Introduction
Bone is a dynamic tissue with a relatively high
turnover. The homeostasis of bone is tightly regulated by its
constant formation and degradation to maintain its proper function
and ensure adaptation to the load of the skeleton. Once this
balance is disturbed, it may lead to bone sclerosis or
osteoporosis. Osteoporosis is currently a major socioeconomic
burden (1). Reduced bone content
caused by various factors accelerates the degradation of bone
microstructure and leads to bone brittleness. Bone density is
reduced, leading to systemic skeletal symptoms, such as severe
pain, multiple fractures, and potentially life-threatening
events.
Osteoporosis may be classified as primary
osteoporosis, which is observed in postmenopausal women, and
secondary osteoporosis, which may be caused by several factors,
such as malnutrition, endocrine problems, diabetes, genetics and
medications, among which glucocorticoid (GC)-induced osteoporosis
(GIO) is considered to be the most common secondary iatrogenic type
of osteoporosis (2,3). Even a daily dose as low as 2.5 mg
prednisone increases the risk of osteoporotic fractures, whereas
inhaled GC therapy is also associated with bone loss. In extreme
cases, a high dose may ultimately lead to systemic side effects,
such as bone loss and osteoporotic osteonecrosis. It was previously
reported that patients exhibited progressive loss of bone mass
several weeks post-GC therapy. The patients’ bone mass was found to
decline rapidly within the first few months, reaching 5–15% in
total per year, and long-term GC therapy (>1 year) was
associated with an osteoporosis incidence rate is as high as 30–50%
(4). Along with the occurrence of
osteoporosis, osteoporotic fractures may occur, most commonly
observed in the spinal vertebral body, proximal femur and ribs.
Although the awareness of osteoporosis has increased in the general
public, the severity of the side effects associated with the
clinical use of GCs requires further elucidation; furthermore, the
complex interplay between GCs and altered bone homeostasis is a
subject of intense clinical interest. GC-induced osteogenic
dysfunction of osteoblasts is considered to be one of the main
pathological mechanisms underlying the development of GIO,
including inhibition of osteoblast proliferation and osteogenic
differentiation, or even induction of adipogenic differentiation
and cell apoptosis (5). Natural
medicines have been a subject of interest in recent years, as they
are well known to exert diverse biological effects and improve bone
quality. The aim of the present study was to investigate the
therapeutic efficacy of gastrodin (GSTD), a natural medicinal
extract, in reducing the adverse effects of high-dose dexamethasone
(DEX) on MC3T3-E1 osteoblasts.
GSTD, also referred to as gastrodia glycosides, has
been isolated from the traditional Chinese herbal agent
Gastrodia elata and confirmed to be one of the major active
components of Rhizoma gastrodiae. The molecular structure of
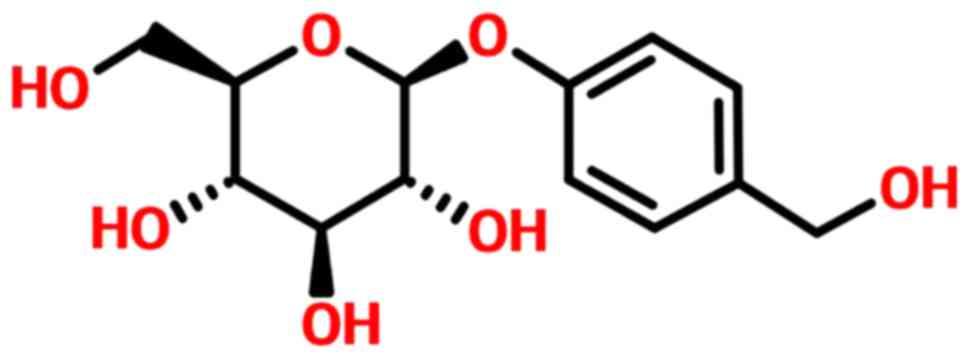
GSTD is shown in Fig. 1, and its
medicinal properties include smooth muscle-relaxing, anti-necrotic,
anti-aging and anti-apoptotic effects (6,7).
GSTD effectively lowers lipid peroxidation levels, removes
oxygen-free radicals, exerts antioxidant effects, reduces coupling
oxidative phosphorylation and increases the levels of
malondialdehyde and superoxide dismutase, as well as the expression
levels of related genes and proteins (8,9).
Therefore, the main objective of the present study was to improve
our knowledge of the therapeutic value of GSTD by examining its
potential application as an inhibitor of GC-induced dysfunction of
MC3T3-E1 cells. Furthermore, the possible molecular mechanisms
underlying the effects of GSTD on MC3T3-E1 cell proliferation and
differentiation under DEX treatment were investigated.
Regulatory transcription factor nuclear factor-like
2 (NRF2) is a transcription activator that can combine with
antioxidant responsive element (ARE) and enhance the expression of
antioxidant enzymes, including NAD(P)H:quinone oxidoreduc-tase-1
(NQO-1) and heme oxygenase-1 (HO-1) (10). NRF2 acts as an important cell
protective mechanism against exogenous or endogenous noxious
stimuli (11), and its expression
is closely correlated with cell differentiation (12). In recent years, there have been
studies indicating that certain natural products, such as
sulforaphane, and bioactive compounds referred to as
avenanthramides, may block the cytotoxic effects of high-dose DEX
through activation of the NRF2 pathway and osteogenic
differentiation, as well as inhibition of the apoptosis-related
caspase protein family in GIO (13,14). However, considering the safety
profile and long history of these drugs or extracts, new
pharmaceuticals must be developed and approved. It was previously
reported that GSTD may activate NRF2 signaling in liver, nerve, and
vascular smooth muscle cells, playing a key role in antioxidation
and anti-inflammation. GSTD may also be used in human bone marrow
mesenchymal stem cells to promote osteogenesis by reducing
osteoclast differentiation (13,15). GSTD may inhibit the key adipogenic
differentiation factor peroxisome proliferator-activated receptor
(PPAR)γ, reducing fat content in hypertensive model rats (16).
Bone formation largely depends on the proliferation
and differentiation of osteoblasts and is required for skeletal
development. It is well known that GCs suppress bone formation by
attenuating osteoblast viability and differentiation (17). The pathophysiology of GIO is
complex, and GCs modify bone mass mainly by directly affecting
osteoblasts (18). Whether GSTD
can activate NRF2 and its downstream transcription factors to
reduce the excessive stimulation induced by DEX in osteoblasts and
reduce GC-induced osteoblast injury requires further investigation.
Our results demonstrated that GSTD effectively promoted osteogenic
differentiation in MC3T3-E1 cells through regulation of the NRF2
pathway. This study may provide a novel strategy for the prevention
of GCs-induced osteoporosis by using natural products.
Materials and methods
Reagents
Purified GSTD (>98%) was purchased from the
National Institute for the Control of Pharmaceutical and Biological
Products (Dalian, China), dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA, St. Louis, MO, USA) and stored at −20°C.
The final concentrations of GSTD were 0 (control), 1 and 5
µM, and the final concentration of DMSO in the culture was
<0.01%. Minimum essential medium alpha modification (α-MEM),
fetal bovine serum (FBS) and trypsin-EDTA were obtained from GE
Healthcare Life Sciences HyClone Laboratories (Logan, UT, USA). The
Cell Counting Kit-8 (CCK-8) was purchased from Sigma-Aldrich; Merck
KGaA. The Annexin V-FITC apoptosis detection kit was obtained from
Beyotime Institute of Biotechnology (Shanghai, China). Rabbit
anti-HO-1 (cat. no. ab68477; 1:1,000), rabbit anti-NQO-1 (cat. no.
ab76956; 1:1,000), and mouse anti-β-actin (cat. no. ab8245;
1:1,000), Anti-OCN (cat. no. ab93876; 1:1,000) monoclonal
antibodies were purchased from Abcam (Cambridge, MA, USA). Rabbit
anti-NRF2 (cat. no. 12721; 1:1,000), rabbit anti-Runx2 (cat. no.
20056S; 1:1,000), and rabbit anti-PPARγ (cat. no. 2430; 1:1,000)
monoclonal antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). TRIzol reagent was obtained
from Thermo Fisher Scientific, Inc. (Carlsbad, CA, USA). Primers
were designed and synthesized by Sangon Biotech Co., Ltd.
(Shanghai, China).
Cell culture
MC3T3-E1 cells were maintained in α-MEM (GE
Healthcare Life Sciences HyClone Laboratories) supplemented with a
10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin at
37°C with 5% CO2 in a humidified atmosphere. Cells in
the exponential phase of growth were selected for the experiments.
For osteogenic differentiation, cells were cultured in inducing
growth medium supplemented with 100 nM DEX, 10 mM
β-glycerophosphate and 50 mg/ml ascorbic acid (Sigma-Aldrich; Merck
KGaA), replenished every 3 days.
