Introduction
Ischemic stroke is a leading cause of mortality and
disability worldwide (1).
Survivors typically suffer from permanent brain damage, resulting
in a reduced quality of life and increased social burden (2). Cerebral blood flow reperfusion is
the primary treatment option for stroke (3). However, cerebral
ischemia/reperfusion (I/R) induces a series of pathophysiological
processes, which may cause further damage, including I/R injury
(4).
Oxidative stress and neuronal apoptosis are
important factors in the pathological process of I/R injury that
occurs in cerebral ischemic stroke followed by reperfusion
(5). Oxidative stress refers to a
comparative surplus of reactive oxygen species (ROS) caused by an
imbalance between oxidants and antioxidants (6). The brain tissue is sensitive to
oxidative stress as it contains low levels of endogenous
antioxidant enzymes, including superoxide dismutase (SOD),
glutathione-peroxidase (GSH-Px), and catalase (CAT), which act as
cellular defenses against ROS (7). Malondialdehyde (MDA) is primarily
induced by ROS and is commonly used as a biomarker of oxidative
stress (8). Experimental and
clinical studies have demonstrated that the expression of SODs,
CATs and GSH-Px are significantly reduced (7,9-12)
and MDA is significantly elevated in animal models of stroke and in
patients that have experienced an ischemic stroke (12).
Oxidative stress is implicated in the initiation of
apoptosis and it has been suggested that the balance between the
anti-apoptotic protein B-cell lymphoma-2 (Bcl-2) and the
pro-apoptotic apoptosis regulator Bax (BAX) protein regulates
apoptosis (13). BAX mediates the
activation of caspase-9, which is upregulated following ischemia in
human brain tissue (14). In
animal models of ischemic stroke, caspase-9 leads to the activation
of caspase-3, which is considered to be a key mediator of
apoptosis. Wagner et al (15) revealed that cleaved caspase-3 was
predominantly associated with cellular responses to stroke and this
has been supported by the results of other studies (16-18). Therefore, modulation of Bcl-2/BAX
and the suppression of caspase-9, caspase-3 and cleaved caspase-3
expression may serve a protective role in the treatment of cerebral
I/R injury.
Rhein is a naturally occurring anthraquinone
compound isolated from Radix et Rhizoma Rhei and is involved
in a number of biological activities, including the suppression of
ROS, downregulation of intracellular calcium concentration,
promotion of microcirculatory function, inhibition of neutrophil
migration and the promotion of anti-inflammatory effects (19-22). The blood-brain barrier is
disrupted during cerebral I/R injury and previous studies have
revealed that segments of rhein are able to pass through the
disrupted blood-brain barrier (23), inhibit cerebral infarction and
improve the neurological functional score (NFS) (24). Therefore, rhein may have potential
beneficial effects in anti-cerebral I/R injury. However, to the
best of our knowledge, the signaling mechanisms that underlie the
protective effects of rhein against I/R remain unknown.
There have been few previous studies investigating
the effect of rhein on cerebral I/R injury. Therefore, the present
study aimed to evaluate the effects of rhein on ischemic injury,
neurological outcomes, oxidative stress and apoptosis biomarkers in
rats following ischemic stroke induced by middle cerebral artery
occlusion (MCAO).
Materials and methods
Reagents
Rhein (molecular formula,
C15H8O6; molecular weight,
284.225) was purchased from the National Institute for Food and
Drug Control (Beijing, China). Nimodipine tablets were provided by
Bayer AG (Leverkusen, Germany) and 2,3,5-triphenyltetrazolium
chloride (TTC) was obtained from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). A Total Extraction Sample kit and a BCA
protein assay kit were purchased from Nanjing KeyGen Biotech Co.,
Ltd. (Nanjing, China). Antibodies against caspase-9 (cat. no.
ab2013), caspase-3 (cat. no. ab44976) and GAPDH (cat. no. ab9485)
were obtained from Abcam (Cambridge, UK). Cleaved caspase-3 (cat.
no. 9661), Bcl-2 (cat. no. 2876), BAX (cat. no. 2772) and β-actin
(cat. no. 4970) antibodies were procured from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin G (cat. no.
ZB-2301) was obtained from OriGene Technologies, Inc. (Beijing,
China). A Total RNA kit was obtained from Tiangen Biotech Co., Ltd.
(Beijing, China). A RevertAid First Strand cDNA Synthesis kit was
obtained from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) was purchased
fromTakara Bio, Inc. (Otsu, Japan). The total superoxide dismutase
(T-SOD) assay kit (cat. no. A001-1-1), glutathione peroxidase
(GSH-PX) assay kit (cat. no. A005), catalase (CAT) assay kit (cat.
no. A007-1-1) and malondialde-hyde (MDA) assay kit (cat. no.
A003-1) were purchased from Nanjing Jiancheng Bioengineering
Institute (Nanjing, China).
High-performance liquid chromatography to
determine rhein content
The content of rhein was determined using an L-2000
Liquid Chromatography system (Hitachi, Ltd., Tokyo, Japan) equipped
with an L-2200 auto-injection device. Chromatographic analysis was
preformed with a reverse-phase YMC-Pack Pro C18 column
(5 μm; 4.6×250 mm). The isocratic mobile phase comprised
0.5% aqueous acetic acid solution and methanol (40:60,
v/v) at a flow rate of 1.0 ml/min and detection was
performed at 435 nm using an L-2455 UV-VIS detector (Hitachi,
Ltd.). Total analysis time was 15 min and analysis was performed at
a temperature of 40°C. The injection volume of rhein was 10
μl. The results revealed that the exact purity of rhein was
98%, which was consistent with the result provided by the
manufacturer.
Animals
A total of 144 male specific pathogen free
Sprague-Dawley rats (weighing 260-300 g, aged 8-10 weeks old) were
obtained from the Laboratory Animal Center of Ningxia Medical
University (Yinchuan, China). All animal handling procedures were
approved by the Animal Research Ethics Committee, School of Ningxia
Medical University (Yinchuan, China). Animals were maintained in a
12 h light/dark cycle at room temperature (23±2°C) in 60% humidity.
The animals had ad libitum access to food and water. Rats
were randomly divided into 6 groups (each, n=24) as follows: Sham,
I/R, I/R + 12 mg/kg/day nimodipine (25,26), and I/R + 25, 50 or 100 mg/kg/day
rhein (27). Nimodipine was used
as a control, as it acts as a neuroprotectant. The sham and I/R
groups received 0.5% sodium carboxymethyl cellulose orally for 3
days following cerebral ischemic/reperfusion induced by MCAO, and
the other groups received nimodipine (12 mg/kg/day) and rhein (25,
50 or 100 mg/kg/day) orally for 3 days following cerebral
ischemic/reperfusion induced by MCAO.
MCAO model
As previously stated, MCAO induces ischemic injury
(28). Rats were anesthetized
with 7% chloral hydrate (350 mg/kg) via intraperitoneal injection.
The right common carotid artery (CCA), the external carotid artery
and the internal carotid artery (ICA) were exposed. A nylon wire
(1.8±0.5 cm) was advanced from the CCA into the ICA until it
blocked the original MCA. Reperfusion was performed by withdrawing
the nylon wire following 2 h ischemia. The sham-operated rats were
treated by the same surgical procedure however the nylon wire was
not introduced.
Determination of the NFS
NFSs were obtained from randomly selected mice from
each group (n=8) 72 h post reperfu-sion and they were evaluated
according to the Zea Longa's score criteria. The deficit criteria
used have been previously described (29). The scoring details were as
follows: 0, No noticeable neurological deficit; 1, failed to extend
opposite forepaw; 2, circled to the contralateral side; 3, tumbled
to its side while walking due to hemiplegia; 4, loss of
consciousness or mortality.
Determination of the area of cerebral
infarction
Rats were anesthetized by intraperitoneal injection
of 7% chloral hydrate (350 mg/kg). Rats were decapitated and then
the whole brain was removed. Subsequently, coronary brain slices
(2-mm sections) were stained using 2% TTC chloride for 30 min at
37°C and then fixed in 10% formalin solution at 25°C for 1 h (n=6).
The infarcted area was stained white, whereas normal brain tissue
was red. Stained cerebral slices were photographed using a Nikon
D7100 camera (Nikon Corporation, Tokyo, Japan). Cerebral infarct
areas were determined using microscope image-analysis software
(Image Pro plus, version 6.0; Media Cybernetics, Rockville, MD,
USA). The ratio of the infarcted area to the total brain area was
calculated using the following equation: Percentage of infarct
volume=infarct volume/(infarct volume + normal volume) ×100.
