Introduction
Hepatic fibrosis, a spontaneous wound healing
response, occurs when the liver has an acute or chronic injury
caused by viral infection, drug damage or immune attack (1,2).
Fibrosis impairs tissue and organ function and is caused by
excessive deposition of the extracellular matrix (ECM) that occurs
as a result of inflammation (3,4).
Liver fibrosis markedly contributes to morbidity in patients with
liver disease (5). Hepatic
stellate cells (HSCs) are pivotal in the development of hepatic
fibrosis (6) and have the ability
to proliferate and differentiate into myofibroblast-like cells
(7). Platelet-derived growth
factor (PDGF) signaling is one of the best characterized pathways
of HSC activation (8) and induces
apoptosis, senescence or quiescence in HSCs (9). Thus, inhibiting the activation of
HSCs may be a feasible therapeutic strategy for hepatic
fibrosis.
Various agents, including malotilate, genistein,
curcumin and silymarin, have been reported to be effective against
fibrosis in vitro and in vivo (10,11); however, none of these are ideal
candidates for inhibiting the activation of HSCs. Currently, an
increasing number of natural products derived from herbal extracts
are used in the treatment of diseases such as liver fibrosis;
however, the different mechanisms through which they work are not
known (12). Baicalin (Fig. 1), is the major bioactive flavonoid
extracted from dried roots of the herb Scutellaria
baicalensis Georgi (13).
Baicalin exhibits a range of pharmacological effects that include
antioxidative, anti-viral, and anti-inflammatory properties, as
well as liver protection (14).
Previous reports have indicated that baicalin has significant
anti-fibrogenic effects that act by suppressing the signaling of
canonical Wnts (15,16). Baicalin also inhibits hepatic
fibrosis by reducing the level of pro-fibrotic cytokines, including
transforming growth factor (TGF)-β1, tumor necrosis factor-α,
interleukin (IL)-6 and IL-22. The reduction of these cytokines
significantly enhances HSC senescence or apoptosis and reduce the
activation of HSCs (17,18). To expand the application of
baicalin, it is necessary to further elucidate the mechanisms of
the anti-fibrotic effect of baicalin.
MicroRNA (miRNA) are short RNA molecules (~22
nucleotides) (19) that typically
bind to the 3′ untranslated region (3′UTR) of specific mRNA to
regulate gene expression, either by repressing mRNA translation or
promoting repression. They are involved in diverse physiological
and developmental processes (20,21) and regulate all biological
processes in all cell types, such as cells in the liver. Previous
research suggests that miRNA are involved in the regulation of HSC
activation (7). Thus, modulating
miRNA represents a novel therapeutic approach for hepatologists in
the treatment of liver disease.
Long-chain-fatty-acid-CoA ligase 4 (ACSL4) is a
member of the long-chain acyl-CoA synthetase (ACSL4) family that
preferentially uses arachidonate and eicosapentaenoate as
substrates, and activates long-chain fatty acids for the synthesis
of cellular lipids (22,23). ACSL4 is associated with various
liver diseases (24) and is
aberrantly regulated in liver cancer (25).
The present study demonstrated the anti-fibrogenic
property of baicalin in PDGF-BB-induced HSCs. miRNA profiles of
PDGF-BB-induced activated HSC-T6 cells treated with and without
baicalin were compared. Results indicated that miR-3595 was
upregulated in baicalin-treated cells compared with the levels in
the untreated cells. It was also revealed that ACSL4 is a potential
functional target of miR-3595. These findings indicate the central
role of baicalin in inhibiting the activation of HSCs and suggest
that baicalin has the potential to be an effective natural medicine
for the treatment of hepatic fibrosis. The present results support
ongoing research to identify potential therapeutic targets for
liver fibrosis.
Materials and methods
Cell culture and treatment
HSC-T6 cells were purchased from American Type
Culture Collection (Manassas, VA, USA) and were cultured in
Dulbecco's modified Eagle medium (DMEM; cat. no. 11965-092;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; cat. no. 10099-141;
Invitrogen; Thermo Fisher Scientific, Inc.). The cells were grown
in a humidified atmosphere maintained at 5% CO2 at 37°C.
The cells were serum-starved in an FBS-free medium for 24 h and
then induced with PDGF-BB (10 ng/ml; cat. no. P3201; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany), following the protocol in a
previous study (26). The cells
were treated with baicalin at different concentrations (50, 100 and
150 µM; cat. no. 572667; Sigma-Aldrich; Merck KGaA) for the
indicated time according to different experiments at 37°C. The same
volume of diluted dimethyl sulfoxide (DMSO) was used as a negative
control. While the baicalin stock solution was in DMSO, it was
ensured that the concentration of DMSO did not influence cell
survival. Plasmids [ACSL4 wild-type (WT) and mutant (MUT);
constructed by Shanghai GenePharma Co., Ltd., Shanghai, China] and
miR-3595 inhibitors (5′-AUCGUAGAGGAAAAUCCAC-3′) or negative control
(NC; 5′-CAGUACUUUUGUGUAGUACAA-3′) (Shanghai GenePharma Co., Ltd.)
were transfected into cells using Lipofectamine® 2000
(cat. no. 11668027; Invitrogen; Thermo Fisher Scientific, Inc.) at
a concentration of 50 nM, following the manufacturer's
instructions. After 48 h of transfection, cells were treated with
different reagents for different experiments.
Cell proliferation assay
The induced HSC-T6 cells (1×104
cells/well) were seeded into 96-well plates and treated with
baicalin for 72 h at 37°C. Cell proliferation was assessed using a
Cell Counting Kit-8 (CCK-8) assay (cat. no. 96992; Sigma-Aldrich;
Merck KGaA), according to the manufacturer's protocol. The optical
density was measured at 450 nm.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from HSC-T6 cells was extracted using
TRIzol reagent (cat. no. 15596026; Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. Total
RNA was reverse-transcribed to cDNA by using a Bestar qPCR RT kit
(cat. no. 210212; Qiagen GmbH, Hilden, Germany) following the
manufacturer's protocol. Following this, cDNA was quantified by
qPCR on a Mx3000p real-time PCR system with DBI Bestar®
SYBRGreen qPCR master mix (cat. no. DBI-2073; DBI Bioscience,
Ludwigshafen, Germany) in a 20-µl PCR reaction. qPCR was
performed using the following settings: Pre-denaturation at 95°C
for 10 sec, followed by 40 cycles of denaturation at 95°C for 10
sec and elongation at 60°C for 30 sec. Results were normalized to
β-actin expression. The 2−ΔΔCq method was used for
calculating the relative expression levels (27). The PCR primer sequences are listed
in Table I.
 | Table IPrimer sequences used for microRNA
and mRNA expression analysis. |
Table I
Primer sequences used for microRNA
and mRNA expression analysis.
| Primer | Sequence
(5′-3′) | Length, bp |
|---|
| β-actin F |
GGAGATTACTGCCCTGGCTCCTA | 150 |
| β-actin R |
GACTCATCGTACTCCTGCTTGCTG | |
| miR-3595 reverse
transcription |
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGATCGTAGA | |
| miR-3595 F |
ACACTCCAGCTGGGGTGGATTTTCCTCTA | |
| All R |
CTCAACTGGTGTCGTGGA | |
| ACSL4 F |
AAGGAGAAGGGCAAAGAGA | 211 |
| ACSL4 R |
GGTGGTTGTAGGAGGCTGA | |
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay
buffer (cat. no. R0278; Sigma-Aldrich; Merck KGaA) supplemented
with protease inhibitor cocktail (cat. no. ab201111; Abcam,
Cambridge, UK) for 30 min on ice. The protein concentration was
determined using a Bio-Rad protein assay kit (cat no. 5000002;
Bio-Rad Laboratories, Inc., Hercules, CA, USA). A total of 20
µg/lane protein was separated on 10% SDS-PAGE and
transferred to a polyvinylidene difluoride membrane. The membranes
were blocked with 5% skim milk for 1 h at room temperature. The
membranes were incubated overnight with the following specific
primary antibodies: α-smooth muscle actin (SMA; 1:1,000; cat. no.
A5228; Sigma-Aldrich; Merck KGaA), collagen (Col) I (1:1,000; cat.
no. NB600-408; Novus Biologicals, LLC, Littleton, CO, USA), Col III
(1:1,000; cat. no. ab6310; Abcam), TGF-β1 (1:1,000; cat. no.
ab92486; Abcam), desmin (1:1,000; cat. no. ab8592; Abcam), GAPDH
(1:4,000; cat. no. 14C10; Cell Signaling Technology, Inc., Danvers,
MA, USA) and ACSL4 (1:1,000; cat. no. sc-365230; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) at 4°C. Following this,
incubation was performed with horseradish peroxidase-conjugated
secondary immunoglobulin antibodies (1:10,000; cat. no. A4416 and
A6154; Sigma-Aldrich; Merck KGaA) for 1 h at room temperature.
Enhanced chemiluminescence chromogenic substrate (cat. no. 32109;
Invitrogen; Thermo Fisher Scientific, Inc.) was used and signals
were recorded on an X-ray film. β-actin was chosen as the loading
control.