Cell viability assay
Cell viability was assessed by the CCK-8 assay,
which is used for detecting cell proliferation with a high
sensitivity. Briefly, MC3T3-E1 cells were seeded at a density of
5×103 cells/well in 96-well plates and incubated in the
growth media for 24 h at 37°C. Subsequently, the cells were exposed
to different concentrations of DEX (0, 1, 5, 25, 50, 100 and 200
µM) and GSTD (0, 0.1, 1, 5, 10, 50 and 100 µM) for 24
h. Furthermore, the cells were also pretreated with GSTD (1 and 5
µM) for 2 h, and then exposed to DEX (200 µM) for 24
h. Subsequently, 10 µl CCK-8 were added to each well and
incubation was continued for 2 h. The optical density was measured
at 450 nm on a microplate reader with a microplate
spectrophotometer (ELX-800; BioTek, VT, USA) and the absorbance of
each well was recorded. The ratio of the mean absorbance was
considered to reflect the relative cellular viability.
Alkaline phosphatase (ALP) activity
assay
To examine the ALP activity in MC3T3-E1 cells, the
cells were seeded into 12-well plates at a density of
5×104 cells/well in different drug-treated
osteogenic-inducing media. The cells were washed twice, harvested
and lysed with 100 µl assay lysis buffer (Beyotime Institute
of Biotechnology) after 7 days of culture. The ALP activity levels
(U/ml) were measured with an ALP reagent kit (Nanjing Jiancheng
Bioengineering Research Institute, Nanjing, China), according to
the manufacturer’s instructions. All samples were examined in
triplicate.
Flow cytometric analysis of osteoblast
apoptosis
MC3T3-E1 cells were incubated in 6-well plates at a
density of 2×105 cells/well for 24 h. The cells were
then exposed to DEX in the presence or absence of different
concentrations of GSTD for indicated time periods. The cells were
harvested and resuspended in 300 µl binding buffer
containing 5 µl Annexin V-FITC and 5 µl propidium
iodide (PI), then rinsed with phosphate-buffered saline (PBS).
After washing twice with PBS, the cells were placed in an ice bath
for 0.5 h and the samples were analyzed by FACScan flow
cytometry.
Determination of mitochondrial outer
membrane potential (MOMP)
The lipophilic cationic fluorescent dye
5,5′,6,6′-tera-chloro-1,1′,3,3′-tetraethylbenzimidazol-carbocyanine
iodide (JC-1) was used to detect the changes in MOMP (Δψm) as an
indicator of mitochondrial function. Carbonyl cyanide
3-chlo-rophenylhydrazone (CCCP 1.0 mM, 2 h; Sigma-Aldrich; Merck
KGaA) was used as positive control. Cells (2×105
cells/well) were cultured in 6-well plates. The groups were as
previously designed, treated with different concentrations of GSTD
and DEX for 24 h. Measurements were performed using a MOMP
detection kit (Beyotime Institute of Biotechnology) according to
the manufacturer’s instructions. In brief, the cells were collected
and then incubated with the MOMP-sensitive fluorescent dye JC-1 for
20 min at 37°C, washed twice in PBS, and subjected to flow
cytometry.
Mineralization assay
Preosteoblast MC3T3-E1 cells were cultured in
complete growth medium in 35-mm dishes. When the cells reached 80%
confluence, mineralization was induced by culturing confluent cells
in growth medium supplemented with 10 mM β-glycerophosphate and 50
mg/ml ascorbic acid (Sigma-Aldrich; Merck KGaA). Inducing media
were replenished every 3 days. After 21 days, the cells were washed
with PBS and fixed in 90% ethanol (Sigma-Aldrich; Merck KGaA) at
room temperature for 30 min. After rinsing with distilled water,
the cells were stained with 2 ml of 10 mM Alizarin Red S (pH 4.2)
per dish at room temperature for 15 min while being gently swayed.
Following aspiration of the unincorporated dye, the cells were
washed four times with 2 ml distilled water to exclude non-specific
staining. Mineralized nodules and stained cells were visualized and
photographed with a phase contrast microscope (Zeiss, Oberkochen,
Germany).
Quantification of gene expression by
quantitative polymerase chain reaction (qPCR)
Cells were cultured in osteogenic-inducing media
with drug treatment for 2 days. Total RNA was extracted with TRIzol
reagent, then used to synthesize cDNA using SuperScript II reverse
transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.) with 5
µg oligo(dT) primers per sample. By using SYBR-Green PCR
master mix (Applied Biosystems; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), qPCR was performed in a total volume of 20
µl in a 7500 Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) as follows: 95°C for 5 min,
followed by 40 cycles of 95°C for 30 sec and 60°C for 45 sec.
Melting curve analysis was used to confirm the specificity of the
amplification and glyceraldehyde 3-phosphate dehydrogenase (GAPDH)
served as the endogenous control for normalization of the amount of
total RNA in each group. The relative levels of gene expression
were calculated as follows: ΔCq = Cqgene −
Cqreference; gene expression was calculated as fold
change according to the 2−ΔΔCq method and measurements
were repeated independently in triplicate. The primer sequences
were as follows: Forward, 5′-CTGACCACCTGAACTCCAC-3′ and reverse,
5′-CATCTAGGTACAACATGGAG-3′ for BMP-2; forward,
5′-GAATGCACTACCCAGCCAC-3′ and reverse, 5′-TGGCAGGTACGTGTGGTAG-3′
for Runx2; forward, 5′-GTCAAGAGTCTTAGCCAAACTC-3′ and reverse,
5′-AAATGATGTGAGGCCAGATGG-3′ for osterix (OSX); forward,
5′-CAATAAGGTAGTGAACAGAC-3′ and reverse, 5′-CTTCAAGCCATACTGGTCT-3′
for osteocalcin (OCN); and forward, 5′-GTGAAGCAGGCATCTGAGGG-3′ and
reverse, 5′-GCCGTATTCATTGTCATACCAGG-3′ for GAPDH.
Western blot analysis
Total protein from each well was harvested in
ice-cold radioimmunoprecipitation (RIPA) lysis buffer (Thermo
Fisher Scientific, Inc.) supplemented with phenylmethanesulfonyl
fluoride for 1 h. The protein concentration was quantified using
the bicinchoninic acid protein assay kit (Sigma-Aldrich; Merck
KGaA) according to the manufacturer’s instructions. Equal proteins
of each treatment were separated on 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (Beyotime Institute of
Biotechnology) and electrophoretically transferred onto
polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA).
The membranes were soaked in 5% skimmed milk as blocking buffer for
1 h, then washed 3 times in Tris-buffered saline Tween-20 [TBST;
150 mM NaCl (pH 7.5), 20 mM Tris-HCl and 0.1% Tween-20] at room
temperature. The membranes were incubated with primary monoclonal
antibodies against Runx2, NRF2, NQO-1, HO-1, PPARγ and caspase-3 at
1:1,000 dilution overnight at 4°C followed by hybridization with
horseradish peroxidase-conjugated secondary antibody (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) and visualized using
enhanced chemiluminescence. The relative protein levels were
calculated based on β-actin as the loading control.
Animals and drug supplementation
A total of 32 female experimental Sprague Dawley
rats, aged 8 weeks and weighing 250±22 g, were obtained from the
Animal Center of Shengjing Hospital of China Medical University
(Shenyang, China). The rats were acclimatized to specific
pathogen-free laboratory conditions (a well-ventilated controlled
room at 20°C on a 12-h light/dark cycle with free access to water
and food) for 1 week prior to the drug treatments. All animal care
and experimental procedures were approved by the Institutional
Animal Care Ethics and Use Committee of China Medical University,
and performed in accordance with the guidelines. The rats were
randomly divided into 4 groups: Control, DEX and DEX with GSTD (1
and 5 mg/kg/day) groups. A GIO rat model (n=8) was induced by
intramuscularly injected DEX (1 mg/kg/day DEX for 60 days), while
the control group (n=8) was administered an equivalent volume of
normal saline. The GSTD therapeutic groups (n=16) received
different doses of GSTD (1 and 5 mg/kg/day) by gavage
simultaneously with induction of the model by DEX. All the
procedures lasted for 60 days to ensure the experimental modeling
of the animals until bone samples (bilateral femora) were removed
for further analysis.
Histology and bone mineral density (BMD)
measurement
Following completion of animal modeling, the left
femora were removed and placed in 4% paraformaldehyde for 48 h
prior to demineralization for 30 days in 10% EDTA solution that was
replenished every 2 days. The femora were then processed in
paraffin and were serially cut sagittally into 5-µm sections
for hematoxylin and eosin (H&E) staining (Beyotime Institute of
Biotechnology). The microstructure photograph and analysis were
conducted in the proximal metaphysis adjacent to the epiphyseal
growth plate. The right femora were directly used for BMD
measurement by dual X-ray absorptiometry using a PIXImus II
densitometer (GE Medical Systems, Lunar Division, Madison, WI, USA)
and data were recorded for further analysis. The measurements were
limited to the proximal femoral area.
Statistical analysis
All the presented data and results were processed
using GraphPad Prism 6.01 software and expressed as mean ± standard
deviation of at least three independent experiments. One-way
analysis of variance was used to determine statistical
significance. P<0.05 or P<0.01 were considered to indicate
statistically significant differences.
