinal (26), endometrial (27), liver (28), prostate (29) and ovarian cancer (30). However, the functional relevance
of miR-152 and ERBB3 in ovarian cancer remains unknown.
The aim of the present study was to investigate
whether miR-152 downregulates ERBB3 in SKOV3 and OVCAR3 ovarian
cancer cells in vitro, suppressing their ability to
proliferate, migrate and invade and promoting their apoptosis, in
order to determine whether miR-152 is a potential therapeutic
target for ovarian cancer.
Materials and methods
Clinical specimens and cell lines
The clinical samples for the present study were
obtained from 36 patients with ovarian cancer in The Affiliated
Tumor Hospital of Harbin Medical University (Harbin, China) between
2015 and 2016. Informed consent was obtained from all patients and
the study protocol was approved by the Ethics Committee of The
Affiliated Tumor Hospital of Harbin Medical University. The
histological diagnosis of ovarian cancer was evaluated according to
the World Health Organization. The ovarian carcinoma cell lines
(SKOV3 and OVCAR3) were obtained from the American Type Culture
Collection (Manassas, VA, USA).
Transfection of miR-152 mimics
Transfection of miR-152 mimics was performed as
previously described (31). The
primer sequence of the negative control (NC) was
5′-UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT-3′. miR-152-mimics
with the following sequence were also designed:
5′-UCAGUGCAUGACAGAACUUGGAAGUUCUGUCAUGCACUGAUU-3′. The RNA
nucleotides were synthesized by GenePharma (Shanghai, China).
EERB3 transfection
According to the manuscript protocol, pcDNA3.1
expressing ERBB3 or control vector were transfected into SKOV3 and
OVCAR3 cells that had been transfected with miR-152-NC and
miR-152-mimics using the FuGENE6 (Roche Diagnostics, Mannheim,
Germany) reagent. Stable transfection cell lines were screened in
medium containing 1 mg/ml G418 (Sigma-Aldrich; Merck KGaA, St.
Louis, MO, USA).
Quantitative polymerase chain reaction
(qPCR)
As described previously (32), the SYBR-Green PCR Master Mix kit
(Takara, Dalian, China) was used to measure the mRNA expression
levels. The primer sequences for glyceraldehyde 3-phosphate
dehydrogenase (GAPDH), which was used as internal control, were as
follows: Forward, 5′-TGTTCGTCATGGGTGTGAAC-3′ and reverse,
5′-ATGGCATGGACTGTGGTCAT-3′; the primer sequences for ERBB3 were:
Forward, 5′-GCAGATCAGTGTGTAGCGTG-3′ and reverse,
5′-CGTGTGCAGTTGAAGTGACA-3′; the primer sequences for miR-152 were:
Forward, 5′-TCAGTGCATGACAGAACTTGGAA-3′ and reverse,
5′-GCTGTCAACGATACGCTACGT-3′; and the primer sequences for U6 were:
Forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. The Bio-Rad IQTM5 Multicolor Real-Time
PCR detection System (Bio-Rad, Hercules, CA, USA) was used for
qPCR. The PCR programs were set as follows: 95°C for 30 sec as the
first step in a loop; 95°C for 5 sec, 60°C for 34 sec as the second
step, a total of 40 cycles. GAPDH or U6 were used as the internal
reference, and the 2−ΔΔCt method was applied for
calculating expression (1).
Western blot analysis
Whole lysates of the treated SKOV3 and OVCAR3 cells
were prepared by scraping cells with RIPA buffer supplemented with
a protease inhibitor cocktail (P8340; Sigma-Aldrich; Merck KGaA),
then separated on 8% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis gels. The primary antibodies used in this study
were anti-ERBB3 (1:200; Abcam, Cambridge, CA, USA) and anti-GAPDH
(1:1,000; Abcam, Cambridge, UK).
Luciferase reporter assay
The binding sites between miRNAs and ERBB3 were
predicted by the TargetScan database (http://www.targetscan.org). SKOV3 and OVCAR3 cells
(5×104 cells/well) that were co-transfected with
wild-type or mutated ERBB3 and miR-152 or anti-miR-152,
respectively, were incubated in 24-well plates and co-transfected
with the constructed ERBB3 plasmids by Lipofectamine 2000 (using
Renilla plasmid as internal reference) for 48 h. According
to the manufacturer's instructions, the luciferase activities were
determined by the Dual-Luciferase Reporter assay kit (Promega,
Madison, WI, USA)
MTT and colony-forming unit (CFU)
assays
The MTT assay was used to determine the
proliferative ability of the cells. First, the treated SKOV3 and
OVCAR3 cells (3,000 cells/well) were prepared and seeded in 96-well
plates with the complete media. At 0, 12, 24 and 48 h, 20 μl
MTT solution (5 mg/ml) were added to each well. After 4 h of
incubation, the supernatant was discarded, and 100 μl
dimethyl sulfoxide solution was added to dissolve the crystals. A
microplate reader (BioTek Instruments, Inc., Winooski, VT, USA) was
used to detect the absorbance at 570 nm. The treated SKOV3 and
OVCAR3 cells were incubated for 10 days. Methanol was used to fix
the forming colonies and Giemsa was used to dye the colonies. The
CFUs in the treatment and the control groups were counted.
Flow cytometric analysis of cell
apoptosis
The Annexin V-FITC Apoptosis Detection kit II (BD
Pharmingen, San Diego, CA, USA) was used to detect apoptotic cells
according to the manufacturer's protocol. Briefly, the treated
SKOV3 and OVCAR3 cells were resuspended with binding buffer at a
density of 1×106 cells/ml. Annexin V-FITC (5 μl)
and propidium iodide (1 μg/ml) were used to stain the cells.