Cell invasion and motility assay
A cell migration assay was performed using Transwell
chambers containing 8-µm pore size filters. For the invasion
assay, the inserts were coated with thin Matrigel. A total of
6×104 HSC-T6 cells in 200 µl FBS-free medium were
added to each upper chamber. A total of 500 µl DMEM
supplemented with 10% FBS was added to the lower chamber as a
chemoattractant. Following incubation for 24 and 36 h at 37°C, the
cells that migrated through the filter pores were fixed with 70%
ethanol at room temperature for 20 min and stained with crystal
violet for 10 min at room temperature. The results were calculated
from five random fields under a light microscope (magnification,
×400).
Cell cycle analysis
The cells were trypsinized, collected by
centrifugation at 300 × g for 5 min at room temperature and washed
with PBS. The harvested cells were subsequently fixed in pre-cooled
70% ethanol, and stored at 4°C overnight. The cells were then
washed with PBS and treated with DNase-free RNase (cat. no.
11119915001; Sigma-Aldrich; Merck KGaA) and stained with propidium
iodide in the dark for 15 min at room temperature. Cell samples
were analyzed by using a FACSCalibur (BD Biosciences, Franklin
Lakes, NJ, USA).
Analysis of apoptosis
The cells were trypsinized and collected by
centrifugation at 300 × g for 5 min at room temperature. The
harvested cells were washed with PBS and stained with annexin
V-fluorescein isothiocyanate and propidium iodide (PI; cat. no.
556547; BD Biosciences), according to the manufacturer's protocol.
Cell samples were analyzed using a flow cytometer within 1 h of
staining. The results were analyzed by FlowJo version 10 (BD
Biosciences).
miRNA microarray
The total RNA extracted using TRIzol was purified
and RNA quality was assessed using a NanoDrop ND-1000 (Thermo
Fisher Scientific, Inc., Wilmington, DE, USA). miRNA array
experiments were conducted, according to the manufacturer's
instructions. Total RNA was labeled using a miRCURY™ Array Power
Labeling kit (cat. no. 208500; Exiqon A/S, Vedbaek, Denmark). An
Axon GenePix 4000B microarray scanner (Molecular Devices, LLC,
Sunnyvale, CA, USA) was utilized to obtain the scanning profiles.
Following normalization, the statistical significance of
differentially expressed miRNA was analyzed by a Student's t-test
(GraphPad Prism 5; GraphPad Software, Inc., La Jolla, CA, USA).
Plasmid construction
To overexpress ACSL4, full-length Homo
sapiens ACSL4 sequences from the National Center for
Biotechnology Information database (Gene ID: 2182; ncbi.nlm.nih.gov/nuccore/NM_001318510.1)
were cloned into the pcDNA3.0 vector (cat. no. V790-20; Invitrogen;
Thermo Fisher Scientific, Inc.). To generate the luciferase
reporter plasmid, the 3′UTR of WT ACSL4 was cloned into the pLUC
vector (cat. no. 42094; Addgene, Inc., Cambridge, MA, USA).
Mutations in the 3′UTR of the WT ACSL4 were designed for four
nucleotides (AAAUCC to ACGUAA) of the 6-mer seed region of the
putative binding site of miR-3595 (pLUC-MUT-UTR). All DNA
constructs were sequenced.
Dual-luciferase reporter assay
The luciferase reporter plasmid pLUC was purchased
from Addgene, Inc., (cat. no. 42094). For the dual-luciferase
