Introduction
Atherosclerosis (AS) is as a chronic inflammatory
disease that exhibits severe complications, including myocardial
infarction and ischemic stroke, which are considered two of the
main causes of mortality worldwide (1). Macrophages (MPs) are the most
abundant inflammatory cell to invade atherosclerotic lesions, and
are essential during the atherogenic process (1,2).
During initiation, inflammatory cytokines produced by MPs stimulate
the generation of endothelial adhesion molecules, proteases and
other mediators that enter the systemic circulation in soluble
forms (3). Furthermore, MPs
internalize modified lipoproteins in order to produce foam cells,
which are considered a hallmark event in the formation of early
atherosclerotic lesions (4).
During the advanced stage of the disease, MPs contribute to the
plaque morphology, affecting the fibrous cap and necrotic core,
which increase the pro-inflammatory responses and the apoptotic
signals (5). Therefore,
strategies focusing on the inhibition of MP activation may be
beneficial for the prevention and treatment of AS.
AS is a multiphase process characterized by
activation of the endothelium with the secretion of monocytic
maturation factors (6). AS is
initiated by endothelial injury due to oxidative stress associated
with several cardiovascular risk factors (7). Endothelial cell dysfunction (ECD)
manifested in lesion-prone areas of arterial vasculature recruits
circulating monocytes from the blood into the tunica intima, which
eventually differentiate into MPs (8-10).
In turn, monocyte-derived MPs trigger an inflammatory cascade,
including the production of pro-inflammatory cytokines, interleukin
(IL)-1β, IL-12, tumor necrosis factor-α (TNF-α), and chemokines,
including monocyte chemoattractant protein-1 (MCP-1). These
mediators attract additional monocytes, which adhere to the
endothelial cells (ECs) (1,11).
In previous years, the interaction between MPs and ECs has been
considered a significant feature of a majority of
pathophysiological conditions (12). However, the precise signaling
pathways between them remain to be fully elucidated.
Toll-like receptor 4 (TLR4) is expressed in MPs, ECs
and smooth muscle cells. The receptor is mediated by myeloid
differentiation factor 88 (MyD88)-dependent and -independent
pathways (13,14). Nuclear factor-κB (NF-κB) is a
transcription factor, which localizes to the cytoplasm and mainly
exists as a heterodimer consisting of the p50 and p65 subunits
(15). NF-κB signaling
upregulates the expression levels of lectin-like oxidized
low-density lipoprotein receptor 1, vascular adhesion molecule 1
and MCP-1 in ECs induced by oxidized low-density lipoprotein
(ox-LDL) in vitro (16).
The TLR4/MyD88/NF-κB signaling pathway is the main signaling
pathway to be activated during TLR4-mediated inflammation (17). Activated TLR4 signaling
upregulates the protein expression levels of TLR4
pathway-associated mediators, including upstream (TLR4, MyD88 and
NF-κB) and downstream (IL-1β, IL-6 and TNF-α) factors (18). IL-1β belongs to the IL-1 cytokine
family together with IL-1α, IL-1Ra, IL-18 and IL-33 (19,20). The generation of mature IL-1β is
tightly controlled by a diverse class of cytosolic protein
complexes, inflammasome components, and the K+ efflux
(19,21,22). However, the exact mechanism by
which the activation of TLR4 affects the interaction between ECs
and MPs, during the oxidative damage of ECs remains to be
elucidated.
Currently, the management of AS aims to reduce
inflammation and adjust dyslipidemia. Therefore, research efforts
have focused on the development of several natural and synthetic
agents in order to attenuate inflammation at the molecular level.
Resveratrol inhibits the IL-1β-induced expression of matrix
metalloproteinase-13, IL-6 and TNF-α via the TLR4/MyD88-dependent
signaling cascades (23,24), and it further attenuates neuronal
autophagy and inflammatory injury in experimental traumatic brain
injury (25). Diallyl trisulfide
exerts anti-inflammatory effects via suppressing the TLR4/NF-κB
signaling pathway in lipopolysaccharide (LPS)-stimulated Raw 264.7
macrophages and isch-emic stroke-induced inflammation (26,27). In addition,
(S,R)-3-phenyl-4,5-dihydro-5-isoxasole acetic acid inhibited
LPS-induced NF-κB and p38 mitogen-activated protein kinase
signaling pathways, and reduced the secretion of IL-1β, TNF-α and
IL-10 from peritoneal cells in vitro and ex vivo
(28,29). However, there are several
disadvantages limiting the clinical application of these agents,
including side effects and poor oral bioavailability (30).
7-difluoromethoxy-5,4′-dimethoxy-genistein (DFMG) is a novel active
chemical entity, which is synthesized by the precursor genistein
(GEN), and exhibits optimal fat-solubility, antioxidant and
anti-inflammatory activities (31,32). DFMG is involved in the regulation
of different inflammatory cytokines, including E-selectin,
inter-cellular adhesion molecule 1 (ICAM-1), IL-6 and TNF-α
(33). In addition, DFMG
interacts with the inhibition of different signaling pathway
activations, including mitochondrial apoptosis and TLR4 signaling
(33-35).
The activation of MPs has been shown to be the
critical step in the development of chronic inflammation.
Therefore, the mechanism involving the activation of MPs is
important for the development of preventive and therapeutic
strategies for AS. The aim of the present study was to: i) identify
the function of injured ECs with regard to the activation of MPs in
a non-contact co-culture model and ii) to investigate the effect
and molecular mechanism of DFMG treatment on the activation of MPs
induced by co-culture with injured ECs.
Materials and methods
Reagents and antibodies
Phorbol 12-myristate 13-acetate (PMA),
lysophosphatidylcholine (LPC) and oil red O powder were purchased
from Sigma-Aldrich; EMD Millipore (Billerica, MA, USA). DFMG
(purity >99%) was synthesized as previously reported (31) and dissolved in dimethyl sulfoxide
(DMSO; Ameresco, Inc., Framingham, MA, USA). This compound was
subsequently sterilized by filtration. CLI-095 was obtained from
Invitrogen; Thermo Fisher Scientific, Inc. (Waltham, MA, USA). The
IL-1 receptor antagonist (IL-1Ra) was purchased from Bioss
(Beijing, China).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was procured from Beyotime Institute of Biotechnology (Shanghai,
China). Hematoxylin staining reagents were obtained from Servicebio
Technology Co., Ltd. (Wuhan, China). Anti-TLR4 and NF-κB p65
antibodies were provided by ProteinTech Group, Inc. (Chicago, IL,
USA, cat. nos. 19811-1-AP and 10745-1-AP). Anti-MyD88 and GAPDH
antibodies were provided by Abgent, Inc. (San Diego, CA, USA cat.
nos. ABO10975 and AM1020b). Goat anti-rabbit IgG and goat
anti-mouse IgG were provided by ComWin Biotech Co., Ltd. (Beijing,
China, cat. nos. CW0156S and CW0102S).
Cell culture and treatment
Human umbilical vein ECs (HUVE-12) and human acute
monocytic leukemia cells (THP-1) were purchased from the China
Center for Type Culture Collection (Wuhan, China) and Institute of
Biochemistry and Cell Biology (Shanghai, China), respectively. The
two cell lines were incubated in a humidified incubator at 37°C
with 5% CO2 and were cultured in Roswell Park Memorial
Institute (RPMI)-1640 medium containing 10% fetal bovine serum
(FBS) and 1% penicillin/streptomycin. The THP-1 cells
(1×106 cells/ml) were seeded in 6-well plates and
activated with 200 nM of PMA for 24 h, which allowed them to
transform into MPs. All cell culture reagents were purchased from
Biological Industries (Beit Haemek, Israel).
Reactive oxygen species (ROS) assay
The HUVE-12 cells were treated at 37°C with various
concentrations of LPC (10, 20, 30, 40 and 50 µM). Following
incubation for 24 h, fresh medium with 10% FBS was used and the
cells were cultured for another 24 h. Subsequently, ROS was
determined using a ROS assay kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol.
Fluorescence intensity was detected at excitation and emission
wavelengths of 488 and 525 nm, respectively. Hematoxylin staining
was used to quantify the number of cells under a light microscope
(CX41RF, Olympus, Tokyo, Japan). The fluorescence intensity of each
cell was obtained from the ratio of total fluorescence intensity to
the total cell count.
Lactate dehydrogenase (LDH) assay
The HUVE-12 cells were incubated with various
concentrations of LPC and subsequently treated as described in the
aforementioned protocol. Subsequently, the supernatants were
collected in order to determine the concentration of LDH using the
LDH cytotoxicity assay kit (Jiancheng Bioengineering Institute,
Nanjing, China). The optical density (OD) was measured at 450 nm
with a microplate reader (Elx800; BioTek Instruments, Inc.,
Winooski, VT, USA). The concentration of LDH was calculated
according to the manufacturer's protocol.