Results
GSTD pretreatment maintains MC3T3-E1 cell
viability and ALP activity while exposed to high-dose DEX
To determine whether GSTD exerted protective effects
on MC3T3-E1 cells against DEX-induced reduced viability, the
effects of GSTD and DEX or combined effect of both drugs on
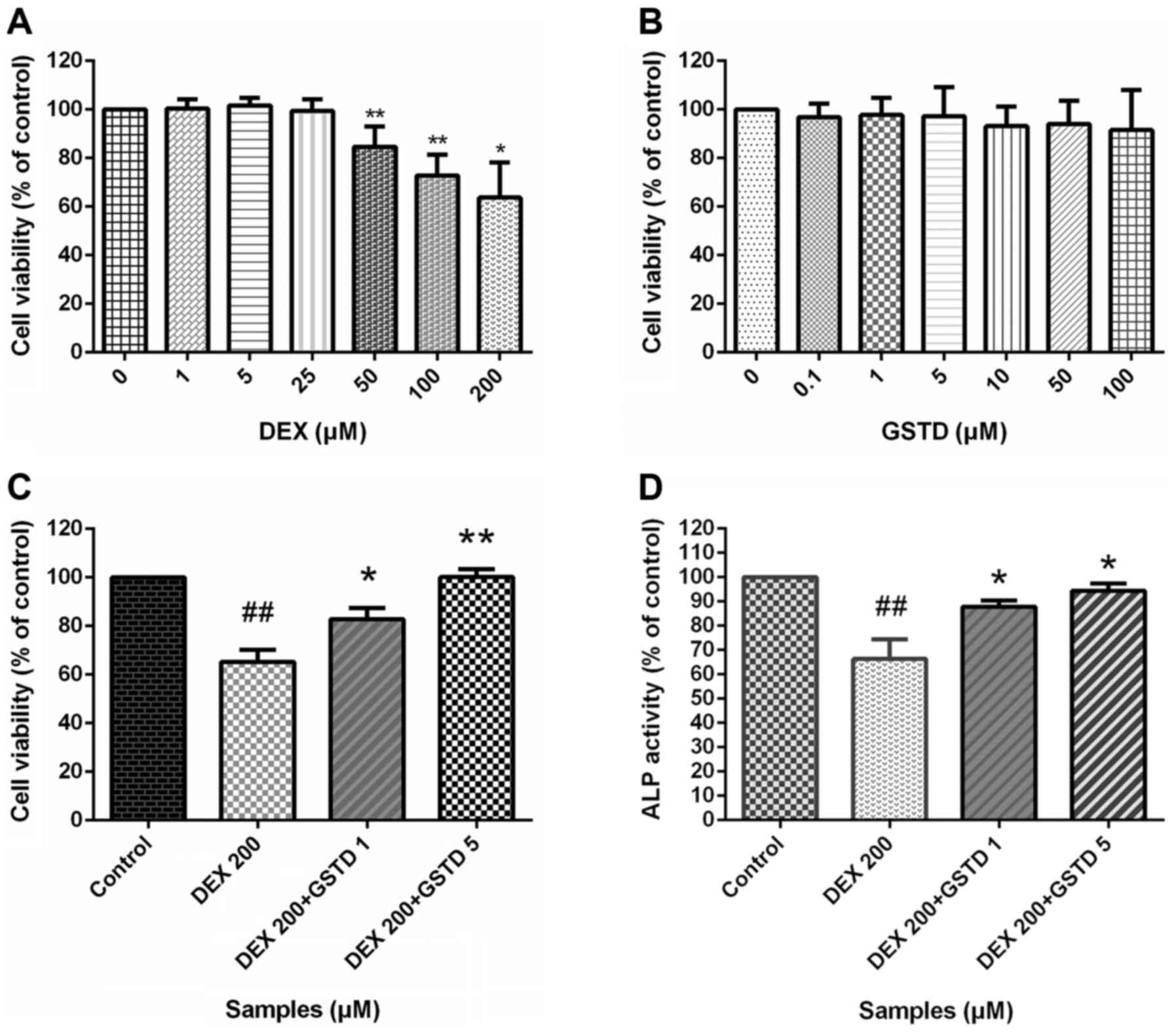
MC3T3-E1 cell viability were evaluated by the CCK-8 assay (Fig. 2). The CCK-8 assay indicated that
GSTD affected the MC3T3-E1 cell viability marginally at
concentrations of ≤100 µM (Fig. 2B). Cell viability exhibited a
noticeable decrease with DEX treatment at a concentration of ≥50
µM (Fig. 2A), and 200
µM was selected as the concentration in the DEX-induced
decreased cell viability model, while GSTD at different
concentrations (1 and 5 µM) was added to relieve this effect
in a dose-dependent manner, indicating that GSTD was able to
partially inhibit high-dose DEX-induced cytotoxicity and exerted a
protective effect, although it did not entirely reverse the toxic
effect.
ALP is a well-recognized indicator of osteoblast
differentiation (19). ALP
activity was examined to evaluate the extent of osteoblast
differentiation. ALP activity exhibited significant differences
between treatment with DEX and GSTD, alone or combined (Fig. 2D). As expected, DEX treatment
significantly decreased ALP activity, while GSTD at 1 or 5
µM was found to increase ALP activity against DEX by ~30%;
these findings indicate that GSTD also enhanced the activity of ALP
compared with the DEX-treated group, suggesting that GSTD
stimulated osteoblast differentiation, even under the inhibitory
influence of DEX.
GSTD protects MC3T3-E1 cells from
apoptosis
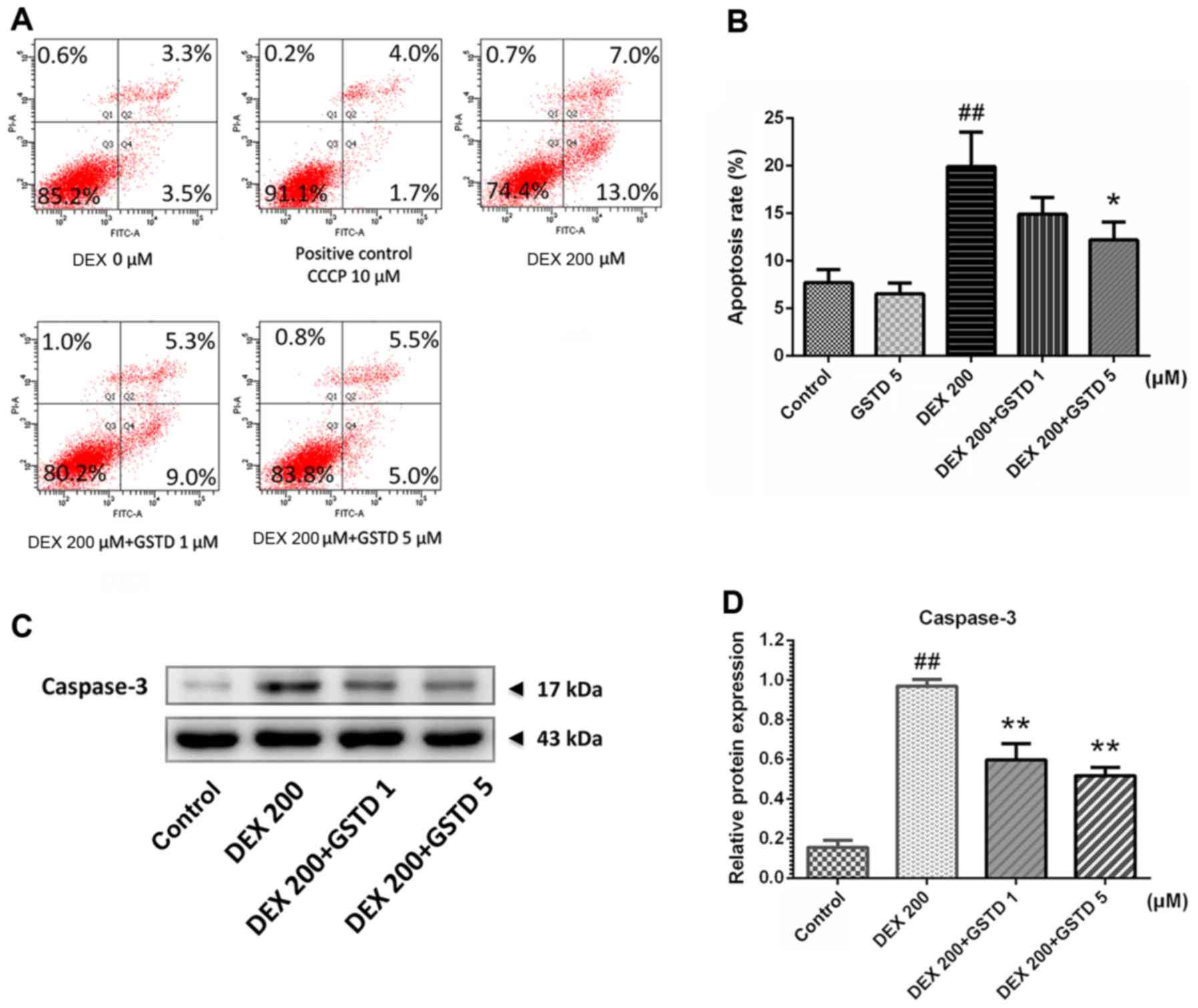
Annexin V-FITC/PI flow cytometric analysis was
performed to determine cell apoptosis (Fig. 3). According to the results, DEX
treatment markedly induced cell apoptosis, as previously reported
(20), which was validated with a
concentration of 200 µM DEX, while combined pretreatment
with GSTD significantly attenuated this effect in a dose-dependent
manner and promoted cell survival. Moreover, the analysis also
confirmed that GSTD per se improved cell survival, without
any external stimulation, in a dose-dependent manner.