Hematoxylin and eosin staining
Prior to this procedure, rats were anesthetized by
intraperitoneal injection of 7% chloral hydrate (350 mg/kg) and
perfused transcardially with 100 ml of 0.9% sodium chloride,
followed by 200 ml 4% paraformaldehyde. Rats were decapitated and
then the whole brain was removed. Rat brains were fixed in 4%
formaldehyde at 25°C for 2 h. Brains were soaked in distilled water
for 4 h, dehydrated in increasing concentrations of alcohol,
hyalinized by dimethylbenzene, embedded in paraffin and sectioned
to a thickness of 4-μm. Sections were adhered to glass
slides prepared with poly-L-Lysine and stored at 4°C. Following
continual dewaxing and routine washing, paraffin-embedded sections
were stained with hematoxylin for 5 min, followed by color
separation using 1% hydrochloric acid alcohol for 20 sec. Sections
were then incubated with 1% ammonia for 30 sec, stained with eosin
for 5 min and subsequently dehydrated using alcohol, hyalinized
using dimethylbenzene and sealed with neutral gum. All the
aforementioned steps were performed at 25°C. A total of five
non-overlapping views of the cortex in each section were randomly
selected and observed under a light microscope for cell counting at
a magnification of ×400. The degenerated cell index (number of
degenerated cells/total cells) indicated the degree of damage.
Determination of oxidative stress
indicators
Levels of SOD, GSH-Px, CAT and MDA in the ischemic
brain were measured 72 h following I/R. The right cortical samples
(n=6 per group) were weighed. The SOD, GSH-Px and CAT activities
and MDA level were obtained using the T-SOD, GSH-Px, CAT and MDA
assay kits according to the manufacturer's protocol (Nanjing
Jiancheng Bioengineering Institute, Co., Ltd., Nanjing, China).
SOD, GSH-Px and CAT activities were expressed as units/mg protein.
The amount of lipid peroxide was obtained as the product of MDA.
MDA concentrations were expressed as nmol/mg protein.
Western blot analysis
Frozen right brains were homogenized in cold whole
cell lysis buffer (cat. no. KGP2100; Nanjing KeyGen Biotech Co.,
Ltd., Nanjing, China), sonicated twice on ice for 5 sec each time
and centrifuged at 12,000 × g for 5 min at 4°C to remove cell
debris. Total protein concentration in the supernatant was
determined using the BCA protein assay kit. A total of 80 μg
protein per lane was separated using 10% SDS-PAGE and transferred
to polyvinylidene difluoride membranes. Membranes were blocked with
5% non-fat dried milk for 1 h at 25°C and subsequently incubated
with primary antibodies against Bcl-2 (1:500), Bax (1:500),
caspase-9 (1:1,000), caspase-3 (1:1,000), cleaved caspase-3
(1:1,000), β-actin (1:1,000) and GAPDH (1:2,000) overnight at 4°C.
Membranes were washed with PBST and subsequently incubated with
horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G
secondary antibodies (1:1,000) for 2 h at room temperature.
Following washing, membranes were developed using a Chemidoc™ XRS
Imaging system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
signal intensities of the bands of interest were quantified and
normalized to β-actin or GAPDH using the Image-Pro Plus software
version 6.0 (Media Cybernetics, Inc.).
Reverse
transcription-quantitative-polymerase chain reaction analysis
(RT-qPCR)
RT-qPCR was used to assess the mRNA expression
levels of caspase-9 and caspase-3 72 h following reperfusion. Total
RNA was extracted using a total RNA kit according to the
manufacturer's protocol. Labeled cDNA was prepared using the
RevertAid First Strand cDNA Synthesis kit from the total RNA
samples. For PCR amplification, a CFX96 Real-Time system was used.
cDNA was added to each reaction to a final reaction volume of 25
μl containing 1X SYBR Premix Ex Taq II. The thermocycling
conditions used were as follows: Initial denaturation at 95°C for
30 sec., followed by 40 cycles comprising of denaturation at 95°C
for 5 sec, annealing at 60°C for 30 sec, elongation at 72°C for 30
sec and a final extension step at 72°C for 5 min. β-actin was used
as the reference gene for the normalization of different transcript
values and normalized mRNA levels were expressed as
2−ΔΔCq (ΔΔCq=Cqtarget-Cqβ-actin) (30). Only one produced PCR product was
subjected to melting curve analysis. Sequences of the primers used
are listed in Table I.
 | Table IThe primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I
The primers for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sequence
(5′-3′) |
|---|
| Caspase-3 | F:
AAAGGATGACTGGGAGTGG |
| R:
ATGACGACCTGGAACATCG |
| Caspase-9 | F:
TATGGCACAGATGGATGCTC |
| R:
CTTTCTGCTCACCACCACAG |
| β-actin | F:
CCCATCTATGAGGGTTACGC |
| R:
TTTAATGTCACGCACGATTTC |
Statistical analysis
Statistical analysis was conducted using SPSS 17.0
(SPSS, Inc., Chicago, IL, USA). The results were presented as the
mean ± standard deviation. Differences among >2 groups were
assessed using one-way analysis of variance followed by the least
significant difference post hoc test. Differences between two
groups were analyzed using an unpaired t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Rhein ameliorates NFSs
NFSs were obtained using the Zea Longa's criteria 72
h following cerebral reperfusion. Severe dysfunction on each test
was indicated by a high NFS score of 2-3. The NFSs of the I/R group
were significantly higher than those of the sham group (P<0.01;
Table II). However, the NFSs
were significantly decreased in the groups treated with 50 and 100
mg/kg rhein (P<0.05 and P<0.01, respectively) and nimodipine
(P<0.05) compared with the I/R group, thereby indicating that
rhein treatment improves neurological function impaired by cerebral
I/R.
| Table IIThe protective effects of rhein on
neurological functional score in rats 72 h post reperfusion. |
Table II
The protective effects of rhein on
neurological functional score in rats 72 h post reperfusion.