The flow cytometry results were analyzed by the FlowJo software
(Tree Star Corp., Ashland, OR, USA). Cells were grouped into the
viable, early-stage apoptotic, late-stage apoptotic and dead cell
groups.
Wound healing assay
As described previously (33), the treated SKOV3 and OVCAR3 cells
were incubated in 6-well plates. Small wounds were formed by
scratching a line of cells. The small fragments of cells were
gently washed with phosphate-buffered saline. After 24 and 72 h,
the wounds were photographed under a microscope. The distance of
cells from the edge of the scratch represented the degree of cell
migration.
Migration and invasion assays
According to the manufacturer's instructions, the
treated SKOV3 and OVCAR3 cells were starved overnight in serum-free
medium and resuspended at a density of 2.5×106 cells/ml
in serum-free medium. Treated cells (200 μl) were seeded
into the top chamber of the 8-μm pore cell culture insert,
and complete medium was added into the bottom chamber. After 24 h,
the upper chamber cells were removed using a cotton swab and the
migrating cells were fixed with 4% paraformaldehyde and stained
with 0.1% crystal violet solution. The migrating cells were counted
at 5 random fields. For the invasion assay, diluted Matrigel (BD
Biosciences, San Diego, CA, USA) was paved into the upper well of
the Transwell chamber (Corning, Inc., Corning, NY, USA) and
incubated for 2 h at 37°C.
Tumor formation in nude mice
This study was approved by the Institutional
Committee for Animal Research and completed in accordance with the
Institutional Animal Care and Use Committee. Treated SKOV3 cells
(100 μl 2×107/mouse) were subcutaneously injected
into the flanks of 5-week-old BALB/c male athymic nude mice for a
total of 5 times over 4 weeks. At the specified time, the treated
nude mice were sacrificed and the tumors were collected and
measured.
Statistical analysis
All data are presented as the mean ± standard
deviation from three independent experiments. P<0.05 indicates
statistically significant differences. Statistical significance was
analyzed using GraphPad (GraphPad Prism Software, La Jolla, CA,
USA) and the SPSS 20.0 software (variance and Student's
t-test).
Results
Screening of miRNAs that are closely
associated with ERBB3 in ovarian cancer
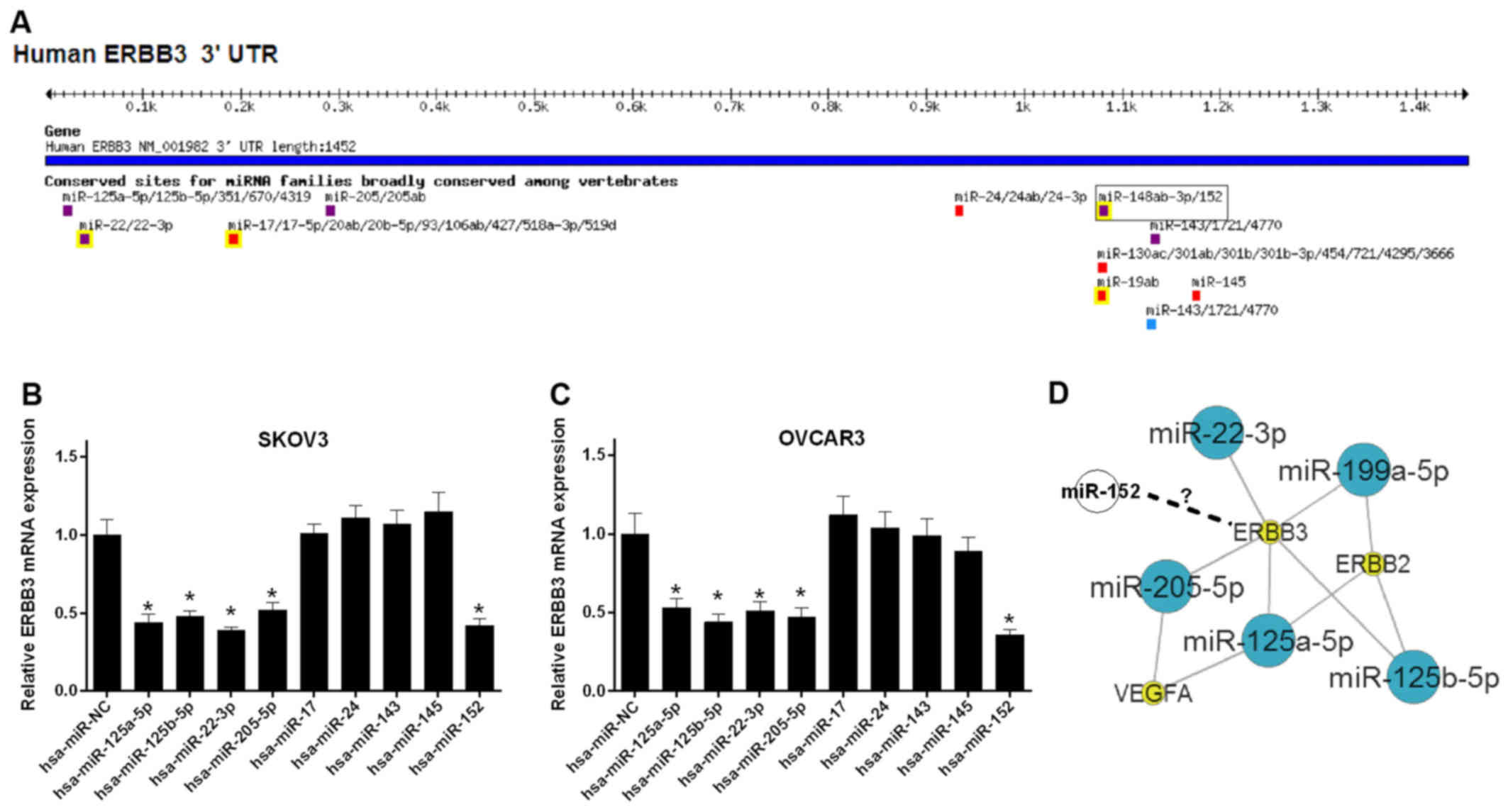
In order to study the regulatory mechanism of the
ERBB3 gene in ovarian cancer, the miRNA target sites in ERBB3 3′UTR
were predicted by TargetScan. We found multiple miRNAs displaying
target sites in ERBB3 3′UTR (Fig.