GSTD alleviated DEX-induced MOMP loss in
MC3T3-E1 cells
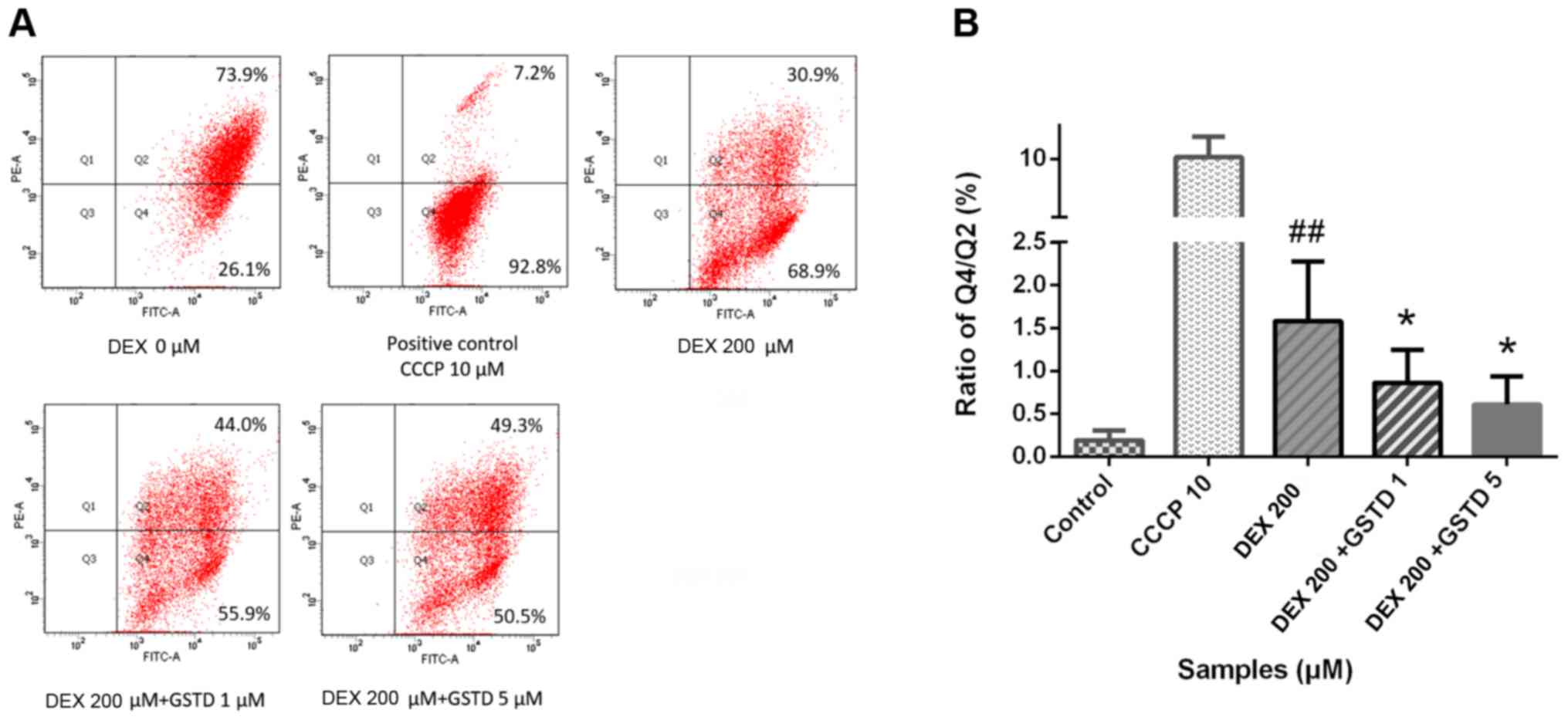
JC-1 is a type of double-fluorescent dye, which has
a good permeability through cellular and mitochondrial membranes.
Once mitochondria are damaged, the mitochondrial membrane pores are
opened, resulting in the loss of MOMP. The dye fluoresces red when
it accumulates in healthy mitochondrial matrix to form J-aggregates
with high membrane potential, whereas it fluoresces green as its
monomeric form in affected mitochondria with diminished membrane
potential. A decrease in the red/green fluorescence ratio manifests
mitochondrial depolarization. CCCP is an inhibitor of the
mitochondrial electron transport chain used as positive control,
which may exert a toxic effect by decreasing MOMP. As shown in
Fig. 4, GSTD increased MOMP
against DEX treatment in a dose-dependent manner.
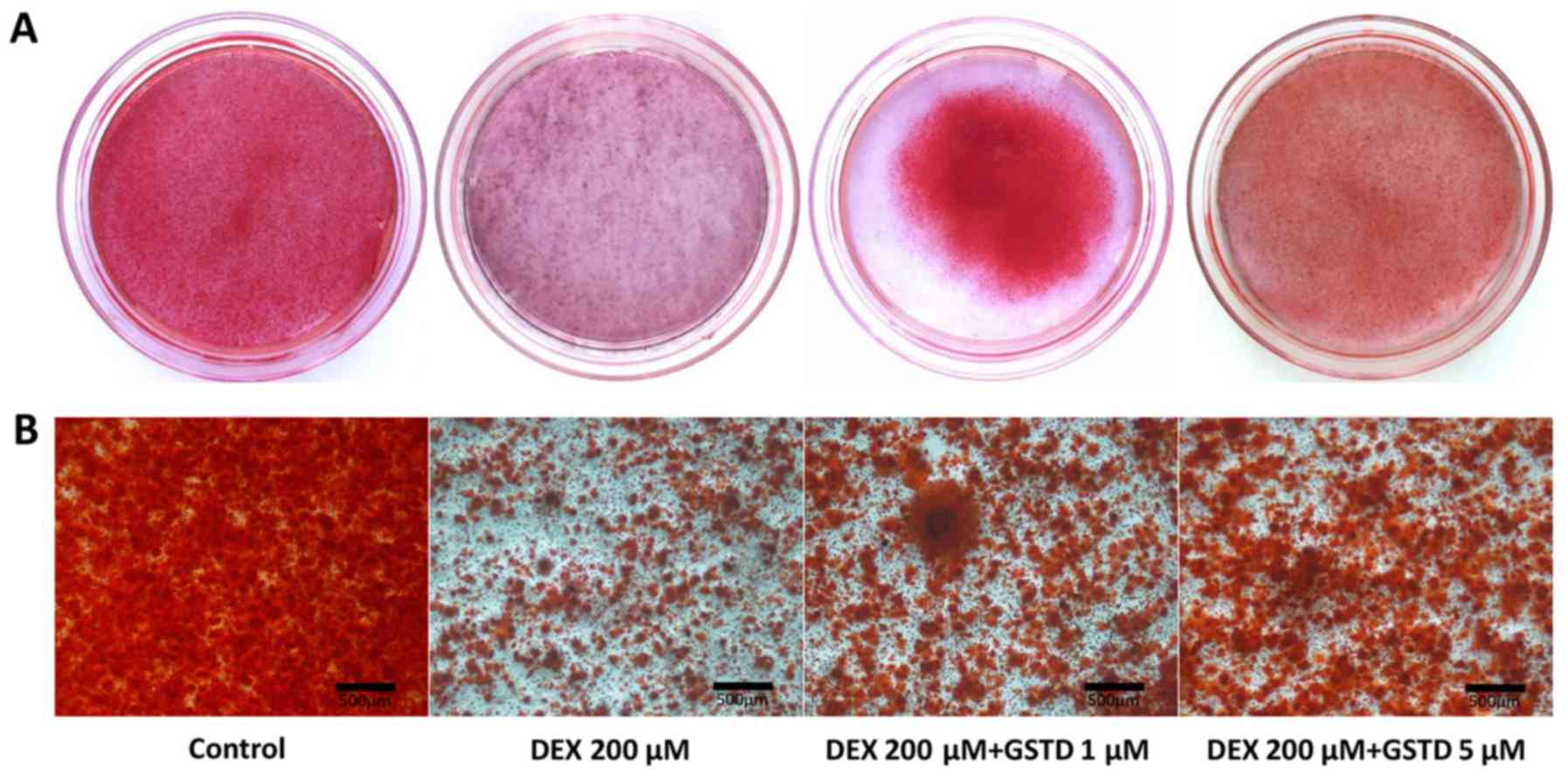
GSTD modulates DEX-induced matrix
mineralization
Osteoblast osteogenic differentiation during the
process of maturation is accompanied by mineralized bone nodule
formation. Alizarin Red staining dye conjugates with mineralized
extracellular matrix to form calcified deposits in the cells. The
effect of GSTD on mineralization in cells was determined by
Alizarin Red staining and microscopic examination. Results reported
by Huang et al indicated that a reduction in reactive oxygen
species by GSTD in human bone marrow mesenchymal stem cells
(hBMMSCs) exerted a good recovery effect on calcium mineralization,
suggesting that GSTD attenuated H2O2-induced
hBMMSC dysfunction, demonstrating its protective effects on
osteoblastogenic differentiation and calcium deposition (6). In the present study, the addition of
DEX led to inhibition of calcified deposit formation, while GSTD
significantly enhanced bone nodule formation and preserved
calcification of MC3T3-E1 cells against high-dose DEX in a
dose-dependent manner compared with that in the control group
(Fig. 5). In conclusion, GSTD
restored the mineralization ability of cells exposed to DEX
treatment.
