Introduction
Gestational diabetes mellitus (GDM) is defined as
abnormal or impaired glucose metabolism before pregnancy and it
initially appears during pregnancy. GDM may lead to early embryo
abnormalities or even death, pregnancy-induced hypertension
syndrome, infection, polyhydramnios and premature labor. It will
also affect the status of the fetus, including effects such as
fetal malformation, macrosomia or stillbirth. The offspring of
woman with GDM are more inclined to develop low blood sugar,
respiratory distress syndrome and polycythemia. Furthermore, these
offspring have a higher risk of developing obesity and impaired
glucose tolerance and the mother with GDM is more likely to develop
diabetes [mainly type 2 diabetes mellitus (DM)] later in life.
In recent years, the incidence of GDM has increased
significantly. In China, the incidence of GDM has increased
significantly and women with GDM account for 3-7% of pregnancies
(1). The pathophysiological basis
of GDM world-wide includes increased insulin resistance and
decreased secretion of β-cell during pregnancy (2,3).
Sex hormone-binding globulin (SHBG) is a glycoprotein produced in
the liver, specifically to bind and transport sex hormones, and
regulate the biological activity and the concentration of sex
hormone in the blood (4).
Previous studies have shown that the level of SHBG was decreased in
women with insulin tolerance and GDM (5). A low SHBG level is an independent
risk factor for type 2 DM, which has been suggested as a predictor
for hyperinsulinemia and insulin tolerance (6). Higher SHBG concentrations can
effectively prevent impaired fasting glucose, the occurrence of
type 2 DM and hyperinsulinemia (7).
Placenta is a combination of tissues of embryo and
mother, carrying the exchange of substances. It consists of
amniotic membrane, chorion frondosum and deciduas. Placenta is
evolved from the fertilized egg and has the same genetic material
with the fetus. Our prior experiments on the changes of SHBG in GDM
placental tissue have been confirmed that SHGB can be synthesized
and secreted by the placenta trophoblast cells, SHBG mRNA and SHBG
protein concentration were significantly decreased in the GDM group
(P<0.01), and that of SHBG played an important role in the
occurrence and development of GDM (8-10).
The single-nucleotide polymorphism, rs6259, is a
functioning gene mutation within a coding region of the SHBG gene.
Nucle otide 5790 within exon 8 of the SHBG gene has a missense
mutation (GAC→AAC), and the carboxyl-terminal globular laminin
domain created a new N-connecting carbohydrate chain, which could
provide for an additional N-glycosylation site (11,12). Glycosylation does not affect the
binding of steroids to SHBG, but it may decrease the plasma
clearance of SHBG and prolong its half-life (13,14). This substitution may lead to
increased SHBG levels, changes in gene polymorphism (TAAAA)n and
has influence on transcription of the non-coding region (11). Studies have confirmed that the
increased SHBG levels in hirsutism (11) and polycystic ovary syndrome
(3) are associated with the
variant Asn327 allele. Ding et al (15) found that Asn327 allele
carriers have higher SHBG levels in plasma (10%) than
Asp327 carriers (P=0.005). This single nucleotide
polymorphism (SNP) rs6259 has different frequencies in ethnic
groups around the world (13). In
healthy Caucasians it was 7.5-12%, in African Americans 2%, and the
highest frequency was found in Chinese women (16-21).
The present study focused on the distribution of the
human SHBG SNP rs6259, its correlation with placental SHBG levels
and the occurrence and development of GDM, to predict the risk of
diabetes in the next generation.
Patients and methods
Patients and sample collection
All subjects were from Shengjing Hospital, which is
affiliated with China Medical University. This study was approved
by the Ethics Committee of Shengjing Hospital, and all participants
provided informed consent.
In total, 180 women with singleton pregnancy,
positive oral glucose tolerance test (OGTT), and regular prenatal
examinations were selected into the study. The normal group was
comprised of 210 cases that were OGTT negative, healthy pregnant
women with singleton pregnancy over the same period. Pregnant women
with a complication, such as gestational hypertension and
pregestational DM were excluded. The GDM group was selected from
the outpatients whose glucose was well-controlled and who received
systematic examinations during the pregnancy from Nove mber 2012 to
September 2014. According to the American Diabetes Association
(22), the recommended GDM
diagnosis is: i) fasting plasma glucose over 5.1 mmol/l during
pregnancy; and ii) blood glucose levels at 1 and 2 h after taking
75-g oral glucose over 10.0 and 8.5 mmol/l during gestational weeks
24 to 28. Either one beyond the diagnostic boundary can be
identified as GDM.
The placental tissues of the two groups were
collected from each patient immediately after delivery. The tissue
was cut into ~0.5 cm3 pieces from placenta on the side
of the mother, central and marginal areas. Cold normal saline was
used to wash the pieces before they were dissected into small
sections, which were blotted dry on filter paper and snap-frozen in
liquid nitrogen for 4 h, before being stored at −80°C.
Clinical index collection
We had statistics on general information in the
selected 390 cases, including in pregnant women the age,
gestational weeks, birth weight, childbearing history and fasting
blood glucose, and the differences were compared between the two
groups.
DNA sample preparation
Genomic DNA was extracted from placental samples by
a blood/cell/tissue genomic DNA extraction kit (Tiangen Biotech
Co., Ltd., Beijing, China) strictly according to the manufacturer's
instructions. The test result determination must take the
microtiter plate reader as a standard.