| Group | Neurological
functional score |
|---|
| Sham | 0.0±0.0 |
| I/R | 2.2±0.8a |
| Nimodipine | 1.0±0.7b |
| Rhein (25
mg/kg) | 2.0±0.9 |
| Rhein (50
mg/kg) | 1.6±0.5b |
| Rhein (100
mg/kg) | 1.1±0.9c |
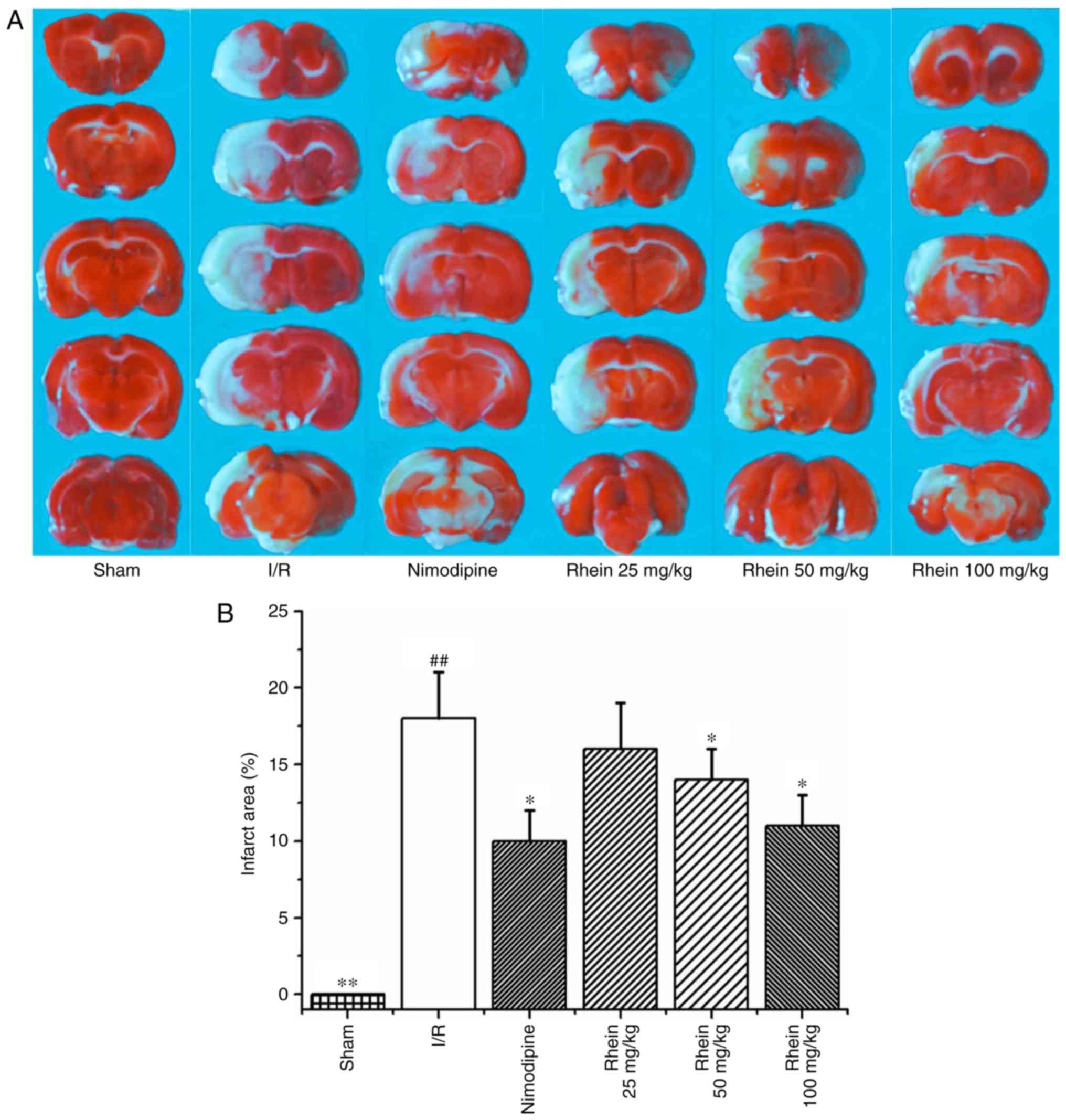
Rhein reduces the area of cerebral
infarction
Infarcted cerebral volume was measured 72 h
following cerebral reperfusion by TTC staining. The infarcted area
(white) was primarily located at the striatum and parts of the
cortex frontoparietal lobe in the right hemisphere (Fig. 1A). The infarcted cerebral area of
the I/R group was significantly greater than that of the sham group
(P<0.01). By contrast, theinfarcted cerebral area of the
nimodipine and 50 and 100 mg/kg rheingroups were significantly
smaller than the I/R group (each P<0.05; Fig. 1B), thereby indicating that
treatment with rhein reduces cerebral injury induced by cerebral
I/R.
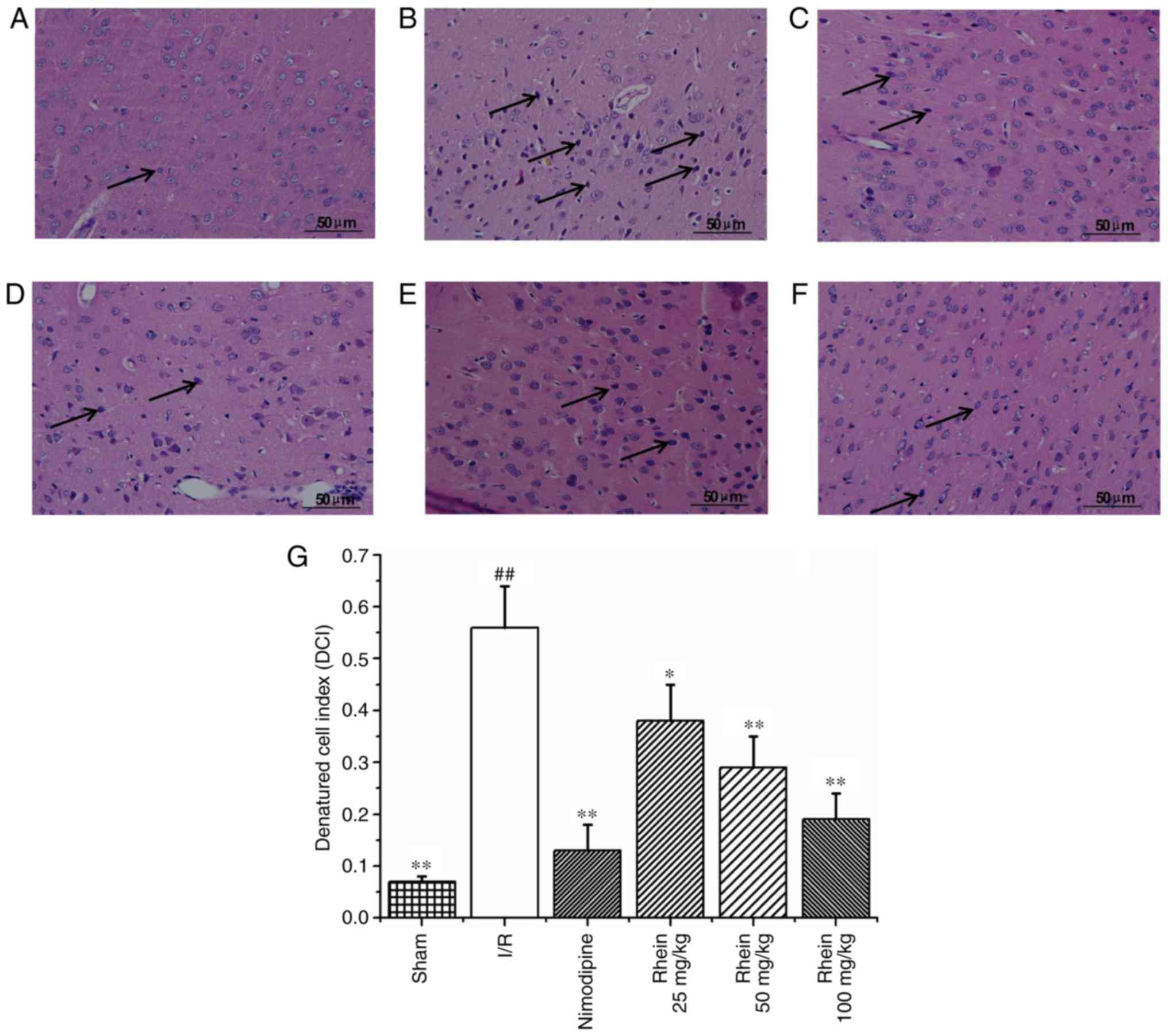
Histological changes
Hematoxylin and eosin staining (Fig. 2) revealed that nerve cell
arrangement was well organized, exhibiting a complete structure in
the control group (Fig. 2A).
However, 72 h following ischemia, in the I/R group the neuron
arrangement became irregular with deeply colored and condensed
nuclei, increased gaps around the nerves and degenerative changes,
including the formation of vacuoles and necrosis in a number of the
neurons (Fig. 2B). In addition,
the denatured cell index (DCI) values in the I/R group were
significantly higher than those of the sham group (P<0.01;
Fig. 2G). In the nimodipine and
rhein groups, nerve cell damage was milder compared with that of
the model group and neurons were well arranged, with clear
outlines, nuclear condensation, and shriveled bodies. DCI values in
all the rhein groups and the nimodipine group were significantly
decreased compared with the I/R group (P<0.05 or P<0.01;
Fig. 2G). Deep coloration was
observed in only a few cells and there were no significant
differences in DCI values between the different rhein groups.