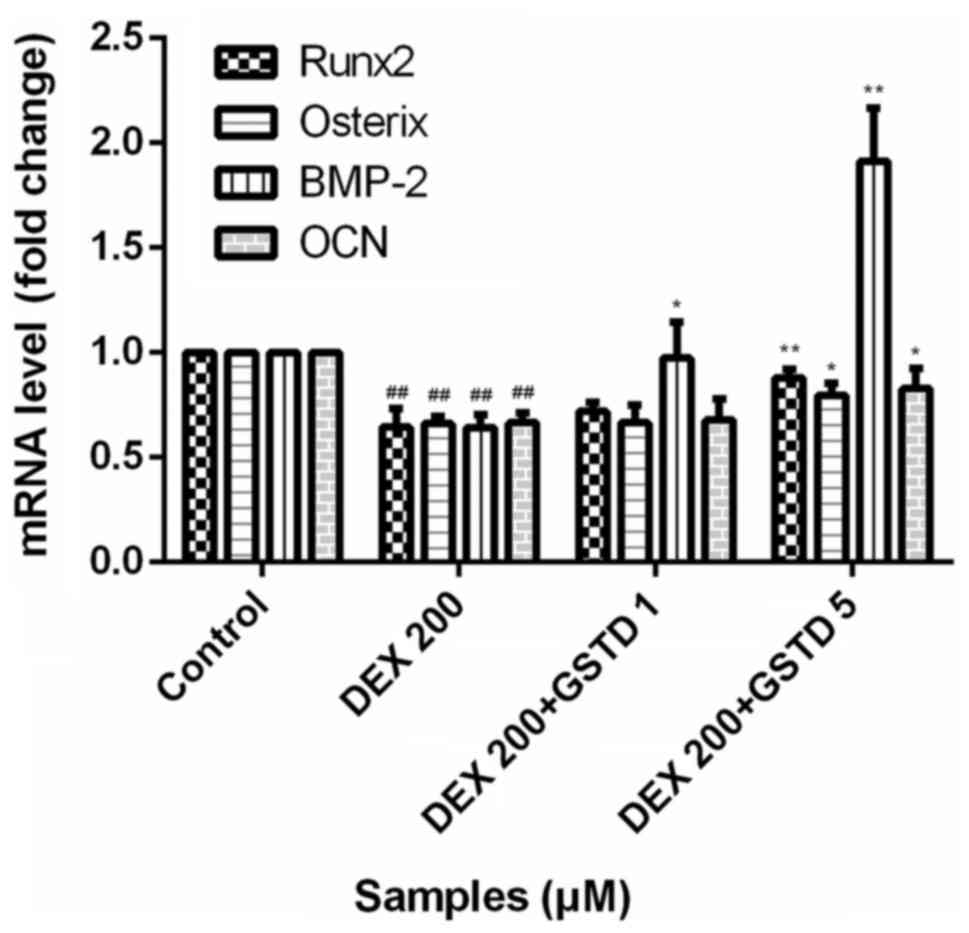
GSTD increases the expression of
osteogenic gene markers in DEX-treated MC3T3-E1 cells
GCs at physiological levels are essential for the
normal development of a wide range of tissues. DEX at physiological
(low) concentrations is indispensable for osteoblastogenic
differentiation and plays a pivotal role during maturity. To
determine the effects of GSTD on the expression of osteogenic gene
markers, the cells were pretreated with various concentrations of
GSTD for 2 h followed by adding DEX at 200 µM and incubating
under osteogenic differentiation conditions for the next 48 h.
Osteogenic differentiation was assessed by using qPCR and measuring
the mRNA expression levels of bone morphogenic protein-2 (BMP-2),
Runx2, osterix and OCN. Based on qPCR, cells treated with 200
µM DEX exhibited decreased mRNA expression levels of BMP-2,
Runx2, OSX and OCN compared with the control, whereas these
decreased expression levels were reversed in cells pretreated with
GSTD, particularly demonstrated as changes in the expression of
BMP-2 (Fig. 6), a cytokine most
widely used to confer osteoinductivity, which accounts for
osteogenic effect of osteoblasts to a great extent.
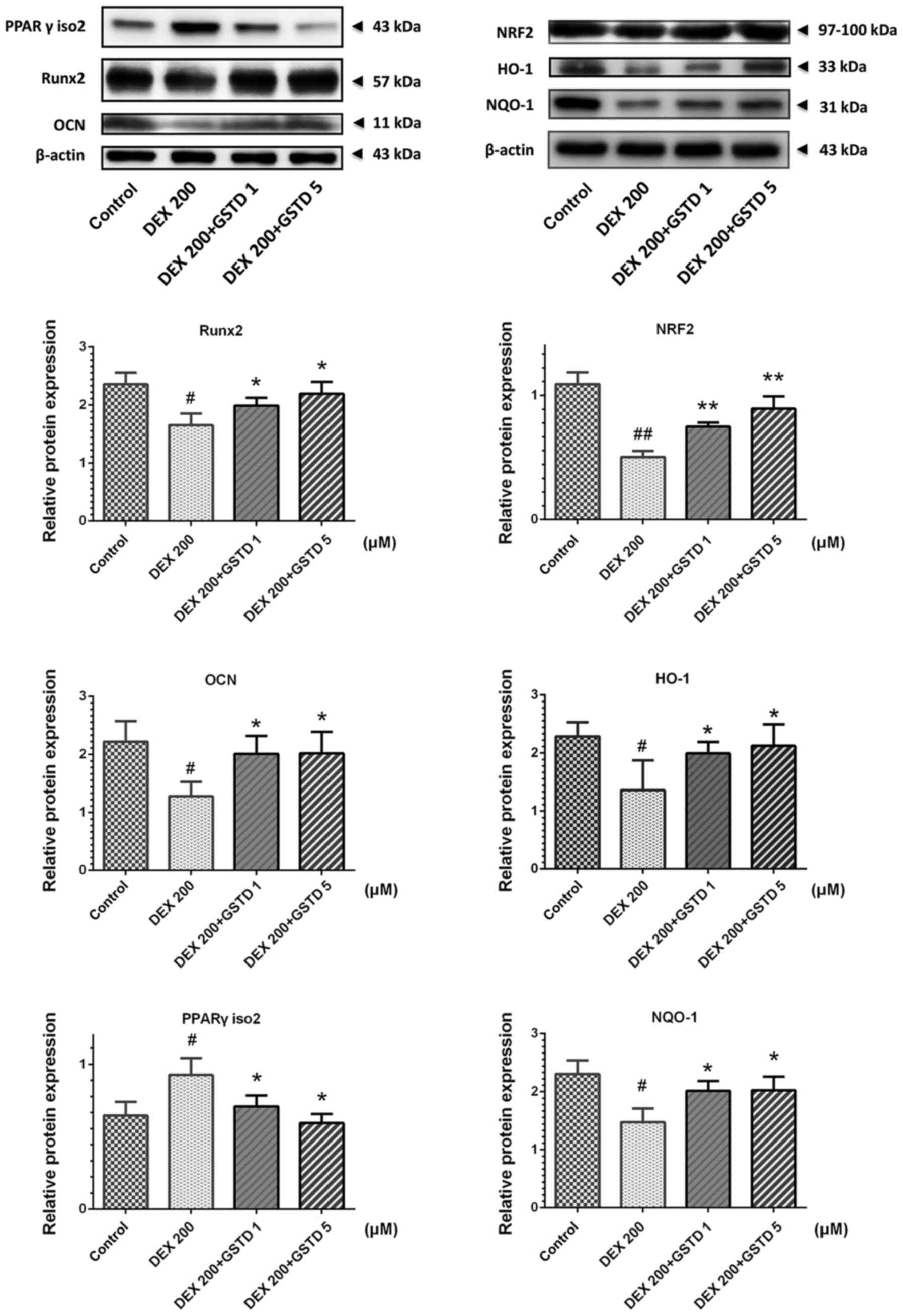
GSTD increases osteogenic transcription
Runx2 and decreases adipogenic factor PPARγ and apoptosis-related
caspase-3 protein expression levels via NRF2 pathway
activation
To further investigate the mechanisms by which GSTD
stimulates osteoblast differentiation under DEX treatment, western
blotting was performed to examine the DEX- and GSTD-induced changes
in Runx2, OCN, PPARγ isoform 2 and NRF2 pathway and its downstream
effector protein NQO-1 and HO-1 expression (Fig. 7). Runx2, OCN, NQO-1 and HO-1
expression levels were markedly increased in MC3T3-E1 cells treated
with different concentrations of GSTD (1 and 5 µM) compared
with the cells treated with DEX alone, particularly in the 5
µM group. On the contrary, PPARγ isoform 2 was
downregulated, as was caspase-3. These findings indicate that GSTD
promoted osteogenic rather than adipogenic differentiation and
blocked apoptosis via activating NRF2 and its downstream effectors’
HO-1 and NQO-1 protein expression.