Polymerase chain reaction-restriction
fragment length polymorphism (PCR-RFLP) and agarose gel
electrophoresis
PCR primer sequences were as follows: forward
primer, 5′-TTCTGGATCCGAGCCACCT-3′ and reverse primer,
5′-AGTGCCTGGTACATTGCTAG-3′ (Invitrogen Life Technologies, Carlsbad,
CA, USA). Each PCR reaction contained, DNA template 2 μl,
Takara Ex Taq (5 U/μl) 0.25 μl, dNTP mixture (2.5 mM)
4 μl, 1 μl (10 μM) each of SHBG forward and
reverse primers, 10X Ex Taq buffer (Mg2+ Plus) 5
μl, with ultrapure water added to 50 μl total volume
(Takara Bio, Inc., Otsu, Japan). The PCR conditions: initial
denaturation at 94°C for 5 min. PCR profiles consisted of 35 cycles
with denaturation at 94°C for 10 sec, annealing at 55°C for 30 sec,
and extension at 72°C for 60 sec, followed by a final extension at
72°C for 7 min. PCR products were electrophoresed in a 2% agarose
gel and visualized by ethidium bromide and UV light (c150; Azure
Biosystems, Inc., Dublin, CA, USA).
Restriction enzyme digestion
technique
According to the SNP mutation site gene, we selected
the restriction enzyme HinfI (Takara Bio, Inc.) for the
treatment of PCR amplified fragments, to carry on the
identification of the gene. All of the 180 GDM and 210 in the
control group PCR amplified fragments were by HinfI. The
reaction system contained Takara HinfI 1 μl, 10X
HinfI buffer 2 μl, PCR template 7-8 μl, add
ultrapure water to 20 μl, kept in 37°C water for 8 h. The
fragments digested by HinfI were electrophoresed in a 3%
agarose gel. Genotypes were determined by the results of various
bands though UV light (c150; Azure Biosystems, Inc.).
Western blot analysis and SDS-PAGE
Placental tissue total protein was extracted by RIPA
with phenylmethylsulfonyl fluoride (PMSF) (Beyotime Institute of
Biotechnology, Shanghai, China). Then put into homogenizer to grind
into tissue homogenate and the supernatant was collected and
centrifuged at 14,000 × g at 4°C for 5 min. The protein
concentration was quantified by BCA assay. Then, samples containing
50 μg were separated by polyacrylamide gel electrophoresis
with a 10% separating gel (pH 8.8) and a 5% stacking gel (pH 6.8)
(Beyotime Institute of Biotechnology) and the proteins were
transferred onto PVDF membranes (Millipore, Billerica, MA, USA).
Membranes were incubated with primary antibody: goat anti-human
SHBG polyclonal antibody (INC sc-32468, 1:2,000 dilution; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and GAPDH monoclonal
antibody (60004-1-lg, 1:10,000 dilution; Proteintech, Wuhan, China)
overnight at 4°C after blocking in 5% skim milk for 2 h. Following
three washes with 1X Tris-buffered saline containing 0.1% Tween-20
(TBST), the membranes were incubated in secondary antibody:
HBP-conjugated rabbit anti-goat IgG (Ab-204-01, 1:5,000 dilution;
Vazyme, Nanjing, China) and peroxidase-conjugated AffiniPure goat
anti-mouse IgG (SA00001-1, 1:2,000 dilution; Proteintech) for 120
min then room temperature for 2 h. Immune complexes were detected
with enhanced chemiluminescence (ECL) (Beyotime Institute of
Biotechnology). A bioanalytical imaging system (c300; Azure
Biosystems, Inc.) was used to catch the bands. All experiments were
repeated at least three times. Data were expressed as a ratio of
SHBG gray value to GAPDH (ImageJ; National Institutes of Health,
Bethesda, MA, USA).
Cell culture
The placenta villus trophoblast cells HTR8-SVneo
were obtained from Canada Queen's University and cultured
conventionally with RPMI-1640 medium, supplemented with 10%
heat-inactivated fetal bovine serum (FBS) (Gibco, Carlsbad, CA,
USA). Standard culture condition: 37°C, 5% humidified
CO2 incubator. Cell culture supplies were purchased from
the Greiner Bio-One GmbH (Frickenhausen, Germany), trypsin
(Gibco).
Transfection
The recombinant lentivirus was designed and
synthesized with all of the SBHG genetic sequence labeled by green
fluorescence (GenePharma, Shanghai, China). For this part,
HTR8-SVneo cells were divided into ten flasks, some of which had
upregulated SHBG expression. A and B were the normal groups
(without transfection), C and D were the blank control groups
(transfected with empty virus LV-5). E-G were negative control
groups (respectively transfected with SHBG rs6259 Asp-1, rs6259
Asp-2 and rs6259 Asp-3). H-J were the positive transfection group
(respectively transfected with SHBG rs6259 Asn-1, rs6259 Asn-2 and
rs6259 Asn-3).
One day before transfection, cells were cultured
with 90% RPMI-1640+10% FBS and seeded in 25 cm2 flasks
(250-500×103 cells/flasks), cultivated at 37°C, in 5%
CO2 incubator. Twenty-four hours later, the cell growth
reached 30-40%, the medium was replaced and the cells were
trans-fected with the lentivirus. Cells were incubated at 37°C,
morphological structure was observed at 12 h and the culture media
was changed at 24 h. After 96 h, transfection efficiency was
determined by using fluorescence imaging. Then, the cells were
collected, SHBG mRNA and protein were assayed via western blot
analysis and RT-qPCR.
Gene expression analysis by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total SHBG mRNA was extracted by TRIzol reagent
(Invitrogen Life Technologies) from transfected cells according to
the manufacturer's instructions. Amplification of SHGB cDNA
fragment was divided into two parts: i) removal of extraneous DNA
contamination: 1 μl RNA template, 2 μl 5X g DNA Erase
buffer, 1 μl g DNA Erase buffer, 6 μl RNase Free
dH2O, at 42°C for 2 min, then for 4°C; ii) cDNA
production: 4 μl 5X g Premix Script buffer 2, 1 μl
Premix Script RT enzyme mix, 1 μl RT Primix Mix and 4
μl RNase Free dH2O into reaction volume from step
1, at 37°C for 15 min, at 85°C for 5 sec, then for 4°C (no. RR047A;
Takara Bio, Inc.).