Establishment of the co-culture
model
The HUVE-12 cells, which had been stimulated by LPC
were co-cultured with MPs in a Transwell chamber consisting of a
10-µm thick porous membrane with 0.4-µm pores in the
culture inserts (Corning Incorporated, Corning, NY, USA). This
methodology was used to construct an indirect co-culture system.
Briefly, the MPs were seeded in the wells of a 6-well
(1×105 cells/well) and/or a 24-well (2.5×104
cells/well) plate (lower layer) at 37°C prior to co-culture with
the HUVE-12 cells. The HUVE-12 cells were individually grown in the
upper layer of the Transwell under the following treatment
conditions: Pretreatment with LPC for 24 h, and/or pretreatment
with DFMG for 1 h, followed by CLI-095 and IL-1Ra for 24 h, and
subsequent treatment with LPC for another 24 h. The HUVE-12 cells
in fresh medium containing the inserts were added to the upper
chamber of the Transwell system, with the MPs located at the
bottom.
Proliferation assay
The MTT assay was used to detect the viability of
the cells. The MPs were co-cultured with the HUVE-12 cells, which
had been pretreated for 24 h in 24-well plates, and 50 µl of
MTT solution (MTT powder dissolved in PBS, 5 mg/ml) was added to
each well. The plates were incubated at 37°C for 4 h. Following
removal of the medium, 750 µl of DMSO was added to each well
for 10 min in order to fully dissolve the formazan crystals. The OD
was measured on a microplate reader (BioTek Instruments, Inc.) at
490 nm. The viability of the cells was assessed in terms of OD
values.
Migration assay
A wound healing assay was used to analyze the
migratory activity of the cells. Prior to co-culture, the MPs were
seeded in 6-well plates and scratched with a pipette tip
(200-µl), followed by rinsing with PBS and incubation in
RPMI-1640 medium in the absence of FBS. Images were captured at the
0 h time point. The pretreated HUVE-12 cells containing inserts
were added to the upper chamber of the Transwell system and
incubated at 37°C. After 24 h, an additional image (24 h) was
captured in the same position. The cell-free area was quantified
using Adobe Photoshop CS6 software (Adobe Systems, Inc., San Jose,
CA, USA).
Oil red O staining
The oil red O working solution was prepared by
diluting the stock solution (0.05 g of oil red O powder dissolved
in 10 ml of isopropanol) with distilled water (3:2), following
which the solution was filtered. The MPs were co-cultured with
HUVE-12 cells for 24 h and fixed in 4% paraformaldehyde for 15 min
in order to prepare the samples for oil red O staining. The samples
were washed with distilled water twice and were incubated with
filtered oil red O working solution at room temperature for 10 min.
Subsequently, the samples were washed twice with distilled water,
and the cells were observed for image capture under an optical
microscope (CX41RF, Olympus, Tokyo, Japan). The cells containing
oil red O-positive fat droplets were considered as foam cells.
Assessment of total cholesterol (TC)
The quantification of TC was performed according to
the protocols provided by the manufacturer (Solarbio, Beijing,
China). The MPs were co-cultured with HUVE-12 cells for 24 h, and
the cells were collected and dissolved in isopropanol (1
ml/4×104 cells), followed by ultrasound treatment for 1
min. The samples were centrifuged at 4°C at 8,000 × g for 10 min.
The OD of the supernatants was detected at 500 nm using a
spectrophotometer. The concentration of TC was calculated according
to the manufacturer's protocol.
Western blot analysis
The pretreated HUVE-12 cells, which were co-cultured
with MPs for 24 h, were digested and lysed in
radioimmunoprecipitation assay buffer containing 1% phenylmethyl
sulfonyl fluoride. The samples were centrifuged at 12,000 × g at
4°C for 10 min. Protein quantitation was performed using the BCA
protein assay kit (Solabio). A total of 25 µg of each
protein lysate was separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (Solarbio), followed by
transfer onto polyvinylidene difluoride membranes (EMD Millipore),
which were blocked with 5% non-fat milk in PBST (PBS containing
0.05% Tween-20) at room temperature for 1 h. The membranes were
incubated with antibodies against TLR4 (1:1,000), MyD88 (1:10,000),
NF-κB p65 (1:2,500) and GAPDH (1:1,000) on a shaking platform at
4°C overnight. Subsequently, the membranes were washed with PBST
and incubated with goat anti-rabbit antibody (1:10,000) and goat
anti-mouse antibody (1:10,000) for 1 h at room temperature,
followed by washing in PBST three times. Finally, the protein bands
were visualized using enhanced chemiluminescence. The results were
analyzed using densitometry with GelPro 3.2 software (Media
Cybernetics, Inc., Rockville, MD, USA).
Enzyme-linked immunosorbent assay
(ELISA)
Following co-culture for 24 h, the concentration of
IL-1β in the supernatants was measured using a human IL-1β ELISA
kit (MultiSciences Biotech Co., Ltd., Hangzhou, China) according to
the manufacturer's protocol. The standards and/or samples were
incubated in the wells at 37°C for 1.5 h. Subsequently, the wells
were washed and primary antibody was added, covered with an
adhesive strip and incubated at 37°C for 1 h. Following washing of
the unbound biotinylated antibody, streptavidin-HRP was added for
incubation at 37°C for 30 min. A cycle of five washing steps (with
PBST for 30 sec each time) was performed and substrate solution was
added to the samples. Following incubation at dark for 30 min, the
stop solution was added. The OD was detected at 450 nm on a
microplate reader (BioTek Instruments, Inc.), and the concentration
of IL-1β was calculated according to the manufacturer's
protocol.
Statistical analysis
All experiments were performed independently at
least three times, and the values are expressed as the mean ±
standard deviation. SPSS 20.0 software (IBM SPSS, Armonk, NY, USA)
and GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA)
were used for statistical analysis. Student's paired t-test was
used for the comparison of two samples, and one-way analysis of
variance was used for multiple group comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
Establishment of the injured HUVE-12
cell-MP co-culture model
Initially, HUVE-12 cells were incubated with
different concentrations (10, 20, 30, 40 and 50 µM) of LPC
for 24 h, and the production of ROS and LDH were detected. The
results indicated that LPC damaged the HUVE-12 cells in a
concentration-dependent manner. LPC significantly promoted the
generation of ROS at a concentration range of 20-50 µM
(Fig. 1A and B). In terms of LDH
activity, the effective concentration range of LPC was estimated to
be between 30 and 50 µM (Fig.
1C). Therefore, 30 µM of LPC was selected in order to
injure the HUVE-12 cells.
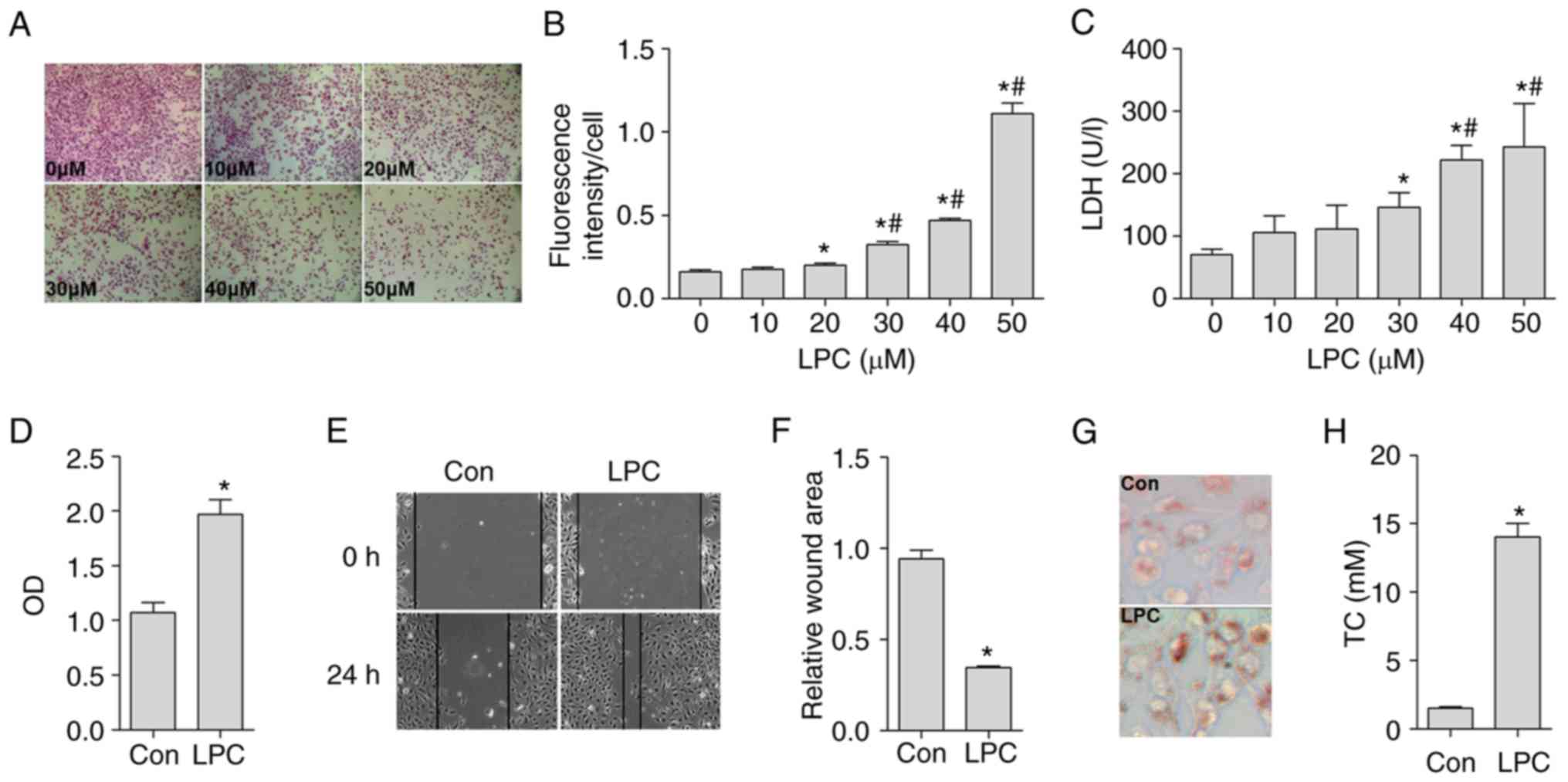 | Figure 1Effects of LPC on the generation of
ROS and LDH in HUVE-12 cells, and activation of MPs induced by
co-culture with LPC-injured HUVE-12 cells. (A) Representative
images of hematoxylin and eosin staining of LPC-injured HUVE-12
cells. The number of stained cells was counted for each group
across four areas at 200× magnification and the mean was estimated.