assay, HSC-T6 cells were transiently transfected with either the
pLUC-WT-UTR plasmid or the pLUC-MUT-UTR plasmid and miR-3595 mimics
or control NC mimics using Lipofectamine® 2000. After 48
h, cells were washed with PBS, lysed with lysis buffer and
luciferase activity was measured by using the Dual-Luciferase
Reporter Assay kit (cat. no. E1910; Promega Corporation, Madison,
WI, USA). Results were normalized to Renilla luciferase
activity.
miRNA targets mRNA prediction
Target mRNA of miRNA were identified using the
following two websites: microRNA.org
and targetscan.org.
Statistical analysis
Data were presented as the mean ± standard
deviation. Statistical analyses were performed using SPSS 17.0
(SPSS, Inc., Chicago, IL, USA) in all the pertinent experiments.
Differences between groups were analyzed using the Student's t-test
or one-way analysis of variance followed by a Tukey's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Baicalin inhibits the proliferation,
activation, apoptosis and cell cycle progression of activated
HSC-T6 cells induced by PDGF-BB
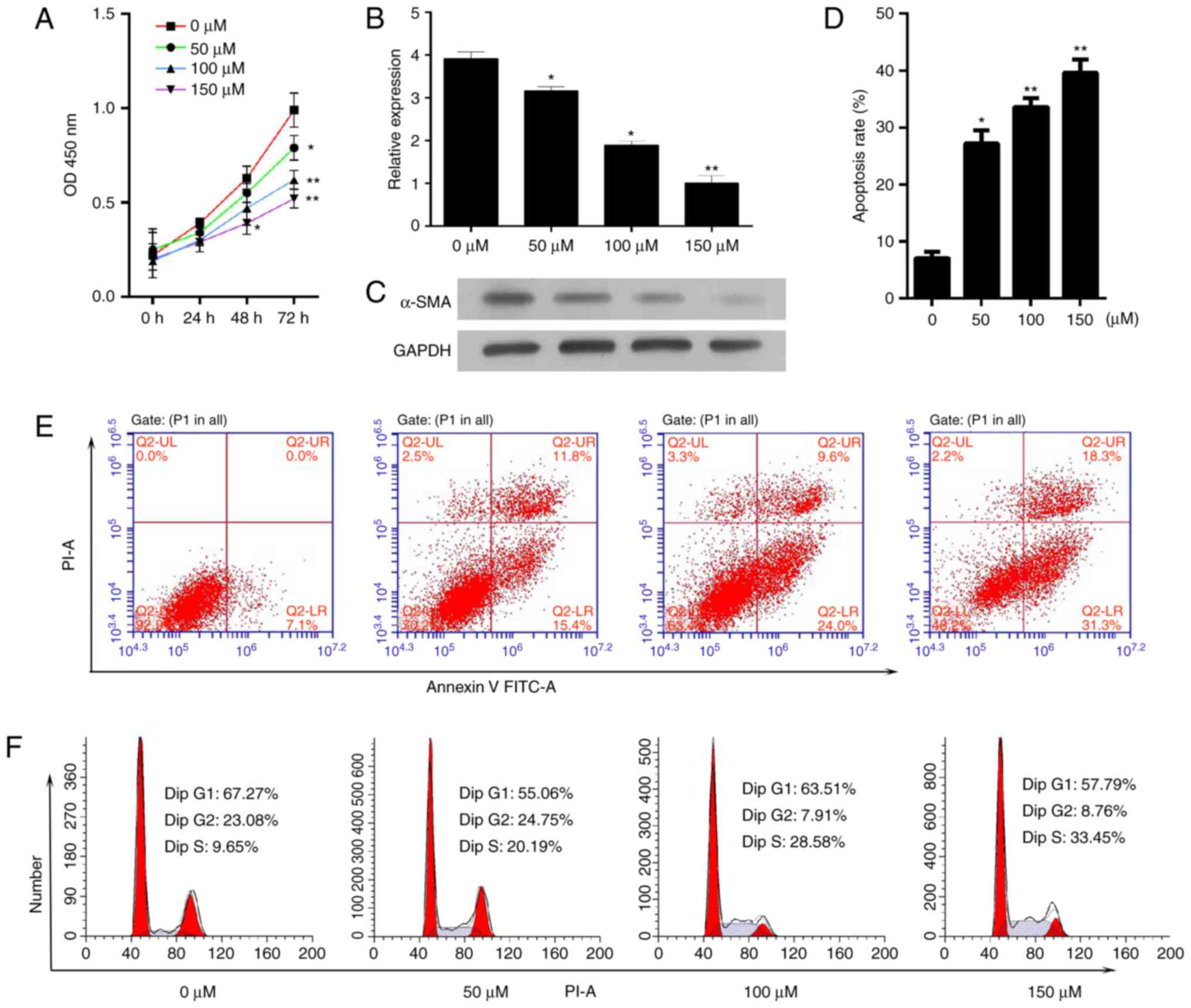
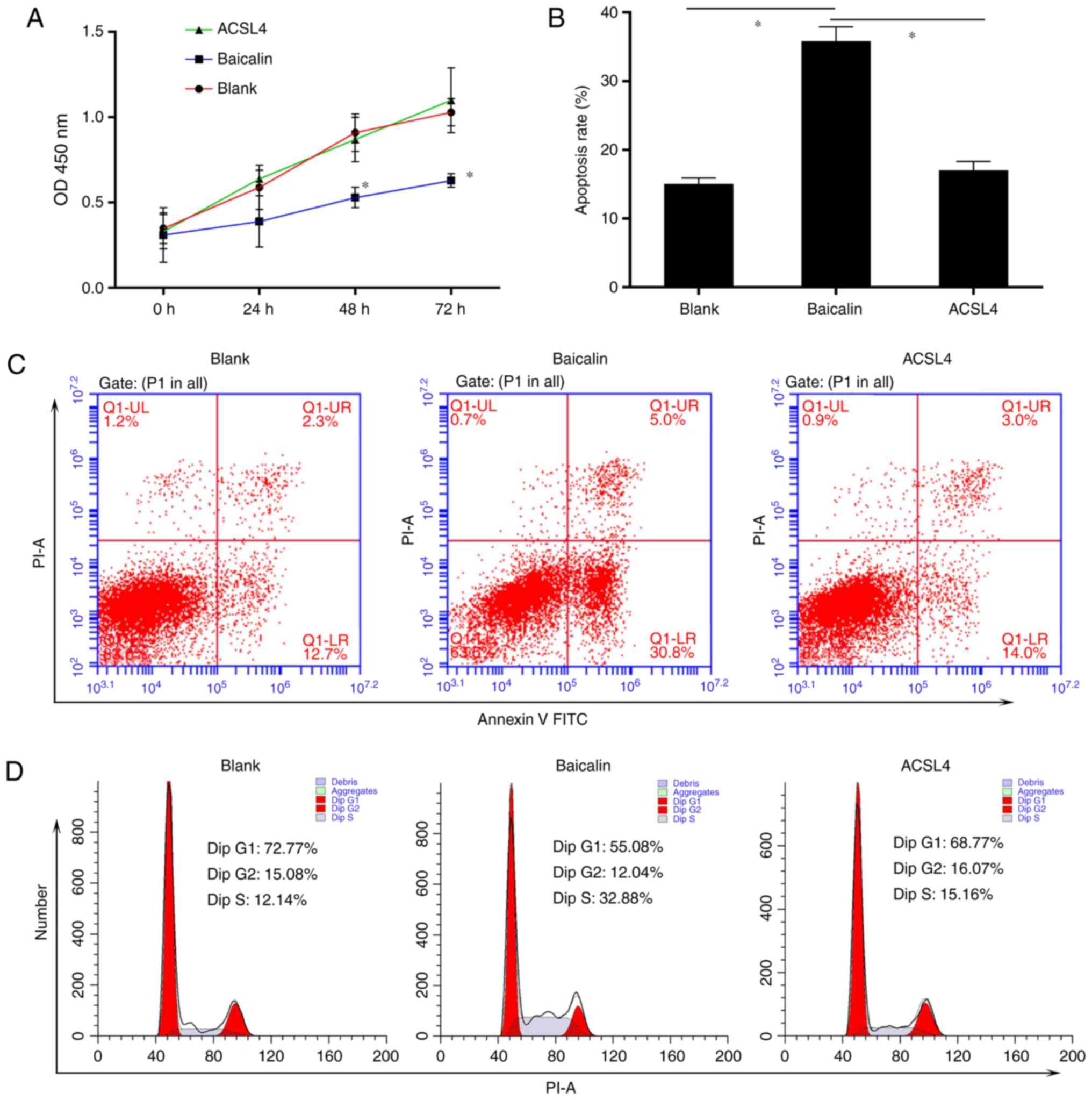
The present study aimed to determine the effect of
baicalin on cell proliferation and survival. Activated HSC-T6 cells
were treated with different concentrations of baicalin (50, 100 or
150 µM) for 72 h. A CCK-8 assay was utilized to measure cell
proliferation and it was demonstrated that baicalin decreased the
proliferative ability of the induced HSC-T6 cells (Fig. 2A). As a critical event in fibrosis
is the upregulation of proteins, such as α-SMA, the mRNA and
protein expression levels of α-SMA in activated HSC-T6 cells
following treatment with different concentrations of baicalin for
48 h were determined. The mRNA and protein expression levels of
α-SMA were significantly decreased compared with the levels in the
untreated group (Fig. 2B and C).
Subsequently, the effect of baicalin on apoptosis and cell cycle
progression was determined. It was demonstrated that baicalin
treatment significantly increased the number of apoptotic cells
(Fig. 2D and E). In addition,
baicalin treatment resulted in a notable increase in the number of
cells in S phase and a decrease in the number of cells in the G0/G1
phase compared with the numbers in the untreated group (Fig. 2F).
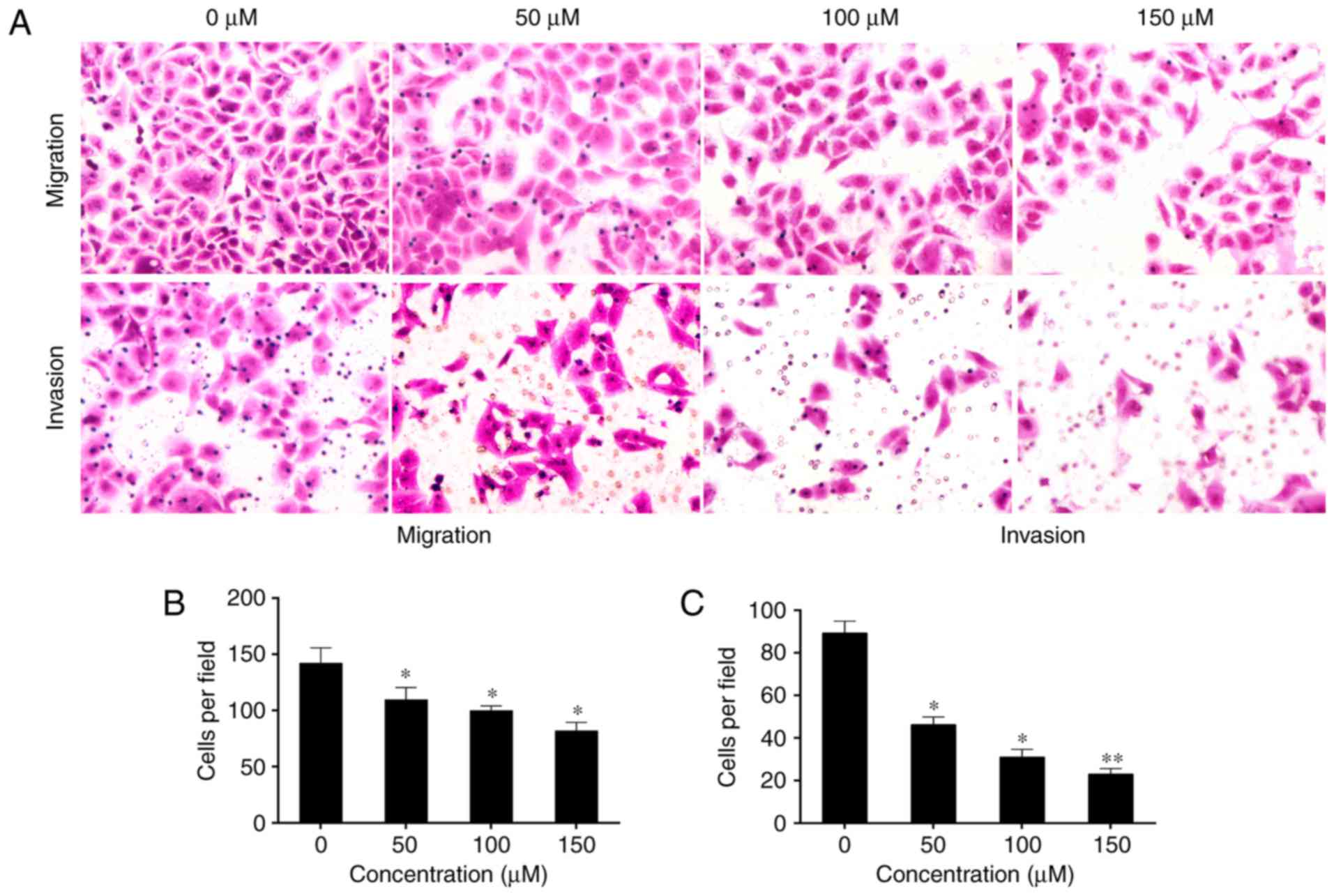
Baicalin decreases
epithelial-to-mesenchymal transition (EMT) of activated HSC-T6
cells induced by PDGF-BB
Epithelial cells lose their polarity as a result of
intercellular adhesion complexes changing their phenotypes. They
may then move through the ECM like mesenchymal cells, and this
process is termed EMT (28).
Activated HSCs exhibit a significant elevation in mesenchymal and
epithelial markers, which suggests that they undergo EMT (12). Therefore, the present study aimed
to determine the effect of baicalin on the invasion and migration
of induced HST-T6 cells. Treatment with baicalin significantly
inhibited the motility and invasive ability of the induced HST-T6
cells in a dose-dependent manner compared with that observed in
untreated cells (P<0.05; Fig.
3).