1A). Furthermore, the mRNA expression levels of ERBB3 were
detected in SKOV3 cells transfected with miRNAs. The results
revealed that the miR-125a-5p, miR-125b-5p, miR-22-3p, miR-205-5p
and miR-152-transfected SKOV3 cells have a significantly lower
level of ERBB3 mRNA (P<0.05; Fig.
1B). A similar tendency was observed in OVCAR3 cells
(P<0.05; Fig. 1C). ERBB3
interacted with miR-125a-5p, miR-125b-5p, miR-22-3p and miR-205-5p
in the oncomiRDB datebase (http://bioinfo.au.tsinghua.edu.cn/member/jgu/oncomirdb/index.php)
(Fig. 1D). However, the
association between miR-152 and ERBB3 remains unclear.
miR-152 downregulates ERBB3 expression in
human ovarian cancer
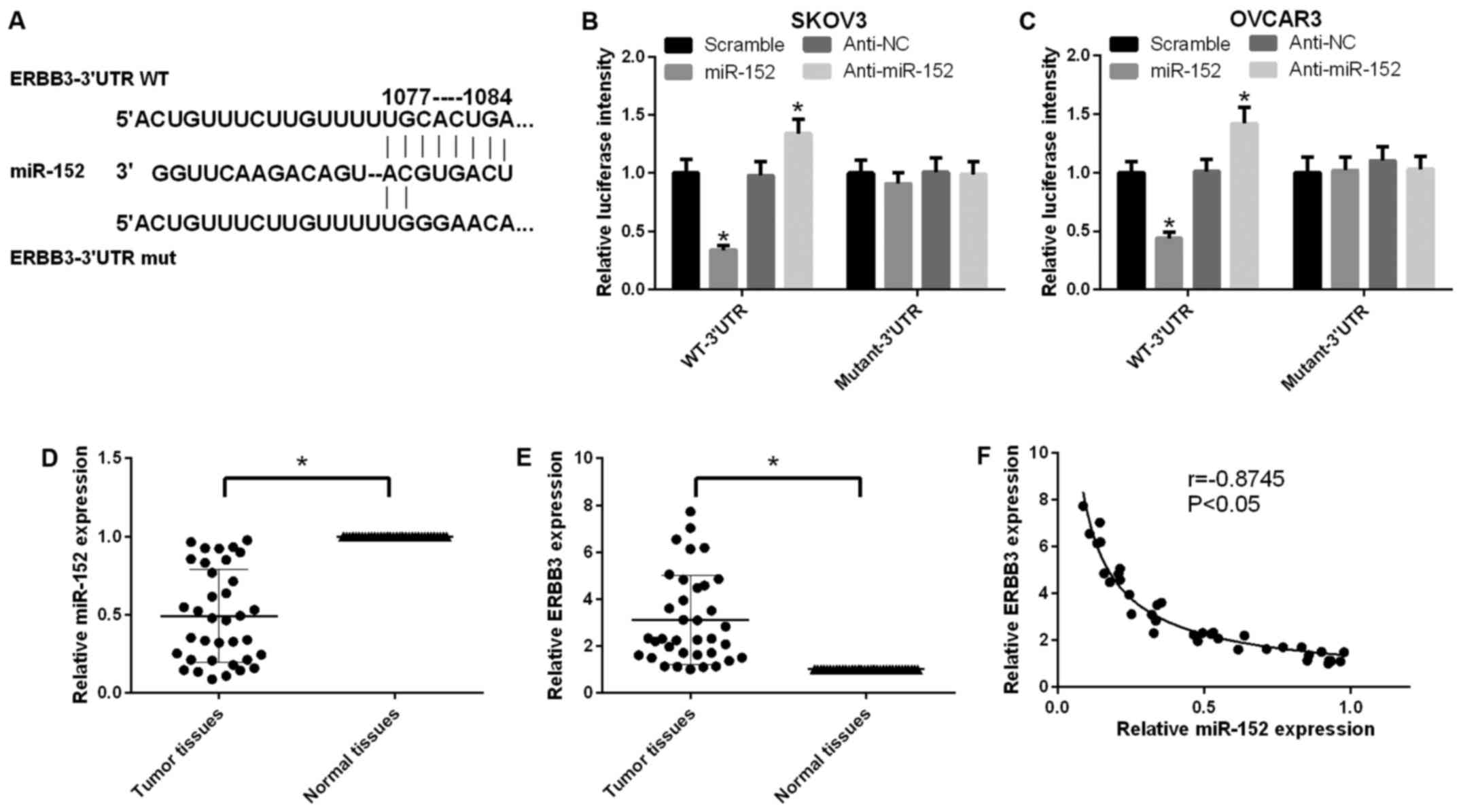
According to the TargetScan prediction, wild-type
and mutated ERBB3 vectors were structured and cotransfected with
miR-152 or anti-miR-152 in SKOV3 cells, respectively. The miR-152
target site in the sequence of ERBB3 and the mutational site of
ERBB3 are shown in Fig. 2A. It
was indicated that miR-152 negatively regulated ERBB3 using the
luciferase reporter gene assay in SKOV3 cells (P<0.05; Fig. 2B). A similar tendency was observed
in OVCAR3 cells (Fig. 2C). In
addition, the expression level of miR-152 in ovarian cancer tissues
was found to be lower compared with that in adjacent non-cancerous
tissues (P<0.05; Fig. 2D).