(B) LPC (20, 30, 40 and 50 µM) significantly enhanced the
fluorescence intensity of each cell in a concentration-dependent
manner. (C) LPC (30, 40 and 50 µM) significantly increased
the release of LDH in a concentration-dependent manner. LPC (30
µM) effectively increased the (D) cell viability, and
decreased the relative wound area of MPs co-cultured with
LPC-injured HUVE-12 cells, as shown in (E) images (magnification,
×100) and (F) graph. (G) Representative images of foam cell
formation. LPC (30 µM) effectively increased lipid
accumulation in MPs (magnification, ×200). (H) LPC (30 µM)
effectively increased the TC content. Data are presented as the
mean ± standard deviation of three separate experiments.
*P<0.05, vs. 0 µM LPC and/or Con (not treated
with LPC) group, #P<0.05, vs. 20 µM and/or 30
µM LPC group. LPC, lysophosphatidylcholine; ROS, reactive
oxygen species; LDH, lactate dehydrogenase; MPs, macrophages; Con,
control; OD, optical density. |
Subsequently, the capacity of MPs to proliferate,
migrate and develop into foam cells following co-culture with
injured HUVE-12 cells was detected. The HUVE-12 cells were
incubated with LPC (30 µM) for 24 h and co-cultured with MPs
in a Transwell system for another 24 h. A significant increase in
the proliferation and migration of MPs was observed, which was
caused by co-culture with injured HUVE-12 cells (Fig. 1D–F). In addition, co-culture with
injured HUVE-12 cells resulted in abundant cytoplasmic lipid
droplet accumulation in the MPs, as demonstrated by oil red O
staining (Fig. 1G) and TC
(Fig. 1H). Consequently, 30
µM of LPC was selected as the optimum concentration in order
to construct the LPC-injured HUVE-12 cell-MP co-culture model.
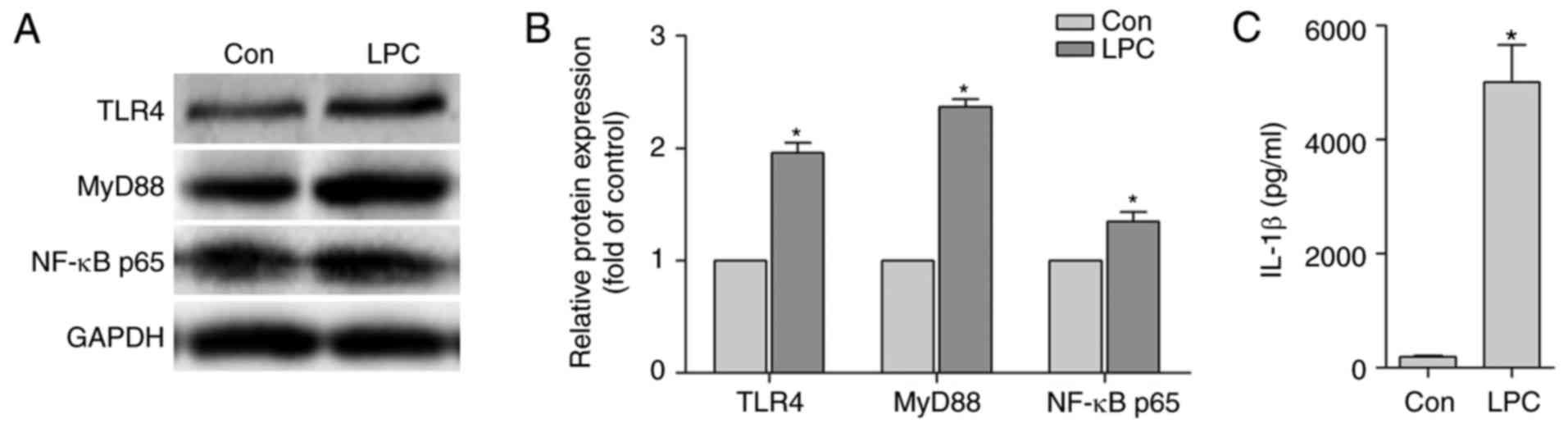
LPC promotes the activation of
TLR4/MyD88/NFκ-B signaling pathway in HUVE-12 cells co-cultured
with MPs
It has been shown that the TLR4/MyD88/NF-κB
transduction pathway is an important event in inflammation and in
the subsequent secretion of IL-1β (36). Consequently, the change in the
expression levels of TLR4 and its downstream signaling molecules
was monitored in LPC-injured HUVE-12 cells. The supernatants
collected from the MPs following co-culture with HUVE-12 cells for
24 h were obtained. It was noted that the expression levels of
TLR4, MyD88 and NF-κB p65 in HUVE-12 cells were significantly
upregulated (Fig. 2A and B) and
the secretion of IL-1β in the co-culture supernatant was
effectively increased (Fig. 2C).
Taken together, these results suggested that LPC promoted the
activation of the TLR4/MyD88/NF-κB signaling pathway in HUVE-12
cells. This pathway is essential in the cross-talk between
LPC-injured-HUVE-12 cells and MPs.
DFMG inhibits the activation of MPs
induced by co-culture with injured HUVE-12 cells
To determine whether DFMG was able to attenuate the
activation of MPs induced by co-culture with LPC-oxidative damaged
HUVE-12 cells, the HUVE-12 cells were treated with various
concentrations (0.3, 1.0 and 3.0 µM) of DFMG for 1 h prior
to LPC (30 µM) treatment. DFMG effectively inhibited the
proliferation of MPs at a concentration range of 0.3–3.0 µM
(Fig. 3A), and treatment at
concentrations of 1.0–3.0 µM DFMG effectively suppressed the
migration (Fig. 3B and C) and the
formation of foam cells (Fig. 3D and
E) in a dose-dependent manner.