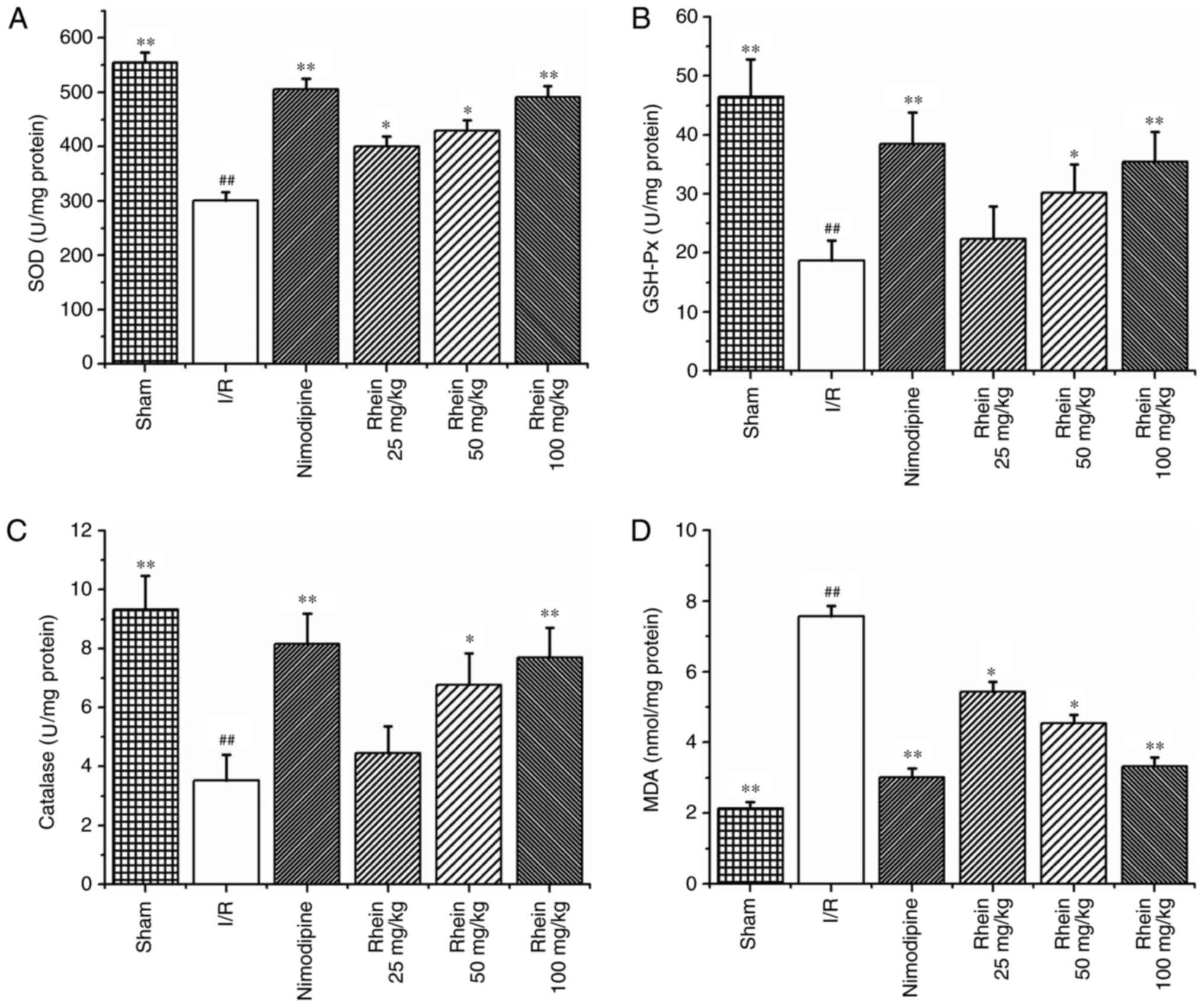
Effect of rhein on oxidative stress
indicators induced by I/R
The activities of the SOD, GSH-Px and CAT enzymes
were significantly reduced in the I/R group compared with those in
the sham group (P<0.01; Fig.
3A–C). However, treatment with nimodipine and 50 and 100 mg/kg
rhein, significantly increased the activities of SOD, GSH-Px and
CAT compared with the I/R group (P<0.01 or P<0.05). Treatment
with 25 mg/kg rhein also significantly increased the activity of
SOD compared with the I/R group (P<0.05). MDA was used as a
biomarker for lipid peroxidation and oxidative stress. MDA content
was significantly higher in the ischemic cortices of the I/R group
compared with the sham group (P<0.01; Fig. 3D). However, following treatment
with nimodipine and all doses of rhein, MDA content was
significantly decreased compared with the I/R group (P<0.01 or
P<0.05).
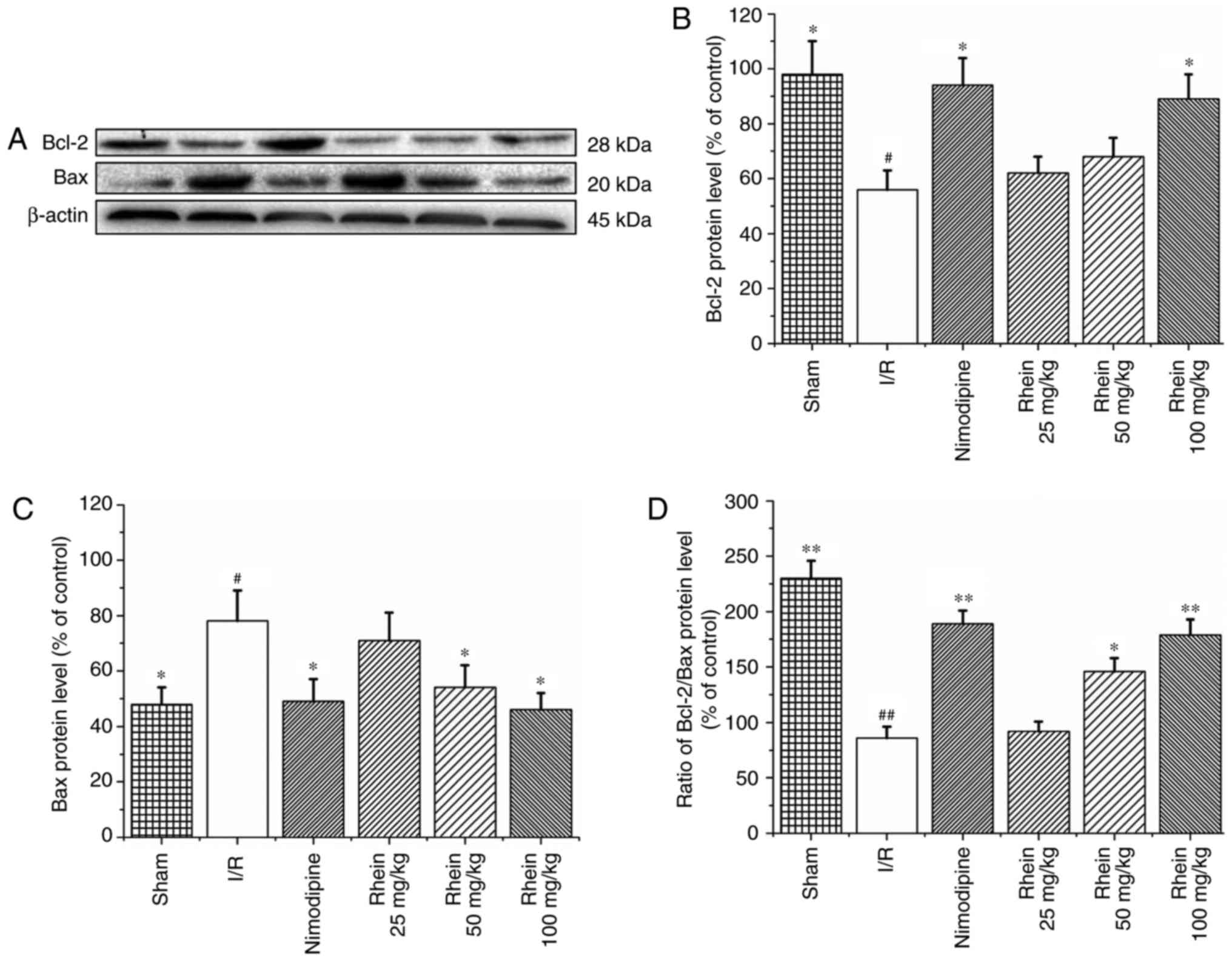
Rhein inhibits BAX expression and
enhances Bcl-2 expression
Western blot analysis revealed that the expression
of Bcl-2 was significantly reduced (P<0.05; Fig. 4A and C) and that BAX protein
expression was significantly enhanced in the I/R group compared
with the sham group (P<0.05; Fig.
4A and D). Compared with the I/R group, treatment with
nimodipine and 100 mg/kg rhein significantly upregulated Bcl-2
expression (P<0.05; Fig. 4A and
C). Furthermore, treatment with nimodipine and 50 and 100 mg/kg
rhein significantly downregulated BAX expression (P<0.05;
Fig. 4A and D). The Bcl-2/BAX
ratio was also significantly increased following treatment with
nimodipine and 50 or 100 mg/kg rhein compared with the I/R group
(P<0.05 or P<0.01; Fig.