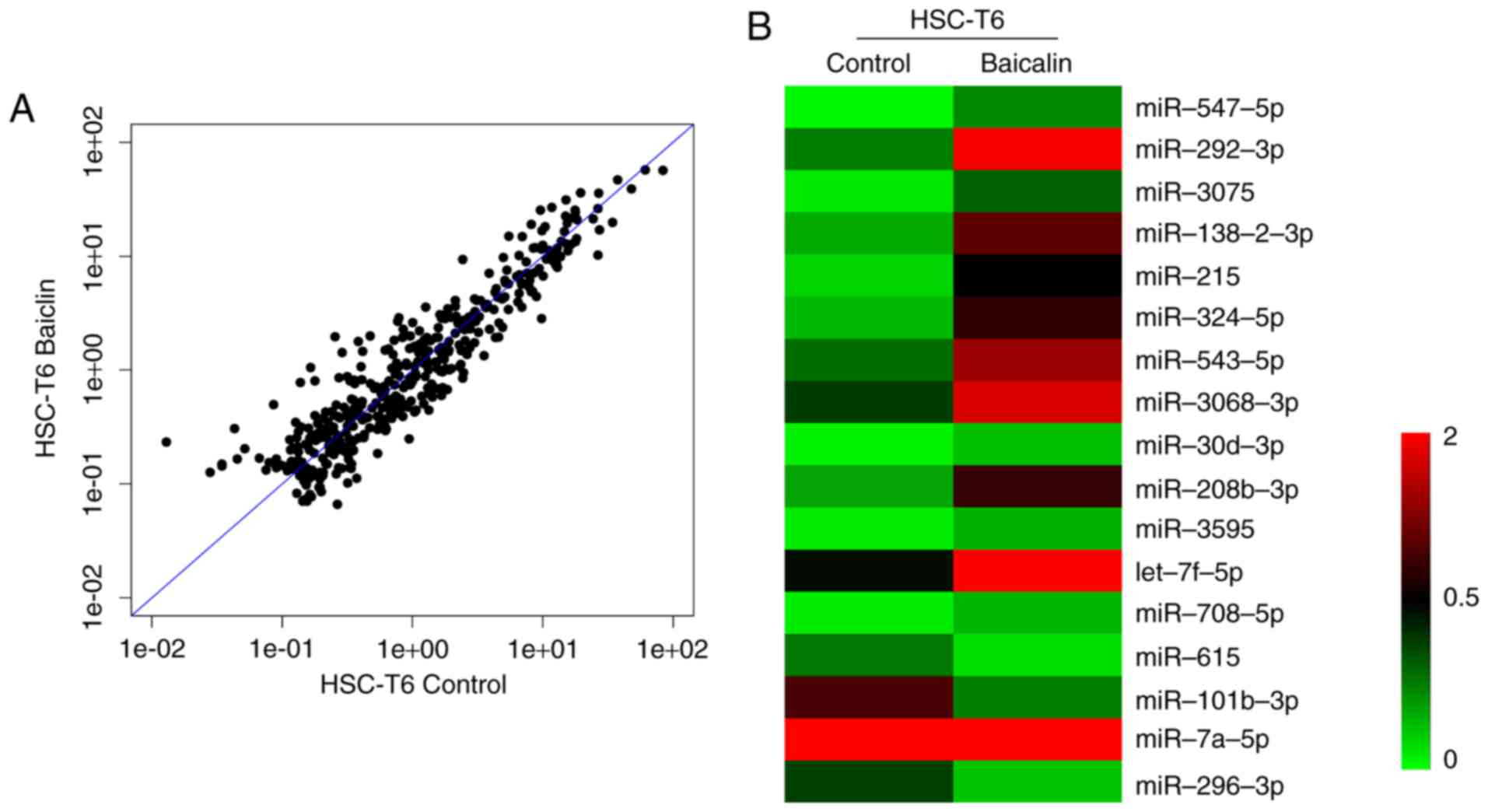
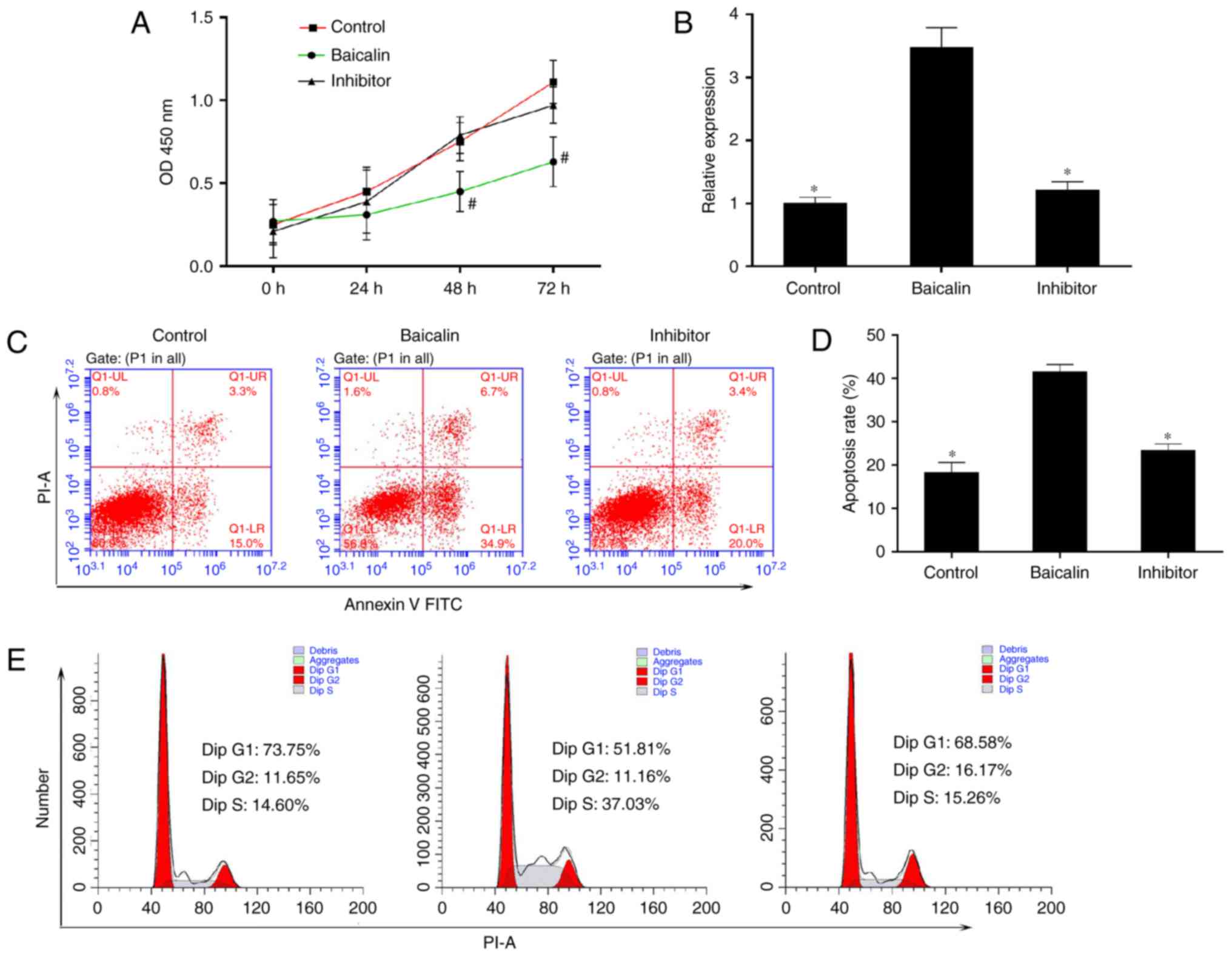
miR-3595 serves an important role in the
anti-fibrotic effect of baicalin
To identify the miRNA involved in the anti-fibrotic
effect of baicalin, miRNA expression profiles were compared between
induced HSC-T6 cells treated with 100 µM baicalin and
untreated cells, using a one-color miRNA array. miR-547-5p,
miR-296-3p and miR-3595 were dysregulated in the baicalin-treated
group when compared with the control group (Fig. 4). To further validate the
microarray data, the expression levels of miR-3595 were compared
between the two groups of cells using RT-qPCR. As demonstrated in
Fig. 5, miR-3595 was
significantly upregulated in baicalin-treated cells compared to the
level in the untreated cells (P<0.05; Fig. 5B). In addition, the cell
proliferation rate was rescued by miR-3595 inhibitor compared with
that in the baicalin-treated group, as measured by a CCK8 assay
(Fig. 5A). Cellular apoptosis and
cell cycle effects of the miR-3395 inhibitor were also detected. As
demonstrated in Fig. 5C and D,
baicalin significantly induced cell apoptosis compared with the
level in the control group, while the miR-3595 inhibitor reversed
baicalin-induced cell apoptosis. Baicalin treatment induced an