PCR primer sequences were as follows: SHBG mRNA
forward primer, 5′-CCTCACCAAGATCACAAAAA-3′ and reverse primer,
5′-TCTCGAAGTCCCAGCATAAACC-3′, giving a fragment length of 120 bp;
β-actin forward primer, 5′-AGCACAATGAAGATCAAGATCAT-3′ and reverse
primer, 5′-ACTCGTCATACTCCTGCTTGC-3′, giving a fragment length of
127 bp (Invitrogen Life Technologis). Real-time PCR amplification
was carried out in 7500 fast thermocycler (Life Technologies) as
follows: 10 μl SYBR Premix Ex Taq, 6 μl RNase Free
dH2O, 1 μl each of SHBG and β-actin forward and
reverse primers (10 μM), 2 μl cDNA template.
Conditions were: 95°C for 5 min (95°C for 10 sec, 60°C for 30 sec)
40 (no. RR820A; Takara Bio, Inc.). Gene expression was determined
by 2−ΔΔCt, where ΔCt = (CtSHBG −
CtGAPDH) and Ct is the threshold cycle.
Transfected HTR8-SVneo cell SHBG
detection by western blot analysis
Expression of SHBG was tested and analyzed in
different transfected groups with the same approach as placental
tissue.
Statistical analysis
The direct counting method for % calculation of the
gene frequency. The differences with alleles and genotypes
distribution compared with Chi-square statistics, P<0.05 was
considered statistically significant. Hardy-Weinberg balance check
(HWE) the reliability of the survey data (gene frequency,
estimation accuracy and reliability) and disease risk with rs6259
through the SHEsis software (http://analysis.bio-x.cn/myAnalysis.php) (23). The correlation between SHBG levels
and genotypes were evaluated by Pearson's correlation coefficient.
Logistic regression models were used to analyse the correlation of
genotype and disease with odds ratios (ORs) and 95% confidence
intervals (95% CIs). T-test and LSD t-test were used with α=0.05
considered to be significant. All analyses used SPSS 13.0 software
(SPSS, Inc., Chicago, IL, USA).
Results
Clinical data analyses
The statistics and analysis of population
characteristics are depicted in Table
I, with the mean ± standard deviation. The differences of age
distribution, gestational weeks, childbearing history and birth
weight had no significant difference, but statistically significant
difference wasfound in fasting blood glucose between normal and GDM
pregnant womean (P<0.05).
 | Table IClinical characteristics of normal
and GDM pregnant women. |
Table I
Clinical characteristics of normal
and GDM pregnant women.
|
Characteristics | GDM group | Control group | P-value |
|---|
| Numbers | 180 | 210 | |
| Age (years) | 31.59±3.89 | 31.73±4.46 | 0.735b |
| Fasting blood
glucose | 5.04±0.86 | 4.32±0.44 | 0.000a |
| Gestational
weeks | 38.69±1.13 | 38.83±0.79 | 0.141b |
| Childbearing
history | 0.10±0.31 | 0.16±0.37 | 0.056b |
| Birth weight
(g) | 3445.6±415.1 | 3394.5±469.1 | 0.308b |
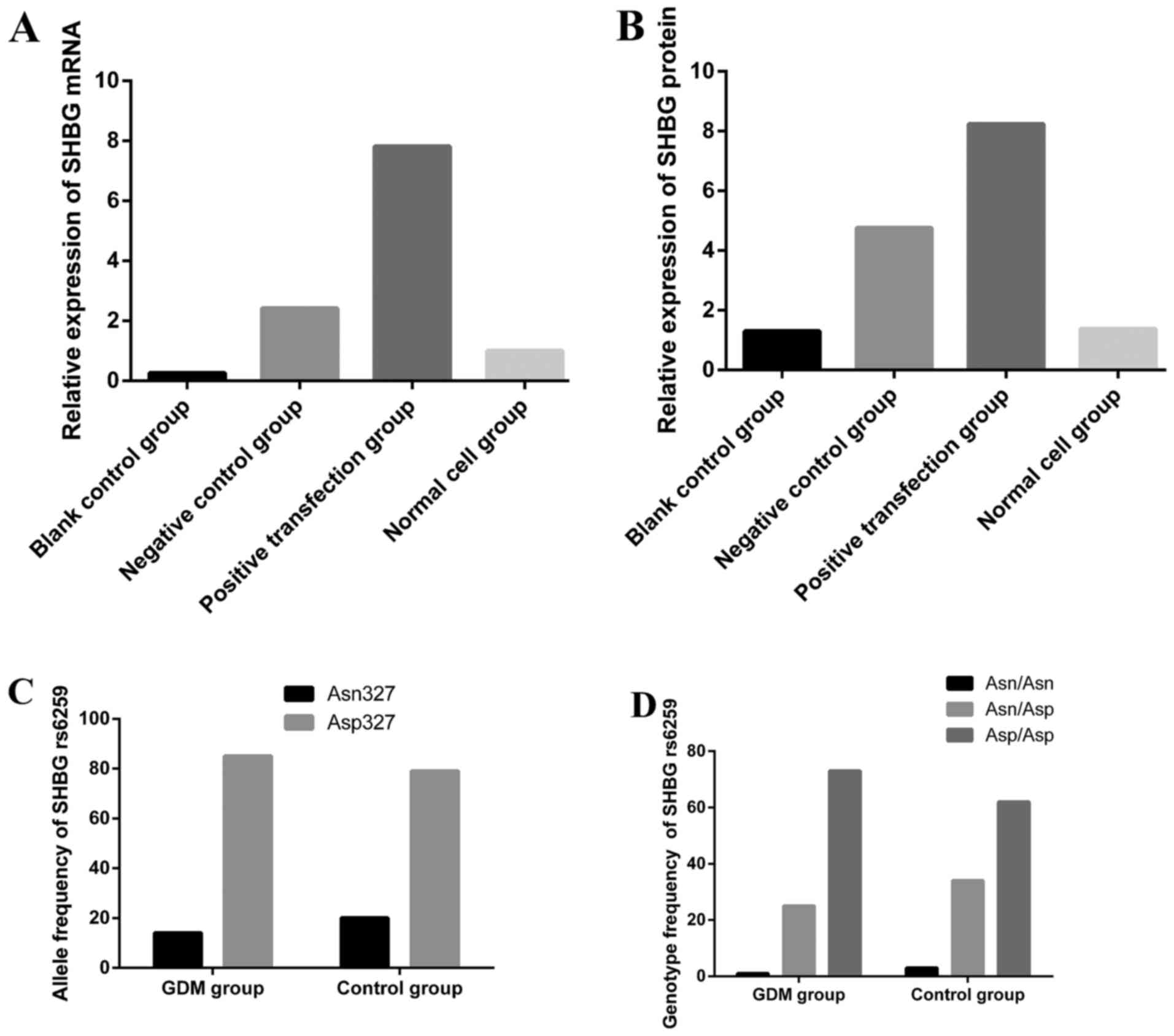
Placenta SHBG gene rs6259 allele
frequencies and genotype distribution
PCR fragment amplification is shown in Fig. 1A. Three genotypes were detected in
two groups: the GG genotype represented three segments, while the
AA genotype was characterized by two parts and GA genotype was
constituted by four segments (Fig.