4B).
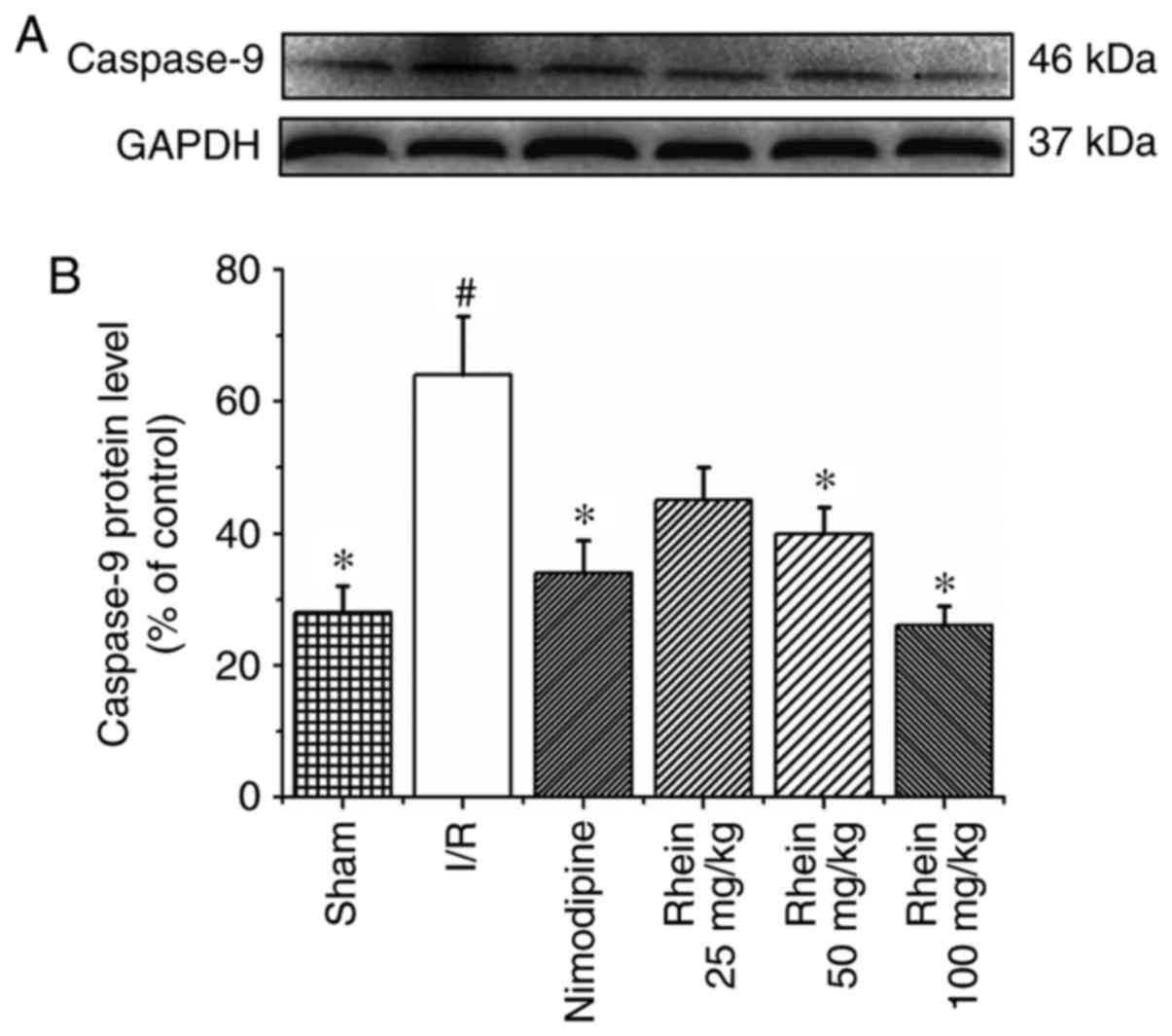
Rhein inhibits the expression of
caspase-9, caspase-3 and cleaved caspase-3
Western blot analysis revealed that the expression
of caspase-9 in the I/R group was significantly higher than in the
sham group (P<0.05; Fig. 5).
Treatment with 50 and 100 mg/kg rhein significantly reduced the
expression of caspase-9 compared with the I/R group (P<0.05). In
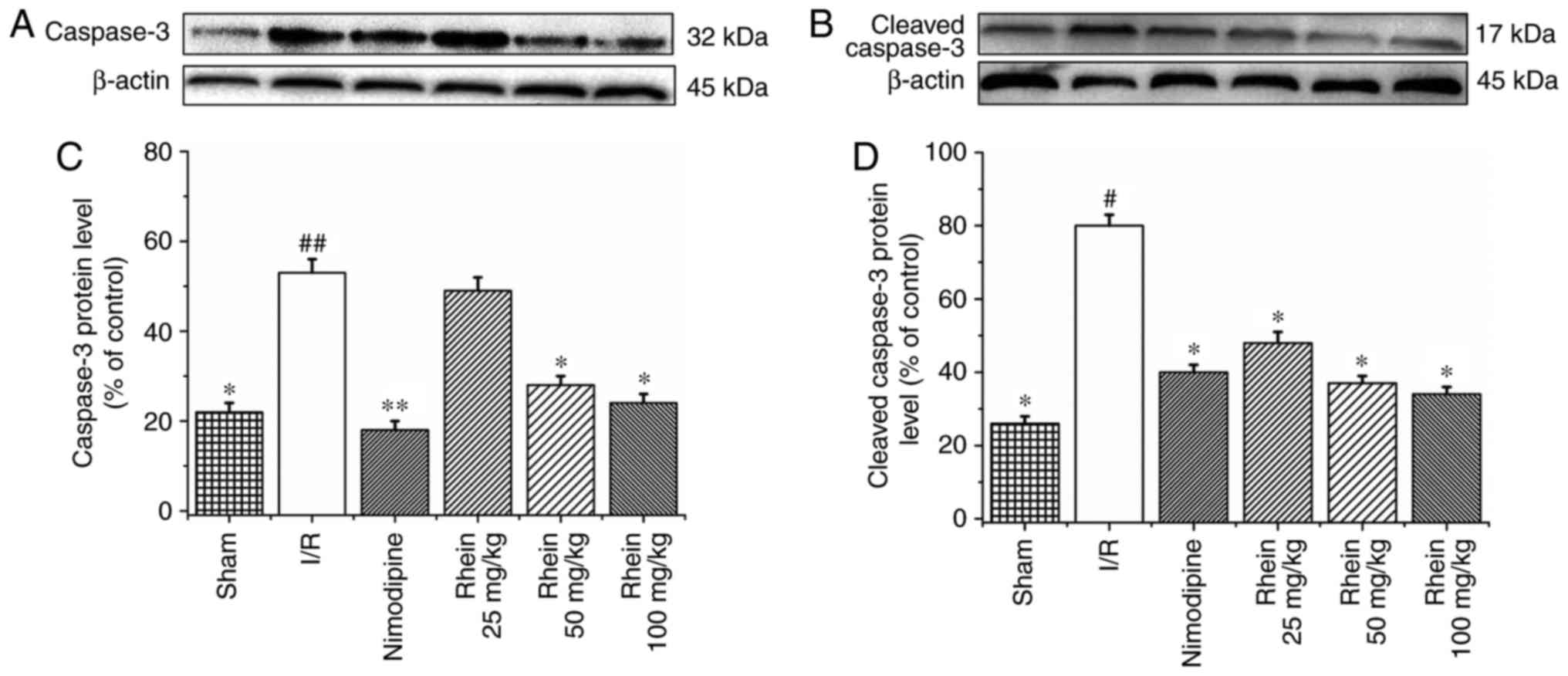
the I/R group, levels of caspase-3 (P<0.01) and cleaved
caspase-3 (P<0.05) expression were significantly higher than in
the sham group (Fig. 6). The
administration of nimodipine and 50 and 100 mg/kg rhein
significantly reduced the expression of caspase-3 (P<0.05),
whereas administration of nimodipine and all doses of rhein
significantly reduced the expression of cleaved caspase-3
(P<0.05).