Furthermore, the expression level of ERBB3 in ovarian cancer
tissues was higher compared with that in adjacent non-cancerous
tissues (P<0.05; Fig. 2E).
Further analysis confirmed a negative correlation between miR-152
and ERBB3 in ovarian cancer tissues (P<0.05, r=0.8745; Fig. 2F).
miR-152 suppresses the ability of ovarian
cancer cell proliferation and promotes apoptosis through inhibiting
ERBB3
To further study the function of miR-152 in ovarian
cancer, functional assays were performed in ovarian cancer cell
lines. The mRNA expression level of ERBB3 was decreased in SKOV3
and OVCAR3 cells transfected with miR-152 compared with scramble,
and was significantly increased in SKOV3 and OVCAR3 cells
transfected with both miR-152 and ERBB3 relative to the control
group (P<0.05) (Fig. 3A).
Similarly, the protein level of ERBB3 was also reduced by ERBB3
vector in SKOV3 and OVCAR3 cells (Fig. 3B and C). Furthermore, the cell
proliferation ability was decreased in SKOV3 cells transfected with
miR-152 compared with scramble, and this ability was significantly
increased in SKOV3 cells transfected with both miR-152 and ERBB3
relative to both miR-152 and vector (Fig. 3D). A similar result was observed
in OVCAR3 cells (Fig. 3E).
Similarly, the CFU assay results indicated that miR-152 inhibited
the proliferation ability of SKOV3 and OVCAR3 cells through ERBB3
(Fig. 3F). Apoptosis assay proved
that miR-152 accelerated the apoptosis of SKOV3 and OVCAR3 cells,
and that ERBB3 can reverse miR-152-mediated promotion of apoptosis
in vitro (Fig. 3G).
Furthermore, to detect the effect of miR-152 on tumorigenesis in
vivo, SKOV3 cells transfected with scramble, miR-152, miR-152
and vector, or miR-152 and ERBB3 were implanted subcutaneously into
nude mice. The results indicated that miR-152 markedly inhibited
tumor growth, while ERBB3 was able to reverse this phenotype
(Fig. 3H).
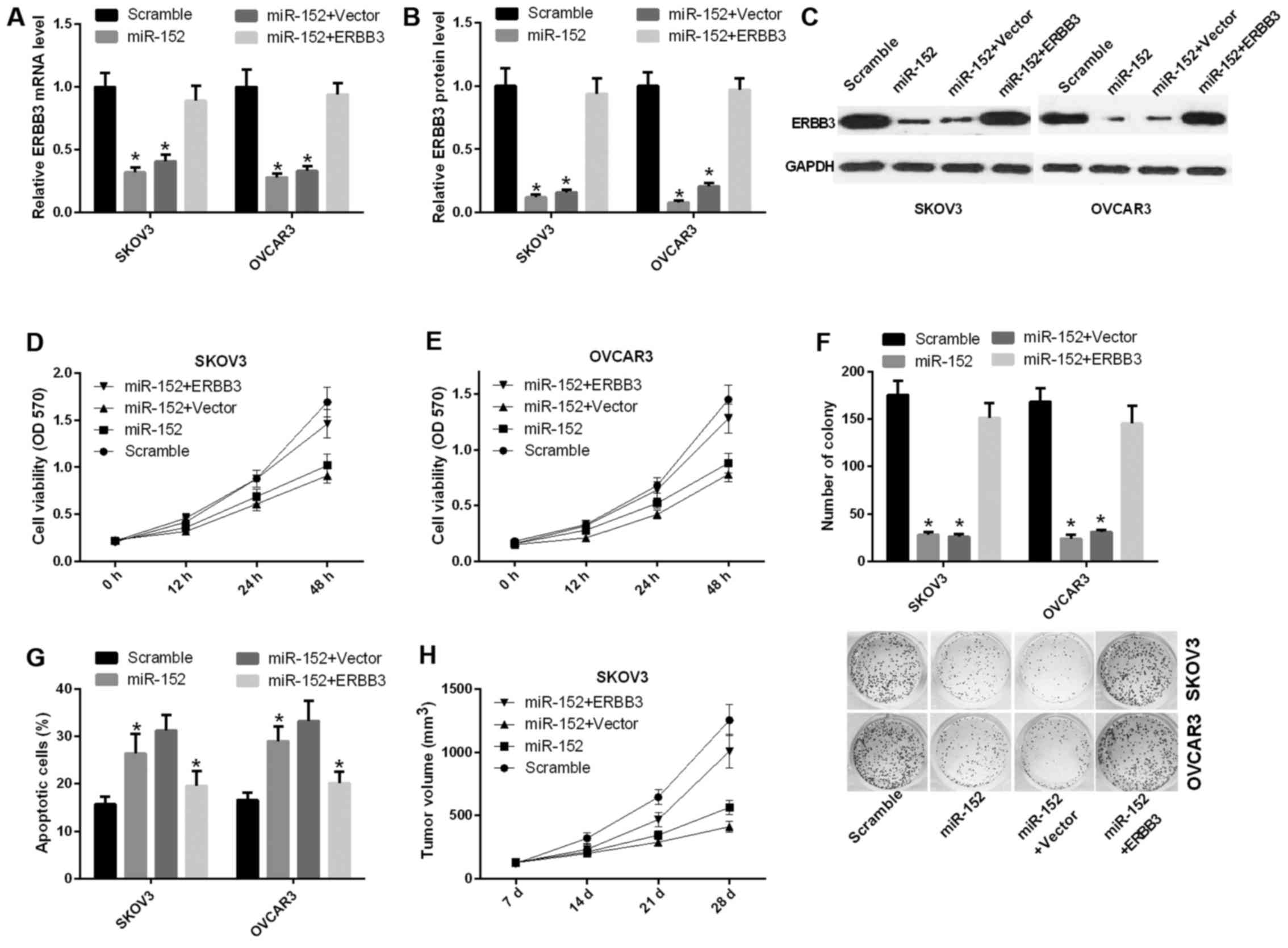 | Figure 3miR-152 suppresses the ability of
ovarian cancer proliferation and promotes apoptosis through
inhibiting ERBB3. (A) Reverse transcription-quantitative polymerase
chain reaction was used to determine the mRNA expression level of