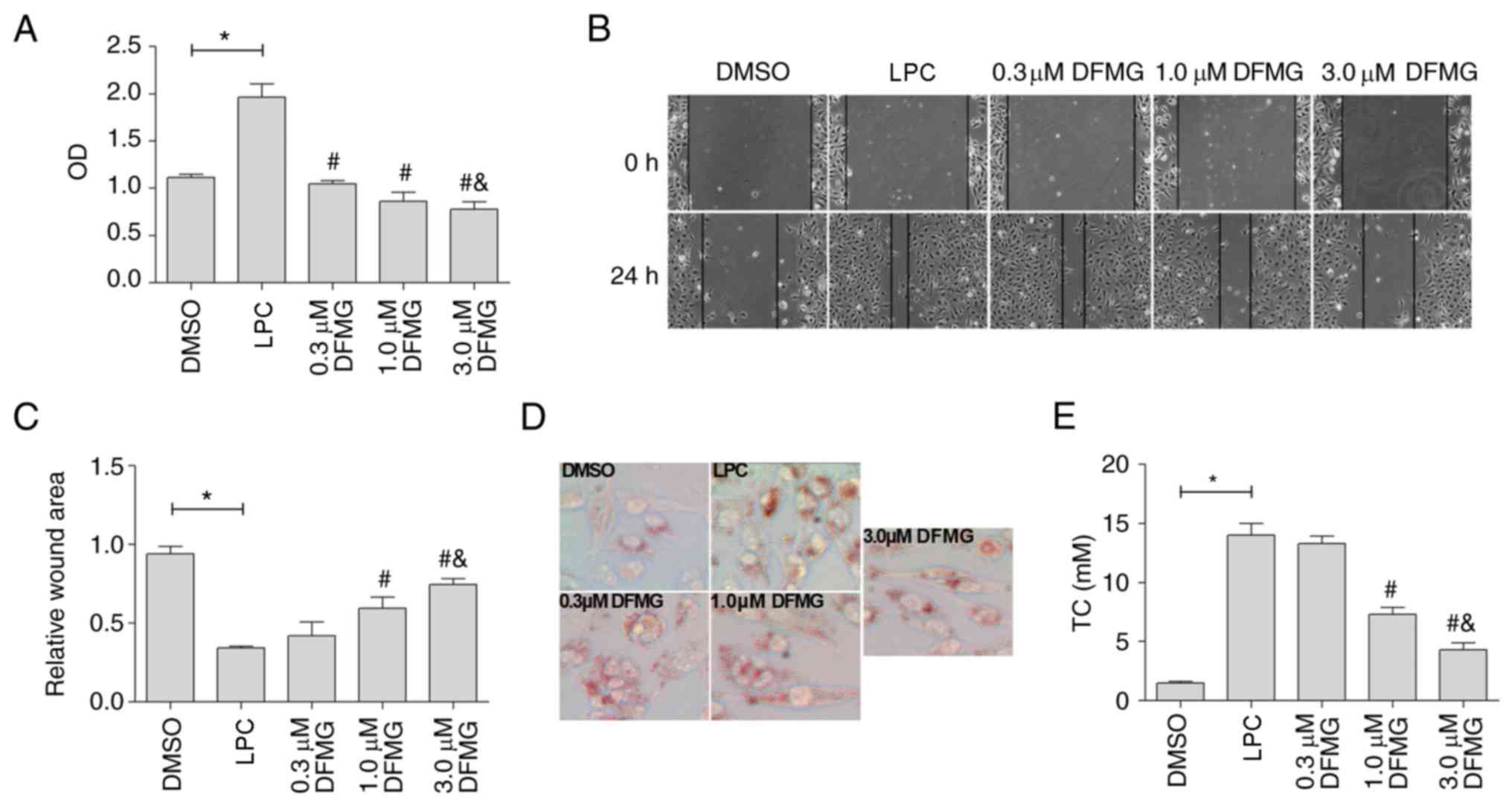 | Figure 3DFMG attenuates the activation of MPs
induced by co-culture with LPC-injured HUVE-12 cells in a
dose-dependent manner. (A) DFMG (0.3, 1.0 and 3.0 µM)
effectively decreased cell viability of MPs co-cultured with
LPC-injured HUVE-12 cells. (B) Images and (C) quantification
showing that DFMG (1.0 and 3.0 µM) effectively increased the
relative wound area of MPs (magnification, ×100). (D) Images and
(E) graph show that DFMG (1.0 and 3.0 µM) significantly
reduced the lipid accumulation in MPs (magnification, ×200). The
data are presented as the mean ± standard deviation of three
separate experiments. *P<0.05, vs. DMSO group,
#P<0.05, vs. LPC group, &P<0.05,
vs. 0.3 or 1.0 µM DFMG group. LPC, lysophosphatidylcholine;
DFMG, 7-difluoromethoxy-5,4′-dimethoxy-genistein; MPs, macrophages;
DMSO, dimethyl sulfoxide. |
DFMG inhibits the TLR4/MyD88/NF-κB
signaling pathway in HUVE-12 cells
As mentioned above, activation of the
TLR4/MyD88/NF-κB signaling pathway in HUVE-12 cells was associated
with the cross-talk between HUVE-12 cells and MPs. The molecular
mechanism underlying the effect of DFMG, in relation to the
activation of MPs induced by co-culture with injured HUVE-12 cells,
was examined by investigating the expression levels of TLR4, MyD88
and NF-κB p65 in the HUVE-12 cells co-cultured with MPs. In
addition, the concentration of IL-1β in the co-culture supernatant
was monitored using ELISA. The data indicated that DFMG (1.0 and/or
3.0 µM) effectively reduced the expression levels of TLR4,
MyD88 and NF-κB p65 (Fig. 4A–D)
and the secretion of IL-1β (Fig.
4E). These observations suggested that DFMG attenuated the
activation of MPs induced by co-culture with injured HUVE-12 cells,
at least partly, via inhibition of the TLR4/MyD88/NF-κB signaling
pathway in the LPC-injured HUVE-12 cells. Accordingly, 3.0
µM was selected as the optimum concentration of DFMG to be
used for the following experiments.
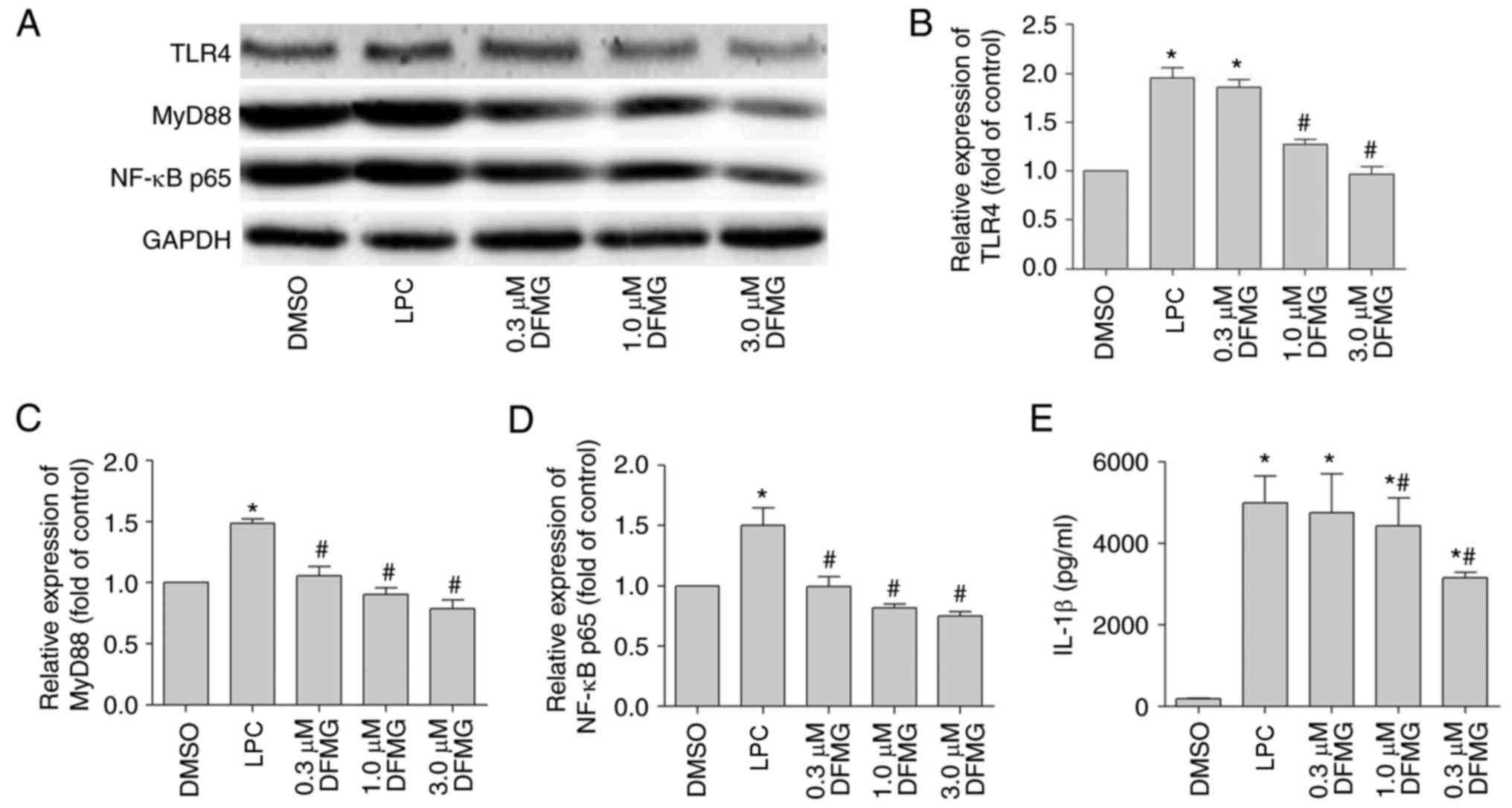 | Figure 4DFMG represses activation of the
TLR4/MyD88/NF-κB pathway in LPC-injured HUVE-12 cells co-cultured
with MPs, and reduces the generation of IL-1β in the co-culture
supernatant. (A) Representative western blots of protein expression
levels of TLR4, MyD88 and NF-κB p65 in HUVE-12 cells co-cultured
with MPs. Densitometric analysis was used to quantify the protein
levels of (B) TLR4, (C) MyD88 and (D) NF-κB p65. DFMG (0.3, 1.0 and
3.0 µM) downregulated the expression of these proteins. (E)
DFMG (1.0 and 3.0 µM) decreased the concentration of IL-1β
in the co-culture supernatant. The data are presented as the mean ±
standard deviation of three separate experiments.