1B). SHEsis online software detected the Hardy-Weinberg
balance, control group: χ2=0.469, P=0.493; GDM group:
χ2=0.141, P=0.707. The gene frequency conforms to the
laws of genetic balance constantly (P>0.05). Allelic frequencies
and genotype distributions are given in Table II. Because of the low proportion
of AA homozygotes, they were considered together with the GA
heterozygotes. In the GDM pregnancy group, 14.17% had the variant
gene, higher than the control group. Compared with the control
group, the decreased tendencies were observed in the GDM group with
AA and GA group. The result [odd ratio <1, 95% CI=(0-1)] shows
that rs6259 is a protective factor for decreased risk of GDM
(P=0.043). Pearson's p=0.707 confirms that rs6259 is highly
correlated with the morbidity of GDM.
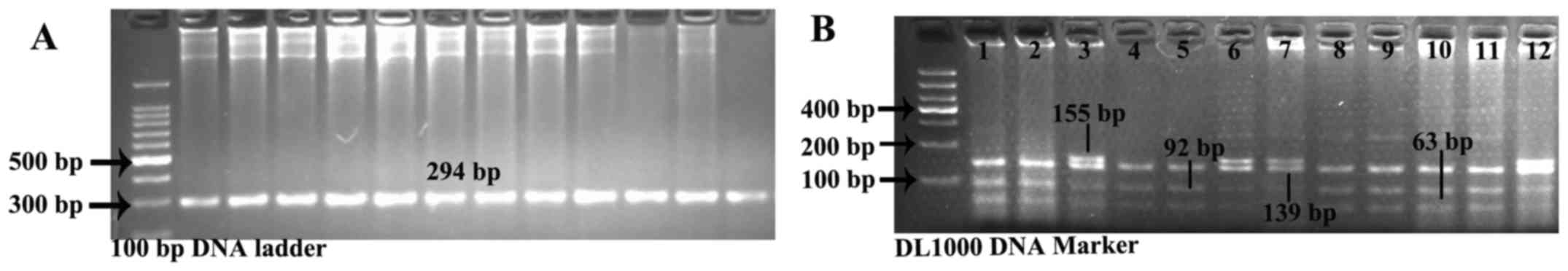 | Figure 1(A) Sex hormone-binding globulin
(SHBG) rs6259 fragment amplification by polymerase chain
reaction-restriction fragment length polymorphism (PCR-RFLP). PCR
fragments, 294 bp. (B) Results after restriction enzyme digestion.
The GG genotype was constituted by segments of 139, 92 and 63 bp,
while the AA genotype was characterized by segments of 155 and 139
bp and the GA genotype was characterized by segments of 155, 139,
92 and 63 bp. Lanes 1, 2, 4, 5, 8, 9, 10, 11: GG homozygote
(Asp/Asp); lanes 3, 6, 12: GA heterozygote (Asn/Asp); lane 7, AA
homozygote (Asn/Asn). |
| Table IIAllele and genotypic frequencies of
SHBG Asp327Asn polymorphism in placenta. |
Table II
Allele and genotypic frequencies of
SHBG Asp327Asn polymorphism in placenta.