ERBB3 in SKOV3 and OVCAR3 cells transfected with scramble, miR-152,
miR-152 and vector, or miR-152 and ERBB3 (P<0.05). (B and C)
Analysis of the ERBB3 protein expression level in SKOV3 and OVCAR3
cells (P<0.05). (D and E) The MTT assay was performed to
determine the proliferation capacity of SKOV3 and OVCAR3 cells
transfected with scramble, miR-152, miR-152 and vector, or miR-152
and ERBB3. (F) The capacity of cell proliferation was analyzed by
colony formation evaluation in SKOV3 and OVCAR3 cells (P<0.05).
(G) The number of apoptotic cells was counted in SKOV3 and OVCAR3
cells transfected with scramble, miR-152, miR-152 and vector, or
miR-152 and ERBB3 (P<0.05). (H) Athymic mice were treated with
SKOV3 cells transfected with scramble, miR-152, miR-152 and vector,
or miR-152 and ERBB3 for 7, 14, 21 and 28 days. The tumor volume
was measured and calculated at the specified timepoints. OD,
optical density. |
miR-152 inhibits the ability of migration
and invasion in ovarian cancer cells through ERBB3
The effect of miR-152 on cell migration and invasion
ability of ovarian cancer cells was further investigated. First,
SKOV3 and OVCAR3 cells were transfected with scramble, miR-152,
miR-152 and vector, or miR-152 and ERBB3. The results of the
invasion assay demonstrated that the invasion ability was
significantly decreased in SKOV3 and OVCAR3 cells transfected with
miR-152 compared with scramble, and significantly increased in
SKOV3 cells transfected with both miR-152 and ERBB3 relative to
both miR-152 and vector (P<0.05; Fig. 4A and B). The results of the
migration assay also demonstrated that the migration ability was
significantly decreased in SKOV3 and OVCAR3 cells transfected with
miR-152 compared with scramble, and significantly increased in
SKOV3 cells transfected with both miR-152 and ERBB3 relative to
both miR-152 and vector (P<0.05; Fig. 4C and D). In addition, the
migration ability of SKOV3 and OVCAR3 cells transfected with
scramble, miR-152, miR-152 and vector, or miR-152 and ERBB3, were
evaluated by the wound healing assay at 1 and 3 days. The results
indicated that miR-152 inhibited the migration ability of SKOV3 and
OVCAR3 cells through ERBB3 (Fig. 4E
and F).