increase in the number of cells in S phase and decreased the number
of cells in the G0/G1 phase compared with the control group;
however, miR-3595 inhibitor reversed these baicalin-induced effects
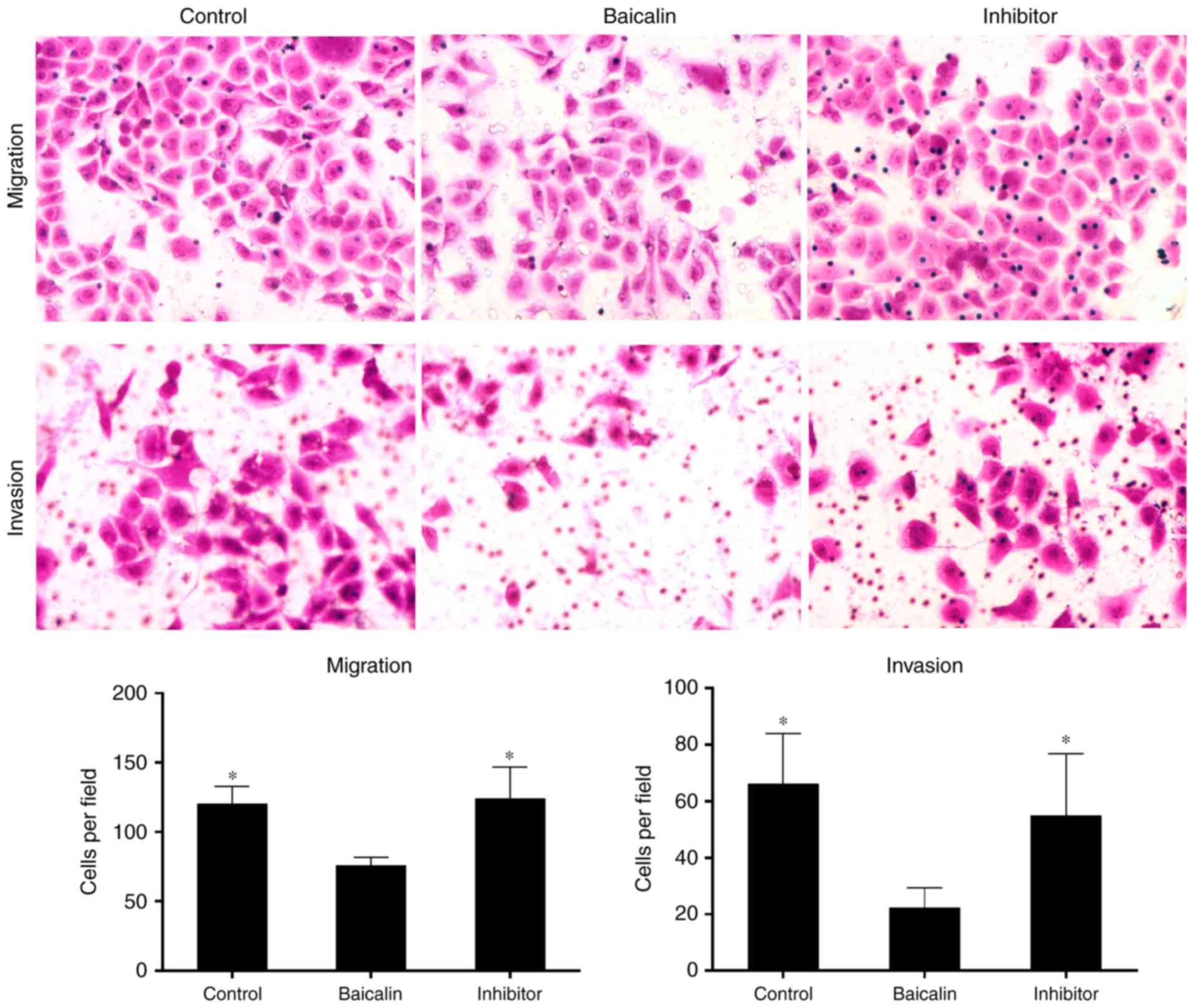
(Fig. 5E). Furthermore, miR-3595
inhibitor recovered the baicalin-induced inhibitory effects on cell
migration and invasion (Fig. 6).
These results suggest that inhibiting miR-3595 significantly
suppressed the anti-fibrotic effect of baicalin.
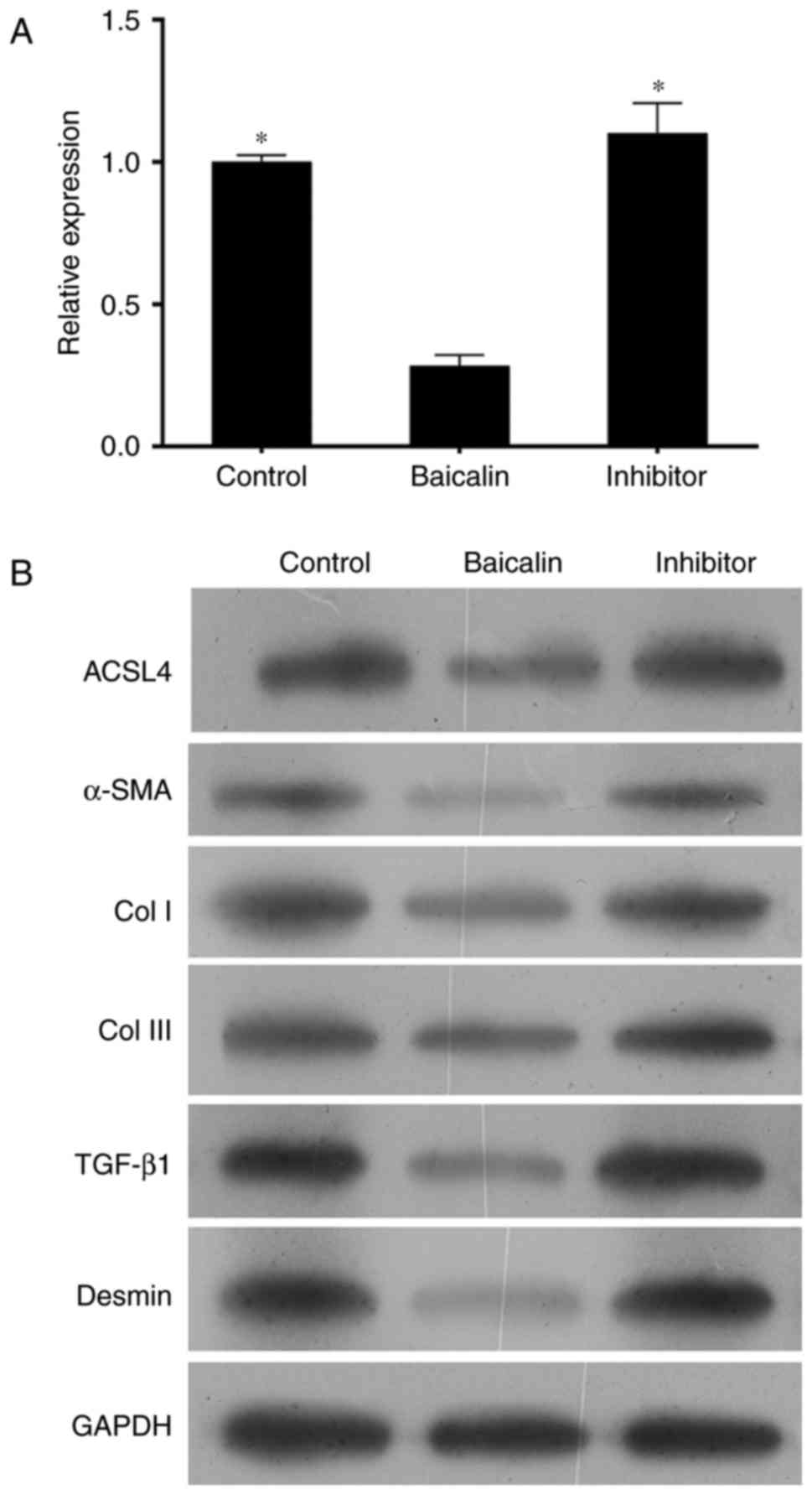
To determine the role of miR-3595 on the
anti-fibrotic effect of baicalin, the protein levels of α-SMA, Col
I, Col III, TGF-β and desmin were measured. As demonstrated in
Fig. 7, the expression levels of
these proteins in activated HCS-T6 cells treated with 100 µM
baicalin and miR-3595 inhibitor were similar to those in the
activated HSC-T6 cells without any treatment, while the expression
levels of these proteins in the baicalin-treated activated HCS-T6
cells were markedly decreased. Taken together, the present results
suggest that miR-3595 regulates the anti-fibrotic effects of
baicalin with respect to cell proliferation, apoptosis, invasion
and migration, and fibrosis-related protein expression levels.