*P<0.05, vs. DMSO group, #P<0.05, vs.
LPC group. LPC, lysophosphatidylcholine; DFMG,
7-difluoromethoxy-5,4′-dimethoxy-genistein; MPs, macrophages; TLR4,
Toll-like receptor 4; MyD88, myeloid differentiation factor 88;
NF-κB, nuclear factor-κB; IL-1β, interleukin-1β; DMSO, dimethyl
sulfoxide; OD, optical density. |
TLR4 signaling pathway is involved in the
activation of MPs induced by co-culture with injured HUVE-12
cells
To further confirm the effect of the TLR4 signaling
pathway on the activation of MPs induced by co-culture with injured
HUVE-12 cells, the HUVE-12 cells were pre-incubated with DFMG (3.0
µM) for 1 h, and CLI-095 (5 µg/ml) or IL-1Ra (10
µg/ml) were added for 24 h. Subsequently, LPC (30 µM)
was added and incubated for another 24 h, followed by co-culture
with MPs for 24 h. The results indicated that DFMG, CLI-095 and
IL-1Ra significantly decreased the proliferation and migration of
MPs (Fig. 5A–C) and the formation
of foam cells (Fig. 5D and E),
which were induced by co-culture with injured HUVE-12 cells. In
addition, DFMG with CLI-095 or IL-1Ra exhibited synergistic effects
in promoting the stability of MPs. These data indicated that the
inhibition of TLR4 or its downstream targets in HUVE-12 cells
effectively attenuated the activation of MPs induced by co-culture
with LPC-injured HUVE-12 cells. The results also confirmed that the
TLR4 signaling pathway in LPC-injured HUVE-12 cells was associated
with the activation of MPs induced by co-culture.
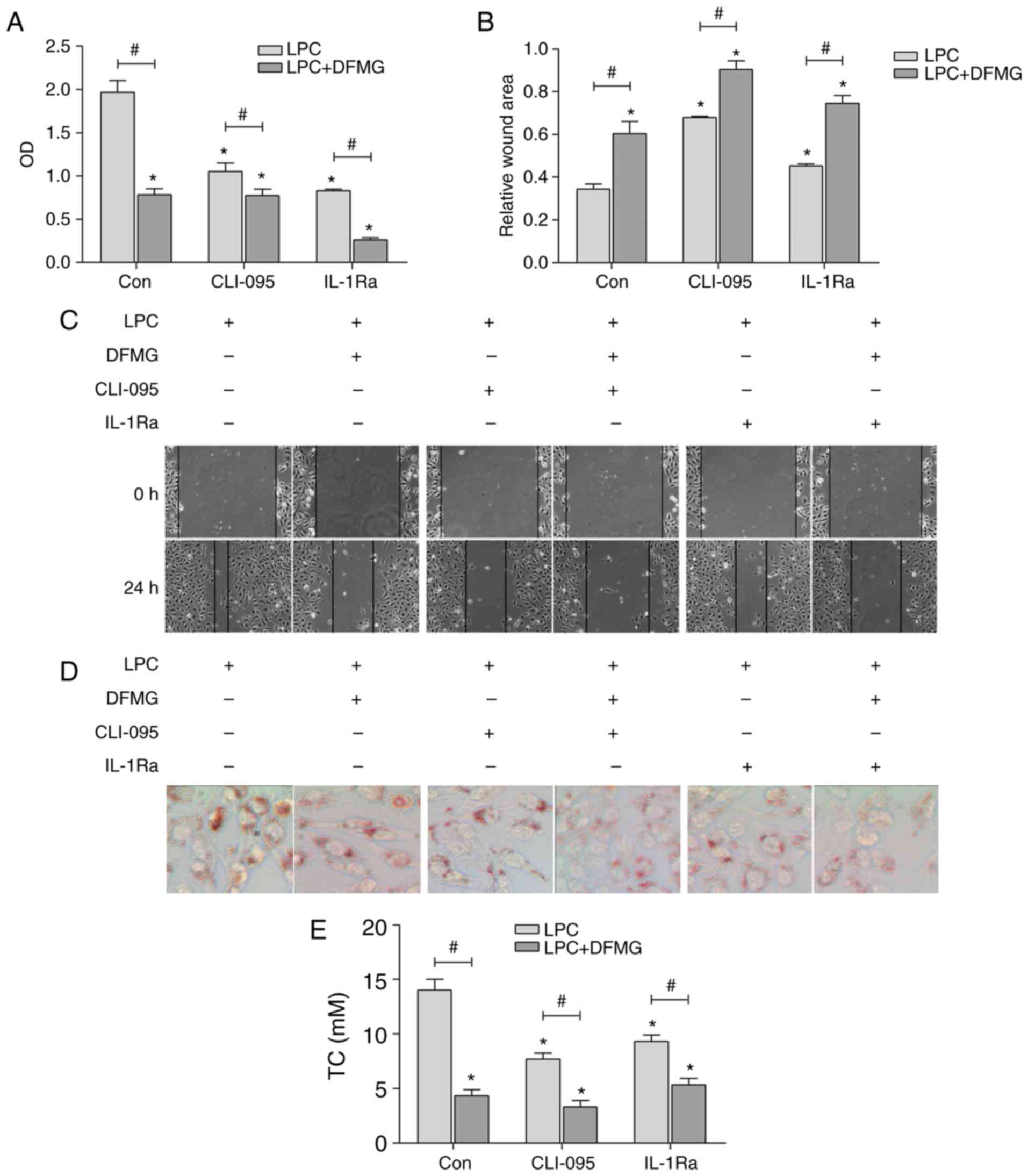 | Figure 5TLR4/MyD88/NF-κB pathway is involved
in the activation of MPs induced by co-culture with LPC-injured
HUVE-12 cells. (A) CLI-095 and IL-1Ra effectively reduced the cell
viability of MPs co-cultured with LPC-injured HUVE-12 cells, which
were enhanced by co-treatment with DFMG (magnification, ×200). (B)
Graph and (C) images of wound healing assay showed that CLI-095 and
IL-1Ra effectively increased the relative wound area of MPs, which
were promoted by DFMG (magnification, ×100). (D) Images and (E)
quantification showed that CLI-095 and IL-1Ra effectively reduced
lipid accumulation in MPs co-cultured with LPC-injured HUVE-12
cells, which were enhanced by co-treatment with DFMG
(magnification, ×200). The data are presented as the mean ±
standard deviation of three separate experiments.
*P<0.05, vs. Con (without DFMG pretreatment) group,
#P<0.05, vs. adjacent group. LPC,
lysophos-phatidylcholine; DFMG,
7-difluoromethoxy-5,4′-dimethoxy-genistein; MPs, macrophages; TLR4,
Toll-like receptor 4; MyD88, myeloid differentiation factor 88;
NF-κB, nuclear factor-κB; IL-1Ra, interleukin-1 receptor
antagonist; OD, optical density. |
DFMG attenuates activation of the
TLR4/MyD88/NF-κB signaling pathway in LPC-injured HUVE-12 cells
co-cultured with MPs
DFMG (3.0 µM), CLI-095 (5 µg/ml),
IL-1Ra (10 µg/ml) and LPC (30 µM) were incubated with
HUVE-12 cells, as described above, in order to investigate the
molecular mechanism underlying the effect of DFMG on the inhibition
of the activation of MPs induced by co-culture with LPC-injured
HUVE-12 cells. The results indicated that DFMG, CLI-095 and IL-1Ra
significantly downregulated the expression levels of TLR4, MyD88
and NF-κB p65 (Fig. 6A–D) in the
HUVE-12 cells and decreased the secretion of IL-1β (Fig. 6E) in the co-culture supernatant.
Furthermore, DFMG and CLI-095 or IL-1Ra exhibited synergistic
effects in the inhibition of the TLR4 signaling pathway.
Consequently, DFMG suppressed activation of the TLR4/MyD88/NF-κB
signaling pathway in LPC-injured HUVE-12 cells following co-culture
with MPs.