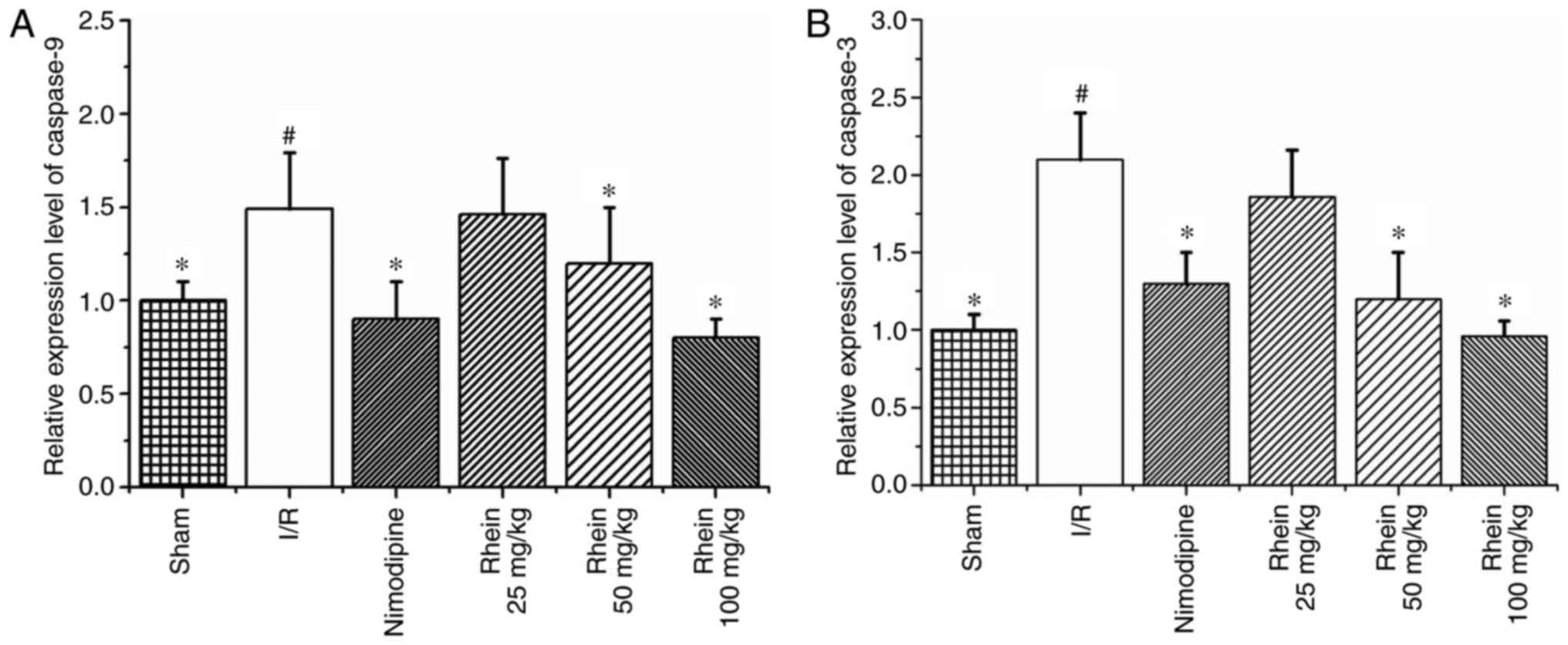
RT-qPCR results revealed that levels of caspases-9
and -3 mRNA expression were significantly increased in the I/R
group compared with the sham group (P<0.05; Fig. 7). However, treatment with 50 and
100 mg/kg rhein significantly decreased levels of caspases-9 and -3
mRNA (P<0.05).
Discussion
In the present study, the infarcted cerebral area
was identified by TTC staining and NFS was evaluated 72 h following
reper-fusion. Treatment with rhein markedly decreased the infarcted
cerebral area and significantly improved the NFS. These results
indicate that rhein may protect against cerebral I/R injury.
Ischemic stroke is the cumulative effect of numerous
mechanisms including oxidative stress, excitotoxicity,
intracellular calcium overload, inflammation and apoptosis
(31). Oxidative stress secondary
to ROS formation and neuron apoptosis is crucial in stimulating the
progression of ischemic stroke (32). Under normal physiological
conditions, low levels of ROS exist within the brain (6). Excessive ROS are removed by
endogenous antioxidant enzymes including SODs (33), GSH-Px and CATs (34). SOD converts superoxide anions into
hydrogen peroxide and GSH-Px uses GSH to reduce
H2O2 to water (35). MDA, which is a poisonous end
product of lipid peroxidation, is able to directly reflect the rate
and extent of lipid peroxidation and indirectly reflects the
capacity of free radical elimination (36). Previous studies have demonstrated
that increasing the SOD, CAT and GSH-Px levels and decreasing MDA
concentrations significantly protects against oxidative stress
during ischemic stroke (7,9-11).
The results of the present study were consistent with the
aforementioned previous studies. The protective effects of rhein in
I/R injury may be associated with its enhancing effect on
endogenous antioxidant capability, which mitigates oxidative stress
during cerebral I/R. Therefore, the enhancement of the antioxidant
system represents a potential target for novel therapies to treat
patients with ischemic stroke.
Ischemic stroke results in ROS overproduction in the
mitochondria (37) and specific
ROS, including H2O2 or superoxide, are
crucial mediators of apoptosis (38,39). Apoptosis is induced via extrinsic
and intrinsic pathways (40). In
the intrinsic pathway, the loss of transmembrane potential leads to
mitochondrial respiration failure and enhanced ROS generation,
which accelerates cytochrome c release from the mitochondria
(41). The release of cytochrome
c via this mechanism may be associated with the Bcl-2 family
of proteins (40,42). When cytochrome c forms a
complex with apoptotic protease-activating factor 1 apoptosomes are
formed, which activate caspase-9 and consequently activate
caspase-3 (43), DNA-breaking
enzymes, including endonucleases (44), and repair enzymes, including poly
ADP-ribose polymerase (45),
leading to cell death. The extrinsic apoptosis pathway activates
caspase-8, which either activates executioner caspases, such as
caspase-3, or the intrinsic apoptotic pathway (46,47).
Apoptosis is caused by an imbalance between
pro-apoptotic and anti-apoptotic signals. Bcl-2 is an
anti-apoptotic protein that intercepts the release of cytochrome
c in response to various apoptotic signals, thereby
inhibiting apoptosis. BAX is a pro-apoptotic protein that
accelerates cytochrome c release from the mitochondria,
thereby leading to cell death (48-51). It has been demonstrated that the
Bcl-2/BAX ratio is decreased in the cerebrum of rats following I/R
(52,53). The results of the present study
illustrated that the expression of Bcl-2 was significantly
inhibited whereas the expression of BAX was significantly increased
following I/R, which is consistent with the results of previous
studies. The Bcl-2/BAX ratio was significantly reduced in the
cerebrum of rats following I/R; however the administration of 100
mg/kg rhein significantly reversed the decrease in Bcl-2
expression. In addition, 50 and 100 mg/kg rhein significantly
inhibited the increase in BAX expression. The ratio of Bcl-2/BAX
was significantly increased following the administration of 50 and
100 mg/kg rhein. The administration of 100 mg/kg rhein was more
effective than 50 mg/kg rhein. The results indicate that rhein may
prevent cerebral I/R injury by upregulating Bcl-2 and
downregulating BAX expression, thereby increasing the Bcl-2/BAX
ratio and inhibiting apoptosis.
Caspases are central regulators of apoptosis and
serve keys roles in ischemia-induced cytotoxicity (54). Caspases may be classified into two
groups, initiators (caspase-9) and effectors (caspase-3), based on
their function (55). Typically,
initiator caspases activate downstream effector caspases through a
proteolytic cascade, resulting in the cleavage of cellular
substrates involved in apoptosis (56). Caspase-3 may be activated by
caspase-9 via the mitochondria-dependent cytochrome
c/caspase-9 intrinsic pathway (57). In addition, increased BAX
expression may induce the activation of caspase-9 within the
mitochondria (58). Caspases-9
and -3 are upregulated and activated in ischemic brain tissues
(59) and pharmacological or
genetic inhibition of caspase-3 reduces neuronal death in the
ischemic brain (60). The present
study demonstrated that oral rhein administration maintains low
caspase-9 levels. Rhein at doses of 50 and 100 mg/kg significantly
inhibited caspase-9 expression. Caspase-3 expression in the I/R
group was significantly higher than in the sham group; however,
administration with 100 mg/kg rhein significantly inhibited its
expression. Cleaved caspase-3 is a key executioner of apoptosis.