Effect of miR-3595 on baicalin-induced
anti-fibrosis is through the regulation of ACSL4 expression
Previous studies indicate that the expression of
ACSL4 is related to the synthesis of eicosanoid-CoA and the
conversion of arachidonic acid into phosphatidyl ethanolamine,
phosphatidyl inositol and triglyceride (29,30), and that it serves a role in liver
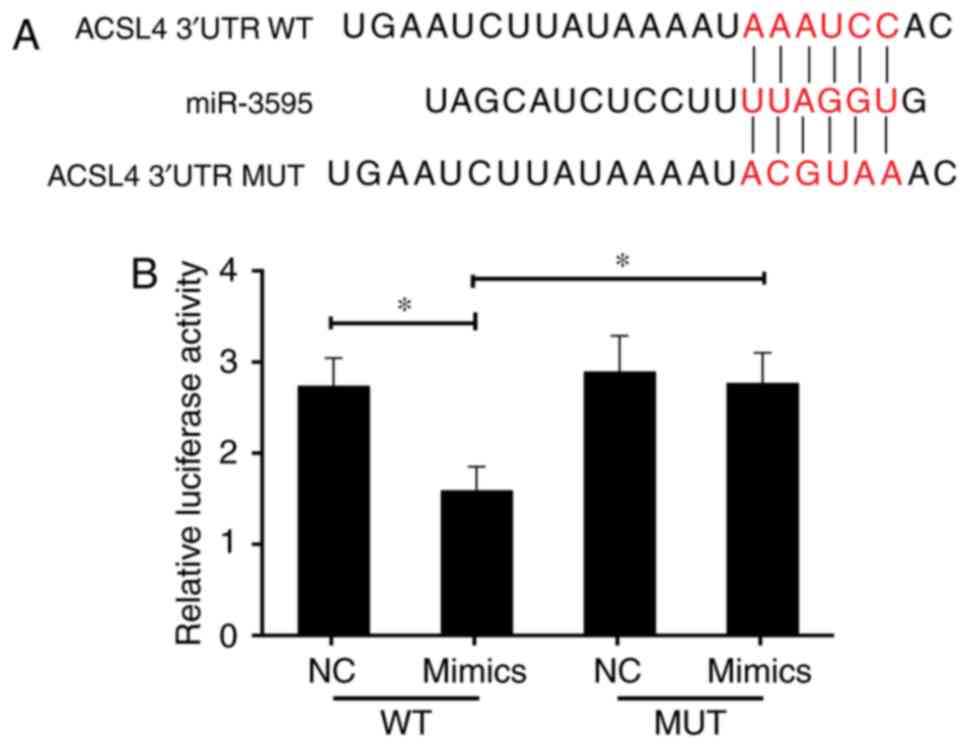
disease. Thus, it was hypothesized that ACSL4 is a direct miR-3595
target. The perfect complementation between the miR-3595 'seed'
sequence and the potential 'recognition' sequences in the 3′UTR of
ACSL4 were observed (Fig. 8A),
suggesting that miR-3595 downregulates ACSL4 expression by
inhibiting the translation of target mRNA. To confirm this
hypothesis, the induced HSC-T6 cells were co-transfected with
ACSL4-3′UTR luciferase reporters and miR-3595 mimics. Results
demonstrated that miR-3595 significantly decreased the luciferase
activity of cells transfected with ACSL4-WT-3′UTR reporter compared
with the level in the NC group (P<0.05); however, transfection
of the ACSL4-MUT-3′UTR had no significant effect on luciferase
activity (Fig. 8B). The mRNA and
protein expression levels of ACSL4 in HSC-T6 cells decreased
significantly on treatment with 100 µM baicalin compared
with the level in the untreated group (P<0.05). These results
indicate that miR-3595 negatively regulates ACSL4. To further
validate these results, ACSL4 was overexpressed via a plasmid in
HSC-T6 cells treated with 100 µM baicalin. Results
demonstrated that the forced expression of ACSL4 attenuated the
anti-fibrotic effect of baicalin by significantly reducing and
apoptosis of drug-treated induced HST-T6 cells (P<0.05), and
increasing the proliferation, invasion and migration of induced
HST-T6 cells (P<0.05), and increasing expression levels of
fibrosis-related proteins (Figs.
9 and 10). Therefore,
miR-3595 targets ACSL4, which is involved in the regulation of the
anti-fibrotic effect of baicalin.
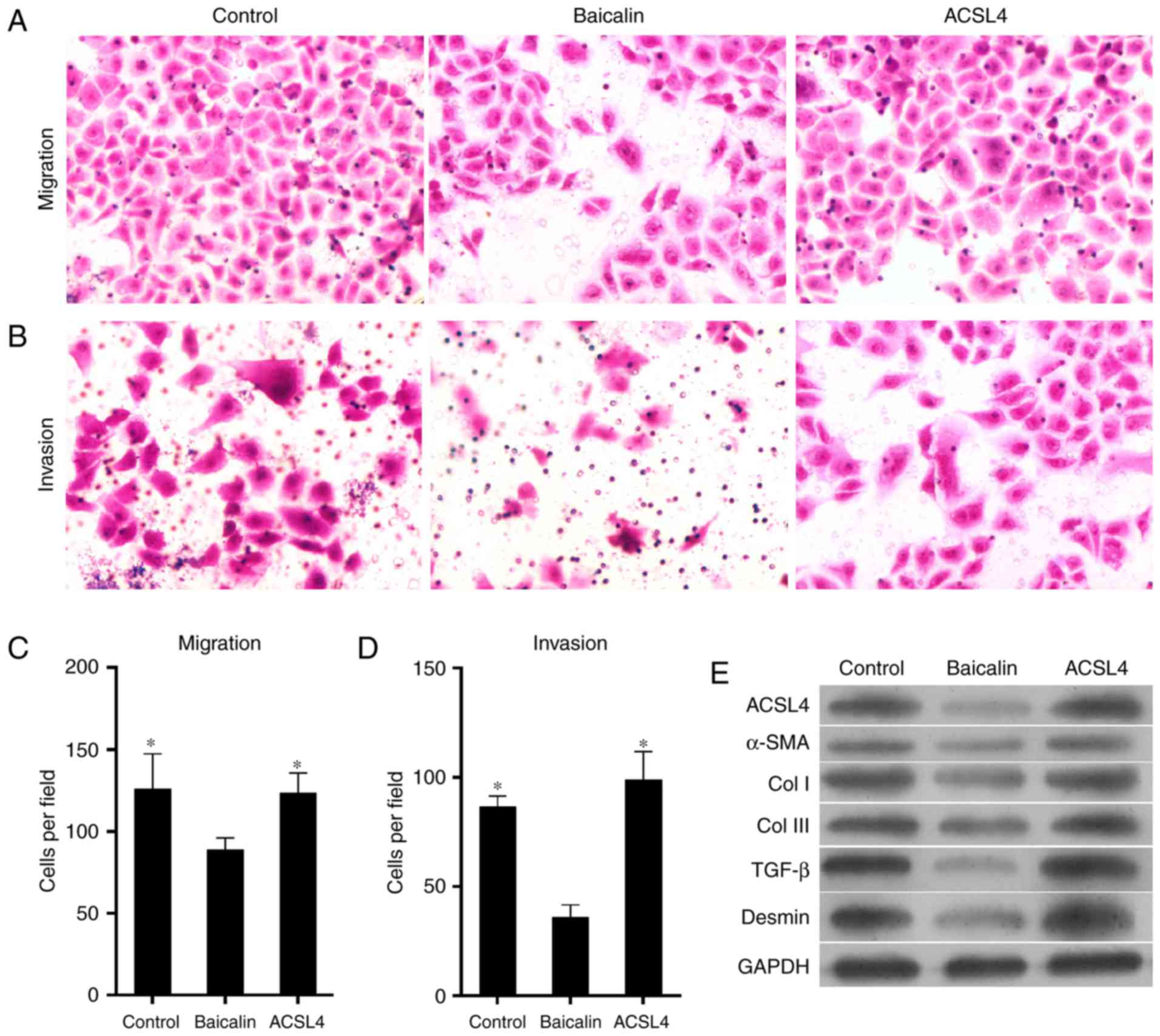 | Figure 10Exogenetic overexpression of ACSL4
attenuates the anti-fibrotic effect of baicalin on cell migration
and invasion, and influences the expression levels of pro-fibrotic
proteins. Platelet-derived growth factor-BB-induced HSC-T6 cells
were treated with 100 µM baicalin. One group of cells was
transfected with ACSL4-overexpressing plasmid in addition to
baicalin. Microscope images of cells undergoing (A) migration and
(B) invasion (magnification, ×400). Representative number of cells
undergoing (C) migration and (D) invasion was counted. (E) Protein
expression levels of fibrosis-promoting proteins, including ACSL4,
α-SMA, Col I, Col III, TGF-β and desmin were verified by western
blotting. *P<0.05 vs. baicalin-treated group. ACSL4,
long-chain-fatty-acid-CoA ligase 4; α-SMA, α-smooth muscle actin;
TGF-β, transforming growth factor-β; Col, collagen. |
Discussion
Liver fibrosis is a wound healing response to a
variety of stimuli (31,32). The progression of liver fibrosis
leads to a number of pathological and biochemical changes, such as
the distortion of normal structural architecture of the liver,
resulting in metabolic abnormalities (33) or hepatocellular carcinoma
(34). Accumulating evidence
suggests that several cell types are involved in the development of
liver fibrosis (35). However,
the activation of HSCs is a particularly critical process in the
pathogenesis of liver fibrosis. Following liver injury, HSCs
increase the expression of α-SMA, c-Myb, and myocyte enhancer
factor-2, and acquire a myofibroblast-like phenotype (35,36). A complex network of intracellular
events occurs during the activation of HSCs (37–43). Thus, one anti-fibrotic therapy is
the modulation of the proliferation, activation and apoptosis of
HSCs.