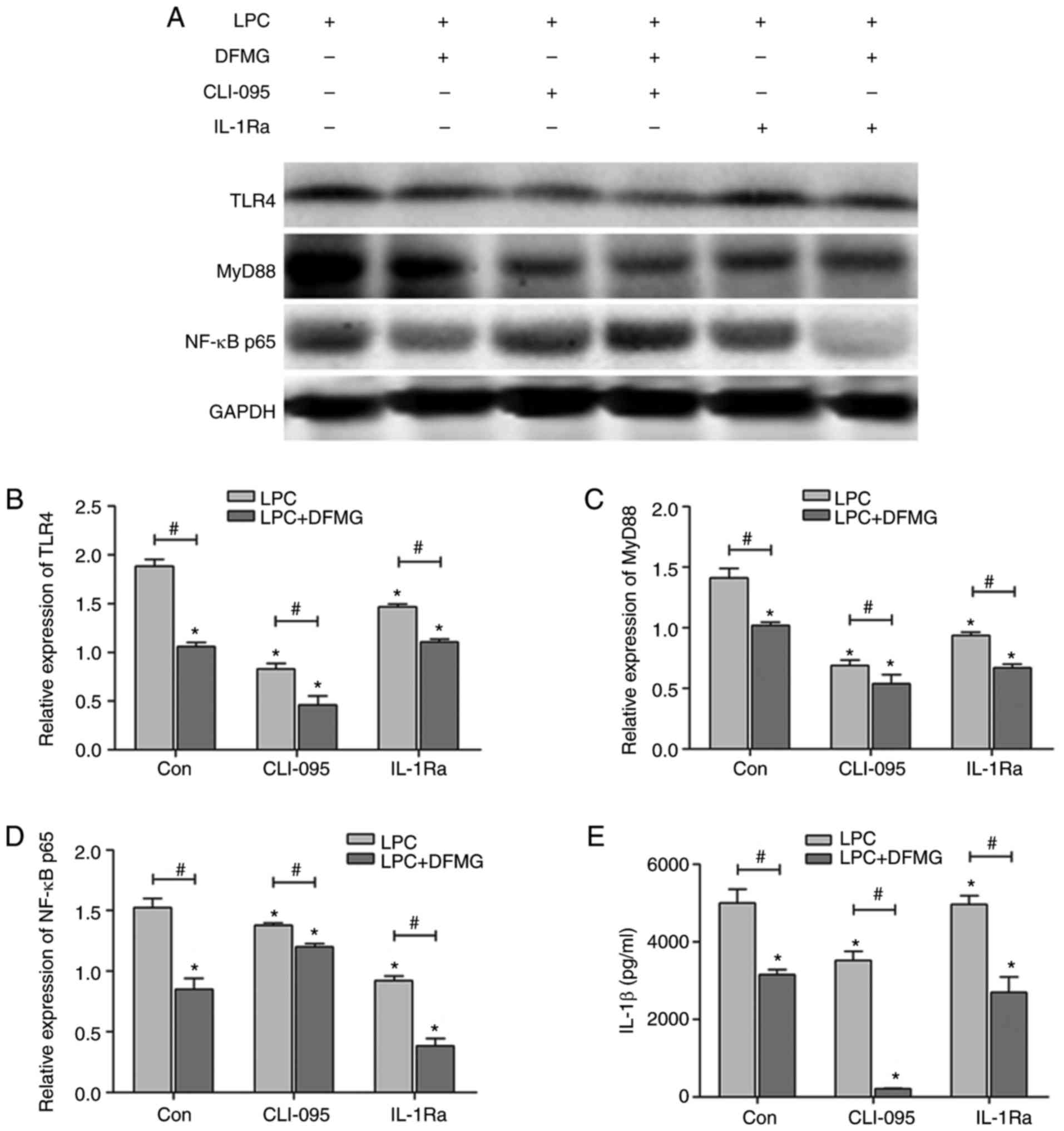 | Figure 6DFMG inhibits the TLR4/MyD88/NF-κB
pathway in LPC-injured HUVE-12 cells co-cultured with MPs, and
inhibits the production of IL-1β in the co-culture supernatant. (A)
Representative western blots of protein expression of TLR4, MyD88
and NF-κB p65 in LPC-injured HUVE-12 cells co-cultured with MPs.
Densitometric analysis and ELISA were used to quantify the
expression levels of (B) TLR4, (C) MyD88, (D) NF-κB p65 and the (E)
concentration of IL-1β in the co-culture supernatant. CLI-095 and
IL-1Ra downregulated the expression of these proteins and reduced
the generation of IL-1β, which was promoted by co-treatment with
DFMG. The data are presented as the mean ± standard deviation of
three separate experiments. *P<0.05, vs. Con (without
DFMG pretreatment), #P<0.05, vs. adjacent group. LPC,
lysophosphatidylcholine; DFMG,
7-difluoromethoxy-5,4′-dimethoxy-genistein; MPs, macrophages; TLR4,
Toll-like receptor 4; MyD88, myeloid differentiation factor 88;
NF-κB, nuclear factor-κB; IL-1Ra, interleukin-1 receptor
antagonist. |
Discussion
MP dysregulation is considered a risk factor for
several inflammatory complications, including AS and cancer
(37). The process of
atherogenesis involves several types of cells, notably ECs, smooth
muscle cells, monocytes and macrophages. AS initiates from the
endothelium of the arterial wall (7). ECD is a vital contributor to the
pathobiology of atherosclerotic cardiovascular disease (8). Previous studies have shown that ECD
induces the functional changes of MPs through the mutual cross-talk
between cells in the vascular microenvironment (12,38). The in vitro Transwell
co-culture model can be used to examine the effects of ECD on MPs.
Oxidative stress is key in the progression of AS (39). LPC is a major component of ox-LDL,
which can cause AS (40). In the
present study, LPC stimulated the generation of ROS and the release
of LDH (cell oxidative damage index) in ECs, but promoted the
activation of MPs induced by co-culture with EC. The results also
indicated that LPC increased the expression levels of TLR4, MyD88
and NF-κB p65 in the ECs co-cultured with THP-1-derived MPs. LPC
also increased the secretion of IL-1β in the co-culture
supernatant. These findings demonstrated that LPC exhibited
oxidative effects on ECs. In addition to these observations, ECD
promoted the activation of MPs, which may be associated with
activation of the TLR4/MyD88/NF-κB signaling pathway in ECs.
AS, the main cause of cardiovascular disease, is
characterized by the accumulation of inflammatory cells in the
artery wall (41). TLRs are
crucial in the initiation of an innate immune response through
activating the inflammatory cells (42). To confirm that the TLR4 signaling
pathway was involved in the activation of macrophages induced by
co-culture with LPC-injured HUVE-12 cells, specific inhibitors of
the proteins and the mediators acting on this pathway were used.
CLI-095, also known as TAK-242, is a novel cyclohexene derivative,
which selectively suppresses TLR4 signaling, potently suppressing
ligand-dependent and ligand-independent signaling of TLR4 (43,44). IL-1 belongs to the innate immune
system and is important in the initiation of the
immunoinflam-matory cascade reaction (45). IL-1Ra is a specific and pure
antagonist of the IL-1 receptor family (46), which has the capacity to inhibit
inflammasome activation and control the balance between the
pro-inflammatory and anti-inflammatory response (47). As expected, treatment of
LPC-injured EC with CLI-095 and IL-1Ra significantly impaired the
proliferation and migration of MPs, and the formation of foam cells
induced by co-culture with injured ECs. Taken together, the data
demonstrated that activation of the TLR4 signaling pathway in ECs
led to the proliferation, migration and lipid accumulation of MPs
in the co-culture model.
GEN, a primary soy isoflavone, has received
considerable attention as a protein kinase inhibitor (48). In addition, studies have
demonstrated that GEN possesses atheroprotective effects via
regulation of the TLR4/NF-κB signaling pathway (49,50). GEN exhibits low bioavailability
and intestinal absorption in vivo, and its clinical
application is limited (51).
DFMG is synthesized using GEN as a precursor. DFMG exhibits
improved bioavailability and absorption. Our previous studies
indicated that DFMG was more efficient than GEN in reducing the
risk of cardiovascular disease by the inhibition of oxidative
damage, mitochondrial apoptosis and/or inhibition of the TLR4
signaling pathway (33-35). In the present study, DFMG was
shown to effectively attenuate the activation of THP-1-derived MPs
induced by co-culture with LPC-injured ECs. Furthermore, inhibition
of the TLR4 signaling pathway using specific inhibitors (CLI-095
and IL-1Ra) increased the anti-inflammatory potential of DFMG.
These observations indicated that DFMG may exert its
anti-inflammatory activity through inhibition of the TLR4 signaling
pathway.
In view of the aforementioned results, the
expression levels of TLR4 were further assessed. In the present
study, DFMG significantly downregulated the protein expression
levels of the aforementioned indicators (TLR4, MyD88 and NF-κB p65)
in ECs, and the levels of IL-1β in the co-culture supernatant.
CLI-095 and IL-1Ra markedly reduced the activation of TLR4, which
was markedly enhanced by DFMG treatment. The endogenous IL-1Ra has
been shown to regulate the extent of TLR9-induced liver damage
(52), whereas IL-1Ra deficiency
can promote TLR4-dependent arthritis (53). The present study hypothesized that
DFMG may act by upregulating the expression of endogenous IL-1ra in
order to inhibit the activity of TLR4.
Considering the above findings, it was suggested
that the atheroprotective effects of DFMG against the activation of
MPs induced by co-culture with LPC-injured ECs was mediated via
regulation of the TLR4/MyD88/NF-κB signaling pathway in ECs.