| Frequencies | Control group
(n=210)
| GDM group (n=180)
| P-value |
|---|
| n | (%) | n | (%) |
|---|
| Allele | | | | | |
| A (Asn) | 86 | 20.48 | 51 | 14.17 | |
| G (Asp) | 334 | 79.52 | 309 | 85.83 | 0.021a |
| Genotype | | | | | |
| GG (Asp/Asp) | 131 | 62.38 | 132 | 73.33 | 0.021b |
| GA+AA | 79 | 37.62 | 48 | 26.67 | 0.021c |
| GA (Asp/Asn) | 72 | 34.29 | 45 | 25 |
0.046d |
| AA (Asn/Asn) | 7 | 3.33 | 3 | 1.67 |
0.299e |
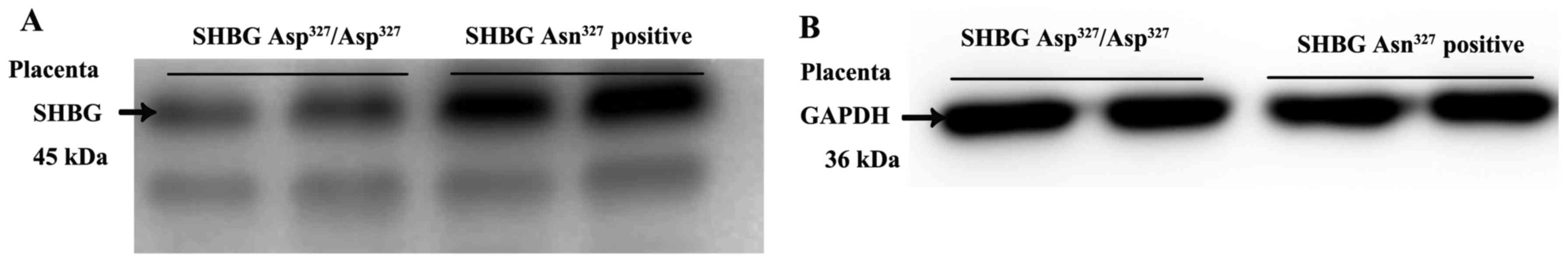
SHBG expression of different genotypes in
placental tissues
Following the results of PCR-RFLP and enzyme
digestion, the placental tissues were divided into two groups. As
shown in Fig. 2, the SHBG
concentration of the GA heterozygote and AA homozygote (SHBG
Asn327 positive group) are significantly higher than the
GG homozygote (SHBG Asp327/Asp327 group). The
difference between the two groups was statistically significant
(t=2.176, P=0.035 <0.001).
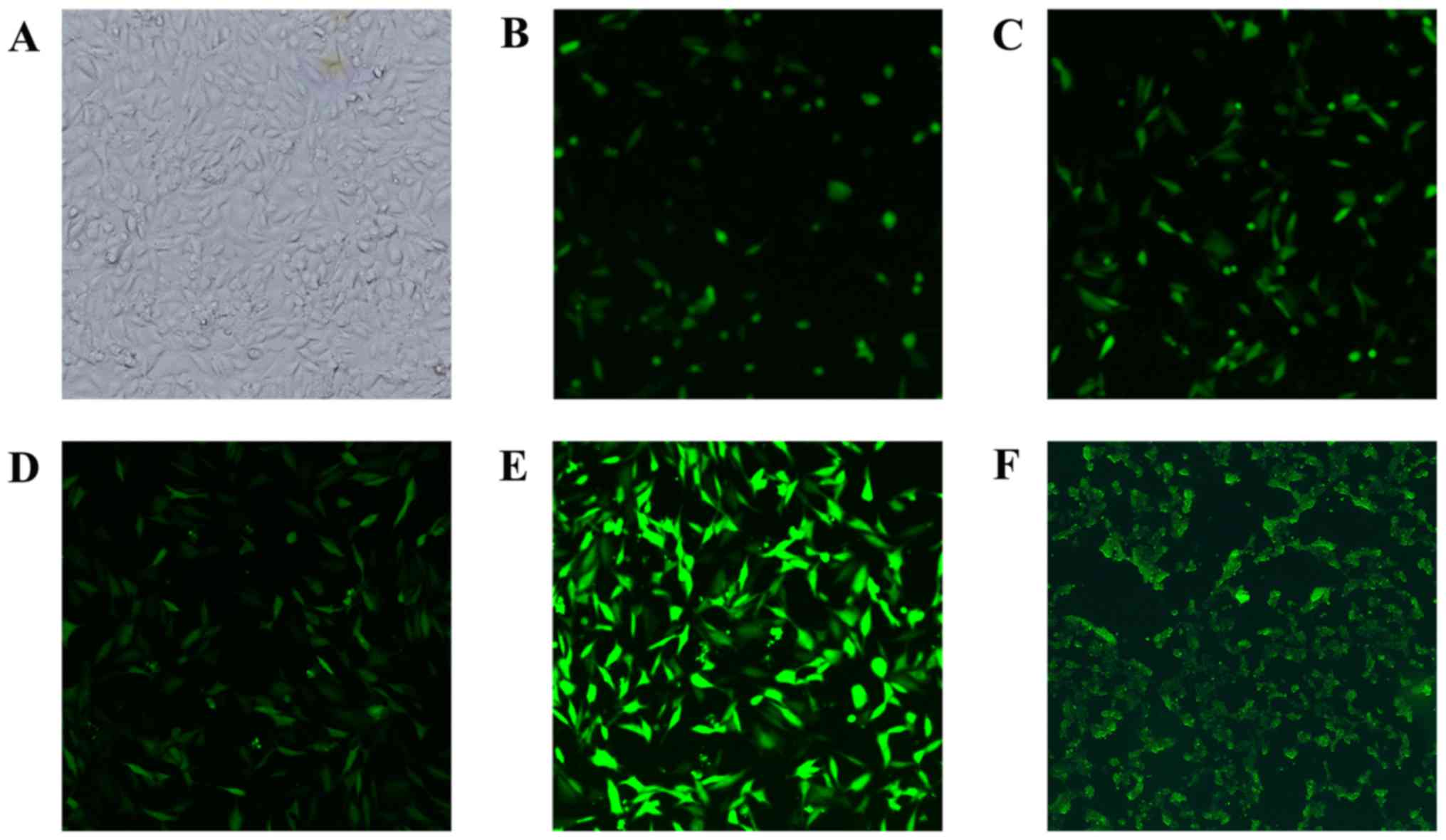
HTR8-SVneo cell transfection
HTR8-SVneo cells were incubated in 5%
CO2, 37°C incubator. After repeated exploration the
optimal transfection conditions were: MOI=80, 5 μg/ml
polybrene, which were successfully transfected with recombinant
lentivirus and upregulation of SHBG expression (Fig. 3).