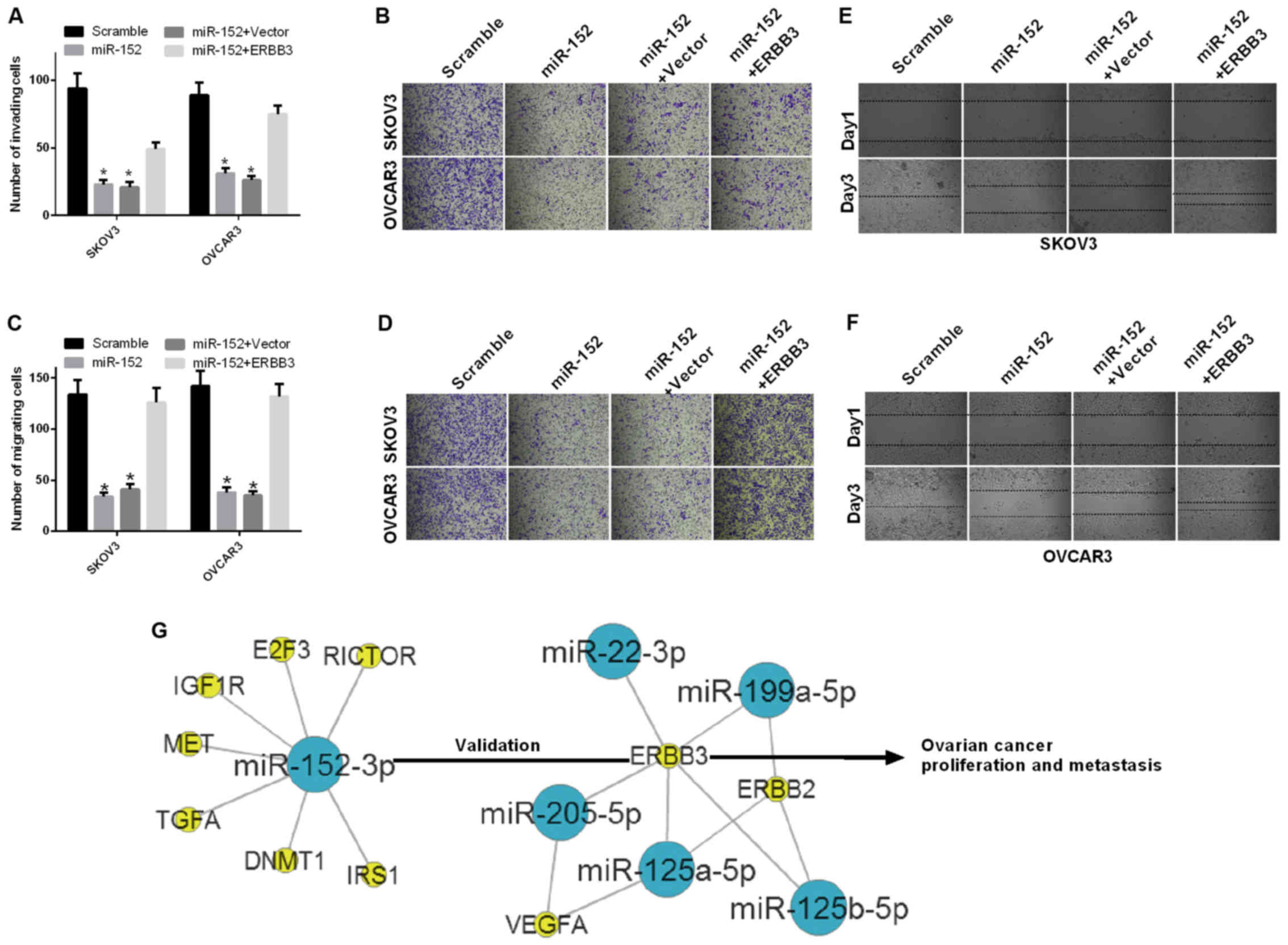 | Figure 4miR-152 inhibits the ability of
migration and invasion of ovarian cancer cells through ERBB3. (A
and B) The number of invading cells was counted among SKOV3 and
OVCAR3 cells transfected with scramble, miR-152, miR-152 and
vector, or miR-152 and ERBB3 (P<0.05). (C and D) The migration
assay was performed in SKOV3 and OVCAR3 cells transfected with
scramble, miR-152, miR-152 and vector, or miR-152 and ERBB3.
Magnification, ×200 (P<0.05). (E and F) The migration ability of
SKOV3 and OVCAR3 cells that were transfected with scramble,
miR-152, miR-152 and vector, or miR-152 and ERBB3 was measured by
the wound healing assay at 1 and 3 days. (G) The gene network
between miR-152 and ERBB3 was established in ovarian cancer. Blue
nodes, miRs; yellow nodes, target genes. |
Regulatory mechanism underlying the
interaction between miR-152 and ERBB3 in ovarian cancer
The gene network between miR-152 and ERBB3 was
established in ovarian cancer as shown in Fig. 4G. miR-152 downregulated the
expression level of ERBB3 by targeting the ERBB3 promoter-binding
site and inhibited ovarian cancer cell proliferation, migration and
invasion, while promoting apoptosis in vitro.
Discussion
Accumulating evidence reported that miRNAs play key
roles in human cancer, including cancer cell proliferation,
metastasis, inflammation and angiogenesis (13,34). Recent studies proved that miR-152
served as a tumor inhibitor, and could be silenced by DNA
hypermethylation in endometrial cancer (27). The combined effect of miR-152 and
miR-185 played an important role in the treatment of ovarian cancer
by targeting DNMT1, independent of decitabine (35). miR-152 affected the proliferation
ability of ovarian cancer cells (30), whereas miR-152 reduced the
invasion and angiogenesis capacity of glioma cells (36). Downregulated miR-152 affected
aberrant DNA methylation in hepatitis B virus-related
hepatocellular carcinoma (28).
miR-152 inhibited migration and invasion through TGF-α in prostate
cancer cells (29). However,
their mechanism of action is not entirely clear in ovarian cancer.
In the present study, miR-152 expression was found to be lower in
ovarian cancer tissues compared with that in adjacent non-cancerous
tissues. miR-152 suppressed the ability of ovarian cancer
proliferation, migration and invasion, and promoted apoptosis in
vitro.