However, the effects of DFMG on MPs were not examined in
vivo. Consequently, the detailed effects of DFMG on the TLR4
signaling network in AS require further investigation using animal
knockout models. TLR4 signaling is also crucial in other diseases,
including systemic lupus erythematosus (29), non-small cell lung cancer
(54), idiopathic pulmonary
fibrosis (55), myocardial
inflammation (56) and type 2
diabetes (57). According to the present study, it was hypothesized
that DFMG may prevent and/or treat these diseases via inhibiting
the TLR4 signaling pathway.
Glossary
Abbreviations
Abbreviations:
|
DFMG
|
7-difluoromethoxy-5,4′-dimethoxy-genistein
|
|
ECs
|
endothelial cells
|
|
MPs
|
macrophages
|
|
HUVE-12
|
human umbilical vein endothelial
cell
|
|
LPC
|
lysophosphatidylcholine
|
|
ROS
|
reactive oxygen species
|
|
LDH
|
lactate dehydrogenase
|
|
AS
|
atherosclerosis
|
|
ECD
|
endothelial cell dysfunction
|
|
TNF-α
|
tumor necrosis factor-α
|
|
MCP-1
|
monocyte chemoattractant protein-1
|
|
TLR4
|
Toll-like receptor 4
|
|
MyD88
|
myeloid differentiation factor 88
|
|
NF-κB
|
nuclear factor-κB
|
|
ox-LDL
|
oxidized low-density lipoprotein
|
|
LPS
|
lipopolysaccharide
|
|
GEN
|
genistein
|
|
PMA
|
phorbol 12-myristate 13-acetate
|
|
DMSO
|
dimethyl sulfoxide
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
THP-1
|
human acute monocytic leukemia
cells
|
|
RPMI
|
Roswell Park Memorial Institute
|
|
FBS
|
fetal bovine serum
|
|
TC
|
total cholesterol ELISA, enzyme-linked
immunosorbent assay
|
Acknowledgments
Not applicable.
Notes
[1]
Funding
Financial support was provided by the Natural
Science Foundation of China (grant no. 81370382).
[2] Availability
of data and materials
All data generated or analysed during this study
were included in this published article.
[3] Authors'
contributions
XF designed the study and prepare the manuscript. LC
and SY performed the experiments. YZ analysed the data. JC revised
the manuscript. All authors read and approved the final
manuscript.
[4] Ethics
approval and consent to participate
Not applicable.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Cochain C and Zernecke A: Macrophages in
vascular inflammation and atherosclerosis. Pflugers Arch.
469:485–499. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gerrity RG, Naito HK, Richardson M and
Schwartz CJ: Dietary induced atherogenesis in swine. Morphology of
the intima in prelesion stages. Am J Pathol. 95:775–792.
1979.PubMed/NCBI
|
|
3
|
Galkina E and Ley K: Immune and
inflammatory mechanisms of atherosclerosis (*). Annu Rev Immunol.
27:165–197. 2009. View Article : Google Scholar
|
|
4
|
Weber C and Noels H: Atherosclerosis:
Current pathogenesis and therapeutic options. Nat Med.
17:1410–1422. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tabas I: Consequences and therapeutic
implications of macrophage apoptosis in atherosclerosis: The
importance of lesion stage and phagocytic efficiency. Arterioscler
Thromb Vasc Biol. 25:2255–2264. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ilhan F and Kalkanli ST: Atherosclerosis
and the role of immune cells. World J Clin Cases. 3:345–352. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Husain K, Hernandez W, Ansari RA and
Ferder L: Inflammation, oxidative stress and renin angiotensin
system in atherosclerosis. World J Biol Chem. 6:209–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gimbrone MA Jr and García-Cardeña G:
Endothelial cell dysfunction and the pathobiology of
atherosclerosis. Circ Res. 118:620–636. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tabas I: Heart disease: Death-defying
plaque cells. Nature. 536:32–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Linton MF, Babaev VR, Huang J, Linton EF,
Tao H and Yancey PG: Macrophage apoptosis and efferocytosis in the
pathogenesis of atherosclerosis. Circ J. 80:2259–2268. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tabas I and Bornfeldt KE: Macrophage
phenotype and function in different stages of atherosclerosis. Circ
Res. 118:653–667. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kalucka J, Bierhansl L, Wielockx B,
Carmeliet P and Eelen G: Interaction of endothelial cells with
macrophages-linking molecular and metabolic signaling. Pflugers
Arch. 469:473–483. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pryshchep O, Ma-Krupa W, Younge BR,
Goronzy JJ and Weyand CM: Vessel-specific Toll-like receptor
profiles in human medium and large arteries. Circulation.
118:1276–1284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takeuchi O and Akira S: Pattern
recognition receptors and inflammation. Cell. 140:805–820. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ghosh G, Wang VY, Huang DB and Fusco A:
NF-κB regulation: Lessons from structures. Immunol Rev. 246:36–58.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Feng Y, Cai ZR, Tang Y, Hu G, Lu J, He D
and Wang S: TLR4/NF-κB signaling pathway-mediated and oxLDL-induced
up-regulation of LOX-1, MCP-1, and VCAM-1 expressions in human
umbilical vein endothelial cells. Genet Mol Res. 13:680–695. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zheng Z, Yuan R, Song M, Huo Y, Liu W, Cai
X, Zou H, Chen C and Ye J: The toll-like receptor 4-mediated
signaling pathway is activated following optic nerve injury in
mice. Brain Res. 1489:90–97. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z and Wu L: Melatonin alleviates
secondary brain damage and neurobehavioral dysfunction after
experimental subarachnoid hemorrhage: Possible involvement of
TLR4-mediated inflammatory pathway. J Pineal Res. 55:399–408.
2013.PubMed/NCBI
|
|
19
|
Palová-Jelínková L, Dáňová K, Drašarová H,
Dvořák M, Funda DP, Fundová P, Kotrbová-Kozak A, Černá M, Kamanová
J, Martin SF, et al: Pepsin digest of wheat gliadin fraction
increases production of IL-1β via TLR4/MyD88/TRIF/MAPK/NF-κB
signaling pathway and an NLRP3 inflammasome activation. PLoS One.
8:e624262013. View Article : Google Scholar
|
|
20
|
Nicoletti F, Patti F, DiMarco R, Zaccone
P, Nicoletti A, Meroni P and Reggio A: Circulating serum levels of
IL-1ra in patients with relapsing remitting multiple sclerosis are
normal during remission phases but significantly increased either
during exacerbations or in response to IFN-beta treatment.
Cytokine. 8:395–400. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Netea MG, Simon A, van de Veerdonk F,
Kullberg BJ, Van der Meer JW and Joosten LA: IL-1beta processing in
host defense: Beyond the inflammasomes. PLoS Pathog.
6:e10006612010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Piccini A, Carta S, Tassi S, Lasiglié D,
Fossati G and Rubartelli A: ATP is released by monocytes stimulated
with pathogen-sensing receptor ligands and induces IL-1beta and
IL-18 secretion in an autocrine way. Proc Natl Acad Sci USA.
105:8067–8072. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gu H, Jiao Y, Yu X, Li X, Wang W, Ding L
and Liu L: Resveratrol inhibits the IL-1β-induced expression of
MMP-13 and IL-6 in human articular chondrocytes via
TLR4/MyD88-dependent and -independent signaling cascades. Int J Mol
Med. 2017.Epub ahead of print. View Article : Google Scholar
|
|
24
|
Liu L, Gu H, Liu H, Jiao Y, Li K, Zhao Y,
An L and Yang J: Protective effect of resveratrol against
IL-1β-induced inflammatory response on human osteoarthritic
chondrocytes partly via the TLR4/MyD88/NF-κB signaling pathway: An
'in vitro study'. Int J Mol Sci. 15:6925–6940. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Feng Y, Cui Y, Gao JL, Li MH, Li R, Jiang
XH, Tian YX, Wang KJ, Cui CM and Cui JZ: Resveratrol attenuates
neuronal autophagy and inflammatory injury by inhibiting the
TLR4/NF-κB signaling pathway in experimental traumatic brain
injury. Int J Mol Med. 37:921–390. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee HH, Han MH, Hwang HJ, Kim GY, Moon SK,
Hyun JW, Kim WJ and Choi YH: Diallyl trisulfde exerts
anti-inflammatory effects in lipopolysaccharide-stimulated RAW
264.7 macrophages by suppressing the Toll-like receptor 4/nuclear
factor-κB pathway. Int J Mol Med. 35:487–495. 2015. View Article : Google Scholar
|
|
27
|
Zhu S, Tang S and Su F: Dioscin inhibits
ischemic stroke-induced inflammation through inhibition of the
TLR4/MyD88/NF-κB signaling pathway in a rat model. Mol Med Rep.
17:660–666. 2018.
|
|
28
|
Stojanovic I, Cuzzocrea S, Mangano K,
Mazzon E, Miljkovic D, Wang M, Donia M, Al Abed Y, Kim J, Nicoletti
F, et al: In vitro, ex vivo and in vivo immunopharmacological
activities of the isoxazoline compound VGX-1027: Modulation of
cytokine synthesis and prevention of both organ-specific and
systemic autoimmune diseases in murine models. Clin Immunol.