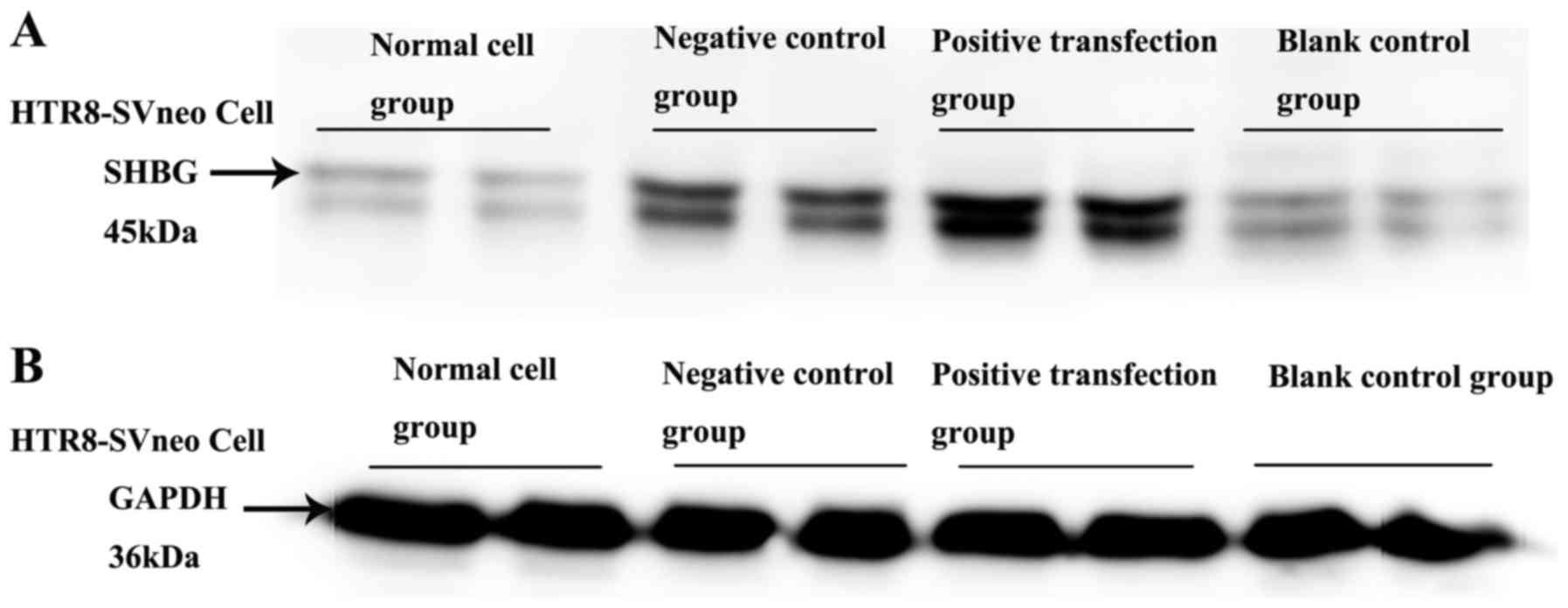
Recombinant intracellular SHBG protein
assay
Western blot analysis and gel image software
analysis demonstrated that the positive transfection groups showed
an increased SHBG level over negative control groups (t=2.675,
P=0.011 <0.001), blank control and normal groups (P<0.001).
There was no significant difference between the blank control group
and the normal group (Table
IIIA; Figs. 4 and 5).
| Table IIIThe result of SHBG protein and mRNA
in different groups of transfected cells. |
Table III
The result of SHBG protein and mRNA
in different groups of transfected cells.
| A, Analysis of SHBG
protein and GAPDH gray ratio in different transfected HTR8-SVneo
cells |
|---|
| LSD t-test | Groups | P-value | 95% CI |
|---|
| NG | LV-5 |
0.950a | −0.225–0.240 |
| rs6259 Asp |
0.003b | 0.116–0.560 |
| rs6259 Asn |
0.000c | 0.463–0.907 |
| rs6259 Asn | rs6259 Asp |
0.000d | 0.148–0.545 |
| LV-5 group | rs6259 Asp |
0.002e | 0.136–0.557 |
| rs6259 Asn |
0.000f | 0.482–0.902 |
|
| B, Analysis of SHBG
mRNA transcription in different trans-fected HTR8-SVneo cells |
|
| LSD t-test | Groups | P-value | 95% CI |
|
| NG | LV-5 |
0.201a | −0.413–1.890 |
| rs6259 Asp |
0.017b | 0.272–2.575 |
| rs6259 Asn |
0.000c | 5.660–7.964 |
| rs6259 Asn | rs6259 Asp |
0.000d | 4.237–6.541 |
| LV-5 group | rs6259 Asp |
0.001e | 1.011–3.314 |
| rs6259 Asn |
0.000f | 6.399–8.703 |
Detection the SHBG mRNA in transfected
HTR8-SVneo cells
According to the LDS-t analysis there were no
significant differences between the blank control group and normal
cell group (P>0.05), in the positive transfection groups a
higher average 2−ΔΔCt for SHBG mRNA were found than the
negative control groups (P<0.001), blank control groups
(P<0.001) and normal cell groups (P<0.001), the differences
were statistically significant (Table IIIB; Fig. 5).
Discussion
The gene encoding human SHBG is located on
chromosome 17p12-13 and is composed of 8 exons and 7 introns (Gene
ID, 6462). The length of SHBG gene is 3.2 kb and it encodes a
polypeptide of 402 amino acids. Human SHBG gene is prone to gene
mutation, deletion and recombination. Because of its fragility and
volatility, it is more likely to accept foreign interference
(24,25). The polymorphic and the protein
level changes may lead to special expression in certain diseases
(26), such as IR damage
(7,27), DM (7,23),
polycystic ovary syndrome (21),
osteoporosis (28), hirsutism
(11), and even some
hormone-dependent tumors, such as breast (16,20), ovarian (17), endometrial (18) and prostate cancers (29,30). rs6259 is one of the functional
SHBG SNPs, which leads to an amino acid substitution of asparagine
for aspartic acid at locus 327 (Asp327Asn, D327N) in the
SHBG polypeptide (11).