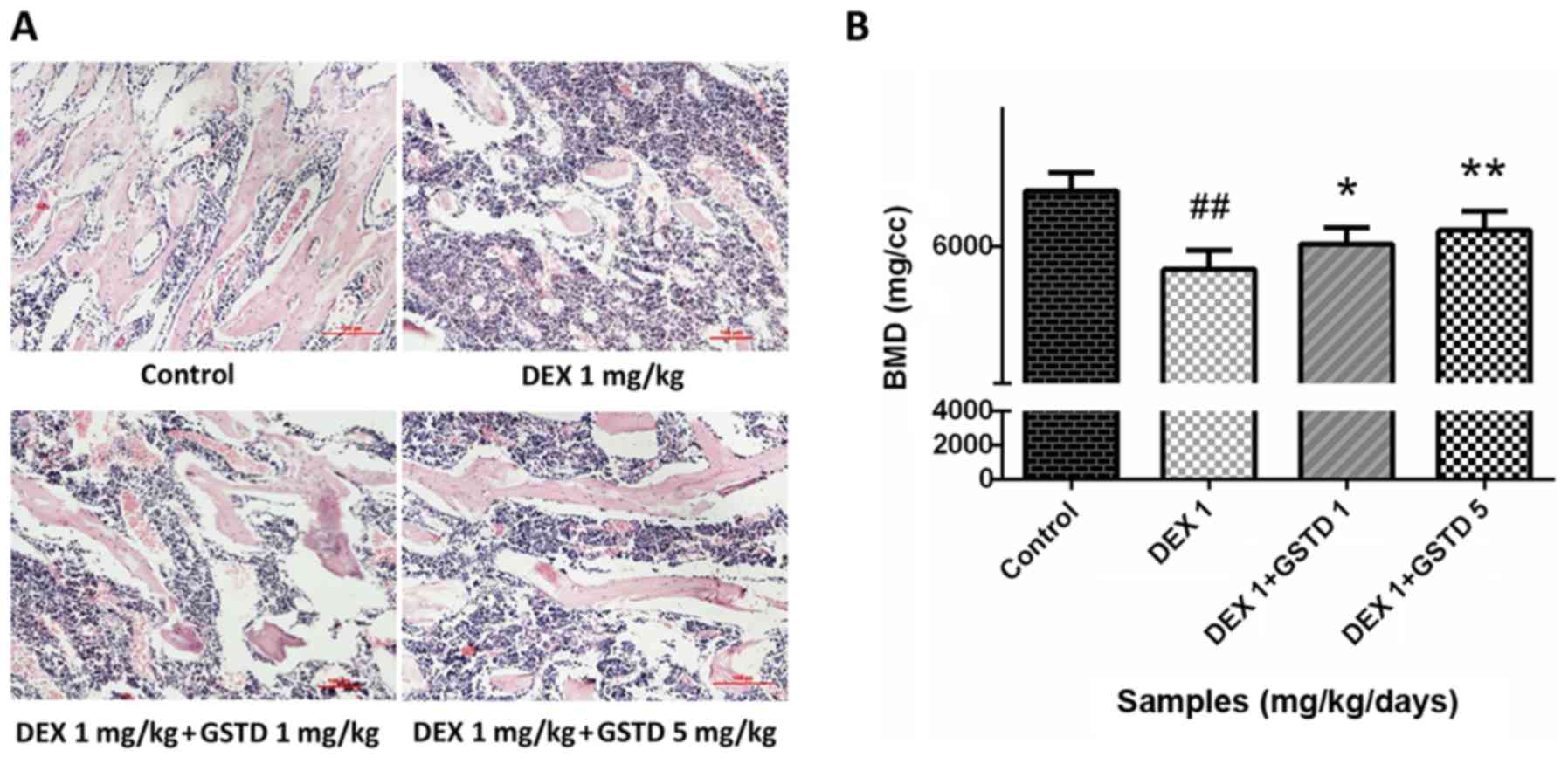
Histological assessment of rat bone
sections with H&E staining and BMD measurement indicate that
GSTD alleviates bone loss induced by DEX
DEX at a high dose reduced bone generation and
destroyed bone trabeculae in rats, leading to osteoporosis.
Microanatomical changes to the bone microarchitecture were observed
with H&E staining (magnification, ×100). The decreased density
and number of trabeculae stained by eosin in the DEX group
demonstrated severe bone loss compared with the control; however,
GSTD alleviated this effect and restored bone mass (Fig. 8A), which was consistent with the
results of the BMD measurement (Fig.
8B).
Discussion
Osteoporosis is a systemic skeletal disease
characterized by decreased bone mass caused by a number of reasons,
leading to weak and brittle bones (22). The clinical administration of GCs
may result in iatrogenic osteoporosis, which disturbs osteoblast
function, even induce apoptosis. GCs at physiological
concentrations are vital for the natural development of bone, while
high-dose administration leads to osteoporosis instead (23). Elucidation of the complex
interplay between GCs and altered bone homeostasis is a subject of
intense clinical interest. As DEX-induced bone loss is primarily
mediated by osteoblast dysfunction, agents that act directly by
either increasing osteoblast proliferation or inducing osteoblast
differentiation are required to enhance bone formation. Several
diverse natural pharmaceuticals exhibiting therapeutic efficacy
through enhancement of bone formation, osteogenic differentiation
and skeletal reconstruction, have been investigated (23,24). The traditional Chinese herb
Gastrodia elata contains an abundance of bioactive compounds
of high pharmaceutical value and biomedical potential, among which,
GSTD, a natural phenol extracted from Gastrodia elata, was
recently demonstrated to exert potential protective effects against
H2O2-induced injury in an oxidative hBMMSC
model, and is considered as a potential candidate for promoting
bone formation (8). However, the
effects of GSTD on the proliferation and differentiation of
osteoblastic cells under GC stimulation have yet to be elucidated.
Therefore, in the present study, MC3T3-E1 cells were selected in
order to investigate whether GSTD could antagonize DEX by
stimulating cell proliferation and osteogenic differentiation, as
well as preserving normal osteoblast function.
As previously stated, a decline in osteoblastic
proliferation is a significant contributor to the pathogenesis of
GIO (25), GCs inhibit osteoblast
viability and proliferation, resulting in failure of osteogenesis.
The CCK-8 assay was used to evaluate the effect of DEX and GSTD on
cell viability and proliferation. A concentration of 200 µM
DEX was selected to construct the low-viability model using
MC3T3-E1 cells. As shown in Fig.
2C, GSTD treatment at concentrations of 1 and 5 µM
restored cell viability in a dose-dependent manner. Therefore,
these results indicated that low-dose GSTD, was not cytotoxic and
promoted cell proliferation, reversing the effect of DEX.
Bone ALP is one of the phenotypic markers of
osteoblasts, and plays a key role in osteogenesis. Its activity may
directly reflect osteoblast activity and/or function. As previously
mentioned, DEX inhibits initial osteoblast proliferation followed
by decreasing ALP activity; thus, ALP activity was measured in
DEX-treated osteoblasts following pretreatment with GSTD at
different concentrations of 1 and 5 µM. The results
indicated that 200 µM DEX decreased ALP activity by ~30%
compared with the control (Fig.
2D), while addition of GSTD restored ALP activity in a
dose-dependent manner.
In addition, the changes in the expression levels of
several key osteogenic genes, such as Runx2, BMP-2, OSX and OCN,
were also investigated. The mRNA expression levels were assessed by
qPCR. GSTD-incubated MC3T3-E1 cells exhibited higher mRNA
expression of osteogenic transcription factors compared with cells
treated with DEX alone, while, as a key signaling component of the
TGF-β superfamily, BMP-2 is indispensable for osteoblast
proliferation and osteogenic differentiation, and may be the main
contributor to this effect (26,27); however, its mechanism of action
requires further elucidation. Therefore, GSTD has the ability to
promote osteogenesis against DEX treatment.