The ideal anti-fibrotic agent should be
liver-specific and have low hepatotoxicity; however, currently
these agents are not commercially available (33). Hence, natural products and herbal
medicines are potential therapies for hepatic fibrosis. Baicalin is
a bioactive flavonoid with a multitude of pharmacological
activities that include inhibiting the effects of several viruses
(44–46), a variety of inflammatory diseases
(47–49) and various types of cancer
(50). Notably, previous studies
have demonstrated that baicalin serves an important role in the
fibrotic liver by reducing oxidative stress and shifting the
balance of cytokines from pro-fibrotic to anti-fibrotic in
experimental animal models (13,51).
However, the mechanism of the anti-fibrotic effect
of baicalin is not understood. To elucidate the anti-fibrotic
function of baicalin and its possible mechanism, the present study
treated activated HSC-T6 cells induced by PDGF-BB with baicalin at
different concentrations (50, 100 or 150 µM). It was
revealed that baicalin reduced the proliferation, invasion and
migration of these cells. Baicalin also reduced the expression
levels of α-SMA (a marker of activated HSCs), and increased the
number of apoptotic cells, in a dose-dependent manner. These data
suggested that baicalin inhibits the proliferation, apoptosis,
invasion, migration and activation of PDGF-BB-treated HSCs.
miRNA have been implicated in various diseases, such
as liver fibrosis, and several current therapeutic strategies
involve the manipulation of miRNA levels (52). Thus, the present study aimed to
verify whether baicalin regulated the expression of miRNA in
induced HSC-T6 cells. miRNA profiling of baicalin-treated cells and
untreated cells was performed, and the data indicated that some
miRNA were differentially expressed between the two groups. One of
these differentially expressed miRNA was miR-3595. The present
study further confirmed the differential expression of miR-3595
using RT-qPCR. The present study, therefore, demonstrated that
baicalin increases the expression of endogenous miR-3595.
Furthermore, the present study indicated that silencing the
expression of miR-3595 attenuated the anti-fibrotic effect of
baicalin. These results suggested that miR-3595 upregulation serves
an important role in the anti-fibrotic effect of baicalin. In
humans, miRNA regulate gene expression by binding to the 3′UTR of
mRNA or miRNA response elements, resulting in translational
repression (53,54).
The liver is the central organ involved in fatty
acid (FA) homeostasis (55).
ACSL4, a member of the ACSL family, catalyzes the cellular
metabolism of polyunsaturated FA in different tissues (56–58). ACSL4 was recently demonstrated to
have various functions in different cell lines (59). ACSL4 also promotes cell invasion
by regulating the generation of eicosanoids in breast cancer
(60). Through prediction and
experimental validation, the present study indicated that ACSL4 was
a direct target of miR-3595. Indeed, exogenous overexpression of
ACSL4 suppressed the anti-fibrotic effect of baicalin in induced
HSC-T6 cells. However, the relevance of the relationship between
ACSL4 and fibrosis remains unclear.
Thus, it may be speculated that the anti-fibrotic
effect of baicalin is associated with the miR-3593/ACSL4 axis.
Furthermore, the present study identified several differentially
expressed miRNA that could be potential targets of baicalin.
However, it is also possible that the anti-fibrotic effect of
baicalin acts through signaling pathways other than miRNA-related
ones. Baicalin may be a promising chemotherapy drug; however,
further work is required to test this hypothesis.
In summary, the present findings suggest that
baicalin inhibits the proliferation and activation observed in
induced HSCs. One potential mechanism through which baicalin
mediates its anti-fibrotic effect could be mediated via the
increased expression of miR-3595, which leads to a decrease in the
expression of ACSL4. Taken together, the present results
demonstrate that baicalin may be a novel therapeutic in the
treatment of hepatic fibrosis. To the best of our knowledge, the
present study is the first to identify a role of miR-3593/ACSL4 in
regulating liver fibrosis, specifically in the activation of
HSCs.
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Lee UE and Friedman SL: Mechanisms of
hepatic fibrogenesis. Best Pract Res Clinical Gastroenterol.
25:195–206. 2011. View Article : Google Scholar
|
|
2
|
Friedman SL: Mechanisms of hepatic
fibrogenesis. Gastroenterology. 134:1655–1669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Trappoliere M, Caligiuri A, Schmid M,
Bertolani C, Failli P, Vizzutti F, Novo E, di Manzano C, Marra F,
Loguercio C and Pinzani M: Silybin, a component of sylimarin,
exerts anti-inflammatory and anti-fibrogenic effects on human
hepatic stellate cells. J Hepatol. 50:1102–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Popov Y and Schuppan D: Targeting liver
fibrosis: Strategies for development and validation of antifibrotic
therapies. Hepatology. 50:1294–1306. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Friedman SL: Hepatic stellate cells:
Protean, multifunctional, and enigmatic cells of the liver. Physiol
Rev. 88:125–172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Szabo G and Bala S: MicroRNAs in liver
disease. Nat Rev Gastroenterol Hepatol. 10:542–552. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wong L, Yamasaki G, Johnson RJ and
Friedman SL: Induction of beta-platelet-derived growth factor
receptor in rat hepatic lipocytes during cellular activation in
vivo and in culture. J Clin Invest. 94:1563–1569. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu CI, Hoffman JA, Shy BR, Ford EM, Fuchs
E, Nguyen H and Merrill BJ: Function of Wnt/β-catenin in
counteracting Tcf3 repression through the Tcf3-β-catenin
interaction. Development. 139:2118–2129. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takase S, Takada A, Yasuhara M, Sato H and
Matsuda Y: Effects of malotilate treatment on the serum markers of
hepatic fibrogenesis in liver cirrhosis. Gastroenterol Jpn.
23:639–645. 1988.PubMed/NCBI
|
|
11
|
Fu Y, Zheng S, Lin J, Ryerse J and Chen A:
Curcumin protects the rat liver from CCl4-caused injury and
fibrogenesis by attenuating oxidative stress and suppressing
inflammation. Mol Pharmacol. 73:399–409. 2008. View Article : Google Scholar
|
|
12
|
Chen SR, Chen XP, Lu JJ, Wang Y and Wang
YT: Potent natural products and herbal medicines for treating liver
fibrosis. Chin Med. 10:72015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Peng XD, Dai LL, Huang CQ, He CM and Chen
LJ: Correlation between anti-fibrotic effect of baicalin and serum
cytokines in rat hepatic fibrosis. World J Gastroenterol.
15:4720–4725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
de Oliveira MR, Nabavi SF, Habtemariam S,
Erdogan Orhan I, Daglia M and Nabavi SM: The effects of baicalein
and baicalin on mitochondrial function and dynamics: A review.
Pharmacol Res. 100:296–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang MD, Chiang YM, Higashiyama R, Asahina
K, Mann DA, Mann J, Wang CC and Tsukamoto H: Rosmarinic acid and
baicalin epigenetically derepress peroxisomal
proliferator-activated receptor gamma in hepatic stellate cells for
their antifibrotic effect. Hepatology. 55:1271–1281. 2012.
View Article : Google Scholar :
|
|
16
|
Zhu QJ: Research advances on baicalin and
baicalein as potential therapeutic agents for fibrotic disease.
Zhongguo Zhong Yao Za Zhi. 42:1271–1276. 2017.In Chinese.
PubMed/NCBI
|
|
17
|
Meng F, Wang K, Aoyama T, Grivennikov SI,
Paik Y, Scholten D, Cong M, Iwaisako K, Liu X, Zhang M, et al:
Interleukin-17 signaling in inflammatory, Kupffer cells, and
hepatic stellate cells exacerbates liver fibrosis in mice.
Gastroenterology. 143:765–776e3. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kong X, Feng D, Wang H, Hong F, Bertola A,
Wang FS and Gao B: Interleukin-22 induces hepatic stellate cell
senescence and restricts liver fibrosis in mice. Hepatology.
56:1150–1159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight. Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Teng KY and Ghoshal K: Role of noncoding
RNAs as biomarker and therapeutic targets for liver fibrosis. Gene
Expr. 16:155–162. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lewin TM, Kim JH, Granger DA, Vance JE and
Coleman RA: Acyl-CoA synthetase isoforms 1,4 and 5 are present in
different subcellular membranes in rat liver and can be inhibited
independently. J Biol Chem. 276:24674–24679. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lewin TM, Van Horn CG, Krisans SK and
Coleman RA: Rat liver acyl-CoA synthetase 4 is a
peripheral-membrane protein located in two distinct subcellular
organelles, peroxisomes, and mitochondrial-associated membrane.
Arch Biochem Biophys. 404:263–270. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yan S, Yang XF, Liu HL, Fu N, Ouyang Y and
Qing K: Long-chain acyl-CoA synthetase in fatty acid metabolism
involved in liver and other diseases: An update. World J
Gastroenterol. 21:3492–3498. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang YC, Wu CH, Chu JS, Wang CK, Hung LF,
Wang YJ, Ho YS, Chang JG and Lin SY: Involvement of fatty acid-CoA
ligase 4 in hepatocellular carcinoma growth: Roles of cyclic AMP
and P38 mitogen-activated protein kinase. World J Gastroenterol.