123:311–323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fagone P, Muthumani K, Mangano K, Magro G,
Meroni PL, Kim JJ, Sardesai NY, Weiner DB and Nicoletti F: VGX-1027
modulates genes involved in the lipopolysaccharide-induced
toll-like receptor 4 activation and in a murine model of systemic
lupus erythematosus. Immunology. 142:594–602. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pangeni R, Sahni JK, Ali J, Sharma S and
Baboota S: Resveratrol: Review on therapeutic potential and recent
advances in drug delivery. Expert Opin Drug Deliv. 11:1285–1298.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fu XH, Wang L, Zhao H, Xiang HL and Cao
JG: Synthesis of genistein derivatives and determination of their
protective effects against vascular endothelial cell damages caused
by hydrogen peroxide. Bioorg Med Chem Lett. 18:513–517. 2008.
View Article : Google Scholar
|
|
32
|
Zhao H, Li C, Cao JG, Xiang HL, Yang HZ,
You JL, Li CL and Fu XH: 7-Difluoromethyl-5,4′-dimethoxygenistein,
a novel genistein derivative, has therapeutic effects on
atherosclerosis in a rabbit model. J Cardiovasc Pharmacol.
54:412–420. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang L, Zheng X, Xiang HL, Fu XH and Cao
JG: 7-Difluoromethyl-5,4′-dimethoxygenistein inhibits oxidative
stress induced adhesion between endothelial cells and monocytes via
NF-kappaB. Eur J Pharmacol. 605:31–35. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu S, Li L, Zhang J, Huang H, Yang S, Ren
C, Fu X and Zhang Y: 7-Difluoromethyl-5, 4′-dimethoxygenistein
reverses LPC-induced apoptosis of HUVE-12 cells through regulating
mitochondrial apoptosis pathway. Curr Signal Transd T. 9:50–58.
2014. View Article : Google Scholar
|
|
35
|
Liu F, Cao JG, Li C, Tan JS and Fu XH:
Protective effects of 7-difluoromethyl-5,4′-dimethoxygenistein
against human aorta endothelial injury caused by lysophosphatidyl
choline. Mol Cell Biochem. 363:147–155. 2012. View Article : Google Scholar
|
|
36
|
Kong F, Ye B, Cao J, Cai X, Lin L, Huang
S, Huang W and Huang Z: Curcumin represses NLRP3 inflammasome
activation via TLR4/MyD88/NF-κB and P2X7R signaling in PMA-induced
macrophages. Front Pharmacol. 7:3692016. View Article : Google Scholar
|
|
37
|
Eaton KV, Yang HL, Giachelli CM and
Scatena M: Engineering macrophages to control the inflammatory
response and angiogenesis. Exp Cell Res. 339:300–309. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Im GI: Coculture in musculoskeletal tissue
regeneration. Tissue Eng Part B Rev. 20:545–554. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang L, Huang Z, Huang W, Chen X, Shan P,
Zhong P, Khan Z, Wang J, Fang Q, Liang G and Wang Y: Inhibition of
epidermal growth factor receptor attenuates atherosclerosis via
decreasing inflammation and oxidative stress. Sci Rep. 8:459172017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Voight BF, Peloso GM, Orho-Melander M,
Frikke-Schmidt R, Barbalic M, Jensen MK, Hindy G, Hólm H, Ding EL,
Johnson T, et al: Plasma HDL cholesterol and risk of myocardial
infarction: A mendelian randomisation study. Lancet. 380:572–580.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Canfrán-Duque A, Rotllan N, Zhang X,
Fernández-Fuertes M, Ramírez-Hidalgo C, Araldi E, Daimiel L, Busto
R, Fernández-Hernando C and Suárez Y: Macrophage deficiency of
miR-21 promotes apoptosis, plaque necrosis, and vascular
inflammation during atherogenesis. EMBO Mol Med. 9:1244–1262. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Szatmary Z: Molecular biology of toll-like
receptors. Gen Physiol Biophys. 31:357–366. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ii M, Matsunaga N, Hazeki K, Nakamura K,
Takashima K, Seya T, Hazeki O, Kitazaki T and Iizawa Y: A novel
cyclohexene derivative, ethyl
(6R)-6-[N-(2-Chloro-4-fluorophenyl)sulfamoyl]
cyclohex-1-ene-1-carboxylate (TAK-242), selectively inhibits
toll-like receptor 4-mediated cytokine production through
suppression of intracellular signaling. Mol Pharmacol.
69:1288–1295. 2006. View Article : Google Scholar
|
|
44
|
Kawamoto T, Ii M, Kitazaki T, Iizawa Y and
Kimura H: TAK-242 selectively suppresses Toll-like receptor
4-signaling mediated by the intracellular domain. Eur J Pharmacol.
584:40–48. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
D ujmovic I, Mangano K, Pekmezovic T,
Quattrocchi C, Mesaros S, Stojsavljevic N, Nicoletti F and Drulovic
J: The analysis of IL-1 beta and its naturally occurring inhibitors
in multiple sclerosis: The elevation of IL-1 receptor antagonist
and IL-1 receptor type II after steroid therapy. J Neuroimmunol.
207:101–106. 2009. View Article : Google Scholar
|
|
46
|
van Oosten BW, Lai M, Hodgkinson S,
Barkhof F, Miller DH, Moseley IF, Thompson AJ, Rudge P, McDougall
A, McLeod JG, et al: Treatment of multiple sclerosis with the
monoclonal anti-CD4 antibody cM-T412: Results of a randomized,
double-blind, placebo-controlled, MR-monitored phase II trial.
Neurology. 49:351–357. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yoon GS, Sud S, Keswani RK, Baik J,
Standiford TJ, Stringer KA and Rosania GR: Phagocytosed Clofazimine
Biocrystals can modulate innate immune signaling by inhibiting TNFα
and boosting IL-1RA secretion. Mol Pharm. 12:2517–2527. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jeong JW, Lee HH, Han MH, Kim GY, Kim WJ
and Choi YH: Anti-inflammatory effects of genistein via suppression
of the toll-like receptor 4-mediated signaling pathway in
lipopoly-saccharide-stimulated BV2 microglia. Chem Biol Interact.
212:30–39. 2014. View Article : Google Scholar : PubMed/NCBI
Zhou X, Yuan L, Zhao X, Hou C, Ma W, Yu H
and Xiao R: Genistein antagonizes inflammatory damage induced by
β-amyloid peptide in microglia through TLR4 and NF-κB. Nutrition.
30:90–95. 2014. View Article : Google Scholar
|
|
49
|
Ma W, Ding B, Yu H, Yuan L, Xi Y and Xiao
R: Genistein alleviates β-amyloid-induced inflammatory damage
through regulating Toll-like receptor 4/nuclear factor κB. J Med
Food. 18:273–279. 2015. View Article : Google Scholar :
|
|
50
|
Cave NJ, Backus RC, Marks SL and Klasing
KC: The bioavailability and disposition kinetics of genistein in
cats. J Vet Pharmacol Ther. 30:327–335. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Petrasek J, Dolganiuc A, Csak T,
Kurt-Jones EA and Szabo G: Type I interferons protect from
Toll-like receptor 9-associated liver injury and regulate IL-1
receptor antagonist in mice. Gastroenterology. 140:697–708.e4.
2011. View Article : Google Scholar :
|
|
52
|
Rogier R, Ederveen THA, Boekhorst J,
Wopereis H, Scher JU, Manasson J, Frambach SJCM, Knol J, Garssen J,
van der Kraan PM, et al: Aberrant intestinal microbiota due to IL-1
receptor antagonist deficiency promotes IL-17 and TLR4-dependent
arthritis. Microbiome. 5:632017. View Article : Google Scholar
|
|
53
|
Li D, Jin Y, Sun Y, Lei J and Liu C:
Knockdown of toll-like receptor 4 inhibits human NSCLC cancer cell
growth and inflammatory cytokine secretion in vitro and in vivo.
Int J Oncol. 45:813–821. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Samara KD, Antoniou KM, Karagiannis K,
Margaritopoulos G, Lasithiotaki I, Koutala E and Siafakas NM:
Expression profiles of Toll-like receptors in non-small cell lung
cancer and idiopathic pulmonary fbrosis. Int J Oncol. 40:1397–1404.
2012.PubMed/NCBI
|
|
55
|
Yang Y, Lv J, Jiang S, Ma Z, Wang D, Hu W,
Deng C, Fan C, Di S, Sun Y and Yi W: The emerging role of Toll-like
receptor 4 in myocardial inflammation. Cell Death Dis. 7:e22342016.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cha JJ, Hyun YY, Lee MH, Kim JE, Nam DH,
Song HK, Kang YS, Lee JE, Kim HW, Han JY and Cha DR: Renal
protective effects of toll-like receptor 4 signaling blockade in
type 2 diabetic mice. Endocrinology. 154:2144–2155. 2013.
View Article : Google Scholar : PubMed/NCBI
|