Runx2, also referred to as the core-binding factor
α1 (Cbfα1), is the most specific marker gene in the earliest stage
of bone formation, and the expression of Runx2 is a sign of
osteogenic differentiation, osteogenesis and bone development. OCN
is synthesized and secreted by osteoblasts, and it is more stable
rather than easily affected by bone resorption factors, more
accurately reflecting osteoblast activity. PPARγ is an adipogenic
transcription factor regulating adipogenesis and is found as two
isoforms, namely isoforms 1 and 2 (28), of which PPARγ isoform 2 directly
regulates cell adipogenic differentiation when highly expressed. In
the present study, treatment with DEX alone decreased Runx2
expression and increased the level of PPARγ isoform 2 in MC3T3-E1
cells. As previously reported, GSTD is considered to effectively
intervene with the effects of PPARγ in hypertensive rats (9). After pretreatment with GSTD, the
cells were confirmed to exhibit reduced PPARγ expression and
upregulated Runx2 and OCN expression, reflecting a reversal of the
DEX effects, demonstrating that GSTD exerts a targeted effect
against DEX-induced osteogenic differentiation dysfunction of
osteoblasts. It has been reported that downregulating elevated
reactive oxygen species levels in human mesenchymal stem cells is
crucial for their proper differentiation to osteoblasts and
osteogenic differentiation (29).
NRF2 is an essential transcription factor that exerts
cytoprotective effects through restoring the intracellular redox
homeostasis, including downstream transcription effectors NQO-1and
HO-1, by binding to ARE (30).
DEX decreases NRF2 pathway protein expression possibly by inducing
oxygen free radical generation; our study demonstrated that
preincubation with GSTD effectively increased NRF2 and its
downstream effectors’ NQO-1 and HO-1 expression (Fig. 7A). This result indicates that
oxidative stress accounts for the changes in the direction of
MC3T3-E1 cell differentiation. In addition, GSTD promotes
osteogenesis and maintains the balance of osteogenesis and
adipogenesis by regulating NRF2 signaling pathway to a certain
extent.
Apart from alterations in the differentiation
characteristics at the molecular level, the process of bone
formation also includes development and maturation of the
extracellular matrix to mineralized deposits. Therefore,
mineralization of extracellular matrix as Ca2+ deposits
for mineralized nodule formation was assessed by Alizarin Red
staining, which combines with Ca2+ ions. GSTD at the
determined concentrations increased the formation of calcium
nodules. These findings demonstrated that GSTD enhances the
maturation process, upregulating osteoblast osteogenic
differentiation in MC3T3-E1 cells. These results are in line with
the findings at the molecular level mentioned above.
Mitochondria are the principal organelles considered
to play a crucial role in osteoblasts, as they are involved in
energy metabolism and calcium metabolism homeostasis, regulating
cell survival and death, reflecting cellular function (31). Imbalance of mitochondrial dynamics
may lead to cellular dysfunction and are the main cause of
GC-induced bone metabolic disorders (32). It has been reported that GSTD may
induce improvement in cellular mitochondrial function in several
types of cells (8). We
hypothesized that GSTD participates in repairing mitochondrial
function, which is involved, at least in part, in the changes in
cell function underlying GIO. Loss of MOMP is an early sign of cell
apoptosis, representing mitochondrial dysfunction. Fluorescence
shifting from red to green by JC-1 can be easily detected upon
reduction of the membrane potential, and is also considered an
indicator of DEX-induced early apoptosis. As shown in Fig. 4, 200 µM DEX induced a
~3-folddecrease in MOMP compared with the control, while GSTD
preincubation for 1 h increased the red/green fluorescence ratio,
indicating that GSTD mitigated the damage in mitochondrial
function. In our study, the MOMP was measured using the JC-1 dye,
and the findings suggested that GSTD attenuated the mitochondrial
membrane injury in osteoblasts induced by DEX in vitro,
indicating that GSTD improves DEX-induced dysfunction of M3T3-E1
cells by stabilizing MOMP.
It is universally acknowledged that DEX-induced
osteoblast apoptosis significantly contributes to the development
of GIO (33). Annexin V-FITC/PI
staining-based flow cytometry analysis was applied to gain further
insight into the effect of GSTD on DEX-induced osteoblast
apoptosis. The results revealed that a large proportion of
osteoblasts underwent apoptosis following exposure to 200 µM
DEX for 48 h (Fig. 3A). However,
pretreatment with GSTD concentration-dependently attenuated
DEX-induced apoptosis of osteoblasts. In particular, GSTD at a
concentration of 5 µM exerted a more potent effect on the
decrease in apoptotic rate. These findings demonstrated that GSTD
protects osteoblasts from DEX-induced apoptosis. Caspase-3 plays a
key role in cell apoptosis, and its expression reflects apoptotic
events, which is in line with the results of Annexin V-FITC/PI flow
cytometry analysis. GSTD was able to reverse DEX-induced increase
of caspase-3 expression (Fig. 3C and
D) and protected osteoblasts from apoptosis.
In order to fully confirm that GSTD alleviates GIO,
in the present study, rats were selected as the model of GIO, and
the results demonstrated that GSTD obviously improved DEX-induced
osteoporosis in a rat model of GIO in vivo. H&E staining
and BMD measurement were applied. The femur is the most vulnerable
body region in terms of mechanical strength, and ~40% of European
women with atypical femoral fractures have received treatment with
GCs at some point (34,35). Hence, the femur was selected to
evaluate the effects of GSTD on GIO. BMD, which is a useful index
reflecting the status of bone metabolism, is broadly used to assess
the changes in bone mass and predict the stiffness of the femur.
GSTD relieved the erosive effects induced by DEX and increased the
BMD, improving bone quality and strength.
GSTD was previously reported to protect
ovariectomized rats against osteoporosis. However, the effect of
GSTD on GIO remains unclear. It has been reported that GSTD may
promote the activation of the NRF2 signaling pathway in other
cells, playing a role in antioxidation and anti-inflammation.
Furthermore, GSTD may also be used in hBMMSCs and promotes
osteogenesis by reducing osteoclast differentiation. GSTD inhibits
the key adipogenic differentiation factor PPARγ, reducing the fat
content in hypertensive model rats (37). In addition, GSTD was also found to
improve mitochondrial function (16,38). These interactions are closely
associated with the mechanism of GIO. The present study mainly
investigated the effects of GSTD on DEX-induced osteoblast
dysfunction, and the findings were also validated in an animal
osteoporotic model. Furthermore, GSTD targetedly modulated some
essential factors in osteoblasts and attenuated the adverse effects
of supra-physiological concentrations of DEX on osteoblasts.
As regards osteoclasts, experimental studies have
established that GCs directly extend the lifespan of mature
osteoclasts by delaying apoptotic signaling. Furthermore, the
effects of GSTD on osteoclasts were well-documented in the study of
Huang et al, who reported that GSTD suppressed osteoclastic
differentiation in hBMMSCs and RAW264.7 cells, a type of
preosteoclast cell (6). This
study will hopefully provide new experimental data in the study of
osteoclasts and help elucidate the mechanism of action of GSTD in
DEX-induced osteoclasts in future studies, which may provide a new
aproach to the development of new drugs for GIO treatment. Further
research is required to provide more insight into DEX-induced
osteoporosis and support the wider application of GSTD in clinical
practice.
In conclusion, the present study demonstrated that
GSTD enhanced osteogenic differentiation and bone formation under
conditions of GC-induced dysfunction of MC3T3-E1 cells in
vitro, through maintaining cell viability, stabilizing
mitochondrial function, promoting osteogenic differentiation and
decreasing cell apoptosis. However, further studies are required to
fully elucidate the mechanism underlying GC-induced apoptosis in
primary osteoblasts and the efficacy of GSTD in vivo for the
treatment of GIO.
Acknowledgments
This study was sponsored by grants from the National
Natural Science Foundation of China (no. 81370981) and the
Outstanding Scientific Fund of Shengjing Hospital (MD31).
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Drake MT, Clarke BL and Lewiecki EM: The
pathophysiology and treatment of osteoporosis. Clin Ther.
37:1837–1850. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Seibel MJ, Cooper MS and Zhou H:
Glucocorticoid-induced osteoporosis: Mechanisms, management, and
future perspectives. Lancet Diabetes Endocrinol. 1:59–70. 2013.