ERBB3 is one of the four members of ERBB family,
which may transduce extracellular signals into the cell, resulting
in several changes in the regulatory processes of various cancer
cells, including proliferation, survival, apoptosis, migration and
invasion (37–39). It has been demonstrated that ERBB3
significantly downregulates kinase activity (40). ERBB3 overexpression has been
proven to take part in the regulation of various physiological and
pathological pathways in several types of cancer, such as melanoma,
breast, prostate, lung and pancreatic cancer (41–44). It has also been reported that
ERBB3 may activate downstream phosphatidylinositol-3-kinase, which
is associated with tumorigenesis and resistance to therapy
(45). Therefore, an increasing
number of studies indicate that ERBB3 is a potential target of
cancer therapy. In the present study, ERBB3 expression was found to
be higher in ovarian cancer tissues compared with that in adjacent
non-cancerous tissues. There was a negative correlation between
miR-152 and ERBB3 expression in ovarian cancer tissues. ERBB3 was
downregulated by miR-152 in ovarian cancer cells. ERBB3 may reverse
miR-152 mediated-inhibition of ovarian cancer proliferation,
migration and invasion, and miR-152-mediated promotion of apoptosis
in vitro. ERBB3 may also reverse miR-152-mediated inhibition
of tumor growth in vivo.
In conclusion, the findings of the present study
indicated that miR-152 is involved in the regulation of the
proliferation and metastasis of ovarian cancer cells through
repression of ERBB3 expression. First, it was demonstrated that the
mRNA expression levels of miR-125a-5p, miR-125b-5p, miR-22-3p,
miR-205-5p and miR-152 were significantly downregulated in SKOV3
cells. Second, a negative correlation between miR-152 and ERBB3 was
identified in ovarian cancer tissues, and miR-152 downregulated
ERBB3 expression in ovarian cancer cells. Furthermore, our results
demonstrated that miR-152 suppressed the ability of ovarian cancer
cells to proliferate, migrate and invade, and promoted their
apoptosis through inhibiting ERBB3 in vitro. Therefore, the
present study demonstrated that miR-152 may be a potential
therapeutic target for the treatment of ovarian cancer.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81472028), the
Scientific Research Project of Hei Long-jiang Provincial Health
Bureau of China [AN1] (no. 2016-105) and the Administration of
Traditional Chinese Medicine of Heilongjiang Province, China (no.
ZHY16-110).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim K, Zang R, Choi SC, Ryu SY and Kim JW:
Current status of gynecological cancer in China. J Gynecol Oncol.
20:72–76. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Legge F, Ferrandina G, Salutari V and
Scambia G: Biological characterization of ovarian cancer:
prognostic and therapeutic implications. Ann Oncol. 16(Suppl 4):
iv95–101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: The impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Seidman JD, Soslow RA, Vang R, Berman JJ,
Stoler MH, Sherman ME, Oliva E, Kajdacsy-Balla A, Berman DM and
Copeland LJ: Borderline ovarian tumors: Diverse contemporary
viewpoints on terminology and diagnostic criteria with illustrative
images. Hum Pathol. 35:918–933. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iorio MV, Visone R, Di Leva G, Donati V,
Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, et
al: MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zaman MS, Maher DM, Khan S, Jaggi M and
Chauhan SC: Current status and implications of microRNAs in ovarian
cancer diagnosis and therapy. J Ovarian Res. 5:442012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heintz AP, Odicino F, Maisonneuve P, Quinn
MA, Benedet JL, Creasman WT, Ngan HY, Pecorelli S and Beller U:
Carcinoma of the ovary. Int J Gynaecol Obstet. 95(Suppl 1):
S161–S192. 2006. View Article : Google Scholar
|
|
9
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. Adv Exp Med Biol. 774:1–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Djuranovic S, Nahvi A and Green R: A
parsimonious model for gene regulation by miRNAs. Science.
331:550–553. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kasinski AL and Slack FJ: Epigenetics and
genetics MicroRNAs en route to the clinic: Progress in validating
and targeting microRNAs for cancer therapy. Nat Rev Cancer.
11:849–864. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheng AM, Byrom MW, Shelton J and Ford LP:
Antisense inhibition of human miRNAs and indications for an
involvement of miRNA in cell growth and apoptosis. Nucleic Acids
Res. 33:1290–1297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Di Leva G and Croce CM: The role of
microRNAs in the tumori-genesis of ovarian cancer. Front Oncol.
3:1532013. View Article : Google Scholar
|
|
18
|
Dahiya N and Morin PJ: MicroRNAs in
ovarian carcinomas. Endocr Relat Cancer. 17:F77–F89. 2010.
View Article : Google Scholar :
|
|
19
|
Mateescu B, Batista L, Cardon M, Gruosso
T, de Feraudy Y, Mariani O, Nicolas A, Meyniel JP, Cottu P,
Sastre-Garau X, et al: miR-141 and miR-200a act on ovarian
tumorigenesis by controlling oxidative stress response. Nat Med.
17:1627–1635. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dahiya N, Sherman-Baust CA, Wang TL,
Davidson B, Shih IeM, Zhang Y, Wood W III, Becker KG and Morin PJ:
MicroRNA expression and identification of putative miRNA targets in
ovarian cancer. PLoS One. 3:e24362008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu X, Macdonald DM, Huettner PC, Feng Z,
El Naqa IM, Schwarz JK, Mutch DG, Grigsby PW, Powell SN and Wang X:
A miR-200 microRNA cluster as prognostic marker in advanced ovarian
cancer. Gynecol Oncol. 114:457–464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheng W, Liu T, Wan X, Gao Y and Wang H:
MicroRNA-199a targets CD44 to suppress the tumorigenicity and
multidrug resistance of ovarian cancer-initiating cells. FEBS J.
279:2047–2059. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Laios A, O'Toole S, Flavin R, Martin C,
Kelly L, Ring M, Finn SP, Barrett C, Loda M, Gleeson N, et al:
Potential role of miR-9 and miR-223 in recurrent ovarian cancer.