In our study, 180 pregnant women with GDM and 210
healthy pregnant women were chosen as participants in the program
at the same period. The distributions of the genotypes and the
allele frequencies had obvious differences in two groups. In GDM
group, the frequency of the mutation gene and genotypes showed
decreasing trends. The frequency of the variant allele A is higher
in healthy pregnant women of northeast China than in healthy
Caucasian and African Americans, confirming the racial diversity in
the distribution of this mutation.
Then, we measured SHBG level in each group. We
discovered the differences between the two groups with disparate
alleles (Asp327 or Asn327). Through many
repeats, we determined that the variant allele Asn327
have a strong connection with the added tendency of SHBG in the
placenta organ. So, considering the result of genotypes screen, we
speculated that Asn327 may lead to a reduced risk of
GDM.
To clarify the mechanism involved, placental villi
trophoblastic cells were cultivated continuously in vitro.
With repeatedly recombinant lentivirus transfection, we found that
both SHBG protein and SHBG mRNA in transfected HTR8-SVneo cells
carried the mutant allele leading to higher level than others
without mutation. Excluding other possible interference factors,
the strong correlation between SHBG rs6259 variant allele A and
SHBG transcription and translation was confirmed. It may be a
genetic molecular basis of the pathogenesis of GDM.
It is well-known that type 2 diabetes is an
endocrine disease with obvious genetic predisposition. If one of
the parents has type 2 diabetes, genetic odds of the offspring is
between 1/7 and 1/13, but if both parents are affected, the genetic
probability is increased to 1/2. Our preliminary experiment
assessed the relevance between placental SHBG and GDM (8-10).
The placenta has the same genetic material as the fetus, therefore,
we anticipate to make a preliminary prediction for the possibility
that the next generation achieve insulin resistance and type 2
diabetes, by testing the SHBG level and SHBG rs6259.
In conclusion, the findings of the present study
support the result that SHBG SNP rs6259 is closely associated with
placental SHBG levels. The mutant Asn327 allele may
affect the transcription of SHBG mRNA and activity of the SHBG
protein by some mechanisms. Associated with the SHBG rs6259 variant
allele A, SHBG protein levels improved in placenta, the risk and
progress of GDM may be delayed. SHBG rs6259 and SHBG are protective
factors in pathogenesis of GDM, and may possiby be used to deduce
the risk of offspring suffering from diabetes in the future.
Acknowledgments
Not applicable.
Notes
[1]
Funding
This study was supported by the National Natural
Science Foundation of China (nos. 81300511 and 81170591).
[2] Availability
of data and material
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
XZ contributed to manuscript writing, data
collection and data analysis. LS contributed to data collection and
financial support. ZJ contributed to project development.
[4] Ethics
approval and consent to participate
This study was in accordance with the ethical
standards of the Shengjing Hospital of China Medical University and
the ethical standards of the 1964 Helsinki declaration and its
later amendments. All individual participants included in the study
provided informed consents.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang HX: Further enhance the research
level of gestational diabetes. Chin J Perinat Med. 8:289–291.
2005.
|
|
2
|
Lewis JG, Shand BI, Elder PA and Scott RS:
Plasma sex hormone-binding globulin rather than
corticosteroid-binding globulin is a marker of insulin resistance
in obese adult males. Diabetes Obes Metab. 6:259–263. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sokup A, Szymański M and Góralczyk K:
Impaired fasting glucose as a marker of heterogeneity of
gestational diabetes mellitus. A study of 1025 women living in the
region of Kuyavia and Pomerania in Poland. Endokrynol Pol.
60:348–352. 2009.PubMed/NCBI
|
|
4
|
Buchanan TA, Xiang A, Kjos SL, Lee WP,
Trigo E, Nader I, Bergner EA, Palmer JP and Peters RK: Gestational
diabetes: antepartum characteristics that predict postpartum
glucose intolerance and type 2 diabetes in Latino women. Diabetes.
47:1302–1310. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morrison JA, Glueck CJ, Daniels S, Wang P
and Stroop D: Adolescent oligomenorrhea in a biracial schoolgirl
cohort: a simple clinical parameter predicting impaired fasting
glucose plus type 2 diabetes mellitus, insulin, glucose, insulin
resistance, and centripetal obesity from age 19 to 25 years.
Metabolism. 60:1285–1293. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nanda S, Savvidou M, Syngelaki A, Akolekar
R and Nicolaides KH: Prediction of gestational diabetes mellitus by
maternal factors and biomarkers at 11 to 13 weeks. Prenat Diagn.
31:135–141. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Perry JR, Weedon MN, Langenberg C, Jackson
AU, Lyssenko V, Sparsø T, Thorleifsson G, Grallert H, Ferrucci L,
Maggio M, et al: MAGIC: Genetic evidence that raised sex hormone
binding globulin (SHBG) levels reduce the risk of type 2 diabetes.
Hum Mol Genet. 19:535–544. 2010. View Article : Google Scholar
|
|
8
|
Jin Z, Guan X, Gao H, Shang L, Gao M, Su D
and Li W: The change in sex hormone binding globulin and the
influence by gestational diabetes mellitus in fetal period. Gynecol
Endocrinol. 25:647–652. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun L, Jin Z, Teng W, Chi X, Zhang Y, Ai W
and Wang P: Expression changes of sex hormone binding globulin in
GDM placental tissues. J Perinat Med. 40:129–135. 2011.PubMed/NCBI
|
|
10
|
Sun L, Jin Z, Teng W, Chi X, Zhang Y, Ai W
and Wang P: SHBG in GDM maternal serum, placental tissues and
umbilical cord serum expression changes and its significance.