The results of the present study revealed that cleaved caspase-3
expression in the I/R group was significantly higher than that of
the sham group, which was consistent with the results obtained by
Wang et al (61). However,
administration of 50 and 100 mg/kg rhein significantly inhibited
the expression of cleaved caspase-3.
In conclusion, the present study demonstrated that
treatment with rhein markedly reduces the area of brain infarction
and reduces neuronal apoptosis in a rat model of I/R injury. The
protective effects of rhein were mediated through the augmentation
of endogenous antioxidant defenses and the inhibition of the
oxidative stress pathway in the ischemic rat brain. In addition,
rhein exhibited anti-apoptotic effects. These results suggest that
rhein may be beneficial in the treatment of ischemic stroke.
However, further studies are required to confirm these results
prior to its use in humans.
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81660700 and
81260679), Ningxia College first-class discipline construction
project (Chinese medicine) funded project (grant no.
NXYLXK2017A06), the college students innovation training program of
Ningxia Hui Autonomous Region (grant nos. NXCX2015183 and
NXCX2016162).
[2] Availability
of data and materials
The analyzed data sets generated during the study
are available from the corresponding author upon reasonable
request.
[3] Authors'
contributions
QZ, XW and XC constructed the MCAO model, performed
hematoxylin and eosin staining, western blot analysis and reverse
transcription-quantitative polymerase chain reaction. AC and GZ
determined the NFS, the area of cerebral infarction and oxidative
stress indicators. YZ and YH analyzed and interpreted the data. JS
revised the manuscript critically for important intellectual
content. YZ and QZ designed the study, supervised the research
group and gave final approval of the version to be published. The
final version of the manuscript has been read and approved by all
authors and each author believes that the manuscript represents
honest work.
[4] Ethics
approval and consent to participate
All procedures were approved by the Animal Research
Ethics Committee, School of Ningxia Medical University (Yinchuan,
China).
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Writing Group Members; Mozaffarian D,
Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de
Ferranti S, Després JP, et al: Heart disease and stroke
statistics-2016 update: A report from the American heart
association. Circulation. 133:e338–e360. 2016. View Article : Google Scholar
|
|
2
|
Hua F, Tang H, Wang J, Prunty MC, Hua XD,
Sayeed L and Stein DG: TAK-242, an antagonist for Toll-like
receptor 4, protects against acute cerebral ischemia/reperfusion
injury in mice. J Cereb Blood Flow Metab. 35:536–542. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El Amki M and Wegener S: Improving
cerebral blood flow after arterial recanalization: A novel
therapeutic strategy in stroke. Int J Mol Sci. 18:E26692017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang X, Chen L, Dang X, Liu J, Ito Y and
Sun W: Neuroprotective effects of total steroid saponins on
cerebral ischemia injuries in an animal model of focal
ischemia/reperfusion. Planta Med. 80:637–644. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Melani A, Dettori I, Corti F, Cellai L and
Pedata F: Time-course of protection by the selective A2A
receptor antagonist SCH58261 after transient focal cerebral
ischemia. Neurol Sci. 36:1441–1448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang R, Xu M, Wang Y, Xie F, Zhang G and
Qin XY: Nrf2-a promising therapeutic target for defensing against
oxidative stress in stroke. Mol Neurobiol. 54:6006–6017. 2017.
View Article : Google Scholar
|
|
7
|
Žitňanová I, Šiarnik P, Kollár B, Chomová
M, Pazderová P, Andrezálová L, JeDoviIová M, Koňariková K,
Laubertová L, Krivošíková Z, et al: Oxidative stress markers and
their dynamic changes in patients after acute ischemic stroke. Oxid
Med Cell Longev. 2016:97616972016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Adibhatla RM and Hatcher JF: Altered lipid
metabolism in brain injury and disorders. Subcell Biochem.
49:241–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guo J, Cheng C, Chen CS, Xing X, Xu G,
Feng J and Qin X: Overexpression of Fibulin-5 attenuates
ischemia/reperfu-sion injury after middle cerebral artery occlusion
in rats. Mol Neurobiol. 53:3154–3167. 2016. View Article : Google Scholar
|
|
10
|
Ashabi G, Khalaj L, Khodagholi F,
Goudarzvand M and Sarkaki A: Pre-treatment with metformin activates
Nrf2 antioxidant pathways and inhibits inflammatory responses
through induction of AMPK after transient global cerebral ischemia.
Metab Brain Dis. 30:747–754. 2015. View Article : Google Scholar
|
|
11
|
Ding Y, Chen M, Wang M, Li Y and Wen A:
Posttreatment with 11-keto-β-Boswellic acid ameliorates cerebral
ischemia reperfusion injury: Nrf2/HO-1 pathway as a potential
mechanism. Mol Neurobiol. 52:1430–1439. 2015. View Article : Google Scholar
|
|
12
|
Milanlioglu A, Aslan M, Ozkol H, Çilingir
V, Nuri Aydın M and Karadas S: Serum antioxidant enzymes activities
and oxidative stress levels in patientswith acute ischemic stroke:
Influence on neurological status and outcome. Wien Klin Wochenschr.
128:169–174. 2016. View Article : Google Scholar
|
|
13
|
Zhao J, Yu S, Zheng W, Feng G, Luo G, Wang
L and Zhao Y: Curcumin improves outcomes and attenuates focal
cerebral ischemic injury via antiapoptotic mechanisms in rats.
Neurochem Res. 35:374–379. 2010. View Article : Google Scholar
|
|
14
|
Chen S, Peng H, Rowat A, Gao F, Zhang Z,
Wang P, Zhang W, Wang X and Qu L: The effect of concentration and
duration of normobaric oxygen in reducing caspase-3 and -9
expression in a rat-model of focal cerebral ischaemia. Brain Res.
1618:205–211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wagner DC, Riegelsberger UM, Michalk S,
Härtig W, Kranz A and Boltze J: Cleaved caspase-3 expression after
experimental stroke exhibits different phenotypes and is
predominantly non-apoptotic. Brain Res. 1381:237–242. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Endres M, Namura S, Shimizu-Sasamata M,
Waeber C, Zhang L, Gómez-Isla T, Hyman BT and Moskowitz MA:
Attenuation of delayed neuronal death after mild focal ischemia in
mice by inhibition of the caspase family. J Cereb Blood Flow Metab.
18:238–247. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma J, Endres M and Moskowitz MA:
Synergistic effects of caspase inhibitors and MK-801 in brain
injury after transient focal cerebral ischaemia in mice. Br J
Pharmacol. 124:756–762. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sugawara T, Lewén A, Gasche Y, Yu FS and
Chan P: Overexpression of SOD1 protects vulnerable motor neurons
after spinal cord injury by attenuating mitochondrial cytochrome c
release. FASEB J. 16:1997–1999. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsang SW and Bian ZX: Anti-fibrotic and
Anti-tumorigenic effects of rhein, a natural anthraquinone
derivative, in mammalian stellate and carcinoma cells. Phytother
Res. 29:407–414. 2015. View
Article : Google Scholar
|
|
20
|
Liu J, Uematsu H, Tsuchida N and Ikeda MA:
Essential role of caspase-8 in p53/p73-dependent apoptosis induced
by etoposide in head and neck carcinoma cells. Mol Cancer.
10:952011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cong XD, Ding MJ, Dai DZ, Wu Y, Zhang Y
and Dai Y: ER stress, p66shc, and p-Akt/Akt mediate
adjuvant-induced inflammation, which is blunted by argirein, a
supermolecule and rhein in rats. Inflammation. 35:1031–1040. 2012.
View Article : Google Scholar
|
|
22
|
Gao Y and Chen X: Rhein exerts pro-and
anti-inflammatory actions by targeting IKKβ inhibition in
LPS-activated macrophages. Free Radic Biol Med. 72:104–112. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Fan R, Luo J, Tang T, Xing Z, Xia
Z, Peng W, Wang W, Lv H, Huang W, et al: An ultra high performance
liquid chromatography with tandem mass spectrometry method for
plasma and cerebrospinal fluid pharmacokinetics of rhein in
patients with traumatic brain injury after administration of
rhubarb decoction. J Sep Sci. 38:1100–1108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lam BY, Lo AC, Sun X, Luo HW, Chung SK and
Sucher NJ: Neuroprotective effects of tanshinones in transient
focal cerebral ischemia in mice. Phytomedicine. 10:286–291. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li H, Deng CQ, Chen BY, Zhang SP, Liang Y
and Luo XG: Total saponins of Panax Notoginseng modulate the
expression of caspases and attenuate apoptosis in rats following
focal cerebral ischemia-reperfusion. J Ethnopharmacol. 121:412–418.