Mol Cancer. 7:352008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nagaraja AK, Creighton CJ, Yu Z, Zhu H,
Gunaratne PH, Reid JG, Olokpa E, Itamochi H, Ueno NT, Hawkins SM,
et al: A link between mir-100 and FRAP1/mTOR in clear cell ovarian
cancer. Mol Endocrinol. 24:447–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu X, Tian J, Zhang L, Chen Y and Hao Q:
Involvement of microRNA-93, a new regulator of PTEN/Akt signaling
pathway, in regulation of chemotherapeutic drug cisplatin
chemosensitivity in ovarian cancer cells. FEBS Lett. 586:1279–1286.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen Y, Song Y, Wang Z, Yue Z, Xu H, Xing
C and Liu Z: Altered expression of MiR-148a and MiR-152 in
gastrointestinal cancers and its clinical significance. J
Gastrointest Surg. 14:1170–1179. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsuruta T, Kozaki K, Uesugi A, Furuta M,
Hirasawa A, Imoto I, Susumu N, Aoki D and Inazawa J: miR-152 is a
tumor suppressor microRNA that is silenced by DNA hypermethylation
in endometrial cancer. Cancer Res. 71:6450–6462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang J, Wang Y, Guo Y and Sun S:
Down-regulated microRNA-152 induces aberrant DNA methylation in
hepatitis B virus-related hepatocellular carcinoma by targeting DNA
methyltransferase 1. Hepatology. 52:60–70. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu C, Li J, Ding Q, Cheng G, Zhou H, Tao
L, Cai H, Li P, Cao Q, Ju X, et al: miR-152 controls migration and
invasive potential by targeting TGFα in prostate cancer cell lines.
Prostate. 73:1082–1089. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou X, Zhao F, Wang Z-N, Song YX, Chang
H, Chiang Y and Xu HM: Altered expression of miR-152 and miR-148a
in ovarian cancer is related to cell proliferation. Oncol Rep.
27:447–454. 2012.
|
|
31
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang L, Lai YK, Zhang J, Wang H, Lin MC,
He ML and Kung HF: Targeting S100P inhibits colon cancer growth and
metastasis by Lentivirus-mediated RNA interference and proteomic
analysis. Mol Med. 17:709–716. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Madhyastha HK, Radha KS, Nakajima Y, Omura
S and Maruyama M: uPA dependent and independent mechanisms of wound
healing by C-phycocyanin. J Cell Mol Med. 12:2691–2703. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Paris PL, Chen J, Ngo V, Yao H,
Frazier ML, Killary AM, Liu CG, Liang H, Mathy C, et al: Next
generation sequencing of pancreatic cyst fluid microRNAs from low
grade-benign and high grade-invasive lesions. Cancer Lett.
356:404–409. 2015. View Article : Google Scholar
|
|
35
|
Xiang Y, Ma N, Wang D, Zhang Y, Zhou J, Wu
G, Zhao R, Huang H, Wang X, Qiao Y, et al: MiR-152 and miR-185
co-contribute to ovarian cancer cells cisplatin sensitivity by
targeting DNMT1 directly: A novel epigenetic therapy independent of
decitabine. Oncogene. 33:378–386. 2014. View Article : Google Scholar
|
|
36
|
Zheng X, Chopp M, Lu Y, Buller B and Jiang
F: MiR-15b and miR-152 reduce glioma cell invasion and angiogenesis
via NRP-2 and MMP-3. Cancer Lett. 329:146–154. 2013. View Article : Google Scholar :
|
|
37
|
Sharma SV and Settleman J: ErbBs in lung
cancer. Exp Cell Res. 315:557–571. 2009. View Article : Google Scholar
|
|
38
|
Hynes NE and Lane HA: ERBB receptors and
cancer: The complexity of targeted inhibitors. Nat Rev Cancer.
5:341–354. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hynes NE and MacDonald G: ErbB receptors
and signaling pathways in cancer. Curr Opin Cell Biol. 21:177–184.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shi F, Telesco SE, Liu Y, Radhakrishnan R
and Lemmon MA: ErbB3/HER3 intracellular domain is competent to bind
ATP and catalyze autophosphorylation. Proc Natl Acad Sci USA.
107:7692–7697. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ocana A, Vera-Badillo F, Seruga B,
Templeton A, Pandiella A and Amir E: HER3 overexpression and
survival in solid tumors: A meta-analysis. J Natl Cancer Inst.
105:266–273. 2013. View Article : Google Scholar
|
|
42
|
Cook RS, Garrett JT, Sánchez V, Stanford
JC, Young C, Chakrabarty A, Rinehart C, Zhang Y, Wu Y, Greenberger
L, et al: ErbB3 ablation impairs PI3K/Akt-dependent mammary
tumorigenesis. Cancer Res. 71:3941–3951. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wimmer E, Kraehn-Senftleben G and Issing
WJ: HER3 expression in cutaneous tumors. Anticancer Res.
28:973–979. 2008.PubMed/NCBI
|
|
44
|
Reschke M, Mihic-Probst D, van der Horst
EH, Knyazev P, Wild PJ, Hutterer M, Meyer S, Dummer R, Moch H and
Ullrich A: HER3 is a determinant for poor prognosis in melanoma.
Clin Cancer Res. 14:5188–5197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Prigent SA and Gullick WJ: Identification
of c-erbB-3 binding sites for phosphatidylinositol 3′-kinase and
SHC using an EGF receptor/c-erbB-3 chimera. EMBO J. 13:2831–2841.
1994.PubMed/NCBI
|