11:2557–2563. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin X, Kong LN, Huang C, Ma TT, Meng XM,
He Y, Wang QQ and Li J: Hesperetin derivative-7 inhibits
PDGF-BB-induced hepatic stellate cell activation and proliferation
by targeting Wnt/β-catenin pathway. Int Immunopharmacol.
25:311–320. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Acloque H, Adams MS, Fishwick K,
Bronner-Fraser M and Nieto MA: Epithelial-mesenchymal transitions:
The importance of changing cell state in development and disease. J
Clin Invest. 119:1438–1449. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Golej DL, Askari B, Kramer F, Barnhart S,
Vivekanandan-Giri A, Pennathur S and Bornfeldt KE: Long-chain
acyl-CoA synthetase 4 modulates prostaglandin E(2) release from
human arterial smooth muscle cells. J Lipid Res. 52:782–793. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cooke M, Orlando U, Maloberti P, Podestá
EJ and Cornejo Maciel F: Tyrosine phosphatase SHP2 regulates the
expression of acyl-CoA synthetase ACSL4. J Lipid Res. 52:1936–1948.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schuppan D: Structure of the extracellular
matrix in normal and fibrotic liver: Collagens and glycoproteins.
Semin Liver Dis. 10:1–10. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Anthony PP, Ishak KG, Nayak NC, Poulsen
HE, Scheuer PJ and Sobin LH: The morphology of cirrhosis
Recommendations on definition, nomenclature, and classification by
a working group sponsored by the World Health Organization. J Clin
Pathol. 31:395–414. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mormone E, George J and Nieto N: Molecular
pathogenesis of hepatic fibrosis and current therapeutic
approaches. Chem Biol Interact. 193:225–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han YP, Zhou L, Wang J, Xiong S, Garner
WL, French SW and Tsukamoto H: Essential role of matrix
metalloproteinases in interleukin-1-induced myofibroblastic
activation of hepatic stellate cell in collagen. J Biol Chem.
279:4820–4828. 2004. View Article : Google Scholar
|
|
35
|
Gressner AM: Transdifferentiation of
hepatic stellate cells (Ito cells) to myofibroblasts: A key event
in hepatic fibrogenesis. Kidney Int. (Suppl 54): S39–S45. 1996.
|
|
36
|
Woodhoo A, Ir uar rizaga-Lejar reta M,
Beraza N, García-Rodríguez JL, Embade N, Fernández-Ramos D,
Martínez-López N, Gutiérrez-De Juan V, Arteta B, Caballeria J, et
al: Human antigen R contributes to hepatic stellate cell activation
and liver fibrosis. Hepatology. 56:1870–1882. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fickert P, Fuchsbichler A, Moustafa T,
Wagner M, Zollner G, Halilbasic E, Stöger U, Arrese M, Pizarro M,
Solís N, et al: Farnesoid X receptor critically determines the
fibrotic response in mice but is expressed to a low extent in human
hepatic stellate cells and periductal myofibroblasts. Am J Pathol.
175:2392–2405. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sharvit E, Abramovitch S, Reif S and Bruck
R: Amplified inhibition of stellate cell activation pathways by
PPAR-gamma, RAR and RXR agonists. PloS One. 8:e765412013.
View Article : Google Scholar
|
|
39
|
Ding N, Yu RT, Subramaniam N, Sherman MH,
Wilson C, Rao R, Leblanc M, Coulter S, He M, Scott C, et al: A
vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic
response. Cell. 153:601–613. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li T, Eheim AL, Klein S, Uschner FE, Smith
AC, Brandon-Warner E, Ghosh S, Bonkovsky HL, Trebicka J and Schrum
LW: Novel role of nuclear receptor Rev-erbα in hepatic stellate
cell activation: Potential therapeutic target for liver injury.
Hepatology. 59:2383–2396. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Beaven SW, Wroblewski K, Wang J, Hong C,
Bensinger S, Tsukamoto H and Tontonoz P: Liver X receptor signaling
is a determinant of stellate cell activation and susceptibility to
fibrotic liver disease. Gastroenterology. 140:1052–1062. 2011.
View Article : Google Scholar :
|
|
42
|
Zhu L, Asahina K, Wang J, Ueno A, Lazaro
R, Miyaoka Y, Miyajima A and Tsukamoto H: Hepatic stellate
cell-derived delta-like homolog 1 (DLK1) protein in liver
regeneration. J Biol Chem. 287:10355–10367. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Delgado I, Carrasco M, Cano E, Carmona R,
García-Carbonero R, Marín-Gómez LM, Soria B, Martín F, Cano DA,
Muñoz-Chápuli R and Rojas A: GATA4 loss in the septum transversum
mesenchyme promotes liver fibrosis in mice. Hepatology.
59:2358–2370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Baylor NW, Fu T, Yan YD and Ruscetti FW:
Inhibition of human T cell leukemia virus by the plant flavonoid
baicalin (7-glucuronic acid, 5,6-dihydroxyflavone). J Infect Dis.
165:433–437. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li BQ, Fu T, Dongyan Y, Mikovits JA,
Ruscetti FW and Wang JM: Flavonoid baicalin inhibits HIV-1
infection at the level of viral entry. Biochem Biophys Res Commun.
276:534–538. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zeng Y, Song C, Ding X, Ji X, Yi L and Zhu
K: Baicalin reduces the severity of experimental autoimmune
encephalomyelitis. Braz J Med Biol Res. 40:1003–1010. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lin CC and Shieh DE: The anti-inflammatory
activity of Scutellaria rivularis extracts and its active
components, baicalin, baicalein and wogonin. Am J Chin Med.
24:31–36. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kubo M, Matsuda H, Tanaka M, Kimura Y,
Okuda H, Higashino M, Tani T, Namba K and Arichi S: Studies on
Scutellariae radix. VII. Anti-arthritic and anti-inflammatory
actions of methanolic extract and flavonoid components from
Scutellariae radix. Chem Pharm Bull (Tokyo). 32:2724–2729. 1984.
View Article : Google Scholar
|
|
49
|
Zhang XP, Tian H, Lai YH, Chen L, Zhang L,
Cheng QH, Yan W, Li Y, Li QY, He Q and Wang F: Protective effects
and mechanisms of Baicalin and octreotide on renal injury of rats
with severe acute pancreatitis. World J Gastroenterol.
13:5079–5089. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ikezoe T, Chen SS, Heber D, Taguchi H and
Koeffler HP: Baicalin is a major component of PC-SPES which
inhibits the proliferation of human cancer cells via apoptosis and
cell cycle arrest. Prostate. 49:285–292. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sun H, Che QM, Zhao X and Pu XP:
Antifibrotic effects of chronic baicalein administration in a CCl4
liver fibrosis model in rats. Eur J Pharmacol. 631:53–60. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hyun J and Jung Y: MicroRNAs in liver
fibrosis: Focusing on the interaction with hedgehog signaling.
World J Gastroenterol. 22:6652–6662. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Akhtar MM, Micolucci L, Islam MS, Olivieri
F and Procopio AD: Bioinformatic tools for microRNA dissection.
Nucleic Acids Res. 44:24–44. 2016. View Article : Google Scholar :
|
|
54
|
Ambros V: microRNAs: Tiny regulators with
great potential. Cell. 107:823–826. 2001. View Article : Google Scholar
|
|
55
|
Babin PJ and Gibbons GF: The evolution of
plasma cholesterol: Direct utility or a 'spandrel' of hepatic lipid
metabolism. Prog Lipid Res. 48:73–91. 2009. View Article : Google Scholar
|
|
56
|
Soupene E and Kuypers FA: Mammalian
long-chain acyl-CoA synthetases. Exp Biol Med (Maywood).
233:507–521. 2008. View Article : Google Scholar
|
|
57
|
Lopes-Marques M, Cunha I, Reis-Henriques
MA, Santos MM and Castro LF: Diversity and history of the
long-chain acyl-CoA synthetase (Acsl) gene family in vertebrates.
BMC Evol Biol. 13:2712013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kang MJ, Fujino T, Sasano H, Minekura H,
Yabuki N, Nagura H, Iijima H and Yamamoto TT: A novel
arachidonate-preferring acyl-CoA synthetase is present in
steroidogenic cells of the rat adrenal, ovary, and testis. Proc
Natl Acad Sci USA. 94:2880–2884. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Doll S, Proneth B, Tyurina YY, Panzilius
E, Kobayashi S, Ingold I, Irmler M, Beckers J, Aichler M, Walch A,
et al: ACSL4 dictates ferroptosis sensitivity by shaping cellular
lipid composition. Nat Chem Biol. 13:91–98. 2017. View Article : Google Scholar :
|
|
60
|
Orlando UD, Garona J, Ripoll GV, Maloberti
PM, Solano ÁR, Avagnina A, Gomez DE, Alonso DF and Podestá EJ: The
functional interaction between Acyl-CoA synthetase
4,5-lipooxygenase and cyclooxygenase-2 controls tumor growth: A
novel therapeutic target. PloS One. 7:e407942012. View Article : Google Scholar
|