2009. View Article : Google Scholar
|
|
26
|
Zhao Q, Cheng X, Wang X, Wang J, Zhu Y and
Ma X: Neuroprotective effect and mechanism of Mu-Xiang-You-Fang on
cerebral ischemia-reperfusion injury in rats. J Ethnopharmacol.
192:140–147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lian Y, Xie L, Chen M and Chen L: Effects
of an astragalus polysaccharide and rhein combination on apoptosis
in rats with chronic renal failure. Evid Based Complement Alternat
Med. 2014:2718622014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Genovese T, Mazzon E, Paterniti I,
Esposito E, Bramanti P and Cuzzocrea S: Modulation of NADPH oxidase
activation in cerebral ischemia/reperfusion injury in rats. Brain
Res. 1372:92–102. 2011. View Article : Google Scholar
|
|
29
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Kaushal V and Schlichter LC: Mechanisms of
microglia-mediated neurotoxicity in a new model of the stroke
penumbra. J Neurosci. 28:2221–2230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Browning JD and Horton JD: Molecular
mediators of hepatic steatosis and liver injury. J Clin Invest.
114:147–152. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rosen DR, Siddique T, Patterson D,
Figlewicz DA, Sapp P, Hentati A, Donaldson D, Goto J, O'Regan JP,
Deng HX, et al: Mutations in Cu/Zn superoxide dismutase gene are
associated with familial amyotrophic lateral sclerosis. Nature.
362:59–62. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vakili A, Sharifat S, Akhavan MM and
Bandegi AR: Effect of lavender oil (Lavandula angustifolia) on
cerebral edema and its possible mechanisms in an experimental model
of stroke. Brain Res. 1548:56–62. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rodrigo R, Fernández-Gajardo R, Gutiérrez
R, Matamala JM, Carrasco R, Miranda-Merchak A and Feuerhake W:
Oxidative stress and pathophysiology of ischemic stroke: Novel
therapeutic opportunities. CNS Neurol Disord Drug Targets.
12:698–714. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schettler V, Methe H, Staschinsky D,
Schuff-Werner P, Müller GA and Wieland E: Review: The
oxidant/antioxidant balance during regular low density lipoprotein
apheresis. Ther Apher. 3:219–226. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fulda S, Galluzzi L and Kroemer G:
Targeting mitochondria for cancer therapy. Nat Rev Drug Discov.
9:447–464. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Carmona-Gutierrez D, Eisenberg T, Büttner
S, Meisinger C, Kroemer G and Madeo F: Apoptosis in yeast:
Triggers, pathways, subroutines. Cell Death Differ. 17:763–773.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fuchs Y and Steller H: Programmed cell
death in animal development and disease. Cell. 147:742–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhen YF, Wang GD, Zhu LQ, Tan SP, Zhang
FY, Zhou XZ and Wang XD: P53 dependent mitochondrial permeability
transition pore opening is required for dexamethasone-induced death
of osteoblasts. J Cell Physiol. 229:1475–1483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Matés JM, Segura JA, Alonso FJ and Márquez
J: Intracellular redox status and oxidative stress: Implications
for cell proliferation, apoptosis, and carcinogenesis. Arch
Toxicol. 82:273–299. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sugawara T, Noshita N, Lewén A, Gasche Y,
Ferrand-Drake M, Fujimura M, Morita-Fujimura Y and Chan PH:
Overexpression of copper/zinc superoxide dismutase in transgenic
rats protects vulnerable neurons against ischemic damage by
blocking the mitochondrial pathway of caspase activation. J
Neurosci. 22:209–217. 2002.PubMed/NCBI
|
|
44
|
Fahmi T, Branch D, Nima ZA, Jang DS,
Savenka AV, Biris AS and Basnakian AG: Mechanism of
graphene-induced cytotoxicity: Role of endonucleases. J Appl
Toxicol. 37:1325–1332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhou X, Patel D, Sen S, Shanmugam V,
Sidawy A, Mishra L and Nguyen BN: Poly-ADP-ribose polymerase
inhibition enhances ischemic and diabetic wound healing by
promoting angiogenesis. J Vasc Surg. 65:1161–1169. 2017. View Article : Google Scholar
|
|
46
|
Broughton BR, Reutens DC and Sobey CG:
Apoptotic mechanisms after cerebral ischemia. Stroke. 40:e331–e339.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rami A, Bechmann I and Stehle JH:
Exploiting endogenous anti-apoptotic proteins for novel therapeutic
strategies in cerebral ischemia. Prog Neurobiol. 85:273–296. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Budihardjo I, Oliver H, Lutter M, Luo X
and Wang X: Biochemical pathways of caspase activation during
apoptosis. Annu Rev Cell Dev Biol. 15:269–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang X: The expanding role of mitochondria
in apoptosis. Genes Dev. 15:2922–2933. 2001.PubMed/NCBI
|
|
50
|
Rossé T, Olivier R, Monney L, Rager M,
Conus S, Fellay I, Jansen B and Borner C: Bcl-2 prolongs cell
survival after BAX-induced release of cytochrome c. Nature.
391:496–499. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
51
|
Van Delft MF and Huang DC: How the Bcl-2
family of proteins interact to regulate apoptosis. Cell Res.
16:203–213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang GH, Lan R, Zhen XD, Zhang W, Xiang J
and Cai DF: An-Gong-Niu-Huang Wan protects against cerebral
ischemia induced apoptosis in rats: Up-regulation of Bcl-2 and
down-regulation of BAX and caspase-3. J Ethnopharmacol.
154:156–162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhao P, Zhou R, Zhu XY, Hao YJ, Li N, Wang
J, Niu Y, Sun T, Li YX and Yu JQ: Matrine attenuates focal cerebral
ischemic injury by improving antioxidant activity and inhibiting
apoptosis in mice. Int J Mol Med. 36:633–644. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lee SR, Lok J, Rosell A, Kim HY, Murata Y,
Atochin D, Huang PL, Wang X, Ayata C, Moskowitz MA and Lo EH:
Reduction of hippocampal cell death and proteolytic responses in
tissue plasminogen activator knockout mice after transient global
cerebral ischemia. Neuroscience. 150:50–57. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pérez-Garijo A: When dying is not the end:
Apoptotic caspases as drivers of proliferation. Semin Cell Dev
Biol. S1084–9521(17): 30500–1. 2017.
|
|
56
|
Liu J, Chen Z, Zhang Y, Zhang M, Zhu X,
Fan Y, Shi S, Zen K and Liu Z: Rhein protects pancreatic β-cells
from dynamin-related protein-1-mediated mitochondrial fission and
cell apoptosis under hyperglycemia. Diabetes. 62:3927–3935. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu H, Zhou Y and Tang L: Caffeine induces
sustained apoptosis of human gastric cancer cells by activating the
caspase9/caspase3 signalling pathway. Mol Med Rep. 16:2445–2454.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Topçu Y, Bayram E, Ozbal S, Yiş U, Tuğyan
K, Karaoğlu P, Kumral A, Yılmaz O and Kurul SH: Zonisamide
attenuates hyperoxia-induced apoptosis in the developing rat brain.
Neurol Sci. 35:1769–1775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rami A, Sims J, Botez G and Winckler J:
Spatial resolution of phospholipid scramblase 1 (PLSCR1), caspase-3
activation and DNA-fragmentation in the human hippocampus after
cerebral ischemia. Neurochem Int. 43:79–87. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Le DA, Wu Y, Huang Z, Matsushita K,
Plesnila N, Augustinack JC, Hyman BT, Yuan J, Kuida K, Flavell RA
and Moskowitz MA: Caspase activation and neuroprotection in
caspase-3 deficient mice after in vivo cerebral ischemia and in
vitro oxygen glucose deprivation. Proc Natl Acad Sci USA.
99:15188–15193. 2002. View Article : Google Scholar
|
|
61
|
Wang N, Zhang Y, Wu L, Wang Y, Cao Y, He
L, Li X and Zhao J: Puerarin protected the brain from cerebral
ischemia injury via astrocyte apoptosis inhibition.
Neuropharmacology. 79:282–289. 2014. View Article : Google Scholar
|