View Article : Google Scholar
|
|
3
|
Frenkel B, White W and Tuckermann J:
Glucocorticoid-induced osteoporosis. Adv Exp Med Biol. 872:179–215.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Henneicke H, Gasparini SJ,
Brennan-Speranza TC, Zhou H and Seibel MJ: Glucocorticoids and
bone: Local effects and systemic implications. Trends Endocrinol
Metab. 25:197–211. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Buehring B, Viswanathan R, Binkley N and
Busse W: Glucocorticoid-induced osteoporosis: An update on effects
and management. J Allergy Clin Immunol. 132:1019–1030. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang Q, Shi J, Gao B, Zhang HY, Fan J, Li
XJ, Fan JZ, Han YH, Zhang JK, Yang L, et al: Gastrodin: An ancient
Chinese herbal medicine as a source for anti-osteoporosis agents
via reducing reactive oxygen species. Bone. 73:132–144. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zheng H, Yang E, Peng H, Li J, Chen S,
Zhou J, Fang H, Qiu B and Wang Z: Gastrodin prevents
steroid-induced osteonecrosis of the femoral head in rats by
anti-apoptosis. Chin Med J (Engl). 127:3926–3931. 2014.
|
|
8
|
Qu LL, Yu B, Li Z, Jiang WX, Jiang JD and
Kong WJ: Gastrodin ameliorates oxidative stress and proinflammatory
response in nonalcoholic fatty liver disease through the AMPK/Nrf2
pathway. Phytother Res. 30:402–411. 2016. View Article : Google Scholar
|
|
9
|
Peng Z, Wang S, Chen G, Cai M, Liu R, Deng
J, Liu J, Zhang T, Tan Q and Hai C: Gastrodin alleviates cerebral
ischemic damage in mice by improving anti-oxidant and
anti-inflammation activities and inhibiting apoptosis pathway.
Neurochem Res. 40:661–673. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun YX, Xu AH, Yang Y and Li J: Role of
Nrf2 in bone metabolism. J Biomed Sci. 22:1012015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Loboda A, Damulewicz M, Pyza E, Jozkowicz
A and Dulak J: Role of Nrf2/HO-1 system in development, oxidative
stress response and diseases: An evolutionarily conserved
mechanism. Cell Mol Life Sci. 73:3221–3247. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun YX, Li L, Corry KA, Zhang P, Yang Y,
Himes E, Mihuti CL, Nelson C, Dai G and Li J: Deletion of Nrf2
reduces skeletal mechanical properties and decreases load-driven
bone formation. Bone. 74:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin H, Wei B, Li G, Zheng J, Sun J, Chu J,
Zeng R and Niu Y: Sulforaphane reverses glucocorticoid-induced
apoptosis in osteoblastic cells through regulation of the Nrf2
pathway. Drug Des Devel Ther. 8:973–982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pellegrini GG, Morales CC, Wallace TC,
Plotkin LI and Bellido T: Avenanthramides prevent osteoblast and
osteocyte apoptosis and induce osteoclast apoptosis in vitro in an
Nrf2-independent manner. Nutrients. 8:4232016. View Article : Google Scholar :
|
|
15
|
Choi EM: Magnolol protects osteoblastic
MC3T3-E1 cells against antimycin A-induced cytotoxicity through
activation of mitochondrial function. Inflammation. 35:1204–1212.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan Z, Li Q, Luo S, Liu Z, Luo D, Zhang
B, Zhang D, Rao P and Xiao J: PPARγ and Wnt signaling in adipogenic
and osteogenic differentiation of mesenchymal stem cells. Curr Stem
Cell Res Ther. 11:216–225. 2016. View Article : Google Scholar
|
|
17
|
Rauch A, Seitz S, Baschant U, Schilling
AF, Illing A, Stride B, Kirilov M, Mandic V, Takacz A,
Schmidt-Ullrich R, et al: Glucocorticoids suppress bone formation
by attenuating osteoblast differentiation via the monomeric
glucocorticoid receptor. Cell Metab. 11:517–531. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hartmann K, Koenen M, Schauer S,
Wittig-Blaich S, Ahmad M, Baschant U and Tuckermann JP: Molecular
actions of glucocor-ticoids in cartilage and bone during health,
disease, and steroid therapy. Physiol Rev. 96:409–447. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu S, Yerges-Armstrong LM, Chu Y, Zmuda JM
and Zhang Y: E2F1 effects on osteoblast differentiation and
mineralization are mediated through upregulation of frizzled-1.
Bone. 56:234–241. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen Z, Xue J, Shen T, Ba G, Yu D and Fu
Q: Curcumin alleviates glucocorticoid-induced osteoporosis by
protecting osteoblasts from apoptosis in vivo and in vitro. Clin
Exp Pharmacol Physiol. 43:268–276. 2016. View Article : Google Scholar
|
|
21
|
Fang J, Yamaza H, Uchiumi T, Hoshino Y,
Masuda K, Hirofuji Y, Wagener FA, Kang D and Nonaka K:
Dihydroorotate dehydrogenase depletion hampers mitochondrial
function and osteogenic differentiation in osteoblasts. Eur J Oral
Sci. 124:241–245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chan CK, Mason A, Cooper C and Dennison E:
Novel advances in the treatment of osteoporosis. Br Med Bull.
119:129–142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kalak R, Zhou H, Street J, Day RE,
Modzelewski JR, Spies CM, Liu PY, Li G, Dunstan CR and Seibel MJ:
Endogenous glucocorticoid signalling in osteoblasts is necessary to
maintain normal bone structure in mice. Bone. 45:61–67. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Z, Xue J, Shen T, Mu S and Fu Q:
Curcumin alleviates glucocorticoid-induced osteoporosis through the
regulation of the Wnt signaling pathway. Int J Mol Med. 37:329–338.
2016. View Article : Google Scholar :
|
|
25
|
Kim J, Lee H, Kang KS, Chun KH and Hwang
GS: Protective effect of Korean Red Ginseng against
glucocorticoid-induced osteoporosis in vitro and in vivo. J Ginseng
Res. 39:46–53. 2015. View Article : Google Scholar
|
|
26
|
Liang W, Lin M, Li X, Li C, Gao B, Gan H,
Yang Z, Lin X, Liao L and Yang M: Icariin promotes bone formation
via the BMP-2/Smad4 signal transduction pathway in the hFOB 1.19
human osteoblastic cell line. Int J Mol Med. 30:889–895. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao H, Ke Y, Zhang Y, Zhang CJ, Qian W and
Zhang GL: Icariin stimulates MC3T3-E1 cell proliferation and
differentiation through upregulation of bone morphogenetic
protein-2. Int J Mol Med. 29:435–439. 2012.
|
|
28
|
Zhuang H, Zhang X, Zhu C, Tang X, Yu F,
Shang GW and Cai X: Molecular mechanisms of PPAR-γ governing MSC
osteogenic and adipogenic differentiation. Curr Stem Cell Res Ther.
11:255–264. 2016. View Article : Google Scholar
|
|
29
|
Gómez-Puerto MC, Verhagen LP, Braat AK,
Lam EW, Coffer PJ and Lorenowicz MJ: Activation of autophagy by
FOXO3 regulates redox homeostasis during osteogenic
differentiation. Autophagy. 12:1804–1816. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Choi EM, Suh KS, Kim YJ, Hong SM, Park SY
and Chon S: Glabridin alleviates the toxic effects of methylglyoxal
on osteoblastic MC3T3-E1 cells by increasing expression of the
glyoxalase system and Nrf2/HO-1 signaling and protecting
mitochondrial function. J Agric Food Chem. 64:226–235. 2016.
View Article : Google Scholar
|
|
31
|
Angelova PR and Abramov AY: Functional
role of mitochondrial reactive oxygen species in physiology. Free
Radic Biol Med. 100:81–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhen YF, Wang GD, Zhu LQ, Tan SP, Zhang
FY, Zhou XZ and Wang XD: P53 dependent mitochondrial permeability
transition pore opening is required for dexamethasone-induced death
of osteoblasts. J Cell Physiol. 229:1475–1483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
O’Brien CA, Jia D, Plotkin LI, Bellido T,
Powers CC, Stewart SA, Manolagas SC and Weinstein RS:
Glucocorticoids act directly on osteoblasts and osteocytes to
induce their apoptosis and reduce bone formation and strength.
Endocrinology. 145:1835–1841. 2004. View Article : Google Scholar
|
|
34
|
de Vries F, Pouwels S, Lammers JW,
Leufkens HG, Bracke M, Cooper C and van Staa TP: Use of inhaled and
oral glucocorticoids, severity of inflammatory disease and risk of
hip/femur fracture: A population-based case-control study. J Intern
Med. 261:170–177. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Giusti A, Hamdy NA and Papapoulos SE:
Atypical fractures of the femur and bisphosphonate therapy: A
systematic review of case/case series studies. Bone. 47:169–180.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang ZC, Su G, Li J, Wu H and Xie XD: Two
new neuroprotective phenolic compounds from Gastrodia elata. J
Asian Nat Prod Res. 15:619–623. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu W, Wang L, Yu J, Asare PF and Zhao YQ:
GSTD reduces blood pressure by intervening with RAAS and PPARγ in
SHRs. Evid Based Complement Alternat Med. 2015:8284272015.
View Article : Google Scholar
|
|
38
|
Wang XL, Xing GH, Hong B, Li XM, Zou Y,
Zhang XJ and Dong MX: Gastrodin prevents motor deficits and
oxidative stress in the MPTP mouse model of Parkinson’s disease:
Involvement of ERK1/2-Nrf2 signaling pathway. Life Sci. 114:77–85.
2014. View Article : Google Scholar : PubMed/NCBI
|