Diabetes Res Clin Pract. 99:168–173. 2013. View Article : Google Scholar
|
|
11
|
Cousin P, Calemard-Michel L, Lejeune H,
Raverot G, Yessaad N, Emptoz-Bonneton A, Morel Y and Pugeat M:
Influence of SHBG gene pentanucleotide TAAAA repeat and D327N
polymorphism on serum sex hormone-binding globulin concentration in
hirsute women. J Clin Endocrinol Metab. 89:917–924. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Power SG, Bocchinfuso WP, Pallesen M,
Warmels-Rodenhiser S, Van Baelen H and Hammond GL: Molecular
analyses of a human sex hormone-binding globulin variant: evidence
for an additional carbohydrate chain. J Clin Endocrinol Metab.
75:1066–1070. 1992.PubMed/NCBI
|
|
13
|
Haiman CA, Riley SE, Freedman ML, Setiawan
VW, Conti DV and Le Marchand L: Common genetic variation in the sex
steroid hormone-binding globulin (SHBG) gene and circulating SHBG
levels among postmenopausal women: The Multiethnic Cohort. J Clin
Endocrinol Metab. 90:2198–2204. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cousin P, Déchaud H, Grenot C, Lejeune H,
Hammond GL and Pugeat M: Influence of glycosylation on the
clearance of recombinant human sex hormone-binding globulin from
rabbit blood. J Steroid Biochem Mol Biol. 70:115–121. 1999.
View Article : Google Scholar
|
|
15
|
Ding EL, Song Y, Manson JE, Hunter DJ, Lee
CC, Rifai N, Buring JE, Gaziano JM and Liu S: Sex hormone-binding
globulin and risk of type 2 diabetes in women and men. N Engl J
Med. 361:1152–1163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Försti A, Jin Q, Grzybowska E, Söderberg
M, Zientek H, Sieminska M, Rogozinska-Szczepka J, Chmielik E,
Utracka-Hutka B and Hemminki K: Sex hormone-binding globulin
polymorphisms in familial and sporadic breast cancer.
Carcinogenesis. 23:1315–1320. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garcia-Closas M, Brinton LA, Lissowska J,
Richesson D, Sherman ME, Szeszenia-Dabrowska N, Peplonska B, Welch
R, Yeager M, Zatonski W, et al: Ovarian cancer risk and common
variation in the sex hormone-binding globulin gene: a
population-based case-control study. BMC Cancer. 7:602007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kataoka N, Cai Q, Xu WH, Xiang YB, Cai H,
Zheng W and Shu XO: Association of endometrial cancer risk with a
functional polymorphism (Asp327Asn) in the sex
hormone-binding globulin gene. Cancer. 109:1296–1302. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eriksson AL, Lorentzon M, Mellström D,
Vandenput L, Swanson C, Andersson N, Hammond GL, Jakobsson J, Rane
A, Orwoll ES, et al: SHBG gene promoter polymorphisms in men are
associated with serum sex hormone-binding globulin, androgen and
androgen metabolite levels, and hip bone mineral density. J Clin
Endocrinol Metab. 91:5029–5037. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui Y, Shu XO, Cai Q, Jin F, Cheng JR, Cai
H, Gao YT and Zheng W: Association of breast cancer risk with a
common functional polymorphism (Asp327Asn) in the sex
hormone-binding globulin gene. Cancer Epidemiol Biomarkers Prev.
14:1096–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bendlová B, Zavadilová J, Vanková M,
Vejrazková D, Lukásová P, Vcelák J, Hill M, Cibula D, Vondra K,
Stárka L, et al: Role of D327N sex hormone-binding globulin gene
polymorphism in the pathogenesis of polycystic ovary syndrome. J
Steroid Biochem Mol Biol. 104:68–74. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
American Diabetes Association: Diagnosis
and classification of diabetes mellitus. Diabetes Care. 36(Suppl
1): S67–S74. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi YY and He L: SHEsis, a powerful
software platform for analyses of linkage disequilibrium, haplotype
construction, and genetic association at polymorphism loci. Cell
Res. 15:97–98. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cousin P, Billotte J, Chaubert P and Shaw
P: Physical map of 17p13 and the genes adjacent to p53. Genomics.
63:60–68. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Toscano V, Balducci R, Bianchi P,
Guglielmi R, Mangiantini A and Sciarra F: Steroidal and
non-steroidal factors in plasma sex hormone binding globulin
regulation. J Steroid Biochem Mol Biol. 43:431–437. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xita N and Tsatsoulis A: Genetic variants
of sex hormone-binding globulin and their biological consequences.
Mol Cell Endocrinol. 316:60–65. 2010. View Article : Google Scholar
|
|
27
|
Pinós T, Barbosa-Desongles A, Hurtado A,
Santamaria- Martínez A, de Torres I, Morote J, Reventós J and
Munell F: Identification, characterization and expression of novel
sex hormone binding globulin alternative first exons in the human
prostate. BMC Mol Biol. 10:592009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Goderie-Plomp HW, van der Klift M, de
Ronde W, Hofman A, de Jong FH and Pols HA: Endogenous sex hormones,
sex hormone-binding globulin, and the risk of incident vertebral
fractures in elderly men and women: The Rotterdam Study. J Clin
Endocrinol Metab. 89:3261–3269. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Selva DM and Hammond GL: Human sex
hormone-binding globulin is expressed in testicular germ cells and
not in sertoli cells. Horm Metab Res. 38:230–235. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Weiss JM, Huang WY, Rinaldi S, Fears TR,
Chatterjee N, Hsing AW, Crawford ED, Andriole GL, Kaaks R and Hayes
RB: Endogenous sex hormones and the risk of prostate cancer: a
prospective study. Int J Cancer. 122:2345–2350. 2008. View Article : Google Scholar : PubMed/NCBI
|